Differential Responses of Colorectal Cancer Cell Lines to Enterococcus faecalis’ Strains Isolated from Healthy Donors and Colorectal Cancer Patients

 , , , , , ,
, , , , , ,  ,
, 
Abstract
:1. Introduction
2. Material and Methods
2.1. Ethical Statement
2.2. CRC Patients and Healthy Donors
2.3. Isolation and Genotyping of Enterococcus faecalis
2.4. Secreted Metabolites
2.5. Gas Chromatography Mass Spectrometry (GC-MS) Analysis of SCFAs
2.6. Cell Lines
2.7. Treatment of Cell Lines with Pre-Fermented Medium and MTT Assay
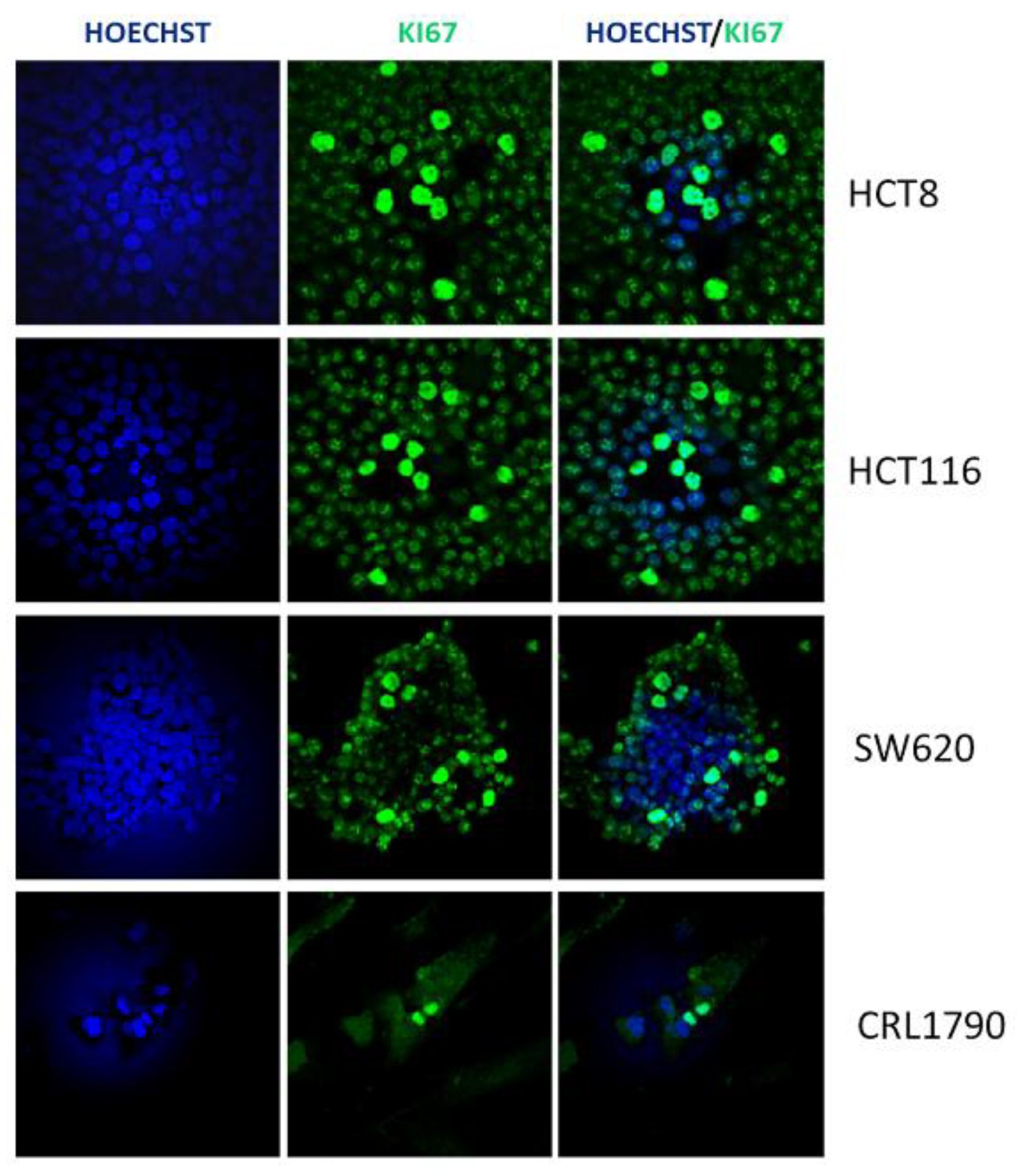
2.8. Ki67 Immunofluorescence Analysis
2.9. Statistical Analysis
3. Results
3.1. Decreased Frequency of E. faecalis on Stool of CRC Patients
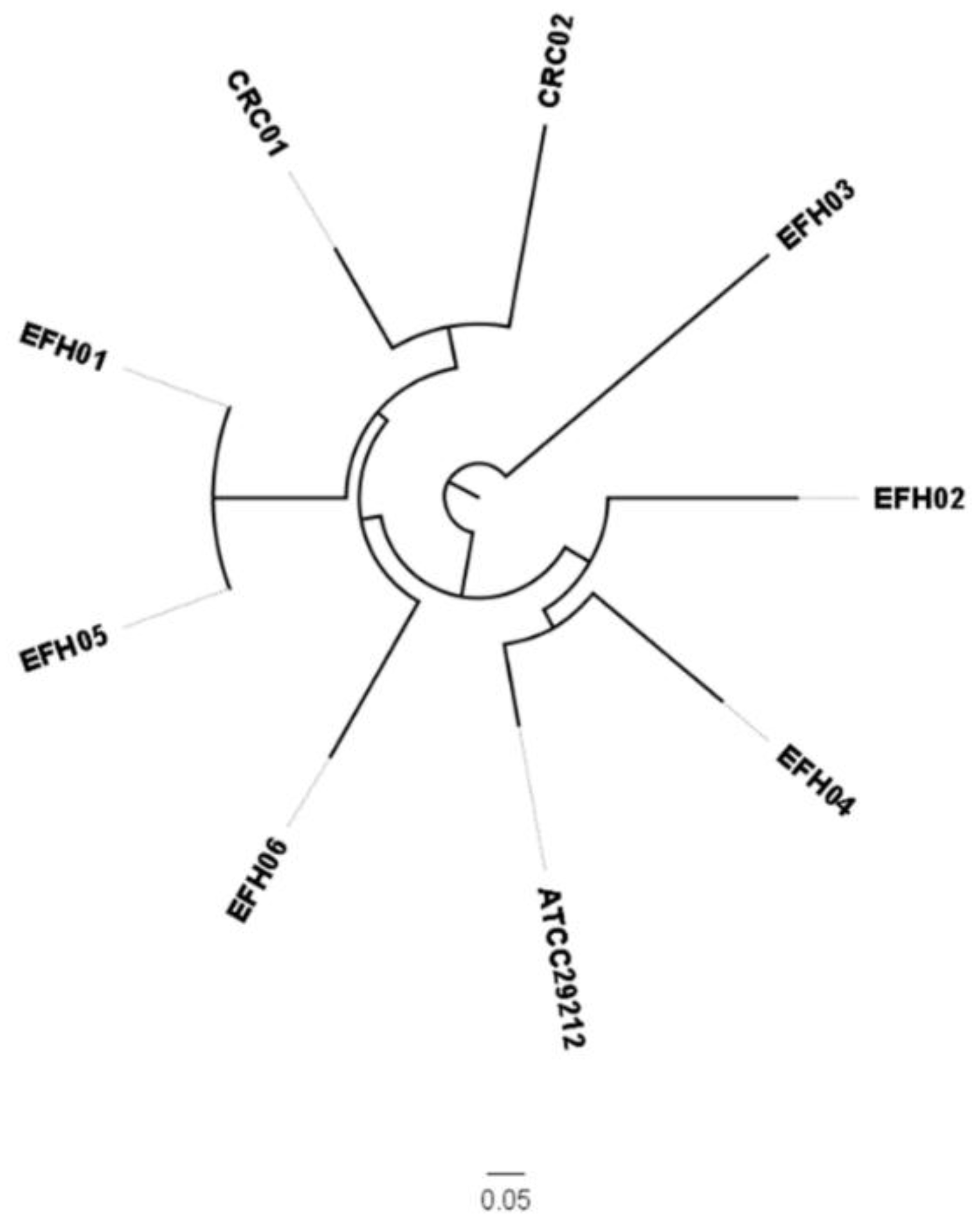
3.2. Genotypic Characterization of E. faecalis’ Strains
3.3. Characterization of Secreted SCFAs
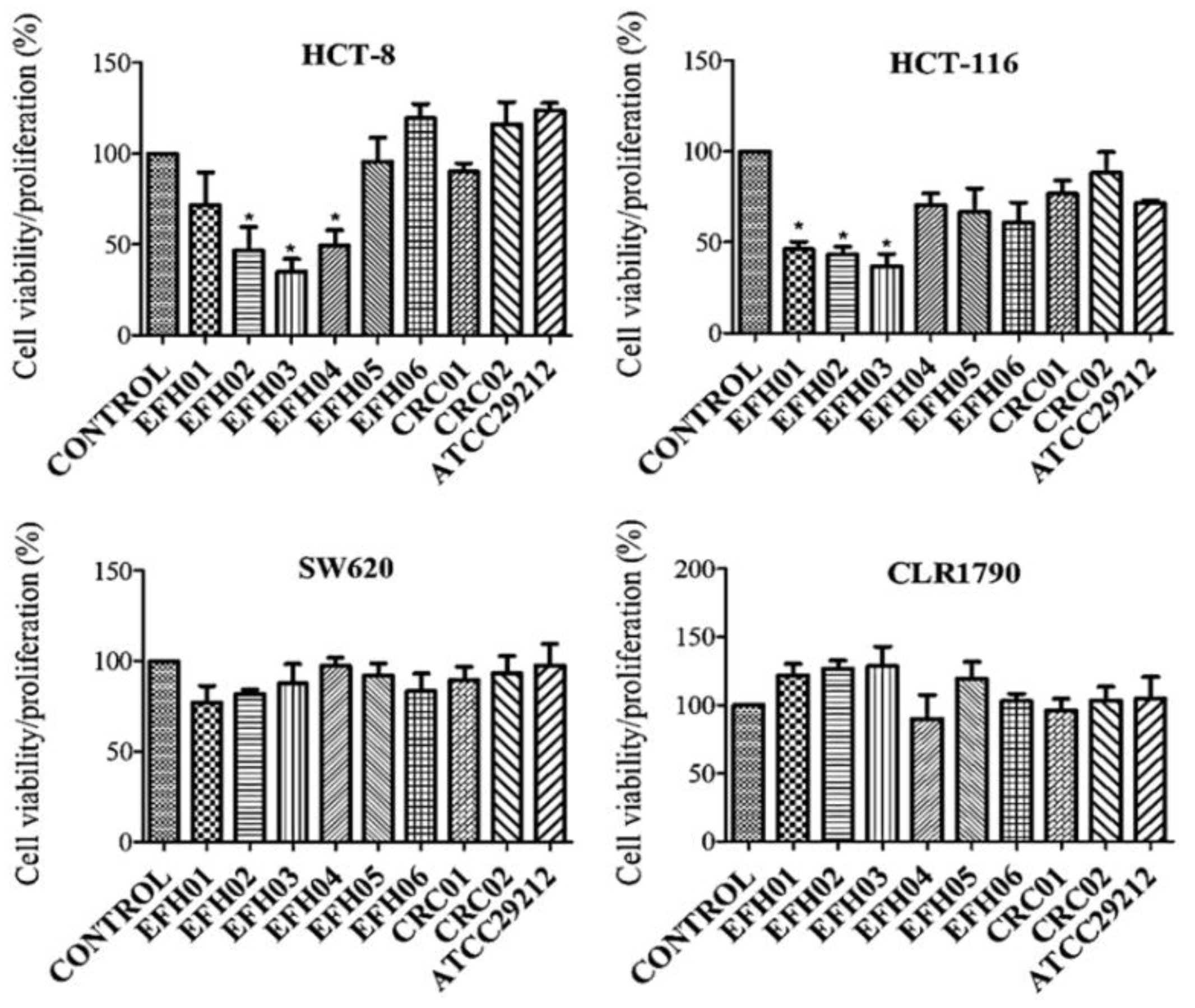
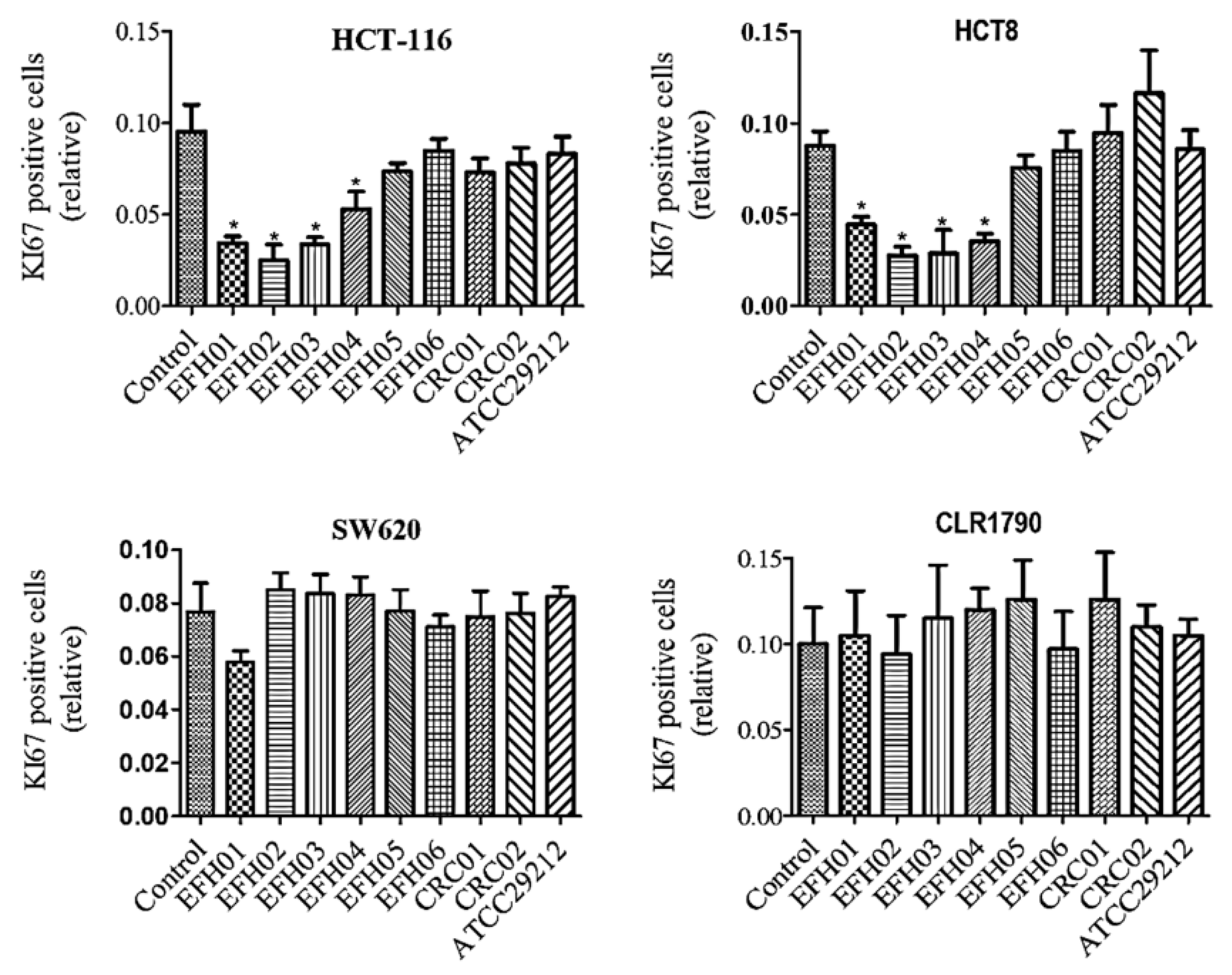
3.4. From the Isolated Strains, Three were Able to Decrease Tumor Cell Growth
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer Statistics, 2018. CA Cancer J. Clin. 2018, 68, 7–30. [Google Scholar] [CrossRef]
- Das Neves, F.J.; Mattos, I.E.; Koifman, R.J. Colon and rectal cancer mortality in Brazilian capitals, 1980-1997. Arq. Gastroenterol. 2005, 42, 63–70. [Google Scholar]
- Ferlay, J.; Shin, H.; Bray, F.; Forman, D.; Mathers, C.; Parkin, D.M. Estimates of worldwide burden of cancer in 2008: GLOBOCAN 2008. Int. J. Cancer. 2010, 127, 2893–2917. [Google Scholar] [CrossRef] [Green Version]
- MacFarlane, A.J.; Stover, P.J. Convergence of genetic, nutritional and inflammatory factors in gastrointestinal cancers. Nutr. Rev. 2007, 65, S157–S166. [Google Scholar] [CrossRef]
- Colorectal Cancer Risk Factors. 2019. Available online: http://www.cdc.gov/cancer/colorectal/basic_info/risk_factors.htm (accessed on 7 February 2019).
- De Almeida, C.V.; Taddei, A.; Amedei, A. The controversial role of Enterococcus faecalis in Colorectal cancer. Therap. Adv. Gastroenterol. 2018, 11, 1–11. [Google Scholar] [CrossRef]
- Rea, D.; Coppola, G.; Palma, G.; Barbieri, A.; Luciano, A.; Del Prete, P.; Rossetti, S.; Berretta, M.; Facchini, G.; Perdonà, S.; et al. Microbiota effects on cancer: From risks to therapies. Oncotarget 2018, 9, 17915–17927. [Google Scholar] [CrossRef]
- Boleij, A.; Tjalsma, H. Gut bacteria in health and disease: A survey on the interface between intestinal microbiology and colorectal cancer. Biol. Rev. Camb. Philos. Soc. 2012, 87, 701–730. [Google Scholar] [CrossRef]
- Umesaki, Y.; Okada, Y.; Matsumoto, S.; Imaoka, A.; Setoyama, H. Segmented filamentous bacteria are indigenous intestinal bacteria that activate intraepithelial lymphocytes and induce MHC class II molecules and fucosylasialo GM1 glycolipids on the small intestinal epithelial cells in the ex-germ-free mouse. Microbiol. Immunol. 1995, 39, 555–562. [Google Scholar] [CrossRef]
- Zur Hausen, H. The search for infectious causes of human cancers: Where and why. Virology 2009, 392, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Davis, C.D.; Milner, J.A. Gastrointestinal microflora, food components and colon cancer prevention. J. Nutr. Biochem. 2009, 20, 743–752. [Google Scholar] [CrossRef] [Green Version]
- Collins, D.; Hogan, A.M.; Winter, D.C. Microbial and viral pathogens in colorectal cancer. Lancet Oncol. 2011, 12, 504–512. [Google Scholar] [CrossRef]
- Russo, E.; Bacci, G.; Chiellini, C.; Fagorzi, C.; Niccolai, E.; Taddei, A.; Ricci, F.; Ringressi, M.N.; Borrelli, R.; Melli, F.; et al. Preliminary Comparison of Oral and Intestinal Human Microbiota in Patients with Colorectal Cancer: A Pilot Study. Front. Microbiol. 2018, 8, 2699. [Google Scholar] [CrossRef] [PubMed]
- Russo, E.; Taddei, A.; Ringressi, M.N.; Ricci, F.; Amedei, A. The interplay between the microbiome and the adaptive immune response in cancer development. Therap. Adv. Gastroenterol. 2016, 9, 594–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Artis, D. Epithelial-cell recognition of commensal bacteria and maintenance of immune homeostasis in the gut. Nat. Rev. Immunol. 2008, 8, 411–420. [Google Scholar] [CrossRef]
- Niccolai, E.; Ricci, F.; Russo, E.; Nannini, G.; Emmi, G.; Taddei, A.; Ringressi, M.N.; Melli, F.; Miloeva, M.; Cianchi, F.; et al. The Different Functional Distribution of “Not Effector” T Cells (Treg/Tnull) in Colorectal Cancer. Front Immunol. 2017, 8, 1900. [Google Scholar] [CrossRef] [PubMed]
- Zou, S.; Fang, L.; Lee, M.H. Dysbiosis of gut microbiota in promoting the development of colorectal cancer. Gastroenterol. Rep. (Oxf). 2018, 6, 1–12. [Google Scholar] [CrossRef]
- Prakash, S.; Rodes, L.; Coussa-Charley, M.; Tomaro-Duchesneau, C. Gut microbiota: Next frontier in understanding human health and development of biotherapeutics. Biologics 2011, 5, 71–86. [Google Scholar] [PubMed]
- Habermann, W.; Zimmermann, K.; Skabaris, H.; Kunze, R.; Rusch, V. The effect of bacterial immunostimulant (human Enterococcus faecalis bacteria) on the occurrence of relapse in patients with chronic bronchitis. Arzneimittel-Forschung. 2001, 51, 931–937. [Google Scholar]
- Chen, C.C.; Kong, M.S.; Lai, M.W.; Chao, H.C.; Chang, K.W.; Chen, S.Y.; Huang, Y.C.; Chiu, C.H.; Li, W.C.; Lin, P.Y.; et al. Probiotics have clinical, microbiological, and immunologic efficacy in acute infectious diarrhea. J. Pediatr. Infect. Dis 2010, 29, 135–138. [Google Scholar] [CrossRef]
- Martinez, R.C.; Bedani, R.; Saad, S.M. Scientific evidence for health effects attributed to the consumption of probiotics and prebiotics: An update for current perspectives and future challenges. Br. J. Nutr. 2015, 114, 1993–2015. [Google Scholar] [CrossRef]
- Huycke, M.M.; Abrams, V.; Moore, D.R. Enterococcus faecalis produces extracellular superoxide and hydrogen peroxide that damages colonic epithelial cell DNA. Carcinogenesis 2002, 23, 529–536. [Google Scholar] [CrossRef]
- Balamurugan, R.; Rajendiran, E.; George, S.; Samuel, G.V.; Ramakrishna, B.S. Real-time polymerase chain reaction quantification of specific butyrate-producing bacteria, Desulfovibrio and Enterococcus faecalis in the feces of patients with colorectal cancer. J. Gastroenterol. Hepatol. 2008, 23, 1298–1303. [Google Scholar] [CrossRef]
- Pericàs, J.M.; Corredoira, J.; Moreno, A.; García-País, M.J.; Falces, C.; Rabuñal, R.; Mestres, C.A.; Alonso, M.P.; Marco, F.; Quintana, E.; et al. Relationship between Enterococcus faecalis infective endocarditis and colorectal neoplasm: Preliminary results from a Cohort of 154 patients. Rev. Esp. Cardiol. 2017, 70, 451–458. [Google Scholar] [CrossRef]
- Hill, M.J.; Drasar, B.S.; Aries, V.; Aries, V.; Crowther, J.S.; Williams, R.E. Bacteria and aetiology of cancer of large bowel. Lancet 1971, 1, 95–100. [Google Scholar] [CrossRef]
- Fanaro, S.; Chierici, R.; Guerrini, P.; Vigi, V. Intestinal microflora in early infancy: Composition and development. Acta Paediatr. Suppl. 2003, 91, 48–55. [Google Scholar] [CrossRef]
- Are, A.; Aronsson, L.; Wang, S.; Greicius, G.; Lee, Y.K.; Gustafsson, J.A.; Pettersson, S.; Arulampalam, V. Enterococcus faecalis from newborn babies regulate endogenous PPARgamma activity and IL-10 levels in colonic epithelial cells. Proc. Natl. Acad. Sci. USA 2008, 105, 1943–1948. [Google Scholar] [CrossRef]
- Wang, S.; Hibberd, M.L.; Pettersson, S.; Lee, Y.K. Enterococcus faecalis from healthy infants modulates inflammation through MAPK signaling pathways. PLoS ONE 2014, 9, e97523. [Google Scholar] [CrossRef]
- Martin, B.; Garriga, M.; Hugas, M.; Aymerich, T. Genetic diversity and safety aspects of enterococci from slightly fermented sausages. J Appl Microbiol. 2005, 98, 1177–1190. [Google Scholar] [CrossRef] [Green Version]
- Sedgley, C.M.; Nagel, A.C.; Shelburne, C.E.; Clewell, D.B.; Appelbe, O.; Molander, A. Quantitative real-time PCR detection of oral Enterococcus faecalis in humans. Arch. Oral. Biol. 2005, 50, 575–583. [Google Scholar] [CrossRef]
- Giani, T.; Arena, F.; Pollini, S.; Di Pilato, V.; D’Andrea, M.M.; Henrici De Angelis, L.; Bassetti, M.; Rossolini, G.M. Pseudomonas aeruginosa Working Group. Italian nationwide survey on Pseudomonas aeruginosa from invasive infections: Activity of ceftolozane/tazobactam and comparators, and molecular epidemiology of carbapenemase producers. J. Antimicrob. Chemother. 2017, 3, 664–671. [Google Scholar]
- Center for Genomic Epidemiology. Available online: https://cge.cbs.dtu.dk/services/MLST/ (accessed on 19 March 2019).
- Center for Genomic Epidemiology. Available online: https://cge.cbs.dtu.dk/services/CSIPhylogeny (accessed on 19 March 2019).
- Arndt, D.; Grant, J.R.; Marcu, A.; Sajed, T.; Pon, A.; Liang, Y.; Wishart, D.S. PHASTER: A better, faster version of the PHAST phage search tool. Nucleic Acids Res. 2016, 44, W16–W21. [Google Scholar] [CrossRef] [PubMed]
- Center for Genomic Epidemiology. Available online: https://cge.cbs.dtu.dk/services/ (accessed on 19 March 2019).
- Basic Local Alignment Search Tool. Available online: http://blast.ncbi.nlm.nih.gov/ (accessed on 19 March 2019).
