
Exploratory Genome-Wide Association Analysis to Identify Pharmacogenetic Determinants of Response to R-CHOP in Diffuse Large B-Cell Lymphoma
 , , , , and add
Show full author list
, , , , and add
Show full author list
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants and Treatment
2.2. DNA Extraction and Genotyping Platform
2.3. Bioinformatic Analysis
2.4. Patient Population and Quality Control of Genotypic Data
2.5. R-CHOP Efficacy and Toxicity Evaluation
2.6. Statistical Analysis
2.7. Functional Enrichment and Network Analysis
3. Results
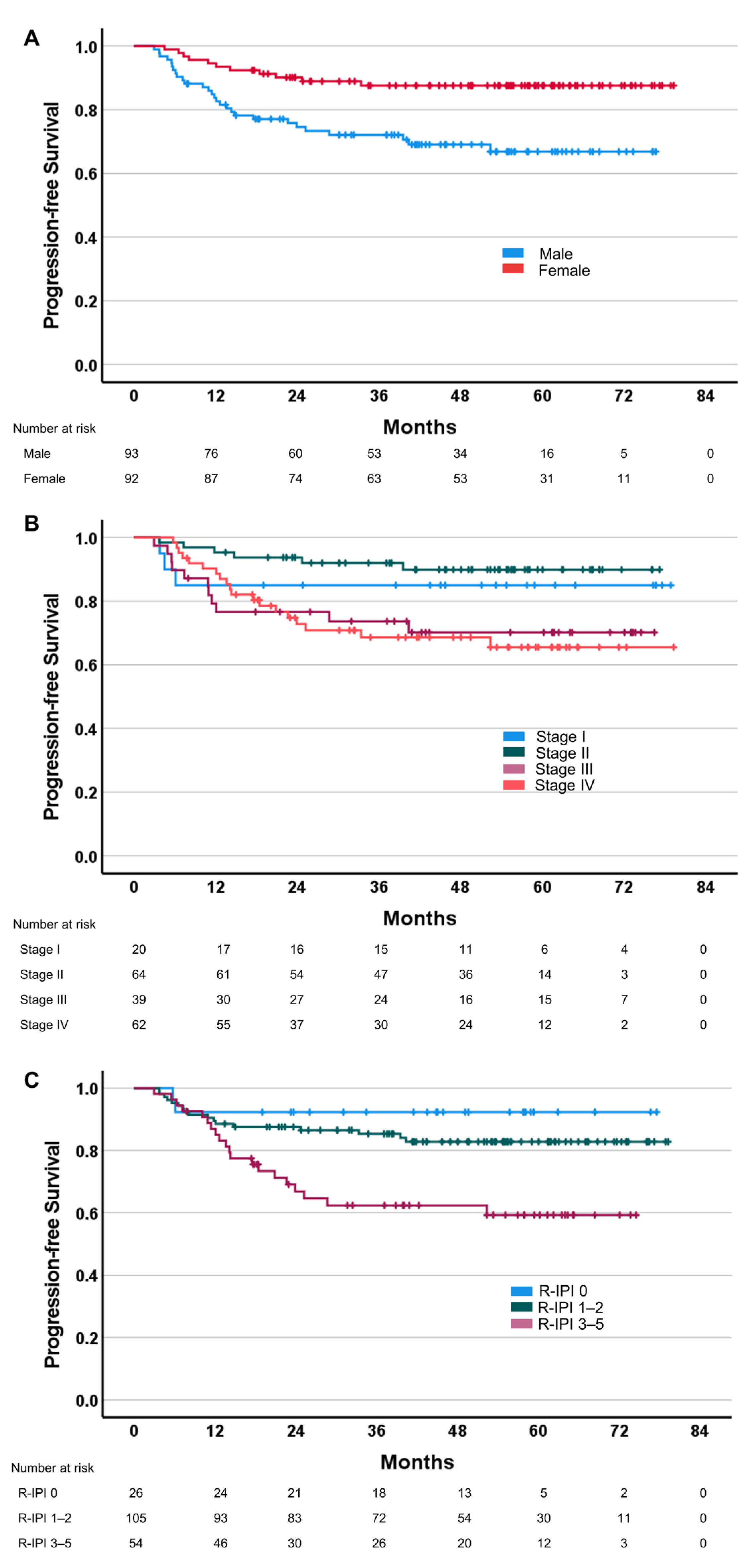
3.1. Relationships between Efficacy Parameters and Clinical/Pathological Characteristics
3.2. Relationships between Efficacy Parameters and SNPs
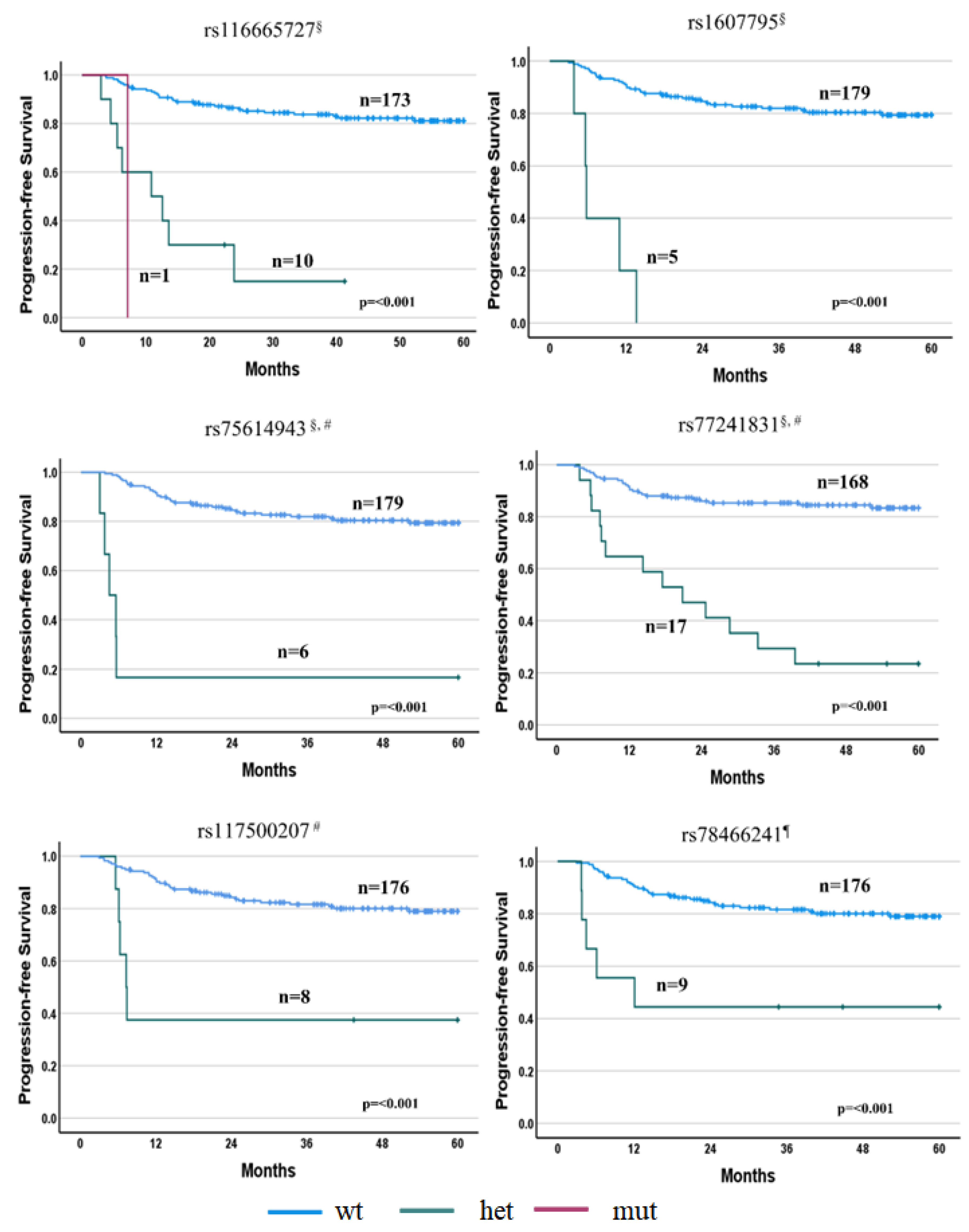
3.2.1. PFS Associations
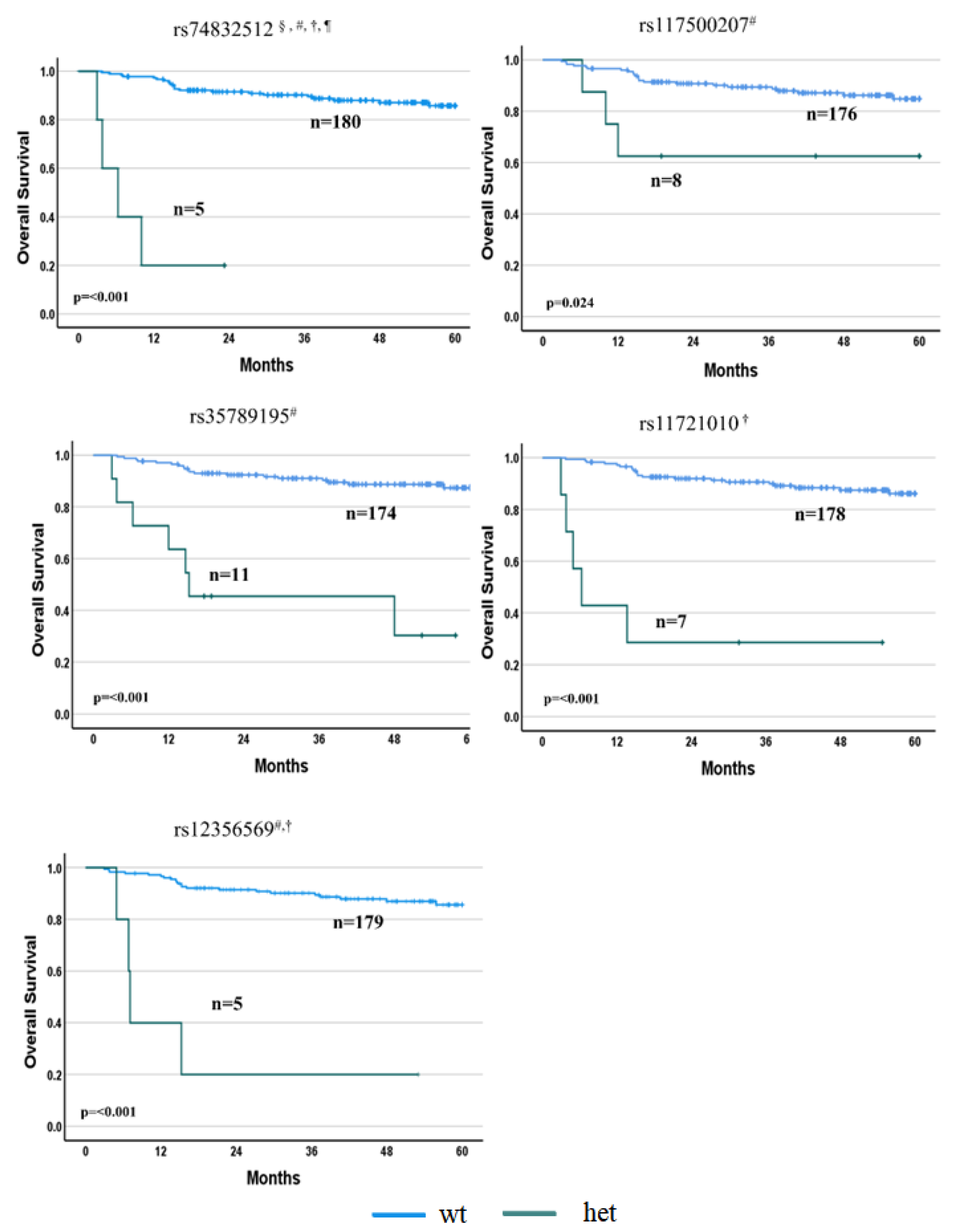
3.2.2. OS Associations
3.3. Relationships between Toxicity and Clinical/Pathological Characteristics
3.4. Relationships between Toxicity and SNPs
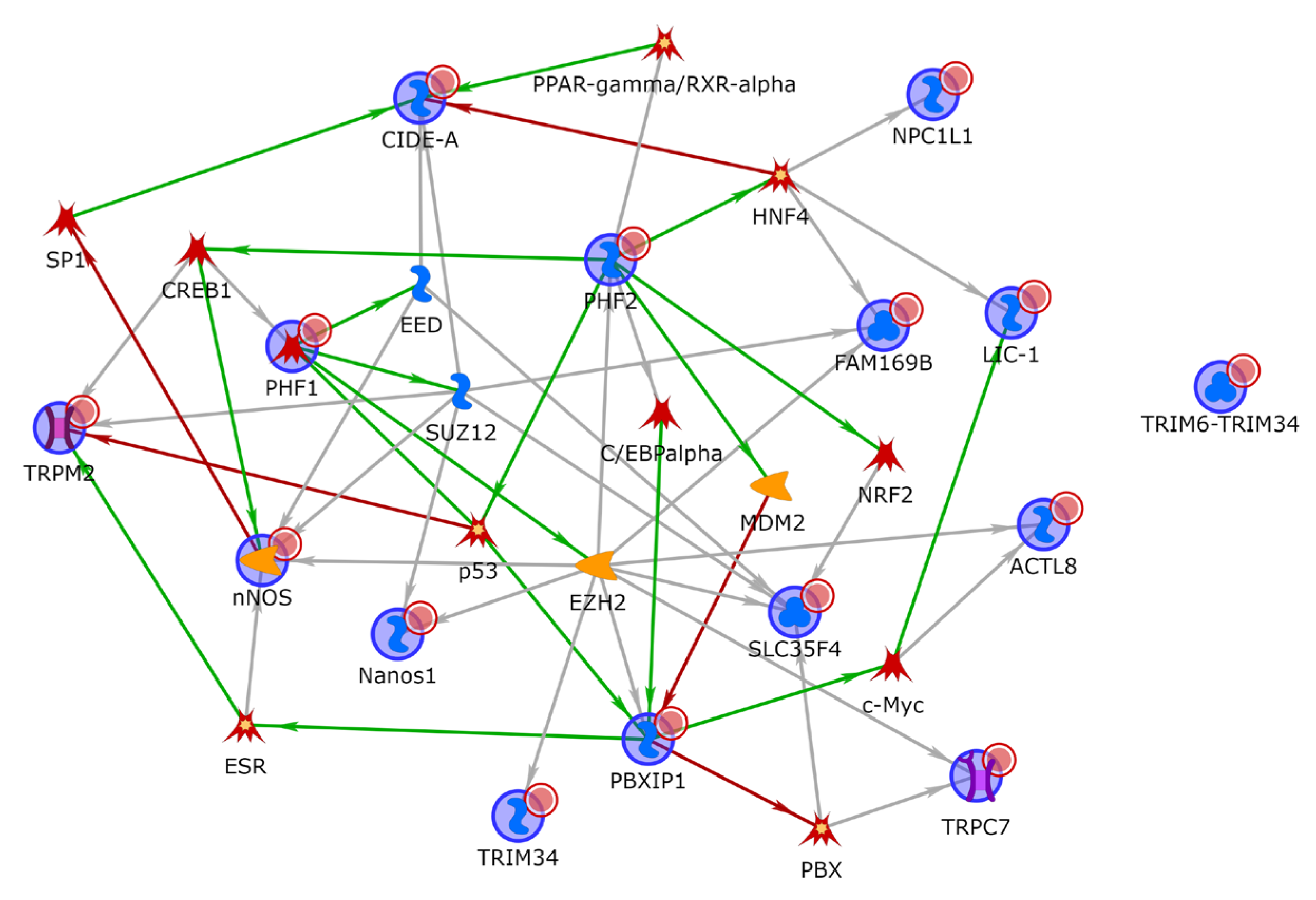
3.5. Functional Enrichment and Network Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Wagle, N.S.; Jemal, A. Cancer statistics, 2023. CA Cancer J. Clin. 2023, 73, 17–48. [Google Scholar] [CrossRef]
- Armitage, J.O.; Gascoyne, R.D.; Lunning, M.A.; Cavalli, F. Non-Hodgkin lymphoma. Lancet 2017, 390, 298–310. [Google Scholar] [CrossRef] [PubMed]
- Alaggio, R.; Amador, C.; Anagnostopoulos, I.; Attygalle, A.D.; Araujo, I.B.O.; Berti, E.; Bhagat, G.; Borges, A.M.; Boyer, D.; Calaminici, M.; et al. The 5th edition of the World Health Organization Classification of Haematolymphoid Tumours: Lymphoid Neoplasms. Leukemia 2022, 36, 1720–1748. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh, A.A.; Eisen, M.B.; Davis, R.E.; Ma, C.; Lossos, I.S.; Rosenwald, A.; Boldrick, J.C.; Sabet, H.; Tran, T.; Yu, X.; et al. Distinct types of diffuse large B-cell lymphoma identified by gene expression profiling. Nature 2000, 403, 503–511. [Google Scholar] [CrossRef] [PubMed]
- Rosenwald, A.; Wright, G.; Chan, W.C.; Connors, J.M.; Campo, E.; Fisher, R.I.; Gascoyne, R.D.; Muller-Hermelink, H.K.; Smeland, E.B.; Giltnane, J.M.; et al. The use of molecular profiling to predict survival after chemotherapy for diffuse large-B-cell lymphoma. N. Engl. J. Med. 2002, 346, 1937–1947. [Google Scholar] [CrossRef]
- Lenz, G.; Wright, G.; Dave, S.S.; Xiao, W.; Powell, J.; Zhao, H.; Xu, W.; Tan, B.; Goldschmidt, N.; Iqbal, J.; et al. Stromal gene signatures in large-B-cell lymphomas. N. Engl. J. Med. 2008, 359, 2313–2323. [Google Scholar] [CrossRef]
- Holmes, A.B.; Corinaldesi, C.; Shen, Q.; Kumar, R.; Compagno, N.; Wang, Z.; Nitzan, M.; Grunstein, E.; Pasqualucci, L.; Dalla-Favera, R.; et al. Single-cell analysis of germinal-center B cells informs on lymphoma cell of origin and outcome. J. Exp. Med. 2020, 217, e20200483. [Google Scholar] [CrossRef]
- Meyer, P.N.; Fu, K.; Greiner, T.C.; Smith, L.M.; Delabie, J.; Gascoyne, R.D.; Ott, G.; Rosenwald, A.; Braziel, R.M.; Campo, E.; et al. Immunohistochemical methods for predicting cell of origin and survival in patients with diffuse large B-cell lymphoma treated with rituximab. J. Clin. Oncol. 2011, 29, 200–207. [Google Scholar] [CrossRef]
- Scott, D.W. Cell-of-Origin in Diffuse Large B-Cell Lymphoma: Are the Assays Ready for the Clinic? Am. Soc. Clin. Oncol. Educ. Book 2015, 35, e458–e466. [Google Scholar] [CrossRef]
- Hans, C.P.; Weisenburger, D.D.; Greiner, T.C.; Gascoyne, R.D.; Delabie, J.; Ott, G.; Müller-Hermelink, H.K.; Campo, E.; Braziel, R.M.; Jaffe, E.S.; et al. Confirmation of the molecular classification of diffuse large B-cell lymphoma by immunohistochemistry using a tissue microarray. Blood 2004, 103, 275–282. [Google Scholar] [CrossRef]
- Chapuy, B.; Stewart, C.; Dunford, A.J.; Kim, J.; Kamburov, A.; Redd, R.A.; Lawrence, M.S.; Roemer, M.G.M.; Li, A.J.; Ziepert, M.; et al. Molecular subtypes of diffuse large B cell lymphoma are associated with distinct pathogenic mechanisms and outcomes. Nat. Med. 2018, 24, 679–690. [Google Scholar] [CrossRef] [PubMed]
- Vermaat, J.S.; Pals, S.T.; Younes, A.; Dreyling, M.; Federico, M.; Aurer, I.; Radford, J.; Kersten, M.J.; EHA Lymphoma Group, a Scientific Working Group of the European Hematology Association. Precision medicine in diffuse large B-cell lymphoma: Hitting the target. Haematologica 2015, 100, 989–993. [Google Scholar]
- Coiffier, B.; Sarkozy, C. Diffuse large B-cell lymphoma: R-CHOP failure-what to do? Hematol. Am. Soc. Hematol. Educ. Program 2016, 2016, 366–378. [Google Scholar] [CrossRef]
- Pasqualucci, L.; Dalla-Favera, R. Genetics of diffuse large B-cell lymphoma. Blood 2018, 131, 2307–2319. [Google Scholar] [CrossRef] [PubMed]
- Sehn, L.H.; Berry, B.; Chhanabhai, M.; Fitzgerald, C.; Gill, K.; Hoskins, P.; Klasa, R.; Savage, K.J.; Shenkier, T.; Sutherland, J.; et al. The revised International Prognostic Index (R-IPI) is a better predictor of outcome than the standard IPI for patients with diffuse large B-cell lymphoma treated with R-CHOP. Blood 2007, 109, 1857–1861. [Google Scholar] [CrossRef] [PubMed]
- Kubuschok, B.; Held, G.; Pfreundschuh, M. Management of diffuse large B-cell lymphoma (DLBCL). Cancer Treat. Res. 2015, 165, 271–288. [Google Scholar] [CrossRef]
- González-Barca, E.; Boumendil, A.; Blaise, D.; Trněný, M.; Masszi, T.; Finel, H.; Michieli, M.G.; Bittenbring, J.T.; Gritti, G.; Snowden, J.A.; et al. Outcome in patients with diffuse large B-cell lymphoma who relapse after autologous stem cell transplantation and receive active therapy. A retrospective analysis of the Lymphoma Working Party of the European Society for Blood and Marrow Transplantation (EBMT). Bone Marrow Transpl. 2020, 55, 393–399. [Google Scholar] [CrossRef]
- Coiffier, B.; Lepage, E.; Briere, J.; Herbrecht, R.; Tilly, H.; Bouabdallah, R.; Morel, P.; Van Den Neste, E.; Salles, G.; Gaulard, P.; et al. CHOP chemotherapy plus rituximab compared with CHOP alone in elderly patients with diffuse large-B-cell lymphoma. N. Engl. J. Med. 2002, 346, 235–242. [Google Scholar] [CrossRef]
- Sehn, L.H.; Donaldson, J.; Chhanabhai, M.; Fitzgerald, C.; Gill, K.; Klasa, R.; MacPherson, N.; O’Reilly, S.; Spinelli, J.J.; Sutherland, J.; et al. Introduction of combined CHOP plus rituximab therapy dramatically improved outcome of diffuse large B-cell lymphoma in British Columbia. J. Clin. Oncol. 2005, 23, 5027–5033. [Google Scholar] [CrossRef]
- Pfreundschuh, M.; Trümper, L.; Osterborg, A.; Pettengell, R.; Trneny, M.; Imrie, K.; Ma, D.; Gill, D.; Walewski, J.; Zinzani, P.L.; et al. CHOP-like chemotherapy plus rituximab versus CHOP-like chemotherapy alone in young patients with good-prognosis diffuse large-B-cell lymphoma: A randomised controlled trial by the MabThera International Trial (MInT) Group. Lancet Oncol. 2006, 7, 379–391. [Google Scholar] [CrossRef]
- Coiffier, B.; Thieblemont, C.; Van Den Neste, E.; Lepeu, G.; Plantier, I.; Castaigne, S.; Lefort, S.; Marit, G.; Macro, M.; Sebban, C.; et al. Long-term outcome of patients in the LNH-98.5 trial, the first randomized study comparing rituximab-CHOP to standard CHOP chemotherapy in DLBCL patients: A study by the Groupe d’Etudes des Lymphomes de l’Adulte. Blood 2010, 116, 2040–2045. [Google Scholar] [CrossRef] [PubMed]
- Pfreundschuh, M.; Kuhnt, E.; Trümper, L.; Osterborg, A.; Trneny, M.; Shepherd, L.; Gill, D.S.; Walewski, J.; Pettengell, R.; Jaeger, U.; et al. CHOP-like chemotherapy with or without rituximab in young patients with good-prognosis diffuse large-B-cell lymphoma: 6-year results of an open-label randomised study of the MabThera International Trial (MInT) Group. Lancet Oncol. 2011, 12, 1013–1022. [Google Scholar] [CrossRef] [PubMed]
- NCCN Clinical Practice Guidelines in Oncology B-Cell lymphomas (V. 1.2023). 2023. Available online: https://www.nccn.org/professionals/physician_gls/pdf/b-cell.pdf (accessed on 14 April 2023).
- Tilly, H.; Morschhauser, F.; Sehn, L.H.; Friedberg, J.W.; Trněný, M.; Sharman, J.P.; Herbaux, C.; Burke, J.M.; Matasar, M.; Rai, S.; et al. Polatuzumab Vedotin in Previously Untreated Diffuse Large B-Cell Lymphoma. N. Engl. J. Med. 2022, 386, 351–363. [Google Scholar] [CrossRef] [PubMed]
- Dunleavy, K. Optimal Management of Double-Hit Lymphoma. J. Oncol. Pract. 2016, 12, 241–242. [Google Scholar] [CrossRef] [PubMed]
- Johnson, N.A.; Slack, G.W.; Savage, K.J.; Connors, J.M.; Ben-Neriah, S.; Rogic, S.; Scott, D.W.; Tan, K.L.; Steidl, C.; Sehn, L.H.; et al. Concurrent expression of MYC and BCL2 in diffuse large B-cell lymphoma treated with rituximab plus cyclophosphamide, doxorubicin, vincristine, and prednisone. J. Clin. Oncol. 2012, 30, 3452–3459. [Google Scholar] [CrossRef]
- Tafasitamab + Lenalidomide + R-CHOP Versus R-CHOP in Newly Diagnosed High-intermediate and High Risk DLBCL Patients (frontMIND). Available online: https://clinicaltrials.gov/ct2/show/NCT04824092 (accessed on 14 April 2023).
- El Hussein, S.; Shaw, K.R.M.; Vega, F. Evolving insights into the genomic complexity and immune landscape of diffuse large B-cell lymphoma: Opportunities for novel biomarkers. Mod. Pathol. 2020, 33, 2422–2436. [Google Scholar] [CrossRef]
- Rossi, D.; Diop, F.; Spaccarotella, E.; Monti, S.; Zanni, M.; Rasi, S.; Deambrogi, C.; Spina, V.; Bruscaggin, A.; Favini, C.; et al. Diffuse large B-cell lymphoma genotyping on the liquid biopsy. Blood 2017, 129, 1947–1957. [Google Scholar] [CrossRef]
- Mini, E.; Nobili, S. Pharmacogenetics of Cancer; OAE Publishing Inc.: Alhambra, CA, USA, 2020. [Google Scholar]
- Lavacchi, D.; Landini, I.; Perrone, G.; Roviello, G.; Mini, E.; Nobili, S. Pharmacogenetics in diffuse large B-cell lymphoma treated with R-CHOP: Still an unmet challenge. Pharmacol. Ther. 2022, 229, 107924. [Google Scholar] [CrossRef]
- Falduto, A.; Cimino, F.; Speciale, A.; Musolino, C.; Gangemi, S.; Saija, A.; Allegra, A. How gene polymorphisms can influence clinical response and toxicity following R-CHOP therapy in patients with diffuse large B cell lymphoma. Blood Rev. 2017, 31, 235–249. [Google Scholar] [CrossRef] [PubMed]
- Low, S.K.; Takahashi, A.; Mushiroda, T.; Kubo, M. Genome-wide association study: A useful tool to identify common genetic variants associated with drug toxicity and efficacy in cancer pharmacogenomics. Clin. Cancer Res. 2014, 20, 2541–2552. [Google Scholar] [CrossRef]
- Kiyotani, K.; Mushiroda, T.; Tsunoda, T.; Morizono, T.; Hosono, N.; Kubo, M.; Tanigawara, Y.; Imamura, C.K.; Flockhart, D.A.; Aki, F.; et al. A genome-wide association study identifies locus at 10q22 associated with clinical outcomes of adjuvant tamoxifen therapy for breast cancer patients in Japanese. Hum. Mol. Genet. 2012, 21, 1665–1672. [Google Scholar] [CrossRef]
- Wu, X.; Ye, Y.; Rosell, R.; Amos, C.I.; Stewart, D.J.; Hildebrandt, M.A.; Roth, J.A.; Minna, J.D.; Gu, J.; Lin, J.; et al. Genome-wide association study of survival in non-small cell lung cancer patients receiving platinum-based chemotherapy. J. Natl. Cancer Inst. 2011, 103, 817–825. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Wu, C.; Zhao, X.; Heist, R.; Su, L.; Zhao, Y.; Han, B.; Cao, S.; Chu, M.; Dai, J.; et al. Genome-wide association study of prognosis in advanced non-small cell lung cancer patients receiving platinum-based chemotherapy. Clin. Cancer Res. 2012, 18, 5507–5514. [Google Scholar] [CrossRef] [PubMed]
- Innocenti, F.; Owzar, K.; Cox, N.L.; Evans, P.; Kubo, M.; Zembutsu, H.; Jiang, C.; Hollis, D.; Mushiroda, T.; Li, L.; et al. A genome-wide association study of overall survival in pancreatic cancer patients treated with gemcitabine in CALGB 80303. Clin. Cancer Res. 2012, 18, 577–584. [Google Scholar] [CrossRef] [PubMed]
- Diekstra, M.H.M.; Swen, J.J.; van der Zanden, L.F.M.; Vermeulen, S.H.; Boven, E.; Mathijssen, R.H.J.; Fukunaga, K.; Mushiroda, T.; Hongo, F.; Oosterwijk, E.; et al. Genome-Wide Meta-Analysis Identifies Variants in DSCAM and PDLIM3 That Correlate with Efficacy Outcomes in Metastatic Renal Cell Carcinoma Patients Treated with Sunitinib. Cancers 2022, 14, 2838. [Google Scholar] [CrossRef]
- Lin, C.L.; Liang, K.H.; Hu, C.C.; Chien, C.H.; Chen, L.W.; Chien, R.N.; Lin, Y.H.; Yeh, C.T. A Single Nucleotide Polymorphism rs1010816 Predicts Sorafenib Therapeutic Outcomes in Advanced Hepatocellular Carcinoma. Int. J. Mol. Sci. 2023, 24, 1681. [Google Scholar] [CrossRef]
- Ghesquieres, H.; Slager, S.L.; Jardin, F.; Veron, A.S.; Asmann, Y.W.; Maurer, M.J.; Fest, T.; Habermann, T.M.; Bene, M.C.; Novak, A.J.; et al. Genome-Wide Association Study of Event-Free Survival in Diffuse Large B-Cell Lymphoma Treated With Immunochemotherapy. J. Clin. Oncol. 2015, 33, 3930–3937. [Google Scholar] [CrossRef]
- Swerdlow, S.H.; Campo, E.; Harris, N.L.; Jaffe, E.S.; Pileri, S.A.; Stein, H.; Thiele, J.; Vardiman, J.W. WHO Classification of Tumours of Haematopoietic and Lymphoid Tissues-WHO Classification of Tumours, 4th ed.; United States Environmental Protection Agency: Washington, DC, USA, 2008; Volume 2. [Google Scholar]
- Swerdlow, S.H.; Campo, E.; Harris, N.L.; Jaffe, E.S.; Pileri, S.A.; Stein, H.; Thiele, J. WHO Classification of Tumours of Haematopoietic and Lymphoid Tissues-WHO Classification of Tumours, revised, 4th ed.; United States Environmental Protection Agency: Washington, DC, USA, 2017; Volume 2. [Google Scholar]
- Thermo Fisher Scientific. Axiom Genotyping Solution Data Analysis User Guide; Publication Number MAN0018363 Tech; Thermo Fisher Scientific: Hong Kong, China, 2020. [Google Scholar]
- Laurie, C.C.; Doheny, K.F.; Mirel, D.B.; Pugh, E.W.; Bierut, L.J.; Bhangale, T.; Boehm, F.; Caporaso, N.E.; Cornelis, M.C.; Edenberg, H.J.; et al. Quality control and quality assurance in genotypic data for genome-wide association studies. Genet Epidemiol. 2010, 34, 591–602. [Google Scholar] [CrossRef]
- Cheson, B.D.; Pfistner, B.; Juweid, M.E.; Gascoyne, R.D.; Specht, L.; Horning, S.J.; Coiffier, B.; Fisher, R.I.; Hagenbeek, A.; Zucca, E.; et al. Revised response criteria for malignant lymphoma. J. Clin. Oncol. 2007, 25, 579–586. [Google Scholar] [CrossRef]
- Cheson, B.D.; Fisher, R.I.; Barrington, S.F.; Cavalli, F.; Schwartz, L.H.; Zucca, E.; Lister, T.A. Recommendations for initial evaluation, staging, and response assessment of Hodgkin and non-Hodgkin lymphoma: The Lugano classification. J. Clin. Oncol. 2014, 32, 3059–3068. [Google Scholar] [CrossRef]
- Nobili, S.; Napoli, C.; Puccini, B.; Landini, I.; Perrone, G.; Brugia, M.; Benelli, G.; Doria, M.; Martelli, M.; Finolezzi, E.; et al. Identification of pharmacogenomic markers of clinical efficacy in a dose-dense therapy regimen (R-CHOP14) in diffuse large B-cell lymphoma. Leuk Lymphoma 2014, 55, 2071–2078. [Google Scholar] [CrossRef] [PubMed]
- Bouroumeau, A.; Bussot, L.; Bonnefoix, T.; Fournier, C.; Chapusot, C.; Casasnovas, O.; Martin, L.; McLeer, A.; Col, E.; David-Boudet, L.; et al. c-MYC and p53 expression highlight starry-sky pattern as a favourable prognostic feature in R-CHOP-treated diffuse large B-cell lymphoma. J. Pathol. Clin. Res. 2021, 7, 604–615. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Fu, D.; Xu, P.P.; Cheng, S.; Wang, L.; Zhang, Y.Z.; Zhao, W.L. Implication of immune cell signature of tumor microenvironment in diffuse large B-cell lymphoma. Hematol. Oncol. 2021, 39, 616–624. [Google Scholar] [CrossRef]
- Kurtz, D.M.; Scherer, F.; Jin, M.C.; Soo, J.; Craig, A.F.M.; Esfahani, M.S.; Chabon, J.J.; Stehr, H.; Liu, C.L.; Tibshirani, R.; et al. Circulating Tumor DNA Measurements As Early Outcome Predictors in Diffuse Large B-Cell Lymphoma. J. Clin. Oncol. 2018, 36, 2845–2853. [Google Scholar] [CrossRef] [PubMed]
- Rivas-Delgado, A.; Nadeu, F.; Enjuanes, A.; Casanueva-Eliceiry, S.; Mozas, P.; Magnano, L.; Castrejón de Anta, N.; Rovira, J.; Dlouhy, I.; Martín, S.; et al. Mutational Landscape and Tumor Burden Assessed by Cell-free DNA in Diffuse Large B-Cell Lymphoma in a Population-Based Study. Clin. Cancer Res. 2021, 27, 513–521. [Google Scholar] [CrossRef]
- Li, M.; Mi, L.; Wang, C.; Wang, X.; Zhu, J.; Qi, F.; Yu, H.; Ye, Y.; Wang, D.; Cao, J.; et al. Clinical implications of circulating tumor DNA in predicting the outcome of diffuse large B cell lymphoma patients receiving first-line therapy. BMC Med. 2022, 20, 369. [Google Scholar] [CrossRef]
- Mendes, R.V.; Martins, A.R.; de Nucci, G.; Murad, F.; Soares, F.A. Expression of nitric oxide synthase isoforms and nitrotyrosine immunoreactivity by B-cell non-Hodgkin’s lymphomas and multiple myeloma. Histopathology 2001, 39, 172–178. [Google Scholar] [CrossRef]
- Han, X.; Zheng, T.; Lan, Q.; Zhang, Y.; Kilfoy, B.A.; Qin, Q.; Rothman, N.; Zahm, S.H.; Holford, T.R.; Leaderer, B. Genetic polymorphisms in nitric oxide synthase genes modify the relationship between vegetable and fruit intake and risk of non-Hodgkin lymphoma. Cancer Epidemiol. Biomark. Prev. 2009, 18, 1429–1438. [Google Scholar] [CrossRef]
- Kim, C.; Zheng, T.; Lan, Q.; Chen, Y.; Foss, F.; Chen, X.; Holford, T.; Leaderer, B.; Boyle, P.; Chanock, S.J.; et al. Genetic polymorphisms in oxidative stress pathway genes and modification of BMI and risk of non-Hodgkin lymphoma. Cancer Epidemiol. Biomark. Prev. 2012, 21, 866–868. [Google Scholar] [CrossRef]
- Slayton, M.; Gupta, A.; Balakrishnan, B.; Puri, V. CIDE Proteins in Human Health and Disease. Cells 2019, 8, 238. [Google Scholar] [CrossRef]
- Laurencikiene, J.; Stenson, B.M.; Arvidsson Nordström, E.; Agustsson, T.; Langin, D.; Isaksson, B.; Permert, J.; Rydén, M.; Arner, P. Evidence for an important role of CIDEA in human cancer cachexia. Cancer Res. 2008, 68, 9247–9254. [Google Scholar] [CrossRef] [PubMed]
- Nititham, J.; Taylor, K.E.; Gupta, R.; Chen, H.; Ahn, R.; Liu, J.; Seielstad, M.; Ma, A.; Bowcock, A.M.; Criswell, L.A.; et al. Meta-analysis of the TNFAIP3 region in psoriasis reveals a risk haplotype that is distinct from other autoimmune diseases. Genes Immun. 2015, 16, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Song, B.; Li, H.; Guo, S.; Yang, T.; Li, L.; Cao, L.; Wang, J. LINC00882 Plays a Tumor-promoter Role in Colorectal Cancer by Targeting miR-3619-5p to Up-regulate CTNNB1. Arch. Med. Res. 2022, 53, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Melenotte, C.; Mezouar, S.; Ben Amara, A.; Benatti, S.; Chiaroni, J.; