
Usefulness of Probiotics in the Management of NAFLD: Evidence and Involved Mechanisms of Action from Preclinical and Human Models

 , ,
, ,
Abstract
:1. Introduction
2. Effects of Probiotic Administration (Single Strain and Mixtures) on NAFLD Prevention: Evidence from Preclinical Studies
2.1. Preclinical Studies Using Single Strain Probiotics
2.2. Preclinical Studies Using Probiotic Mixtures
3. Effects of Probiotic Administration (Single Strain and Mixtures) in NAFLD Prevention: Evidence from Clinical Trials
4. Limitations of Probiotic Administration and Potential Alternatives
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Byass, P. The global burden of liver disease: A challenge for methods and for public health. BMC Med. 2014, 12, 159. [Google Scholar] [CrossRef] [Green Version]
- Engin, A. Non-Alcoholic Fatty Liver Disease. In Obesity and Lipotoxicity; Engin, A.B., Engin, A., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 443–467. [Google Scholar]
- Younossi, Z.; Anstee, Q.M.; Marietti, M.; Hardy, T.; Henry, L.; Eslam, M.; George, J.; Bugianesi, E. Global burden of NAFLD and NASH: Trends, predictions, risk factors and prevention. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Angulo, P. Nonalcoholic fatty liver disease. N. Engl. J. Med. 2002, 346, 1221–1231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunt, E.M.; Wong, V.W.S.; Nobili, V.; Day, C.P.; Sookoian, S.; Maher, J.J.; Bugianesi, E.; Sirlin, C.B.; Neuschwander-Tetri, B.A.; Rinella, M.E. Nonalcoholic fatty liver disease. Nat. Rev. Dis. Primers 2015, 1, 15080. [Google Scholar] [CrossRef] [PubMed]
- Buzzetti, E.; Pinzani, M.; Tsochatzis, E.A. The multiple-hit pathogenesis of non-alcoholic fatty liver disease (NAFLD). Metabolism 2016, 65, 1038–1048. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Zheng, J.; Zhang, S.; Wang, B.; Wu, C.; Guo, X. Advances in the Involvement of Gut Microbiota in Pathophysiology of NAFLD. Front. Med. 2020, 7, 361. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Zorita, S.; Aguirre, L.; Milton-Laskibar, I.; Fernández-Quintela, A.; Trepiana, J.; Kajarabille, N.; Mosqueda-Solís, A.; González, M.; Portillo, M.P. Relationship between changes in microbiota and liver steatosis induced by high-fat feeding—A review of rodent models. Nutrients 2019, 11, 2156. [Google Scholar] [CrossRef] [Green Version]
- Softic, S.; Cohen, D.E.; Kahn, C.R. Role of Dietary Fructose and Hepatic De Novo Lipogenesis in Fatty Liver Disease. Dig. Dis. Sci. 2016, 61, 1282–1293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lian, C.; Zhai, Z.; Li, Z.; Wang, L. High fat diet-triggered non-alcoholic fatty liver disease: A review of proposed mechanisms. Chem.-Biol. Interact. 2020, 330, 109199. [Google Scholar] [CrossRef]
- Larson-Meyer, D.E.; Newcomer, B.R.; Heilbronn, L.K.; Volaufova, J.; Smith, S.R.; Alfonso, A.J.; Lefevre, M.; Rood, J.C.; Williamson, D.A.; Ravussin, E.; et al. Effect of 6-month calorie restriction and exercise on serum and liver lipids and markers of liver function. Obesity 2008, 16, 1355–1362. [Google Scholar] [CrossRef] [Green Version]
- Elias, M.C.; Parise, E.R.; de Carvalho, L.; Szejnfeld, D.; Netto, J.P. Effect of 6-month nutritional intervention on non-alcoholic fatty liver disease. Nutrition 2010, 26, 1094–1099. [Google Scholar] [CrossRef] [PubMed]
- Divella, R.; Mazzocca, A.; Daniele, A.; Sabbà, C.; Paradiso, A. Obesity, Nonalcoholic Fatty Liver Disease and Adipocyto-kines Network in Promotion of Cancer. Int. J. Biol. Sci. 2019, 15, 610–616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandenplas, Y.; Huys, G.; Daube, G. Probiotics: An update. J. Pediatr. 2015, 91, 6–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perumpail, B.J.; Li, A.A.; John, N.; Sallam, S.; Shah, N.D.; Kwong, W.; Cholankeril, G.; Kim, D.; Ahmed, A. The Therapeutic Implications of the Gut Microbiome and Probiotics in Patients with NAFLD. Diseases 2019, 7, 27. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.; Park, K.; Ji, Y.; Park, S.; Holzapfel, W.; Hyun, C. Protective effects of Lactobacillus rhamnosus GG against dyslipidemia in high-fat diet-induced obese mice. Biochem. Biophys. Res. Commun. 2016, 473, 530–536. [Google Scholar] [CrossRef]
- Yao, F.; Jia, R.; Huang, H.; Yu, Y.; Mei, L.; Bai, L.; Ding, Y.; Zheng, P. Effect of Lactobacillus paracasei N1115 and fructooligosaccharides in nonalcoholic fatty liver disease. Arch. Med. Sci. 2019, 15, 1336–1344. [Google Scholar] [CrossRef]
- Lee, N.Y.; Yoon, S.J.; Han, D.H.; Gupta, H.; Youn, G.S.; Shin, M.J.; Ham, Y.L.; Kwak, M.J.; Kim, B.Y.; Yu, J.S.; et al. Lactobacillus and Pediococcus ameliorate progression of non-alcoholic fatty liver disease through modulation of the gut microbiome. Gut Microbes 2020, 11, 882–899. [Google Scholar] [CrossRef]
- Machado, A.S.; Oliveira, J.R.; Lelis, D.D.F.; de Paula, A.M.B.; Guimarães, A.L.S.; Andrade, J.M.O.; Brandi, I.V.; Santos, S.H.S. Oral Probiotic Bifidobacterium Longum Supplementation Improves Metabolic Parameters and Alters the Expression of the Renin-Angiotensin System in Obese Mice Liver. Biol. Res. Nurs. 2021, 23, 100–108. [Google Scholar] [CrossRef]
- Mu, J.; Tan, F.; Zhou, X.; Zhao, X. Lactobacillus fermentum CQPC06 in naturally fermented pickles prevents non-alcoholic fatty liver disease by stabilizing the gut-liver axis in mice. Food Funct. 2020, 11, 8707–8723. [Google Scholar] [CrossRef]
- Naudin, C.R.; Maner-Smith, K.; Owens, J.A.; Wynn, G.M.; Robinson, B.S.; Matthews, J.D.; Reedy, A.R.; Luo, L.; Wolfarth, A.A.; Darby, T.M.; et al. Lactococcus lactis Subspecies cremoris Elicits Protection Against Metabolic Changes Induced by a Western-Style Diet. Gastroenterology 2020, 159, 639–651.e5. [Google Scholar] [CrossRef]
- Oh, J.; Schueler, K.L.; Stapleton, D.S.; Alexander, L.M.; Yen, C.E.; Keller, M.P.; Attie, A.D.; van Pijkeren, J.P. Secretion of Recombinant Interleukin-22 by Engineered Lactobacillus reuteri Reduces Fatty Liver Disease in a Mouse Model of Diet-Induced Obesity. mSphere 2020, 5, e00183-20. [Google Scholar] [CrossRef]
- Yau, Y.F.; El-nezami, H.; Galano, J.; Kundi, Z.M.; Durand, T.; Lee, J.C. Lactobacillus rhamnosus GG and Oat Beta-Glucan Regulated Fatty Acid Profiles along the Gut-Liver-Brain Axis of Mice Fed with High Fat Diet and Demonstrated Antioxidant and Anti-Inflammatory Potentials. Mol. Nutr. Food Res. 2020, 64, 2000566. [Google Scholar] [CrossRef]
- Lee, N.Y.; Shin, M.J.; Youn, G.S.; Yoon, S.J.; Choi, Y.R.; Kim, H.S.; Gupta, H.; Han, S.H.; Kim, B.K.; Lee, D.Y.; et al. Lactobacillus attenuates progression of nonalcoholic fatty liver disease by lowering cholesterol and steatosis. Clin. Mol. Hepatol. 2021, 27, 110–124. [Google Scholar] [CrossRef]
- Xu, R.; Wan, Y.; Fang, Q.; Lu, W.; Cai, W. Supplementation with probiotics modifies gut flora and attenuates liver fat accumulation in rat nonalcoholic fatty liver disease model. J. Clin. Biochem. Nut. 2012, 50, 72–77. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Nie, S.; Zhu, K.; Ding, Q.; Li, C.; Xiong, T.; Xie, M. Lactobacillus plantarum NCU116 improves liver function, oxidative stress and lipid metabolism in rats with high fat diet induced non-alcoholic fatty liver disease. Food Funct. 2014, 5, 3216–3223. [Google Scholar] [CrossRef] [PubMed]
- Ye, H.; Li, Q.; Zhang, Z.; Sun, M.; Zhao, C.; Zhang, T. Effect of a novel potential probiotic Lactobacillus paracasei Jlus66 isolated from fermented milk on nonalcoholic fatty liver in rats. Food Funct. 2017, 8, 4539–4546. [Google Scholar] [CrossRef] [PubMed]
- Lew, L.-C.; Hor, Y.-Y.; Jaafar, M.-H.; Lau, A.-S.-Y.; Lee, B.-K.; Chuah, L.-O.; Yap, K.-P.; Azlan, A.; Azzam, G.; Choi, S.-B.; et al. Lactobacillus Strains Alleviated Hyperlipidemia and Liver Steatosis in Aging Rats via Activation of AMPK. Int. J. Mol. Sci. 2020, 21, 5872. [Google Scholar] [CrossRef] [PubMed]
- Park, E.-J.; Lee, Y.-S.; Kim, S.M.; Park, G.-S.; Lee, Y.H.; Jeong, D.Y.; Kang, J.; Lee, H.-J. Beneficial Effects of Lactobacillus plantarum Strains on Non-Alcoholic Fatty Liver Disease in High Fat/High Fructose Diet-Fed Rats. Nutrients 2020, 12, 542. [Google Scholar] [CrossRef] [Green Version]
- Yan, Y.; Liu, C.; Zhao, S.; Wang, X.; Wang, J.; Zhang, H.; Wang, Y.; Zhao, G. Probiotic Bifidobacterium lactis V9 attenuates hepatic steatosis and inflammation in rats with non-alcoholic fatty liver disease. AMB Express 2020, 10, 101. [Google Scholar] [CrossRef]
- Luo, M.; Yan, J.; Wu, L.; Wu, J.; Chen, Z.; Jiang, J.; Chen, Z.; He, B. Probiotics Alleviated Nonalcoholic Fatty Liver Disease in High-Fat Diet-Fed Rats via Gut Microbiota/FXR/FGF15 Signaling Pathway. J. Immunol. Res. 2021, 2021, 2264737. [Google Scholar] [CrossRef]
- Ritze, Y.; Bárdos, G.; Claus, A.; Ehrmann, V.; Bergheim, I.; Schwiertz, A.; Bischoff, S.C. Lactobacillus rhamnosus GG Protects against Non-Alcoholic Fatty Liver Disease in Mice. PLoS ONE 2014, 9, e80169. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Liu, L.; Liu, Q.; Li, F.; Zhang, L.; Zhu, F.; Shao, T.; Barve, S.; Chen, Y.; Li, X.; et al. Fibroblast growth factor 21 is required for the therapeutic effects of Lactobacillus rhamnosus GG against fructose-induced fatty liver in mice. Mol. Metab. 2019, 29, 145–157. [Google Scholar] [CrossRef]
- Zhao, Z.; Wang, C.; Zhang, L.; Zhao, Y.; Duan, C.; Zhang, X.; Gao, L.; Li, S. Lactobacillus plantarum NA136 improves the non-alcoholic fatty liver disease by modulating the AMPK/Nrf2 pathway. Appl. Microbiol. Biotechnol. 2019, 103, 5843–5850. [Google Scholar] [CrossRef]
- Azarang, A.; Farshad, O.; Ommati, M.M.; Jamshidzadeh, A.; Heidari, R.; Abootalebi, S.N.; Gholami, A. Protective Role of Probiotic Supplements in Hepatic Steatosis: A Rat Model Study. Biomed. Res. Int. 2020, 2020, 5487659. [Google Scholar] [CrossRef]
- Endo, H.; Niioka, M.; Kobayashi, N.; Tanaka, M.; Watanabe, T. Butyrate-Producing Probiotics Reduce Nonalcoholic Fatty Liver Disease Progression in Rats: New Insight into the Probiotics for the Gut-Liver Axis. PLoS ONE 2013, 8, e63388. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Chen, L.; Zhao, Y.; Wang, C.; Duan, C.; Yang, G.; Niu, C.; Li, S. Lactobacillus plantarum NA136 ameliorates nonalcoholic fatty liver disease by modulating gut microbiota, improving intestinal barrier integrity, and attenuating inflammation. Appl. Microbiol. Biotechnol. 2020, 104, 5273–5282. [Google Scholar] [CrossRef] [PubMed]
- Rakhshandehroo, M.; Knoch, B.; Müller, M.; Kersten, S. Peroxisome Proliferator-Activated Receptor Alpha Target Genes. PPAR Res. 2010, 2010, 612089. [Google Scholar] [CrossRef] [Green Version]
- Cantó, C.; Auwerx, J. PGC-1alpha, SIRT1 and AMPK, an energy sensing network that controls energy expenditure. Curr. Opin. Lipidol. 2009, 20, 98–105. [Google Scholar] [CrossRef] [Green Version]
- Arulselvan, P.; Fard, M.T.; Tan, W.S.; Gothai, S.; Fakurazi, S.; Norhaizan, M.E.; Kumar, S.S. Role of Antioxidants and Natural Products in Inflammation. Oxid. Med. Cell. Longev. 2016, 2016, 5276130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, C.J.; Sellmann, C.; Engstler, A.J.; Ziegenhardt, D.; Bergheim, I. Supplementation of sodium butyrate protects mice from the development of non-alcoholic steatohepatitis (NASH). Br. J. Nutr. 2015, 114, 1745–1755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chambers, E.S.; Preston, T.; Frost, G.; Morrison, D.J. Role of Gut Microbiota-Generated Short-Chain Fatty Acids in Metabolic and Cardiovascular Health. Curr. Nutr. Rep. 2018, 7, 198–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dangana, E.O.; Omolekulo, T.E.; Areola, E.D.; Olaniyi, K.S.; Soladoye, A.O.; Olatunji, L.A. Sodium acetate protects against nicotine-induced excess hepatic lipid in male rats by suppressing xanthine oxidase activity. Chem.-Biol. Interact. 2020, 316, 108929. [Google Scholar] [CrossRef]
- Kim, B.; Kwon, J.; Kim, M.; Park, H.; Ji, Y.; Holzapfel, W.; Hyun, C. Protective effects of Bacillus probiotics against high-fat diet-induced metabolic disorders in mice. PLoS ONE 2018, 13, e0210120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.; Kim, J.; Kim, J.; Jang, S.; Han, M.J.; Kim, D.H. Lactobacillus plantarum LC27 and Bifidobacterium longum LC67 simultaneously alleviate high-fat diet-induced colitis, endotoxemia, liver steatosis, and obesity in mice. Nutr. Res. 2019, 67, 78–89. [Google Scholar] [CrossRef]
- Ahmadi, S.; Wang, S.; Nagpal, R.; Wang, B.; Jain, S.; Razazan, A.; Mishra, S.P.; Zhu, X.; Wang, Z.; Kavanagh, K.; et al. A human-origin probiotic cocktail ameliorates aging-related leaky gut and inflammation via modulating the microbiota/taurine/tight junction axis. JCI Insight 2020, 5, e132055. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Liu, F.; Lu, J.; Shi, J.; Guan, J.; Yan, F.; Li, B.; Huo, G. Probiotic Mixture of Lactobacillus plantarum Strains Improves Lipid Metabolism and Gut Microbiota Structure in High Fat Diet-Fed Mice. Front. Microbiol. 2020, 11, 512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, J.; Xiong, J.; Ni, J.; Chen, C.; Wang, K. Live Combined B. subtilis and E. faecium Alleviate Liver Inflammation, Improve Intestinal Barrier Function, and Modulate Gut Microbiota in Mice with Non-Alcoholic Fatty Liver Disease. Med. Sci. Monit. 2021, 27, e931143. [Google Scholar] [CrossRef] [PubMed]
- Plaza-Diaz, J.; Gomez-Llorente, C.; Abadia-Molina, F.; Saez-Lara, M.J.; Campaña-Martin, L.; Muñoz-Quezada, S.; Romero, F.; Gil, A.; Fontana, L. Effects of Lactobacillus paracasei CNCM I-4034, Bifidobacterium breve CNCM I-4035 and Lactobacillus rhamnosus CNCM I-4036 on Hepatic Steatosis in Zucker Rats. PLoS ONE 2014, 9, e98401. [Google Scholar] [CrossRef]
- Al-Muzafar, H.M.; Amin, K.A. Probiotic mixture improves fatty liver disease by virtue of its action on lipid profiles, leptin, and inflammatory biomarkers. BMC Complement. Altern. Med. 2017, 17, 43. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.-J.; Park, S.H.; Sin, H.-S.; Jang, S.-H.; Lee, S.-W.; Kim, S.-Y.; Kwon, B.; Yu, K.-Y.; Kim, S.Y.; Yang, D.K. Hypocholesterolemic effects of probiotic mixture on diet-induced hypercholesterolemic rats. Nutrients 2017, 9, 293. [Google Scholar] [CrossRef] [Green Version]
- Candelli, M.; Franza, L.; Pignataro, G.; Ojetti, V.; Covino, M.; Piccioni, A.; Gasbarrini, A.; Franceschi, F. Interaction between Lipopolysaccharide and Gut Microbiota in Inflammatory Bowel Diseases. Int. J. Mol. Sci. 2021, 22, 6242. [Google Scholar] [CrossRef] [PubMed]
- Aller, R.; De Luis, D.A.; Izaola, O.; Conde, R.; Gonzalez Sagrado, M.; Primo, D.; De La Fuente, B.; Gonzalez, J. Effect of a probiotic on liver aminotransferases in nonalcoholic fatty liver disease patients: A double blind randomized clinical trial. Eur. Rev. Med. Pharmacol. Sci. 2011, 15, 1090–1095. [Google Scholar] [PubMed]
- Famouri, F.; Shariat, Z.; Hashemipour, M.; Keikha, M.; Kelishadi, R. Effects of Probiotics on Nonalcoholic Fatty Liver Disease in Obese Children and Adolescents. J. Pediatr. Gastroenterol. Nutr. 2017, 64, 413–417. [Google Scholar] [CrossRef] [PubMed]
- Kobyliak, N.; Abenavoli, L.; Mykhalchyshyn, G.; Kononenko, L.; Boccuto, L.; Kyriienko, D.; Dynnyk, O. A Multi-strain Probiotic Reduces the Fatty Liver Index, Cytokines and Aminotransferase levels in NAFLD Patients: Evidence from a Randomized Clinical Trial. J. Gastrointest. Liver Dis. 2018, 27, 41–49. [Google Scholar] [CrossRef] [Green Version]
- Ahn, S.B.; Jun, D.W.; Kang, B.; Lim, J.H.; Lim, S.; Chung, M.J. Randomized, Double-blind, Placebo-controlled Study of a Multispecies Probiotic Mixture in Nonalcoholic Fatty Liver Disease. Sci. Rep. 2019, 9, 5688. [Google Scholar] [CrossRef] [Green Version]
- Duseja, A.; Acharya, S.K.; Mehta, M.; Chhabra, S.; Rana, S.; Das, A.; Dattagupta, S.; Dhiman, R.K.; Chawla, Y.K. High potency multistrain probiotic improves liver histology in non-alcoholic fatty liver disease (NAFLD): A randomised, double-blind, proof of concept study. BMJ Open Gastroenterol. 2019, 6, e000315. [Google Scholar] [CrossRef] [Green Version]
- Behrouz, V.; Aryaeian, N.; Zahedi, M.J.; Jazayeri, S. Effects of probiotic and prebiotic supplementation on metabolic parameters, liver aminotransferases, and systemic inflammation in nonalcoholic fatty liver disease: A randomized clinical trial. J. Food Sci. 2020, 85, 3611–3617. [Google Scholar] [CrossRef]
- Nabavi, S.; Rafraf, M.; Somi, M.H.; Homayouni-Rad, A.; Asghari-Jafarabadi, M. Effects of probiotic yogurt consumption on metabolic factors in individuals with nonalcoholic fatty liver disease. J. Dairy Sci. 2014, 97, 7386–7393. [Google Scholar] [CrossRef]
- Chong, P.L.; Laight, D.; Aspinall, R.J.; Higginson, A.; Cummings, M.H. A randomised placebo controlled trial of VSL#3® probiotic on biomarkers of cardiovascular risk and liver injury in non-alcoholic fatty liver disease. BMC Gastroenterol. 2021, 21, 144. [Google Scholar]
- Mohamad Nor, M.H.; Ayob, N.; Mokhtar, N.M.; Raja Ali, R.A.; Tan, G.C.; Wong, Z.; Shafiee, N.H.; Wong, Y.P.; Mustangin, M.; Nawawi, K.N.M. The Effect of Probiotics (MCP® BCMC® Strains) on Hepatic Steatosis, Small Intestinal Mucosal Immune Function, and Intestinal Barrier in Patients with Non-Alcoholic Fatty Liver Disease. Nutrients 2021, 13, 3192. [Google Scholar] [CrossRef]
- Xu, C.; Zhu, H.; Qiu, P. Aging progression of human gut microbiota. BMC Microbiol. 2019, 19, 236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, A.; Zhang, Y.N.; Boehringer, A.S.; Montes, V.; Andre, M.P.; Erdman, J.W., Jr.; Loomba, R.; Valasek, M.A.; Sirlin, C.B.; O’Brien, W.D., Jr. Assessment of Hepatic Steatosis in Nonalcoholic Fatty Liver Disease by Using Quantitative US. Radiology 2020, 295, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Yadav, H.; Lee, J.; Lloyd, J.; Walter, P.; Rane, S.G. Beneficial Metabolic Effects of a Probiotic via Butyrate-induced GLP-1 Hormone Secretion. J. Biol. Chem. 2013, 288, 25088–25097. [Google Scholar] [CrossRef] [Green Version]
- Everard, A.; Belzer, C.; Geurts, L.; Ouwerkerk, J.P.; Druart, C.; Bindels, L.B.; Guiot, Y.; Derrien, M.; Muccioli, G.G.; Delzenne, N.M.; et al. Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proc. Natl. Acad. Sci. USA 2013, 110, 9066–9071. [Google Scholar] [CrossRef] [Green Version]
- Minami, J.; Kondo, S.; Yanagisawa, N.; Odamaki, T.; Xiao, J.; Abe, F.; Nakajima, S.; Hamamoto, Y.; Saitoh, S.; Shimoda, T. Oral administration of Bifidobacterium breve B-3 modifies metabolic functions in adults with obese tendencies in a randomised controlled trial. J. Nutr. Sci. 2015, 4, e17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Firouzi, S.; Majid, H.A.; Ismail, A.; Kamaruddin, N.A.; Barakatun-Nisak, M.Y. Effect of multi-strain probiotics (multi-strain microbial cell preparation) on glycemic control and other diabetes-related outcomes in people with type 2 diabetes: A randomized controlled trial. Eur. J. Nutr. 2017, 56, 1535–1550. [Google Scholar] [CrossRef]
- Asad, F.; Anwar, H.; Yassine, H.M.; Ullah, M.I.; Rahman, A.; Kamran, Z.; Sohail, M.U. White Button Mushroom, Agaricus bisporus (Agaricomycetes), and a Probiotics Mixture Supplementation Correct Dyslipidemia without Influencing the Colon Microbiome Profile in Hypercholesterolemic Rats. Int. J. Med. Mushrooms 2020, 22, 235–244. [Google Scholar] [CrossRef]
- Yao, M.; Qv, L.; Lu, Y.; Wang, B.; Berglund, B.; Li, L. An Update on the Efficacy and Functionality of Probiotics for the Treatment of Non-Alcoholic Fatty Liver Disease. Engineering 2021, 7, 679–686. [Google Scholar] [CrossRef]
