
The First Evidence of Bacterial Foci in the Hair Part and Dermal Papilla of Scalp Hair Follicles: A Pilot Comparative Study in Alopecia Areata
 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Results
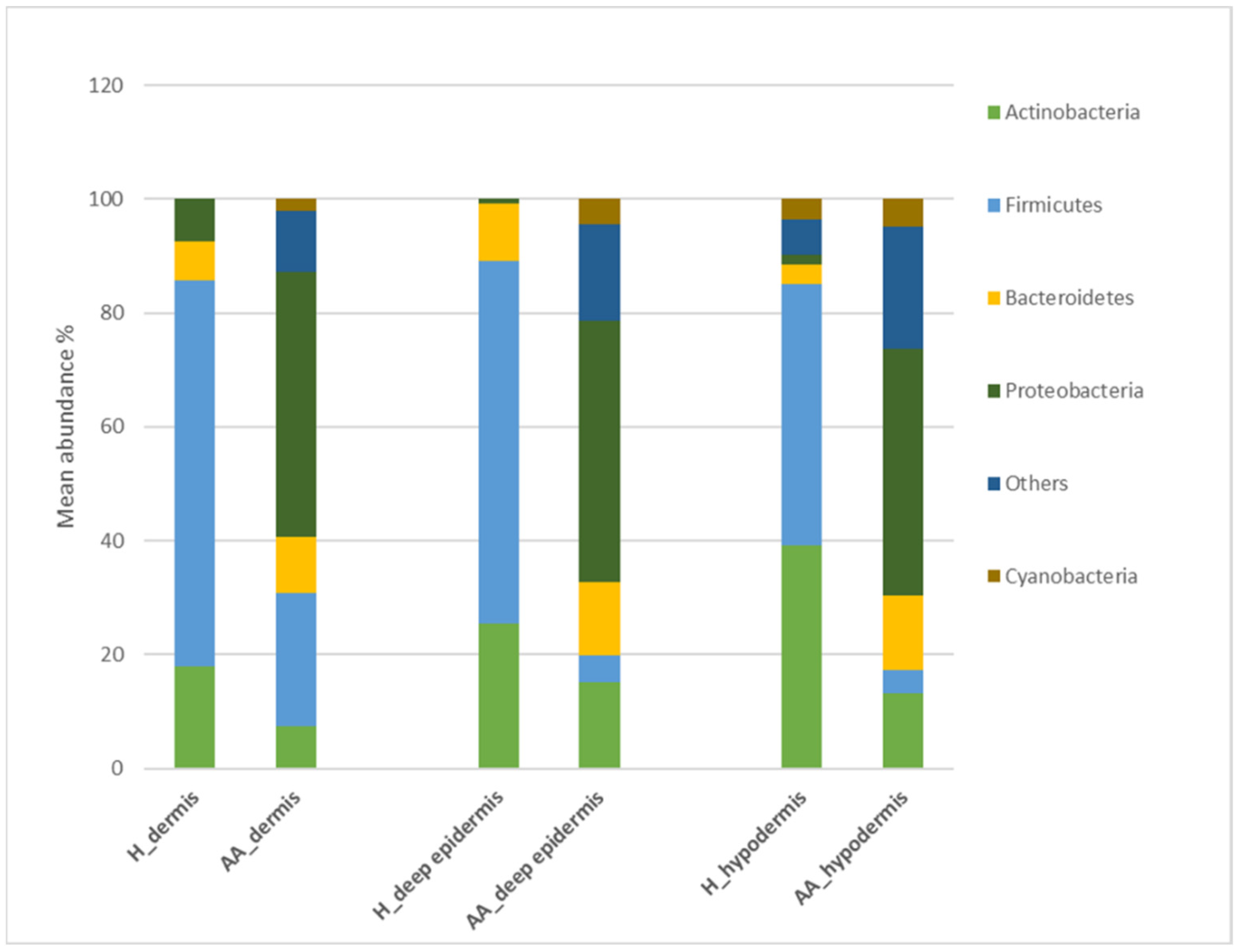
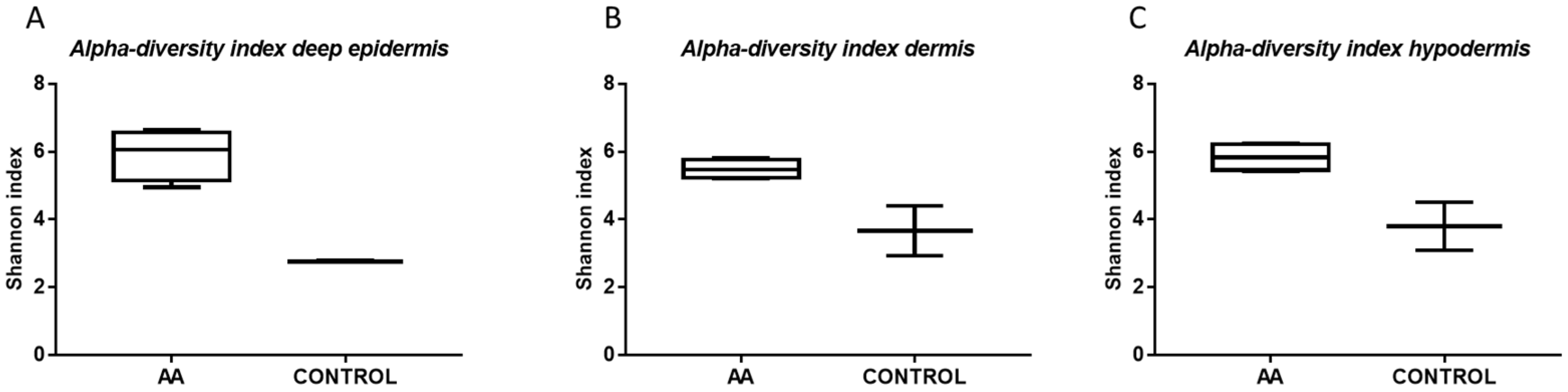
2.1. Microbiota Profiling of the Scalp in AA Patients
2.2. Bacterial Infection in AA Subjects
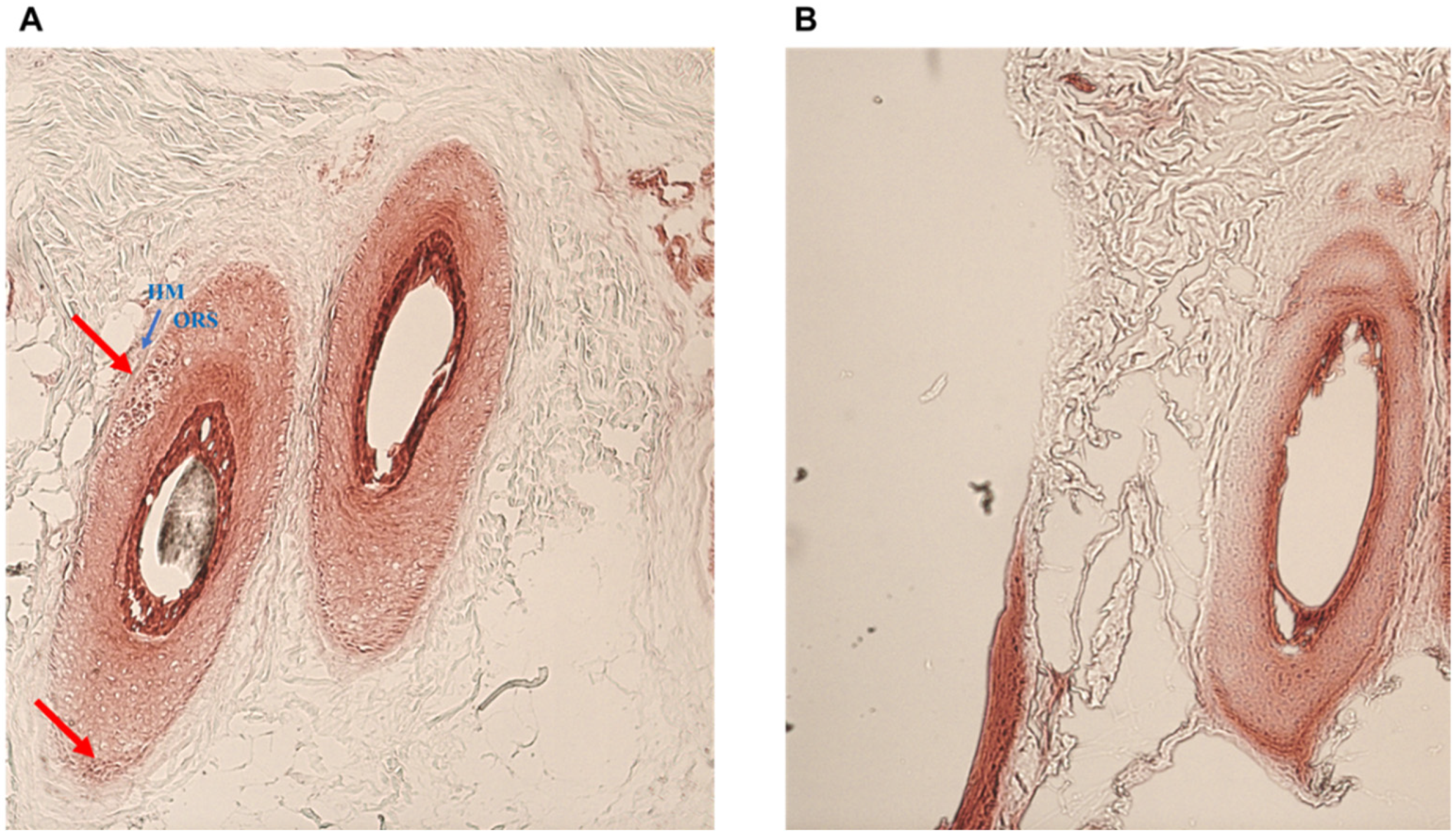
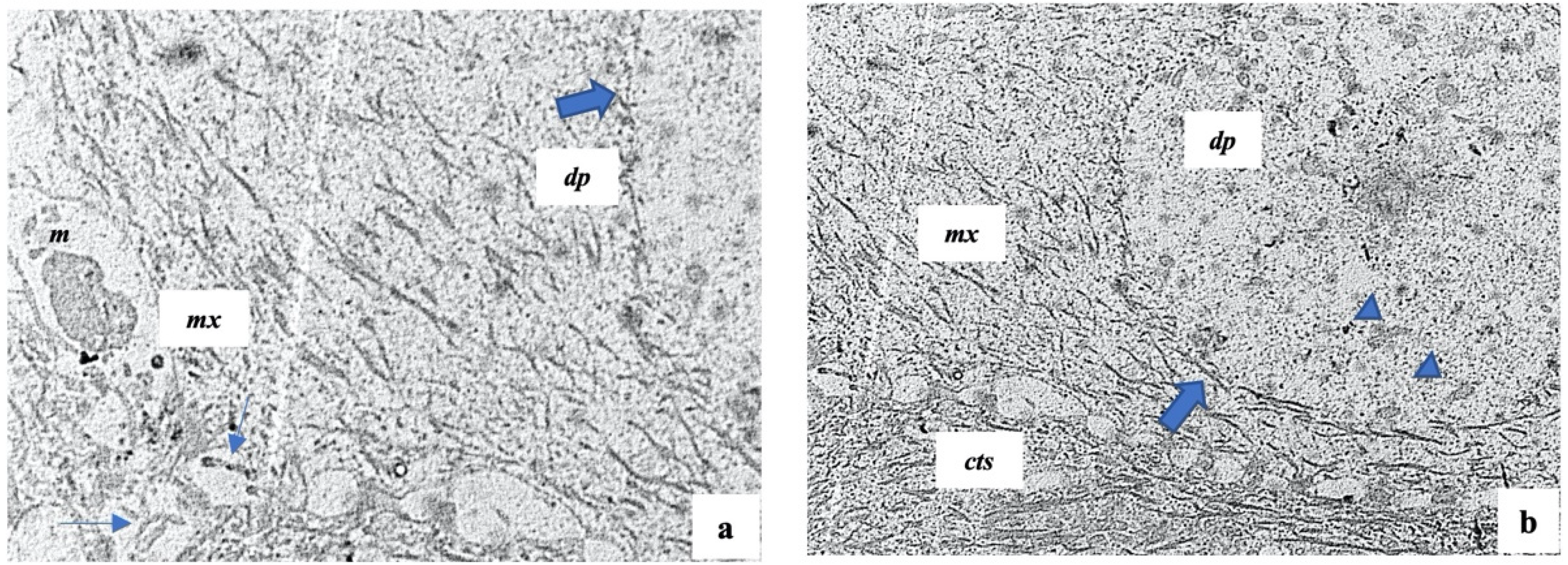
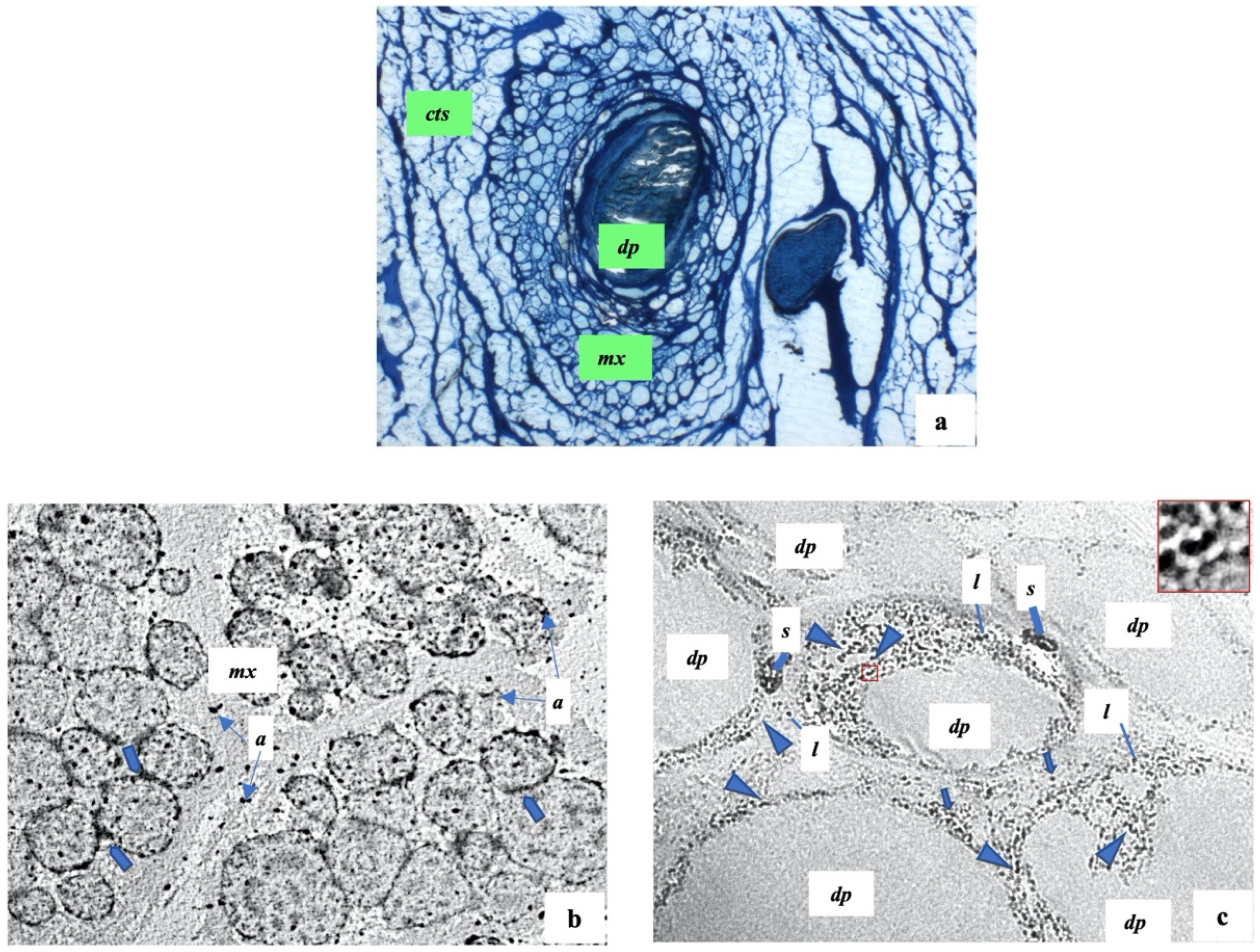
2.3. Histology and Ultrastructure of Normal and Alopecia Aerate Scalp Hair Bulb
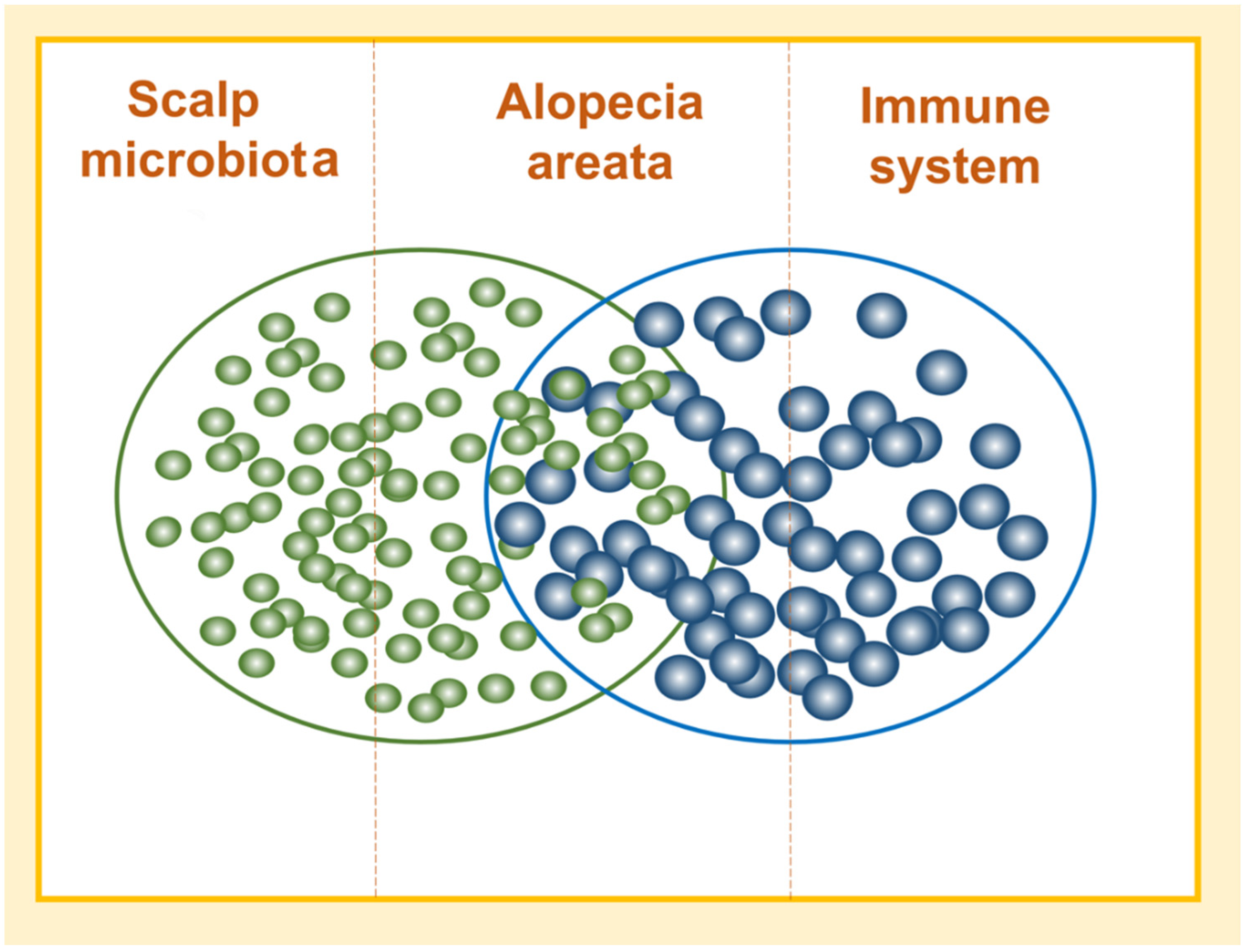
3. Discussion
4. Materials and Methods
4.1. Subjects’ Recruitment
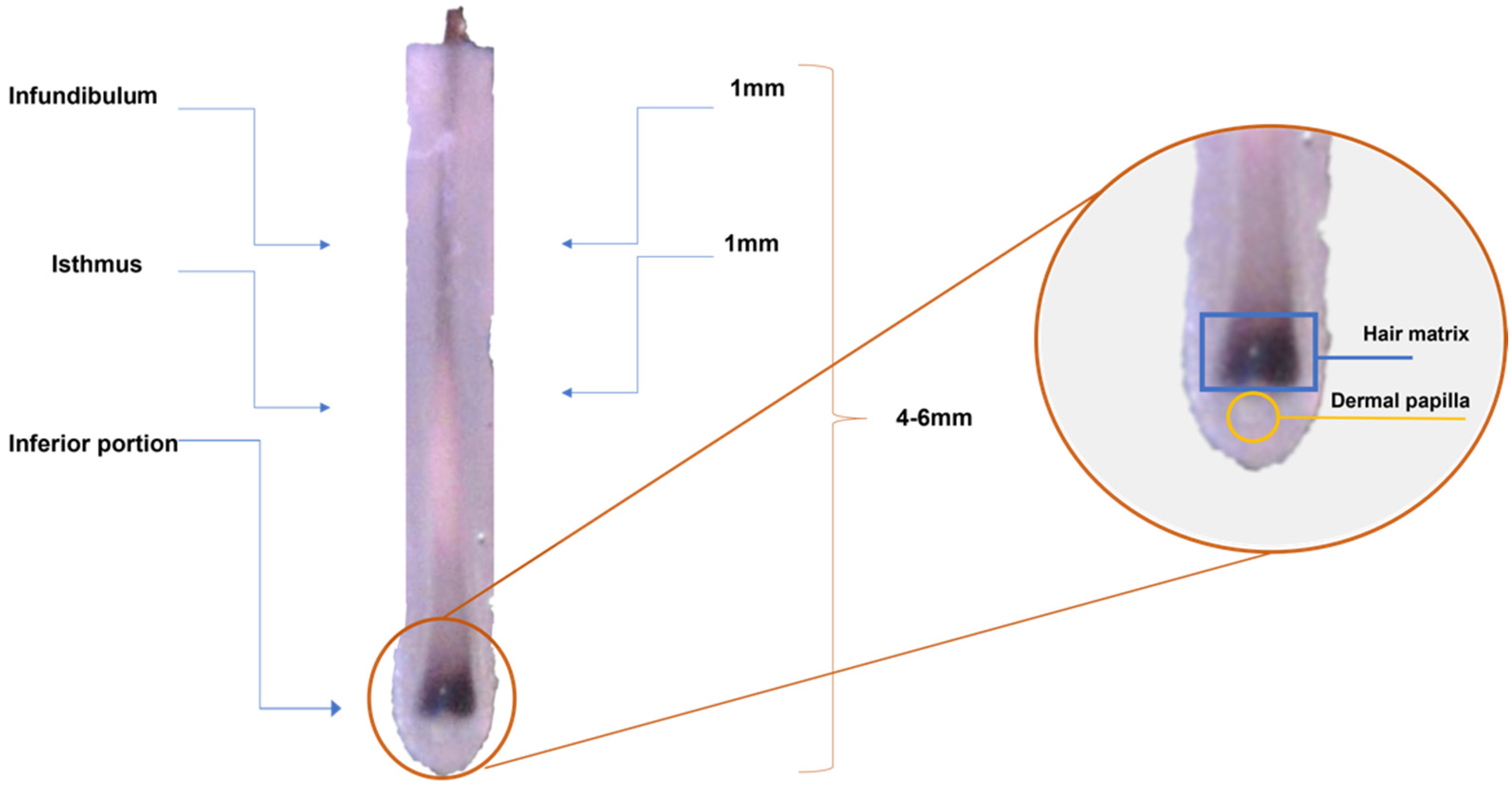
4.2. Sample Collection
4.3. DNA Extraction and 16S Amplicon Generation, Sequencing, and Analysis-Illumina Sequencing
4.4. Transmission Electron Microscopy (TEM)
4.5. Tissue Gram Staining
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Villasante Fricke, A.C.; Miteva, M. Epidemiology and burden of alopecia areata: A systematic review. Clin. Cosmet. Investig. Dermatol. 2015, 8, 397–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harries, M.J.; Meyer, K.C.; Chaudhry, I.H.; Griffiths, C.E.; Paus, R. Does collapse of immune privilege in the hair-follicle bulge play a role in the pathogenesis of primary cicatricial alopecia? Clin. Exp. Dermal. 2010, 35, 637–644. [Google Scholar] [CrossRef] [PubMed]
- Paus, R.; Bertolini, M. The role of hair follicle immune privilege collapse in alopecia areata: Status and perspectives. J. Investig. Dermatol. Symp. Proc. 2013, 16, S25–S27. [Google Scholar] [CrossRef]
- Bertolini, M.; McElwee, K.; Gilhar, A.; Bulfone-Paus, S.; Paus, R. Hair follicle immune privilege and its collapse in alopecia areata. Exp. Dermatol. 2020, 29, 703–725. [Google Scholar] [CrossRef] [PubMed]
- Paus, R.; Bulfone-Paus, S.; Bertolini, M. Hair Follicle Immune Privilege Revisited: The Key to Alopecia Areata Management. J. Investig. Dermatol. Symp. Proc. 2018, 19, S12–S17. [Google Scholar] [CrossRef] [Green Version]
- Constantinou, A.; Kanti, V.; Polak-Witka, K.; Blume-Peytavi, U.; Spyrou, G.M.; Vogt, A. The Potential Relevance of the Microbiome to Hair Physiology and Regeneration: The Emerging Role of Metagenomics. Biomedicines 2021, 9, 236. [Google Scholar] [CrossRef] [PubMed]
- Rinaldi, F.; Pinto, D.; Marzani, B.; Rucco, M.; Giuliani, G.; Sorbellini, E. Human microbiome: What’s new in scalp diseases. J. Transl. Sci. 2018, 4, 1–4. [Google Scholar]
- Naik, P.P.; Farrukh, S.N. Association between alopecia areata and thyroid dysfunction. Postgrad. Med. 2021, 133, 895–898. [Google Scholar] [CrossRef]
- Puavilai, S.; Puavilai, G.; Charuwichitratana, S.; Sakuntabhai, A.; Sriprachya-Anunt, S. Prevalence of thyroid diseases in patients with alopecia areata. Int. J. Dermatol. 1994, 33, 632–633. [Google Scholar] [CrossRef]
- Lewiński, A.; Broniarczyk-Dyła, G.; Sewerynek, E.; Zerek-Mełeń, G.; Szkudliński, M. Abnormalities in structure and function of the thyroid gland in patients with alopecia areata. J. Am. Acad. Dermatol. 1990, 23, 768–769. [Google Scholar] [CrossRef]
- Thomas, E.A.; Kadyan, R.S. Alopecia areata and autoimmunity: A clinical study. Indian J. Dermatol. 2008, 53, 70–74. [Google Scholar] [CrossRef]
- Cogen, A.L.; Nizet, V.; Gallo, R.L. Skin microbiota: A source of disease or defence? Br. J. Dermatol. 2008, 158, 442–455. [Google Scholar] [CrossRef] [Green Version]
- Byrd, A.; Belkaid, Y.; Segre, J. The human skin microbiome. Nat. Rev. Microbiol. 2018, 16, 143–155. [Google Scholar] [CrossRef]
- Lousada, M.B.; Lachnit, T.; Edelkamp, J.; Rouillé, T.; Ajdic, D.; Uchida, Y.; Di Nardo, A.; Bosch, T.C.G.; Paus, R. Exploring the human hair follicle microbiome. Br. J. Dermatol. 2021, 184, 802–815. [Google Scholar] [CrossRef]
- Constantinou, A.; Polak-Witka, K.; Tomazou, M.; Oulas, A.; Kanti, V.; Schwarzer, R.; Helmuth, J.; Edelmann, A.; Blume-Peytavi, U.; Spyrou, G.M.; et al. Dysbiosis and Enhanced Beta-Defensin Production in Hair Follicles of Patients with Lichen Planopilaris and Frontal Fibrosing Alopecia. Biomedicines 2021, 9, 266. [Google Scholar] [CrossRef]
- Ho, B.S.; Ho, E.X.P.; Chu, C.W.; Ramasamy, S.; Bigliardi-Qi, M.; de Sessions, P.F.; Bigliardi, P.L. Microbiome in the hair follicle of androgenetic alopecia patients. PLoS ONE 2019, 14, e0216330. [Google Scholar] [CrossRef] [Green Version]
- Carmona-Cruz, S.; Orozco-Covarrubias, L.; Sáez-de-Ocariz, M. The Human Skin Microbiome in Selected Cutaneous Diseases. Front. Cell. Infect. Microbiol. 2022, 12, 834135. [Google Scholar] [CrossRef]
- Zheng, D.; Liwinski, T.; Elinav, E. Interaction between microbiota and immunity in health and disease. Cell Res. 2020, 30, 492–506. [Google Scholar] [CrossRef]
- Racine, P.J.; Janvier, X.; Clabaut, M.; Catovic, C.; Souak, D.; Boukerb, A.M.; Groboillot, A.; Konto-Ghiorghi, Y.; Duclairoir-Poc, C.; Lesouhaitier, O.; et al. Dialog between skin and its microbiota: Emergence of “Cutaneous Bacterial Endocrinology”. Exp. Dermatol. 2020, 29, 790–800. [Google Scholar] [CrossRef]
- Kabashima, K.; Honda, T.; Ginhoux, F.; Egawa, G. The immunological anatomy of the skin. Nat. Rev. Immunol. 2019, 19, 19–30. [Google Scholar] [CrossRef]
- Belkaid, Y.; Segre, J.A. Dialogue between skin microbiota and immunity. Science 2014, 346, 954–959. [Google Scholar] [CrossRef]
- Sanford, J.A.; O’Neill, A.M.; Zouboulis, C.C.; Gallo, R.L. Short-Chain Fatty Acids from Cutibacterium acnes Activate Both a Canonical and Epigenetic Inflammatory Response in Human Sebocytes. J. Immunol. 2019, 202, 1767–1776. [Google Scholar] [CrossRef] [Green Version]
- Pinto, D.; Sorbellini, E.; Marzani, B.; Rucco, M.; Giuliani, G.; Rinaldi, F. Scalp bacterial shift in Alopecia areata. PLoS ONE 2019, 14, e0215206. [Google Scholar] [CrossRef] [Green Version]
- Pinto, D.; Calabrese, F.M.; De Angelis, M.; Celano, G.; Giuliani, G.; Gobbetti, M.; Rinaldi, F. Predictive Metagenomic Profiling, Urine Metabolomics, and Human Marker Gene Expression as an Integrated Approach to Study Alopecia Areata. Front. Cell. Infect. Microbiol. 2020, 10, 146. [Google Scholar] [CrossRef]
- Naik, S.; Bouladoux, N.; Wilhelm, C.; Molloy, M.J.; Salcedo, R.; Kastenmuller, W.; Deming, C.; Quinones, M.; Koo, L.; Conlan, S.; et al. Compartmentalized control of skin immunity by resident commensals. Science 2012, 337, 1115–1119. [Google Scholar] [CrossRef] [Green Version]
- Campbell, D.J.; Koch, M.A. Living in Peace: Host-Microbiota Mutualism in the Skin. Cell Host Microbe 2017, 4, 419–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scharschmidt, T.C.; Vasquez, K.S.; Pauli, M.L.; Leitner, E.G.; Chu, K.; Truong, H.A.; Lowe, M.M.; Sanchez Rodriguez, R.; Ali, N.; Laszik, Z.G.; et al. Commensal Microbes and Hair Follicle Morphogenesis Coordinately Drive Treg Migration into Neonatal Skin. Cell Host Microbe 2017, 21, 467–477.e5. [Google Scholar] [CrossRef] [Green Version]
- Polak-Witka, K.; Rudnicka, L.; Blume-Peytavi, U.; Vogt, A. The role of the microbiome in scalp hair follicle biology and disease. Exp. Dermatol. 2020, 29, 286–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rolig, A.S.; Carter, J.E.; Ottemann, K.M. Bacterial chemotaxis modulates host cell apoptosis to establish a T-helper cell, type 17 (Th17)-dominant immune response in Helicobacter pylori infection. Proc. Natl. Acad. Sci. USA 2011, 108, 19749–19754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paus, R.; Slominski, A.; Czarnetzki, B.M. Is alopecia areata an autoimmune-response against melanogenesis-related proteins, exposed by abnormal MHC class I expression in the anagen hair bulb? Yale J. Biol. Med. 1993, 66, 541–554. [Google Scholar] [PubMed]
- Crew, P.E.; McNamara, L.; Waldron, P.E.; McCulley, L.; Jones, S.C.; Bersoff-Matcha, S.J. Unusual Neisseria species as a cause of infection in patients taking eculizumab. J. Infect. 2019, 78, 113–118. [Google Scholar] [CrossRef]
- Humbert, M.V.; Christodoulides, M. Atypical, Yet Not Infrequent, Infections with Neisseria Species. Pathogens 2019, 9, 10. [Google Scholar] [CrossRef] [Green Version]
- Boldock, E.; Surewaard, B.G.J.; Shamarina, D.; Na, M.; Fei, Y.; Ali, A.; Williams, A.; Pollitt, E.J.G.; Szkuta, P.; Morris, P.; et al. Human skin commensals augment Staphylococcus aureus pathogenesis. Nat. Microbiol. 2018, 3, 881–890. [Google Scholar] [CrossRef] [Green Version]
- Jahns, A.C.; Lundskog, B.; Ganceviciene, R.; Palmer, R.H.; Golovleva, I.; Zouboulis, C.C.; McDowell, A.; Patrick, S.; Alexeyev, O.A. An increased incidence of Propionibacterium acnes biofilms in acne vulgaris: A case-control study. Br. J. Dermatol. 2012, 167, 50–58. [Google Scholar] [CrossRef]
- Matard, B.; Meylheuc, T.; Briandet, R.; Casin, I.; Assouly, P.; Cavelier-balloy, B.; Reygagne, P. First evidence of bacterial biofilms in the anaerobe part of scalp hair follicles: A pilot comparative study in folliculitis decalvans. J. Eur. Acad. Dermatol. Venereol. 2013, 27, 853–860. [Google Scholar] [CrossRef]
- Thaiss, C.A.; Zmora, N.; Levy, M.; Elinav, E. The microbiome and innate immunity. Nature 2016, 535, 65–74. [Google Scholar] [CrossRef]
- Lavrinienko, A.; Tukalenko, E.; Mappes, T. Skin and gut microbiomes of a wild mammal respond to different environmental cues. Microbiome 2018, 6, 209. [Google Scholar] [CrossRef] [PubMed]
- Negroni, A.; Pierdomenico, M.; Cucchiara, S.; Stronati, L. NOD2 and inflammation: Current insights. J. Inflamm. Res. 2018, 11, 49–60. [Google Scholar] [CrossRef] [Green Version]
- Garzorz, N.; Alsisi, M.; Todorova, A.; Atenhan, A.; Thomas, J.; Lauffer, F.; Ring, J.; Schmidt-Weber, C.; Biedermann, T.; Eyerich, S.; et al. Dissecting susceptibility from exogenous triggers: The model of alopecia areata and associated inflammatory skin diseases. J. Eur. Acad. Dermatol. Venereol. 2015, 29, 2429–2435. [Google Scholar] [CrossRef] [PubMed]
- Tharumanathan, S. Understanding the biological mechanism of alopecia areata. Am. J. Dermatol. Venereol 2015, 4, 1–4. [Google Scholar] [CrossRef]
- Hull, C.M.; Nickolay, L.E.; Estorninho, M.; Richardson, M.W.; Riley, J.L.; Peakman, M.; Maher, J.; Tree, T.I. Generation of human islet-specific regulatory T cells by TCR gene transfer. J. Autoimmun. 2017, 79, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Messenger, A.G.; Bleehen, S.S. Alopecia areata: Light and electron microscopic pathology of the regrowing white hair. Br. J. Dermatol. 1984, 110, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Pratt, C.H.; King, L.E., Jr.; Messenger, A.G.; Christiano, A.M.; Sundberg, J.P. Alopecia areata. Nat. Rev. Dis. Prim. 2017, 3, 17011. [Google Scholar] [CrossRef] [Green Version]
- Coroaba, A.; Chiriac, A.E.; Sacarescu, L.; Pinteala, T.; Minea, B.; Ibanescu, S.A.; Pertea, M.; Moraru, A.; Esanu, I.; Maier, S.S.; et al. New insights into human hair: SAXS, SEM, TEM and EDX for Alopecia Areata investigations. PeerJ 2020, 8, e8376. [Google Scholar] [CrossRef] [Green Version]
- Olsen, E.A.; Hordinsky, M.K.; Price, V.H.; Roberts, J.L.; Shapiro, J.; Canfield, D.; Duvic, M.; King, L.E., Jr.; McMichael, A.J.; Randall, V.A.; et al. National Alopecia Areata Foundation. Alopecia areata investigational assessment guidelines–Part II. national alopecia areata foundation. J. Am. Acad. Dermatol. 2004, 51, 440–447. [Google Scholar] [CrossRef]
- Gao, Z.; Perez-Perez, G.I.; Chen, Y.; Blaser, M.J. Quantitation of major human cutaneous bacterial and fungal populations. J. Clin. Microbiol. 2010, 48, 3575–3581. [Google Scholar] [CrossRef] [Green Version]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Takahashi, S.; Tomita, J.; Nishioka, K.; Hisada, T.; Nishijima, M. Development of a prokaryotic universal primer for simultaneous analysis of Bacteria and Archaea using next-generation sequencing. PLoS ONE 2014, 9, e105592. [Google Scholar] [CrossRef] [Green Version]
- Apprill, A.; McNally, S.; Parsons, R.; Weber, L. Minor revision to V4 region SSU rRNA 806R gene primer greatly increases detection of SAR11 bacterioplankton. Aquat. Microb. Ecol. 2015, 75, 129–137. [Google Scholar] [CrossRef] [Green Version]
- Parada, A.E.; Needham, D.M.; Fuhrman, J.A. Every base matters: Assessing small subunit rRNA primers for marine microbiomes with mock communities, time series and global field samples. Environ. Microbiol. 2016, 18, 1403–1414. [Google Scholar] [CrossRef]
- Walters, W.; Hyde, E.R.; Berg-Lyons, D.; Ackermann, G.; Humphrey, G.; Parada, A. Improved Bacterial 16S rRNA Gene (V4 and V4-5) and Fungal Internal Transcribed Spacer Marker Gene Primers for Microbial Community Surveys. mSystems 2016, 1, e00009-15. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2011, 108, 4516–4522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a dual-index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the MiSeq Illumina sequencing platform. Appl. Environ. Microbiol. 2013, 79, 5112–5120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Desantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a chimerachecked 16S rRNA gene database and workbench compatible with ARB. Appl. Environ. Microbiol. 2006, 72, 5069–5072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tobin, D.J. Morphological analysis of hair follicles in alopecia areata. Microsc. Res. Tech. 1997, 38, 443–451. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Layer | Taxa Level | Feature | Healthy | AA | p-Value |
|---|---|---|---|---|---|
| deep epidermis | family | Enterobacteriaceae | 0.00 | 17.09 | 0.500 |
| deep epidermis | genus | Streptococcus | 2.45 | 16.30 | 0.551 |
| deep epidermis | genus | Gemella | 0.00 | 2.40 | 0.500 |
| deep epidermis | genus | Porphyromonas | 0.00 | 2.14 | 0.500 |
| deep epidermis | genus | Granulicatella | 0.00 | 2.50 | 0.500 |
| deep epidermis | genus | Staphylococcus | 49.60 | 0.00 | 0.227 |
| deep epidermis | genus | Flavobacterium | 8.00 | 0.00 | 0.156 |
| dermis | phylum | Firmicutes | 71.92 | 22.50 | 0.009 |
| dermis | phylum | Proteobacteria | 7.48 | 46.55 | 0.027 |
| dermis | family | Micrococcaceae | 0.00 | 6.41 | 0.137 |
| dermis | genus | Staphylococcus | 48.35 | 1.35 | 0.278 |
| dermis | genus | Flavobacterium | 5.00 | 0.00 | 0.275 |
| hypodermis | phylum | Firmicutes | 45.75 | 4.05 | 0.033 |
| hypodermis | phylum | Proteobacteria | 1.8 | 43.35 | 0.009 |
| hypodermis | family | Bacteroidetes | 2.96 | 12.74 | 0.015 |
| hypodermis | family | Micrococcaceae | 0.00 | 4.85 | 0.06 |
| hypodermis | family | Neisseriaceae | 0.00 | 1.00 | 0.500 |
| hypodermis | genus | Staphylococcus | 10.20 | 0.90 | 0.021 |
| hypodermis | genus | Flavobacterium | 7.10 | 0.00 | 0.269 |
| hypodermis | genus | Veillonella | 0.00 | 4.30 | 0.500 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rinaldi, F.; Pinto, D.; Borsani, E.; Castrezzati, S.; Amedei, A.; Rezzani, R. The First Evidence of Bacterial Foci in the Hair Part and Dermal Papilla of Scalp Hair Follicles: A Pilot Comparative Study in Alopecia Areata. Int. J. Mol. Sci. 2022, 23, 11956. https://doi.org/10.3390/ijms231911956
Rinaldi F, Pinto D, Borsani E, Castrezzati S, Amedei A, Rezzani R. The First Evidence of Bacterial Foci in the Hair Part and Dermal Papilla of Scalp Hair Follicles: A Pilot Comparative Study in Alopecia Areata. International Journal of Molecular Sciences. 2022; 23(19):11956. https://doi.org/10.3390/ijms231911956
Chicago/Turabian StyleRinaldi, Fabio, Daniela Pinto, Elisa Borsani, Stefania Castrezzati, Amedeo Amedei, and Rita Rezzani. 2022. "The First Evidence of Bacterial Foci in the Hair Part and Dermal Papilla of Scalp Hair Follicles: A Pilot Comparative Study in Alopecia Areata" International Journal of Molecular Sciences 23, no. 19: 11956. https://doi.org/10.3390/ijms231911956