- Grootaert, C.; Van de Wiele, T.; Van Roosbroeck, I.; Possemiers, S.; Vercoutter-Edouart, A.S.; Verstraete, W.; Bracke, M.; Vanhoecke, B. Bacterial monocultures, propionate, butyrate and H2O2 modulate the expression, secretion and structure of the fasting-induced adipose factor in gut epithelial cell lines. Environ. Microbiol. 2011, 13, 1778–1789. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 16, 55–63. [Google Scholar] [CrossRef]
- Wayne Rasband. NationalInstitutes of Health, Bethesda. Available online: http://rsbweb.nih.gov/ij/index.html (accessed on 19 March 2019).
- Yu, J.; Feng, Q.; Wong, S.H.; Zhang, D.; Liang, Q.Y.; Qin, Y.; Tang, L.; Zhao, H.; Stenvang, J.; Li, Y.; et al. Metagenomic analysis of faecal microbiome as a tool towards targeted non-invasive biomarkers for colorectal cancer. Gut 2017, 66, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Feng, Q.; Liang, S.; Jia, H.; Stadlmayr, A.; Tang, L.; Lan, Z.; Zhang, D.; Xia, H.; Xu, X.; Jie, Z.; et al. Gut microbiome development along the colorectal adenoma-carcinoma sequence. Nat Commun. 2015, 6, 6528. [Google Scholar] [CrossRef]
- Zhou, Y.; He, H.; Xu, H.; Li, Y.; Li, Z.; Du, Y.; He, J.; Zhou, Y.; Wang, H.; Nie, Y. Association of oncogenic bacteria with colorectal cancer in South China. Oncotarget 2016, 7, 80794–80802. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Cai, G.; Qiu, Y.; Fei, N.; Zhang, M.; Pang, X.; Jia, W.; Cai, S.; Zhao, L. Structural segregation of gut microbiota between colorectal cancer patients and healthy volunteers. ISME J. 2012, 6, 320–329. [Google Scholar] [CrossRef] [PubMed]
- Sabatino, L.; Pancione, M.; Votino, C.; Colangelo, T.; Lupo, A.; Novellino, E.; Lavecchia, A.; Colantuoni, V. Emerging role of the β-catenin-PPARγ axis in the pathogenesis of colorectal cancer. World J. Gastroenterol. 2014, 20, 7137–7151. [Google Scholar] [CrossRef] [PubMed]
- Peyrin-Biroulet, L.; Beisner, J.; Wang, G.; Nuding, S.; Oommen, S.T.; Kelly, D.; Parmentier-Decrucq, E.; Dessein, R.; Merour, E.; Chavatte, P.; et al. Peroxisome proliferator-activated receptor gamma activation is required for maintenance of innate antimicrobial immunity in the colon. Proc. Natl. Acad. Sci. USA 2010, 107, 8772–8777. [Google Scholar] [CrossRef] [Green Version]
- den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.-J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [Green Version]
- McLoughlin, R.F.; Berthon, B.S.; Jensen, M.E.; Baines, K.J.; Wood, L.G. Short-chain fatty acids, prebiotics, synbiotics, and systemic inflammation: A systematic review and meta-analysis. Am. J. Clin. Nutr. 2017, 106, 930–945. [Google Scholar] [CrossRef] [PubMed]
- Le Poul, E.; Loison, C.; Struyf, S.; Springael, J.Y.; Lannoy, V.; Decobecq, M.E.; Brezillon, S.; Dupriez, V.; Vassart, G.; Van Damme, J.; et al. Functional characterization of human receptors for short chain fatty acids and their role in polymorphonuclear cell activation. J. Biol. Chem. 2003, 278, 25481–25489. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.M.; de Souza, R.; Kendall, C.W.; Emam, A.; Jenkins, D.J. Colonic health: Fermentation and short chain fatty acids. J. Clin. Gastroenterol. 2006, 40, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Kespohl, M.; Vachharajani, N.; Luu, M.; Harb, H.; Pautz, S.; Wolff, S.; Sillner, N.; Walker, A.; Schmitt-Kopplin, P.; Boettger, T.; et al. The Microbial Metabolite Butyrate Induces Expression of Th1-Associated Factors in CD4+ T Cells. Front. Immunol. 2017, 8, 1036. [Google Scholar] [CrossRef] [PubMed]
- Thirabunyanon, M.; Hongwittayakorn, P. Potential probiotic lactic acid bacteria of human origin induce antiproliferation of colon cancer cells via synergic actions in adhesion to cancer cells and short-chain fatty acid bioproduction. Appl. Biochem. Biotechnol. 2013, 169, 511–525. [Google Scholar] [CrossRef]
- Velayos, F.S.; Liu, L.; Lewis, J.D.; Allison, J.E.; Flowers, N.; Hutfless, S.; Abramson, O.; Perry, G.S.; Herrinton, L.J. Prevalence of colorectal cancer surveillance for ulcerative colitis in an integrated health care delivery system. Gastroenterology 2010, 139, 1511–1518. [Google Scholar] [CrossRef]
- Hijova, E.; Chmelarova, A. Short chain fatty acids and colonic health. Bratisl. Lek. Listy. 2007, 108, 354–358. [Google Scholar]
- Cummings, J.H.; Pomare, E.W.; Branch, W.J.; Naylor, C.P.; Macfarlane, G.T. Short chain fatty acids in human large intestine, portal, hepatic and venous blood. Gut 1987, 28, 1221–1227. [Google Scholar] [CrossRef]
- Tan, J.; McKenzie, C.; Potamitis, M.; Thorburn, A.N.; Mackay, C.R.; Macia, L. The role of short-chain fatty acids in health and disease. Adv. Immunol. 2014, 121, 91–119. [Google Scholar]
- Garner, C.E.; Smith, S.; de Lacy Costello, B.; White, P.; Spencer, R.; Probert, C.S.; Ratcliffe, N.M. Volatile organic compounds from feces and their potential for diagnosis of gastrointestinal disease. FASEB J. 2007, 21, 1675–1688. [Google Scholar] [CrossRef] [Green Version]
- Singh, M.P.; Rai, S.; Suyal, S.; Singh, S.K.; Singh, N.K.; Agarwal, A.; Srivastava, S. Genetic and epigenetic markers in colorectal cancer screening: Recent advances. Expert Rev. Mol. Diagn. 2017, 17, 665–685. [Google Scholar] [CrossRef] [PubMed]
- González-Sarrías, A.; Giménez-Bastida, J.A.; Núñez-Sánchez, M.Á.; Larrosa, M.; García-Conesa, M.T.; Tomás-Barberán, F.A.; Espín, J.C. Phase-II metabolism limits the antiproliferative activity of urolithins in human colon cancer cells. Eur. J. Nutr. 2014, 53, 853–864. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi, E.S.; Li, X.Q.; Ghorbani, M.; Azadi, B.; Kubow, S. Chlorogenic Acid and Its Microbial Metabolites Exert Anti-Proliferative Effects, S-Phase Cell-Cycle Arrest and Apoptosis in Human Colon Cancer Caco-2 Cells. Int. J. Mol. Sci. 2018, 19, 723. [Google Scholar] [CrossRef] [PubMed]
- Hullar, M.A.; Burnett-Hartman, A.N.; Lampe, J.W. Gut microbes, diet, and cancer. Cancer Treat Res. 2014, 159, 377–399. [Google Scholar]
- Myzak, M.C.; Hardin, K.; Yan, M.; Tong, P.; Dashwood, R.; Ho, E. Sulforaphane inhibits HDAC activity in prostate cancer cells, retards growth of PC3 xenografts, and inhibits HDAC activity in vivo. FASEB J. 2006, 20, A150. [Google Scholar]
- Archimbaud, C.; Shankar, N.; Forestier, C.; Baghdayan, A.; Gilmore, M.S.; Charbonné, F.; Joly, B. In vitro adhesive properties and virulence factors of Enterococcus faecalisstrains. Res. Microbiol. 2002, 153, 75–80. [Google Scholar] [CrossRef]
- Creti, R.; Imperi, M.; Bertuccini, L.; Fabretti, F.; Orefici, G.; Di Rosa, R.; Baldassarri, L. Survey for virulence determinants among Enterococcus faecalis isolated from different sources. J. Med. Microbiol. 2004, 53, 13–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Donors (Healthy—HD Colon Cancer—CC) | Age/Gender (♀/♂) | Histotype/Stage |
|---|---|---|
| HD01 | 57 ♀ | n/a |
| HD02 | 60 ♂ | n/a |
| HD03 | 56 ♂ | n/a |
| HD04 | 53 ♀ | n/a |
| HD05 | 68 ♀ | n/a |
| HD06 | 65 ♂ | n/a |
| HD07 | 68 ♀ | n/a |
| HD08 | 48 ♂ | n/a |
| HD09 | 52 ♂ | n/a |
| CC01 | 78 ♂ | Colorectal adenocarcinoma with moderate differentiation (pT3N0) |
| CC02 | 79 ♂ | Colorectal adenocarcinoma with moderate differentiation (pT3N0) |
| CC03 | 68 ♀ | Colorectal adenocarcinoma with moderate differentiation (pT2N0) |
| CC04 | 78 ♂ | Colon intramucosal adenocarcinoma (pT2N0) |
| CC05 | 40 ♂ | Colorectal adenocarcinoma with moderate differentiation (pT3aN1aMx) |
| CC06 | 78 ♂ | Colorectal adenocarcinoma with moderate differentiation (pT1N0) |
| CC07 | 81 ♀ | Colorectal adenocarcinoma with moderate differentiation (pT2N0Mx) |
| CC08 | 62 ♂ | Colorectal adenocarcinoma with moderate differentiation (pT2N0Mx) |
| CC09 | 63 ♂ | Colorectal adenocarcinoma with moderate differentiation (pT3N0Mx) |
| Cell Line | Medium | Other Information |
|---|---|---|
| HCT-116 | DMEM + 2 mM l-Glutamine + 100 U/mL Penicillin + 100 μg/mL Streptomycin + 10% FBS | (CCL-247™) ATCC®, Manassas, VA, USA. Colon; colorectal carcinoma, male, epithelial, primary tumor. |
| HCT-8 | DMEM + 2 mM l-Glutamine + 100 U/mL Penicillin + 100 μg/mL Streptomycin + 10% FBS | (CCL-244™) ATCC®, Manassas, VA, USA. Colon; ileocecal colorectal adenocarcinoma; primary tumor, epithelial. |
| SW-620 | DMEM + 2 mM l-Glutamine + 100 U/mL Penicillin + 100 μg/mL Streptomycin + 10% FBS | (CCL-227™) ATCC®, Manassas, VA, USA. Colon; derived from metastatic site: lymph node; Dukes’ type C, colorectal adenocarcinoma; male; epithelial |
| CLR-1790 | 50% DMEM + 50% Ham’s F12 Nutrient Mixture + 2 mM l-Glutamine + 100 U/mL Penicillin + 100 μg/mL Streptomycin + 10% FBS | (CCD 841 CoN) ATCC®, Manassas, VA, USA. Colon; normal; 21 weeks gestation fetus; epithelial. |
| Donors | E. faecalis/Total Enterococci | E. faecalis Strains Used | ST | |
|---|---|---|---|---|
| CRC patients | CC1 | -/- | - | |
| CC2 | -/- | - | ||
| CC3 | -/- | - | ||
| CC4 | -/- | - | ||
| CC5 | -/- | - | ||
| CC6 | 2/3 | CRC01 | 40 | |
| CC7 | -/- | - | ||
| CC8 | 2/2 | CRC02 | 59 | |
| CC9 | -/- | - | ||
| Healthy donors | HD1 | 3/8 | EFH01 | 21 |
| HD2 | 4/10 | EFH02 | 47 | |
| EFH05 | 21 * | |||
| EFH06 | 16 | |||
| HD3 | 3/8 | EFH03 | unk | |
| HD4 | -/- | - | ||
| HD5 | -/4 | - | ||
| HD6 | 2/5 | EFH04 | 209 | |
| HD7 | -/5 | - | ||
| HD8 | -/3 | - | ||
| HD9 | -/6 | - | ||
| Total isolated strains | 16/54 | 8 |
| Samples | Acetic ± SD (ug/mL) | Propionic ± SD (ug/mL) | Butyric ± SD (ug/mL) | Iso-Butyric ± SD (ug/mL) | Iso-Valeric ± SD (ug/mL) | 2-MethylButyric ± SD (ug/mL) | Valeric ± SD (ug/mL) |
|---|---|---|---|---|---|---|---|
| RPMI | n.q. | n.q. | n.q. | n.q. | n.q. | n.q. | n.q. |
| EFH01 | 393.5 ± 71.1 | n.q. | n.q. | n.q. | 4.7 ± 1.0 | 4.2 ± 0.8 | n.q. |
| EFH02 | 268.7 ± 19.7 | 2.7 ± 2.5 | 4.2 ± 1.1 | n.q. | 3.3 ± 0.4 | 2.8 ± 0.2 | 0.3 ± 0.6 |
| EFH03 | 379.7 ± 43.6 | n.q | 2.1 ± 1.8 | n.q. | 3.3 ± 0.5 | 3.3 ± 0.4 | n.q. |
| EFH04 | 324.2 ± 23.9 | n.q | 1.0 ± 1.7 | n.q. | 4.8 ± 0.2 | 3.6 ± 0.2 | n.q. |
| EFH05 | 394.1 ± 56.1 | 0.8 ± 1.4 | 1.0 ± 1.7 | n.q. | 4.6 ± 0.4 | 4.1 ± 0.4 | n.q. |
| EFH06 | 381.2 ± 16.1 | n.q. | n.q. | n.q. | 6.4 ± 0.5 | 6.1 ± 0.6 | n.q. |
| CRC01 | 417.5 ± 89.3 | 1.2 ± 2.1 | 2.6 ± 2.3 | n.q. | 4.2 ± 0.9 | 4.0 ± 0.8 | n.q. |
| CRC02 | 399.7 ± 44.5 | n.q. | 3.3 ± 0.1 | n.q. | 3.7 ± 1.1 | 2.4 ± 0.4 | n.q. |
| ATCC | 380.6 ± 45.9 | n.q. | n.q. | n.q. | 4.7 ± 0.8 | 3.5 ± 0.6 | n.q. |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Almeida, C.V.; Lulli, M.; di Pilato, V.; Schiavone, N.; Russo, E.; Nannini, G.; Baldi, S.; Borrelli, R.; Bartolucci, G.; Menicatti, M.; et al. Differential Responses of Colorectal Cancer Cell Lines to Enterococcus faecalis’ Strains Isolated from Healthy Donors and Colorectal Cancer Patients. J. Clin. Med. 2019, 8, 388. https://doi.org/10.3390/jcm8030388
De Almeida CV, Lulli M, di Pilato V, Schiavone N, Russo E, Nannini G, Baldi S, Borrelli R, Bartolucci G, Menicatti M, et al. Differential Responses of Colorectal Cancer Cell Lines to Enterococcus faecalis’ Strains Isolated from Healthy Donors and Colorectal Cancer Patients. Journal of Clinical Medicine. 2019; 8(3):388. https://doi.org/10.3390/jcm8030388
Chicago/Turabian StyleDe Almeida, Carolina Vieira, Matteo Lulli, Vincenzo di Pilato, Nicola Schiavone, Edda Russo, Giulia Nannini, Simone Baldi, Rossella Borrelli, Gianluca Bartolucci, Marta Menicatti, and et al. 2019. "Differential Responses of Colorectal Cancer Cell Lines to Enterococcus faecalis’ Strains Isolated from Healthy Donors and Colorectal Cancer Patients" Journal of Clinical Medicine 8, no. 3: 388. https://doi.org/10.3390/jcm8030388