- Etxeberria, U.; Hijona, E.; Aguirre, L.; Milagro, F.I.; Bujanda, L.; Rimando, A.M.; Martínez, J.A.; Portillo, M.P. Pterostilbene-induced changes in gut microbiota composition in relation to obesity. Mol. Nutr. Food Res. 2017, 61, 1500906. [Google Scholar] [CrossRef]
- Moreira, G.V.; Azevedo, F.F.; Ribeiro, L.M.; Santos, A.; Guadagnini, D.; Gama, P.; Liberti, E.A.; Saad, M.J.A.; Carvalho, C.R.O. Liraglutide modulates gut microbiota and reduces NAFLD in obese mice. J. Nutr. Biochem. 2018, 62, 143–154. [Google Scholar] [CrossRef]
- Cani, P.D.; de Vos, W.M. Next-Generation Beneficial Microbes: The Case of Akkermansia muciniphila. Front. Microbiol. 2017, 8, 1765. [Google Scholar] [CrossRef] [PubMed]
- Seo, B.; Jeon, K.; Moon, S.; Lee, K.; Kim, W.K.; Jeong, H.; Cha, K.H.; Lim, M.Y.; Kang, W.; Kweon, M.N.; et al. Roseburia spp. Abundance Associates with Alcohol Consumption in Humans and Its Administration Ameliorates Alcoholic Fatty Liver in Mice. Cell Host Microbe 2020, 27, 25–40.e6. [Google Scholar] [CrossRef] [PubMed]
- Doron, S.; Snydman, D.R. Risk and Safety of Probiotics. Clin. Infect. Dis. 2015, 60, S129–S134. [Google Scholar] [CrossRef] [Green Version]
- Cuevas-González, P.F.; Liceaga, A.M.; Aguilar-Toalá, J.E. Postbiotics and paraprobiotics: From concepts to applications. Food Res. Int. 2020, 136, 109502. [Google Scholar] [CrossRef]
- Nataraj, B.H.; Ali, S.A.; Behare, P.V.; Yadav, H. Postbiotics-parabiotics: The new horizons in microbial biotherapy and functional foods. Microb. Cell Fact. 2020, 19, 168. [Google Scholar] [CrossRef] [PubMed]
- Vallejo-Cordoba, B.; Castro-López, C.; García, H.S.; González-Córdova, A.F.; Hernández-Mendoza, A. Postbiotics and paraprobiotics: A review of current evidence and emerging trends. Adv. Food Nutr. Res. 2020, 94, 1–34. [Google Scholar]
- Piqué, N.; Berlanga, M.; Miñana-Galbis, D. Health Benefits of Heat-Killed (Tyndallized) Probiotics: An Overview. Int. J. Mol. Sci. 2019, 20, 2534. [Google Scholar] [CrossRef] [Green Version]
- Kang, X.; Liang, H.; Luo, Y.; Li, Z.; He, F.; Han, X.; Zhang, L. Anti-adipogenesis and metabolism-regulating effects of heat-inactivated Streptococcus thermophilus MN-ZLW-002. Lett. Appl. Microbiol. 2020, 72, 677–687. [Google Scholar] [CrossRef]
- Nakamura, F.; Ishida, Y.; Aihara, K.; Sawada, D.; Ashida, N.; Sugawara, T.; Aoki, Y.; Takehara, I.; Takano, K.; Fujiwara, S. Effect of fragmentedLactobacillus amylovorusCP1563 on lipid metabolism in overweight and mildly obese individuals: A randomized controlled trial. Microb. Ecol. Health Dis. 2016, 27, 30312. [Google Scholar] [CrossRef] [Green Version]
- Zorzela, L.; Ardestani, S.K.; Mcfarland, L.V.; Vohra, S. Is there a role for modified probiotics as beneficial microbes: A systematic review of the literature. Benef. Microbes 2017, 8, 739–754. [Google Scholar] [CrossRef]
- Peluzio, M.D.C.G.; Martinez, J.A.; Milagro, F.I. Postbiotics: Metabolites and mechanisms involved in microbiota-host interactions. Trends Food Sci. Technol. 2021, 108, 11–26. [Google Scholar] [CrossRef]
- Wang, S.; Ahmadi, S.; Nagpal, R.; Jain, S.; Mishra, S.P.; Kavanagh, K.; Zhu, X.; Wang, Z.; McClain, D.A.; Kritchevsky, S.B.; et al. Lipoteichoic acid from the cell wall of a heat killed Lactobacillus paracasei D3-5 ameliorates aging-related leaky gut, inflammation and improves physical and cognitive functions: From C. elegans to mice. GeroScience 2020, 42, 333–352. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Ni, Y.; Wang, Z.; Tu, W.; Ni, L.; Zhuge, F.; Zheng, A.; Hu, L.; Zhao, Y.; Zheng, L.; et al. Spermidine improves gut barrier integrity and gut microbiota function in diet-induced obese mice. Gut Microbes 2020, 12, 1832857. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Ni, Y.; Hu, L.; Zhao, Y.; Zheng, L.; Yang, S.; Ni, L.; Fu, Z. Spermidine ameliorates high-fat diet-induced hepatic steatosis and adipose tissue inflammation in preexisting obese mice. Life Sci. 2021, 265, 118739. [Google Scholar] [CrossRef] [PubMed]



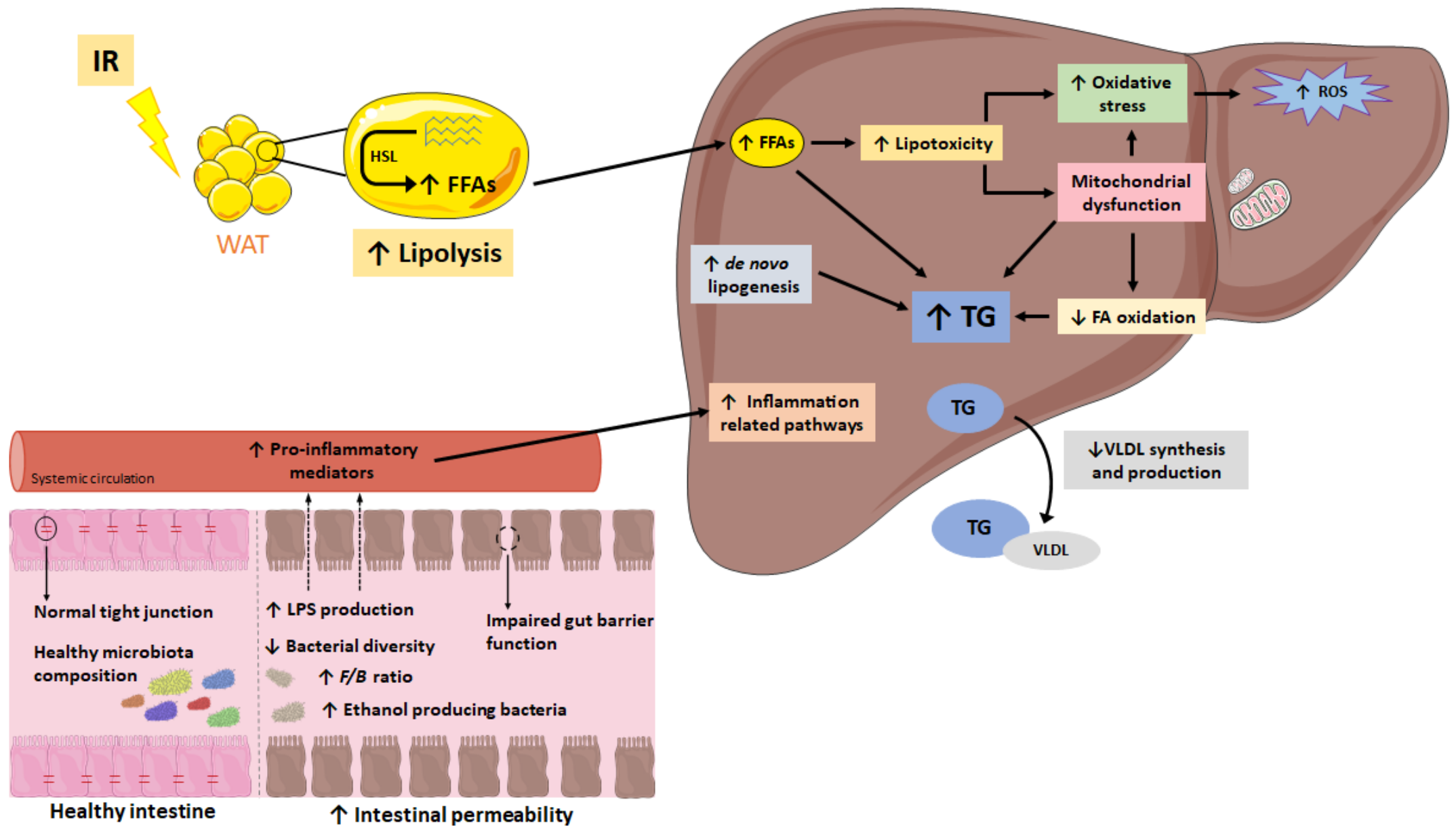{kind=link}
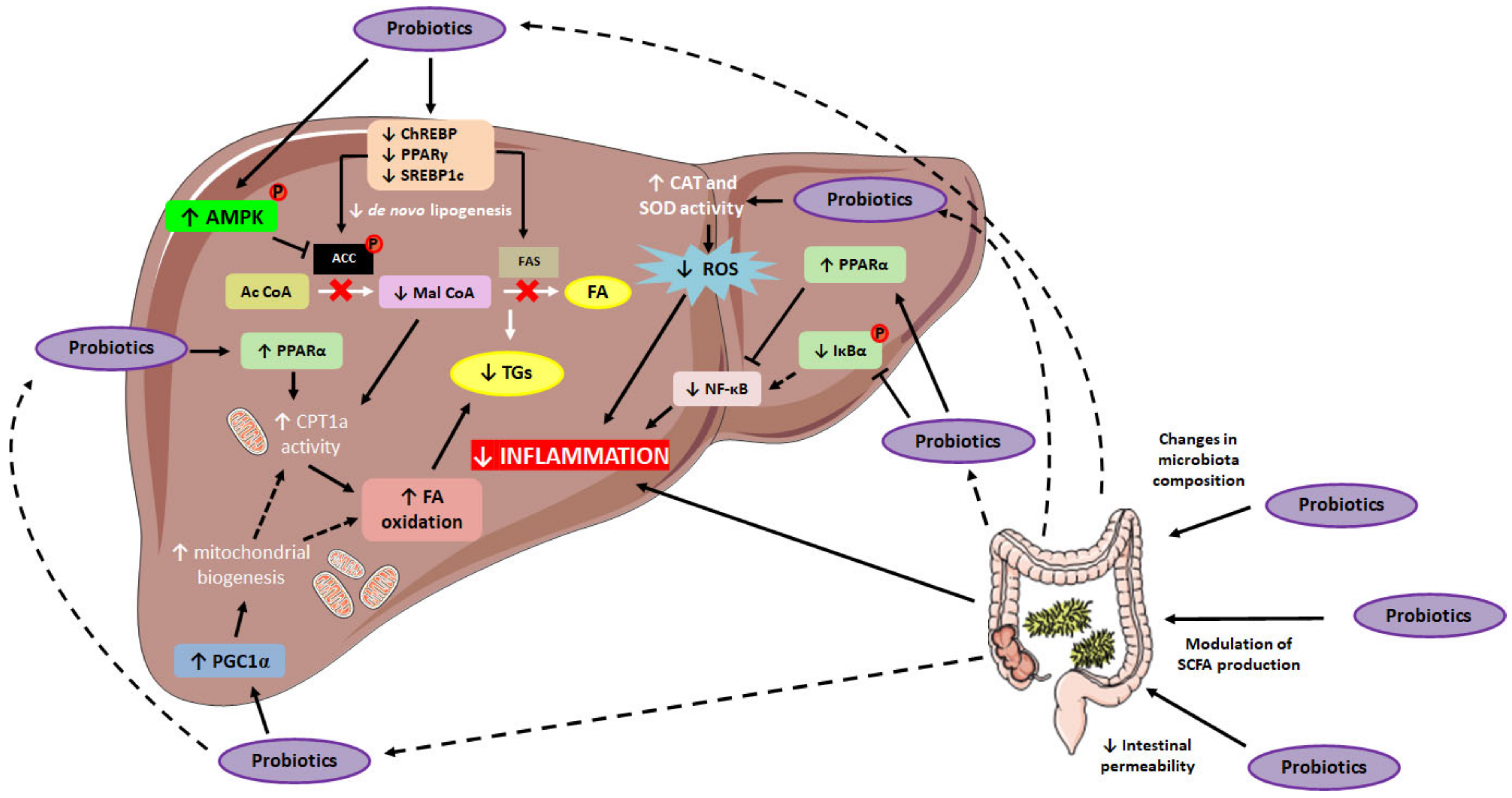{kind=link}
| Reference | Animal Model | Experimental Conditions | Probiotic Treatment | Effects on Liver | Mechanisms of Action |
|---|---|---|---|---|---|
| [32] | Female C57BL/J6 mice 6-week-old | STD diet with 30% fructose in drinking water. | L. rhamnosus GG—LGG Daily administration Dose: 5.2 × 107 CFU/bw g/d. Diluted in drinking water Treatment length: 8 w. | ↓ Liver fat accumulation ↓ Liver TG content ↓ Serum ALT levels ↓ Liver inflammation | Down-regulation of lipogenic markers in the liver: ↓ Gene expression of Acc, Fas and Chrebp. Down-regulation of pro-inflammatory markers and mediators in the liver: ↓ Gene expression of Il-1β, Il-8R, Tnfα and Il-12. Decreased portal levels of LPS. Up-regulation of markers of intestinal mucosa integrity: ↑ Protein expression of Occludin-1 and Claudin-1. |
| [16] | Male C57BL/6 mice 4-week-old | HFD (60% energy from fat). | L. rhamnosus GG—LGG Oral daily administration Dose: 1 × 108 CFU/day Treatment length: 13 w. | ↓ Liver weight ↓ Liver fat accumulation ↓ Liver inflammation | Down-regulation of lipogenic markers in the liver: ↓ Gene expression of Srebp-1 and Ppar-γ. Down-regulation of genes related to long-chain fatty acid uptake and lipoprotein synthesis: ↓ Gene expression of Cd36 and ApoB100. Down-regulation of pro-inflammatory markers and mediators in the liver: ↓ Gene expression of Il-6, Il-12, F4/80 and Cd11b. Modulation of gut microbiota composition: ↑ Proportion of Bacteroidetes. |
| [17] | Male C57BL/6 mice 4-week-old | HFD. | L. paracasei N1115 Oral daily administration Dose: 2.2 × 109 CFU/mL diluted in normal saline (0.5 mL/day). Treatment length: 16 w. | ↓ Liver fat accumulation ↓ Liver inflammation ↓ Liver fibrosis | Decreased content of hepatic inflammatory mediators (Tnfα and IL-1β). Down-regulation of pro-inflammatory markers and mediators in the liver: ↓ Gene expression of Nf-κB, Tlr-4 and Lps. Decreased serum levels of liver fibrosis markers (MAO). Up-regulation of markers of intestinal mucosa integrity: ↑ Protein expression of Occludin-1 and Claudin-1. |
| [33] | Female C57BL/6N mice 6–8-week-old | STD diet with 30% fructose in drinking water. | L. rhamnosus Oral daily gavage Dose:1 × 109 CFU/day. Treatment length: 5 or 12 w. | ↓ Liver fat accumulation ↓ Liver TG, TC and VLDL content ↓ Liver inflammation ↓ Liver apoptotic cells | Down-regulation of liver injury protection markers: ↑ Gene expression of Fgf21. Down-regulation of pro-inflammatory markers and mediators in the liver: ↓ Gene expression of Tnfα and Cxcl10. Down-regulation of lipogenic markers in the liver: ↓ Gene expression of Fas, Srebp1c and Scd1. ↓ Protein expression of SREBP1c and ChREBP. Up-regulation of fatty acid oxidation markers in the liver: ↑ Gene expression of CPT1 and PPARα. Down-regulation of markers involved in hepatic ceramide content: ↓ Methylation of PP2AC. |
| [34] | Male C57BL/6N mice 8-week-old | HFD/F (65% energy from fat and 30% fructose solution). | L. plantarum NA136 group Oral daily administration Dose: 1 × 109 CFU/day. Treatment length: 16 w. | ↓ Liver fat accumulation ↓ Liver FFA content ↓ Liver inflammation ↓ Liver ALT and AST levels | Down-regulation of lipogenic markers in the liver: ↓ Protein expression of FAS and SREBP-1. ↑ Phosphorylation of ACC. Up-regulation of energy yielding pathways in the liver: ↑ Phosphorylation of AMPK. Down-regulation of oxidative stress in the liver: ↓ Content of MDA. ↑ Protein expression of HO-1 andNrf2. ↑ Content of CAT. ↑ Activity of SOD. |
| [18] | Male SPF C57BL/6J mice 6-week-old | Normal or Western diet (42% energy from fat). | L. bulgaricus L. casei L. helveticus P. pentosaceus KID7 Daily administration Dose: 1 × 109 CFU/g suspended in distilled water. Treatment length: 8 w. | ↓ Liver steatosis grade (all treated groups) ↓ Liver inflammation (all treated groups except animals receiving L. casei) ↓ Liver/bw ratio (groups treated with L. bulgaricus, L. helveticus and P. pentosaceus) ↓ Liver AST levels (groups treated with L. bulgaricus and L. helveticus) ↓ Liver ALT levels (group treated with L. bulgaricus) ↓ NAS (groups treated with L. bulgaricus, L. helveticus and P. pentosaceus) | Down-regulation of macrophage markers in the liver: ↓ Expression of Cd68 (groups treated with L. bulgaricus, L. helveticus, L. casei and P. pentosaceus). Modulation of gut microbiota composition: ↓ F/B ratio (groups treated with L. bulgaricus, L. helveticus, P. pentosaceus and L. casei). ↑ Content of A. muciniphila (groups treated with L. bulgaricus, L. helveticus and L. casei). Down-regulation of pro-inflammatory markers and mediators in the liver: ↓ Gene expression of Tnfα Il-6 and Il-1β (in all the treated groups). |
| [19] | Male Swiss mice 4-week-old | HFD (61% energy from fat). | B. longum Daily oral gavage Dose: 5 × 109 CFU/kg bw/d Treatment length: 4 w. | ↓ Liver lipid droplet size | Up-regulation of RAS related genes in the liver: ↑ Gene expression of Ace2 and Masr. |
| [20] | Male C57BL/6N mice 6-week-old | HFD. Animals also received a 10% fructose solution. | L. fermentum—CQPC06 L. delbrueckii subsp. Bulgaricus—LDSB Daily oral gavage Dose of 1 × 109 CFU/kg bw/d (L. fermentum—CQPC06) or 1 × 1010 CFU/kg bw/d (L. fermentum—CQPC06 and L. delbrueckii subsp. Bulgaricus—LDSB) suspended in sterile saline. Treatment length: 8 w. | ↓ Liver weight and index ↓ Liver TG ↓ Serum and liver AST and ALT levels ↓ Serum and liver AKP levels | Decreased ROS levels in the livers of animals receiving L. fermentum—CQPC06 (at both doses) and L. delbrueckii subsp. Bulgaricus—LDSB. Up-regulation of fatty acid oxidation markers in the liver (L. fermentum—CQPC06 (at both doses)): ↑ Gene expression of Cpt1 and Ppar-α. ↑ Protein expression of CPT1 and PPAR-α. Down-regulation of lipogenic markers in the liver (L. fermentum—CQPC06 (at both doses)): ↓ Gene expression of C/ebp-α and Ppar-γ. ↓ Protein expression of C/EBP-α and PPAR-γ. Up-regulation of markers of intestinal mucosa integrity (L. fermentum—CQPC06 (at both doses)): ↑ Protein expression of ZO-1, Occludin and Claudin-1. Modulation of gut microbiota composition (L. fermentum—CQPC06 (at both doses) and L. delbrueckii subsp. Bulgaricus—LDSB): ↓ F/B ratio. ↑ Content of Akkermansia. |
| [21] | Female C57BL/6 mice | WSD (40% energy from fat). | L. rhamnosus GG ATCC 53103 L. lactis subsp. cremoris ATCC 19257 Oral gavage Thrice weekly Dose: 1 × 109 CFU Treatment length: 16 w. | ↓ Liver weight (group treated with L. cremoris) ↓ Liver PC, PS, TG and TG content (group treated with L. cremoris) ↓ Liver lipid droplet area (group treated with L. cremoris) ↓ Liver inflammation (group treated with L. cremoris) | Down-regulation of hepatic content of lipids related to pro-inflammatory response: ↓ Levels of ARA containing lipids (group treated with L. cremoris). Down-regulation of inflammation associated metabolites in the liver: ↓ Levels of Resolvin E1, 9-HETE and 9HpODE (group treated with L. cremoris). |
| [22] | Male C57BL/6J mice 6-week-old | HFD (45% energy from fat). | L. reuteri 6475 L. reuteri VPL3461 Daily oral gavage (in a volume of 100 μL) Dose of 1 × 109 CFU/mL Treatment length: 8 w. | ↓ Liver TG content (all groups) | Not specified. |
| [23] | Male C57BL/6N mice 3–4-weeks-old | HFD (42% energy from fat). | L. rhamnosus GG Daily administration Dose: 1 × 108 CFU/day mixed in the experimental diet. Treatment length: 17 w. | ↓ Liver weight ↓ Liver TG content | Modulation of SCFA levels in the cecum: ↑ Acetate levels. Modulation of SCFA in the liver: ↑ Acetate levels. Modulation of anti-inflammatory lipid mediator levels: ↓ ώ6/ώ 3 PUFA ratio. |
| [37] | Male C57BL/6N mice 8-week-old | HFD/F (65% energy from fat and 30% dietary volume provided as fructose solution). | L. plantarum NA136 Daily oral administration daily Dose: 1 × 109 CFU/day. Treatment length: 16 w. | ↓ Liver lipid content | Modulation of gut microbiota composition: ↑ Bacterial richness and diversity. Up-regulation of intestinal mucosa integrity markers: ↑ Protein expression of tight-junction markers (ZO-1, Occludin, Claudin-1). ↓ Protein expression mucosal dysfunction markers (HIF-1α). Decreased serum levels of pro-inflammatory cytokines (TNF-α, IL-6, and IL-1β) and LPS. Down-regulation of pro-inflammatory markers and mediators in the liver: ↓ Protein expression of NF-κB. ↓ Phosphorylation of p38. |
| [24] | Male SPF C57BL/6J mice 6-week-old | WSD (42% energy from fat). | L. acidophilus L. fermentum L. paracasei L. plantarum Daily administration Dose: 1 × 109 CFU suspended in drinking water. Treatment length: 8 w. | ↓ Liver steatosis score (groups treated with L. paracasei, L. plantarum and L. acidophilus) ↓ Liver TG content (groups treated with L. acidophilus, L. fermentum and L. paracasei) | Modulation of microbiota composition: ↑ Bacteroidetes content (group treated with L. paracasei). ↓ Firmicutes content (group treated with L. paracasei). |
| [25] | Male Sprague-Dawley rats | HFD | L. acidophilus CGMCC 2106. B. longum CGMCC 2107. Daily administration Dose: 1 × 1010 CFU/mL suspended in drinking water. Treatment length: 12 w. | ↓ Liver fat accumulation (group treated with B. longum) | Modulation of fecal microbiota composition: ↑ Bifidobacterium content (group treated with B. longum). ↑ Lactobacillus content (group treated with L. acidophilus). |
| [36] | Male Fischer 344 rats | CDAA diet (30% energy from fat). Animals were fed ad libitum and had free access to drinking water during the whole experiment. | C. butyricum Daily administration Dose: 8.5 × 109 CFU/g mixed in the diet. Treatment length: 42 w. | ↓ Liver total lipid and TG content ↓ Liver inflammation ↓ NAFLD progression (fibrosis) ↓ Serum ALT levels ↓ Liver lipid peroxidation ↓ Oxidative stress | Up-regulation of energy yielding pathways in the liver: ↑ Phosphorylation of AMPK. Up-regulation of fatty acid oxidation markers in the liver: ↑ Protein expression of PPARα. Down-regulation of lipogenic markers in the liver: ↓ Protein expression of SREBP-1c and PPAR-γ. Down-regulation of pro-inflammatory markers and mediators in the liver: ↓ Protein expression of NF-kB and TNF-α. Down-regulation of lipid peroxidation markers in the liver: ↓ Content of 4-HNE and MDA. Up-regulation of antioxidant markers in the liver: ↑ Protein expression of Nrf2 and HO-1. Up-regulation of markers of intestinal mucosa integrity: ↑ Protein expression of ZO1 and Ocln. |
| [26] | Male Sprague-Dawley rats | HFD. | L. plantarum NCU116-L L. plantarum NCU116-H Daily administration Dose: 1 × 108 CFU/mL (L. plantarum NCU116-L) or 1 × 109 CFU/mL (L. plantarum NCU116-H) suspended in a sterile saline solution. Treatment length: 5 w. | ↓ Liver AST levels (group treated with L. plantarum NCU116-H) ↓ Liver oxidative stress ↓ Liver TC and TG content ↓ Liver inflammation | Down-regulation of oxidative stress markers in liver: ↓ MDA content (group treated with L. plantarum NCU116-H). Up-regulation of antioxidant markers in the liver: ↑ Activity of SOD and GPx (all groups). ↑ Activity of CAT (group treated with L. plantarum NCU116-H). ↑ T-AOC (all groups). Down-regulation of serum pro-inflammatory cytokines: ↓ Levels of LPS and IL-6 (all groups). ↓ Levels of TNFα (L. plantarum NCU116-H). Up-regulation of fatty acid oxidation and lipolysis markers in the liver: ↑ Gene expression of Pparα, Pparγ, Pparδ, Pgc1α and Cpt1α (all groups). Down-regulation of lipogenic markers in the liver: ↓ Gene expression of Fas, Acc and Scd1 (all groups). Modulation of colonic microbiota composition: ↓ Gene expression of Bacteroides (all groups). ↑ Gene expression of Lactobacillus spp. and Bifidobacterium spp. (all groups). |
| [24] | Male Wistar rats | HFD (60% energy from fat). | L. paracasei Jlus66 Daily oral administration Doses: 1, 2 or 4 × 1010 CFU/d. Treatment length: 20 w. | ↓ Liver weight ↓ Liver fat accumulation ↓ Liver inflammation ↓ Serum ALT levels (high dose) | Not specified. |
| [35] | Male Sprague-Dawley Rats 42-day-old | STD plus 20% fructose in drinking water. | L. acidophilus B. coagulans L. casei L. reuteri Daily administration Dose: 1 × 109 CFU/mL suspended in drinking water. Treatment length: 16 w. | ↓ Liver TG content (groups treated with L. acidophilus and L. reuteri) ↓ Serum ALT levels (all groups) ↓ Liver oxidative stress (all groups) | Up-regulation of antioxidant response in the liver: ↑ Content of glutathione (groups treated with L. acidophilus and L. casei). ↓ Liver ROS formation (groups treated with L. acidophilus, L. casei and B. coagulans). ↓ Liver protein-carbonylation (all groups). ↓ Liver lipid peroxidation (all groups). |
| [28] | Sprague-Dawley rats 8-week-old | HFD 54% energy from fat). Animals were injected with 600 mg/kg/day of D-galactose daily. | L. fermentum DR9 L. plantarum DR7 L. reuteri 8513d Daily administration Dose: 1 × 1010 CFU/day dissolved in 100 μL of saline and mixed into 1 g of experimental diet. Treatment length: 12 w. | ↓ Liver lipid content (groups treated with L. fermentum DR9, L. plantarum DR7 and L. reuteri 8513d) ↓ Liver inflammation (groups treated with L. fermentum DR9, L. plantarum DR7 and L. reuteri 8513d) ↓ Liver ALP content (groups treated with L. fermentum DR9 and L. plantarum DR7) | Down-regulation of lipogenic markers in the liver: ↓ Gene expression of Scd1 gene expression (groups treated with L. fermentum DR9 and L. plantarum DR7). Decreased liver content of pro-inflammatory cytokines: ↓ IL-4 levels (groups treated with L. fermentum DR9 and L. plantarum DR7). Up-regulation of energy yielding pathways in the liver: ↑ Gene expression of Ampkα1 (groups treated with L. fermentum DR9 and L. plantarum DR7) and Ampkα2 gene expression (group treated with L. plantarum DR7). |
| [16] | Male Wistar Rats 6-week-old | HFD (45% of energy from fat). Animals also received 10% fructose in drinking water. | L. Plantarum strain ATG-K2 L. Plantarum strain ATG-K6 Daily oral gavage Dose: 5 × 108 CFU/d. Treatment length: 8 w. | ↓ Liver TG and TC content ↓ Serum AST and ALT levels (all groups) ↓ Serum ALP levels (all groups) ↓ Liver lipid peroxidation | Down-regulation of lipogenic markers in the liver: ↓ Gene expression of Srebp-1c and Fas (all groups). ↓ Protein expression of SREBP-1c (group treated with L. Plantarum strain ATG-K6). ↓ Protein expression of FAS (all groups). ↓ Protein expression of C/EBP (group treated with L. Plantarum strain ATG-K2). ↑ Phosphorylation of ACC (group treated with L. Plantarum strain ATG-K2). Up-regulation of energy yielding pathways in the liver: ↑ Phosphorylation of AMPK (group treated with L. Plantarum strain ATG-K2). Up-regulation of fatty acid oxidation markers in the liver: ↑ Protein expression of CPT-1 (group treated with L. Plantarum strain ATG-K2). Decreased liver MDA content. Modulation of gut microbiota composition: ↓ Relative abundance of Firmicutes (all groups). ↑ Relative abundance of Bacteroidetes (all groups). |
| [30] | Male Wistar rats | HFD (60% energy from fat). | B. animalis subsp. Lactis V9 Daily oral gavage Dose: 1 × 109 CFU/mL. Treatment length: 4 w. | ↓ Liver TG and FFA content ↓ Serum AST and ALT levels ↓ Liver inflammation ↓ Progression to NASH | Down-regulation of lipogenic markers in the liver: ↓ Gene expression of Srebp-1c and Fas. Up-regulation of fatty acid oxidation markers in the liver: ↑ Gene expression of Pparα. Up-regulation of energy yielding pathways in the liver: ↑ Phosphorylation of AMPK. Down-regulation of NASH progression markers in the liver: ↓ Gene expression of Nlrp3, Asc, Tlr-4 and Tlr-9. Down-regulation of pro-inflammatory markers and mediators in the liver: ↓ Gene expression of Tnfα, IL-1β and IL-6. ↓ Phosphorylation of JNK, NF-kB, ERK and AKT. |
| [31] | Male Sprague-Dawley rats | HFD (45% energy from fat). | Eosinophil-Lactobacillus Daily oral gavage (312 mg/kg). Dose: 1 × 107 CFU/g. Treatment length: 8 w. | ↓ Liver lipid content ↓ Liver inflammation ↓ Serum and liver ALT and AST levels | Modulation of gut microbiota composition: ↑ Bacterial diversity. ↓ Pathogenic bacteria content. Up-regulation of liver lipogenesis inhibitors: ↑ Protein expression of FGF15. |
| Reference | Animal Model | Experimental Conditions | Probiotic Treatment | Effects in Liver | Mechanisms of Action |
|---|---|---|---|---|---|
| [44] | Male C57BL/6J mice 5-week-old | HFD (60% energy from fat). | Probiotic mixtures: Bacillus mixture: B. sonorensis JJY12–3, B. paralicheniformis JJY12–8, B. sonorensis JJY13–1, B. sonorensis JJY 13–3 and B. sonorensis JJY 13–8. VSL#3: L. acidophilus, L. plantarum, L. casei, L. delbrueckii subspecies bulgaricus, B. breve, B. longum, B. infantis and S. salivarius subspecies thermophilus Daily administration Dose of 1 × 108 CFU/day Treatment length: 13 w. | ↓ Liver weight (group treated with Bacillus mixture) ↓ Liver fat accumulation (group treated with Bacillus mixture) ↓ Liver TG content (group treated with Bacillus mixture) ↓ Liver inflammation (group treated with Bacillus mixture) | Up-regulation of markers related to fatty acid oxidation in the liver (group treated with Bacillus mixture): ↑ Gene expression of Acox1 and Cpt1. ↑ Protein expression of PCG1α. Down-regulation of pro-inflammatory markers and mediators in the liver (group treated with Bacillus mixture): ↓ Gene expression of Tnfα, Infγ, Mcp-1 and Il-12. Up-regulation of markers of intestinal mucosa integrity (group treated with Bacillus mixture): ↑ Gene expression of Zo1 and Ocln. ↑ Protein expression of Occludin. Modulation of cecum SCFA content and hepatic receptors: ↓ Cecum acetate levels (group treated with Bacillus mixture). ↓ Gene expression of acetate receptor Gpr43 (group treated with Bacillus mixture). |
| [45] | Male C57BL/6 mice 6-week-old | HFD (60% energy from fat). | Probiotic mixture: L. plantarum LC27 B. longum LC67 Daily oral gavage 3:1 proportion (0.75 × 109 CFU of L. plantarum and 0.25 × 109 CFU of B. longum) Treatment length: 4 w. | ↓ Liver weight ↓ Liver lipid accumulation ↓ Liver TG content (group treated with L. plantarum LC27) ↓ NAS (all groups) ↓ Serum AST and ALT levels | Down-regulation of pro-inflammatory markers and mediators in the liver: ↓ Levels of TNFα. ↓ Activity of MPO. ↓ Protein expression of iNOS and COX-2. ↓ Protein expression of p65. ↓ Phosphorylation of p65. ↑ Protein expression of Iκβα. ↓ Phosphorylation of Iκβα. Up-regulation of energy yielding pathways in the liver: ↑ Phosphorylation of AMPK. Down-regulation of periportal fibrogenesis markers in the liver: ↓ Protein expression of α-SMA. ↓ Phosphorylation of α-SMA. Up-regulation of markers of intestinal mucosa integrity: ↑ Protein expression of Occludin-1 and Claudin-1. Modulation of gut microbiota composition: ↓ Proportion of Firmicutes, Bacteroidetes, Proteobacteria, Deferribacteria. ↓ F/B ratio. ↓ P/B ratio. ↑ Proportion of Actinobacteria. ↓ Gut content of LPS. |
| [46] | Male C57BL/6N mice >78-week-old | HFD (60% energy from fat). | Human origin probiotic mixture: 5 strains of Lactobacillus 5 strains of Enterococcus Daily administration Dose: 1 × 109 CFU/mL suspended in the drinking water. Treatment length: 10 w. | ↓ Liver fat accumulation ↓ Liver inflammation | Enhanced microbial diversity enriched in beneficial commensal bacteria. Up-regulation of markers of intestinal mucosa integrity: ↑ Gene expression of Zo1 and Ocln. Modulation of microbial metabolites in the gut: ↑ Abundance of butyrate and propionate. |
| [18] | Male SPF C57BL/6J mice 6-week-old | Normal or a Western diet (42% energy from fat). | Probiotic mixtures: Mix 1: L. casei + L. helveticus Mix 2: L. casei + L. helveticus + P. pentosaceus KID7 Mix 3: L. casei + L. helveticus + L. bulgaricus Daily administration Dose of 1 × 109 CFU/g suspended in distilled water. Treatment length: 8 w. | ↓ Liver steatosis grade (all groups) ↓ Liver inflammation (all groups) ↓ Liver/bw ratio (groups treated with Mix 1 and Mix 2) ↓ NAS (all groups) | Down-regulation of pro-inflammatory markers and mediators in the liver: ↓ Gene expression of Tnfα (all groups). ↓ Gene expression of Il-6 (groups treated with Mix 2 and Mix 3). ↓ Gene expression of Il-1β (group treated with Mix 3). |
| [47] | Male SPF C57BL/6J mice 3–4-week-old | HFD (60% energy from fat). | Probiotic mixture: L. plantarum KLDS1.0344 L. plantarum KLDS1.0386 Daily oral gavage Dose: 108 CFU Treatment length: 8 w. | ↓ Serum ALT and AST levels ↓ Liver TG content | Down-regulation of oxidative stress markers in the liver: ↓ Content of MDA. ↑ Content of GSH-Px. ↑ Content of CAT. ↑ Content of SOD. Modulation of gut microbiota composition: - Relative abundance of Parabacteroides, Eubacterium xylanophilum group, GCA-900066575, Lachnoclostridium, Lachnospiraceae UCG-006 and Rombustia. ↑ Richness of Lachnospiraceae NK4A136 group and Bacteroides. Modulation of gut SCFA production: ↑ Levels of acetic and butyric acids. |
| [48] | Male C57BL/6J mice 5-week-old | HFD (60% energy from fat). | Probiotic mixture: B. subtilis (1.4 × 109 CFU) E. faecium (1.55 × 1010 CFU) Daily oral gavage. Treatment length: 16 w. | ↓ Liver index ↓ Serum ALT and AST levels ↓ Liver lipid accumulation | Up-regulation of fatty acid oxidation markers in the liver: ↑ Protein expression of CPT1 and PPARα. Down-regulation of pro-inflammatory markers and mediators in the liver: ↓ LPS levels. ↓ Gene expression of Il-1β, Il-6 and Tnf-α. ↓ Protein expression of TLR-4 and NF-κβ. Up-regulation of markers of intestinal mucosa integrity: ↑ Protein expression of Occludin-1 and Claudin-1. Modulation of gut microbiota composition: ↓ F/B ratio. ↓ Relative abundance of Firmicutes. ↑ Relative abundance of Bacteroidetes, Verrucomicrobia, Akkermansia and Oscillibacter. |
| [49] | Male Zucker-Leprfa/fa rats | Chow diet. | Probiotic mixture: L. paracasei CNCM I-4034 B. breve CNCM I-4035 Daily oral gavage Dose: 1010 CFU. Treatment length: 30 days. | ↓ Liver TG content | Decreased serum levels of pro-inflammatory markers cytokines and mediators: ↓ Levels of TNF-α. ↓ Levels of LPS. |
| [50] | Male albino rats 6-week-old | HFSD (59% energy from fat). | Probiotic mixture: L. acidophilus (10 × 108 CFU/g). L. plantarum (9.8 × 107 CFU/g). B. bifidum (2 × 106 CFU/g). B. subtilis fermentation extract (50 g per kg of product). A. oryzae fermentation extract (50 g per kg of product). Daily administration Dose: 1 g of probiotic mixture/kg diet Treatment length: 4 w. | ↓ Serum ALT levels ↓ NAS ↓ Liver inflammation | Not specified. |
| [51] | Male Sprague-Dawley rats 7-week-old | HCD (15% energy from fat). | Probiotic mixture: B. longum CBG-C11 B. lactis CBG-C10 B. breve CBG-C2 L. reuteri CBG-C15 L. plantarum CBG-C21 Daily by oral gavage Doses: Low dose (1.65 × 109 CFU/kg/d) Medium dose (5.5 × 109 CFU/kg/d) High dose (1.65 × 1010 CFU/kg/d) Treatment length: 8 w. | ↓ Hepatic steatosis score (group treated with the high dose) ↓ Liver TG and TC content (all groups) ↓ Liver AST and ALT levels (all groups) | Down-regulation of lipogenic markers in the liver: ↓ Protein expression of FAS, ACC and SREBP-1 (all doses). |
| [35] | Male Sprague-Dawley rats 42-day-old | STD + 20% fructose in drinking water. | Probiotic mixture: L. acidophilus B. coagulans L. casei L. reuteri Daily administration Dose: 1 × 109 CFU/mL suspended in drinking water. Treatment length: 16 w. | ↓ Liver TG content ↓ Serum ALT levels ↓ Liver oxidative stress | Up-regulation of antioxidant response in the liver: ↑ Content of glutathione. ↓ Liver ROS formation. ↑ Liver total antioxidant level. ↓ Liver protein-carbonylation. ↓ Liver lipid peroxidation. |
| Reference | Population | Intervention | Comparison | Outcome |
|---|---|---|---|---|
| [53] | 28 adults with NAFLD 20 men 8 women. | Daily consumption Probiotic mixture (5 × 108 CFU): L. bulgaricus S. thermophilus Treatment period: 3 months. | Placebo. | ↓ Serum ALT, AST and GGT levels. |
| [59] | 72 obese adults with NAFLD 33 men 39 women Age: 23–63 years old. BMI: 25–40 kg/m2 | Daily consumption Probiotic-enriched yogurt (300 g/d): L. bulgaricus S. thermophilus L. acidophilus (6.46 × 106 CFU/g) B. lactis Bb12 (4.97 × 106 CFU/g) Treatment period: 8 weeks | Daily consumption Conventional yogurt (300 g/d) L. bulgaricus S. thermophilus | ↓ Serum ALT and AST levels. |
| [54] | 64 obese adolescents with NAFLD Age: 10–18 years old BMI >85th percentile (age and sex specific) | Daily consumption Probiotic mixture (1 capsule): L. acidophilus ATCC B3208 (3 × 109 CFU) B. lactis DSMZ 32269 (6 × 109 CFU) B. bifidum ATCC SD6576 (2 × 109 CFU) L. rhamnosus DSMZ 21690 (2 × 109 CFU) Treatment period: 12 weeks | Placebo. | ↓ Serum ALT and AST levels. ↓ Fatty liver grade (sonographic grading). |
| [55] | 58 adult patients with NAFLD and T2DM Age: 18–65 years old BMI > 25 kg/m2 | Daily consumption Probiotic mixture (1 sachet of 10 g). Lactobacillus + Lactococcus (6 × 1010 CFU/g) Bifidobacterium (1 × 1010 CFU/g) Propionibacterium (3 × 1010 CFU/g) Acetobacter (1 × 106 CFU/g) Treatment period: 8 weeks | Placebo. | ↓ FLI and LS. ↓ Serum AST and GGT levels. Decreased circulating levels of proinflammatory markers: ↓ Serum TNF-α and IL-6 levels. |
| [56] | 65 obese adults with NAFLD 33 men 32 women Age: 19–75 years old BMI >25 kg/m2 Mean hepatic MRI-PDFF 16.2% | Daily consumption Probiotic mixture (containing 1 × 109 CFU/1.4 g) L. acidophilus CBT LA1 L. rhamnosus CBT LR5 (human feces) L. paracasei CBT LPC5 (Korean fermented food—jeotgal) P. pentosaceus CBT SL4 (Korean fermented vegetable product—kimchi) B. lactis CBT BL3 B. breve CBT BR3 (Korean infant feces) Treatment period: 12 weeks | Placebo. | ↓ IHF fraction. Modulation of gut microbiota composition: ↑ Relative abundances of L. acidophilus L. rhamnosus P. pentosaceus B. lactis B. breve |
| [57] | 30 adults with NAFLD: [ALT] and [AST] >1.5-fold normal levels | Daily consumption Probiotic mixture (2 capsules containing 11.25 × 1010 CFU, each) L. paracasei DSM 24733, L. plantarum DSM 24730 L. acidophilus DSM 24735 L. delbrueckii subsp. Bulgaricus DSM 24734 B. longum DSM 24736 B. infantis DSM 24737 B. breve DSM 24732 S. thermophilus DSM 24731 Treatment period: 12 months | Placebo. | Improvement of liver histology: ↓ Hepatocyte ballooning. ↓ Lobular inflammation. ↓ NAS score. ↓ Serum ALT levels. ↓ Serum ALP levels. ↓ Serum pro-inflammatory cytokines: Il-1β, IL-6 and TNF-α. |
| [58] | 60 adults with NAFLD 43 men 17 women Age: 20–60 years old BMI: 20–40 kg/m2 | Daily consumption Probiotic mixture (1 capsule containing 5 × 109 CFU) L. casei L. rhamnosus L. acidophilus B. longum B. breve Treatment period: 12 weeks | Placebo. | ↓ Serum ALT, AST and GGT levels ↓ Serum ALP levels. |
| [60] | 35 adults with NAFLD 28 men 7 women Age: 25–70 years old Mean BMI 32.6 ± 5.0 kg/m2 | Daily consumption Probiotic mixture (2 sachets of VSL#3 twice daily) S. thermophilus B. breve B. infantis B. longum L. acidophilus L. plantarum L. paracasei L delbrueckii subsp. bulgaricus Treatment period: 10 weeks | Placebo. | No significant improvements in markers of liver injury. |
| [61] | 39 obese adults 28 men 11 women with NAFLD: Fatty liver score >263 dB/m | Daily consumption Probiotic mixture (1 sachet containing 3 × 1010 CFU twice daily) L. acidophilus BCMC 12,130 (107 mg) L. lactis MCMC 12,451 (107 mg) B. bifidum BCMC 02290 (107 mg) B. infantis BCMC 02129 (107 mg) B. longum BCMC 02120 (107 mg) Treatment period: 6 months | Placebo. | No significant improvements in liver fibrosis parameters or serum markers of inflammation. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arellano-García, L.; Portillo, M.P.; Martínez, J.A.; Milton-Laskibar, I. Usefulness of Probiotics in the Management of NAFLD: Evidence and Involved Mechanisms of Action from Preclinical and Human Models. Int. J. Mol. Sci. 2022, 23, 3167. https://doi.org/10.3390/ijms23063167
Arellano-García L, Portillo MP, Martínez JA, Milton-Laskibar I. Usefulness of Probiotics in the Management of NAFLD: Evidence and Involved Mechanisms of Action from Preclinical and Human Models. International Journal of Molecular Sciences. 2022; 23(6):3167. https://doi.org/10.3390/ijms23063167
Chicago/Turabian StyleArellano-García, Laura, María P. Portillo, J. Alfredo Martínez, and Iñaki Milton-Laskibar. 2022. "Usefulness of Probiotics in the Management of NAFLD: Evidence and Involved Mechanisms of Action from Preclinical and Human Models" International Journal of Molecular Sciences 23, no. 6: 3167. https://doi.org/10.3390/ijms23063167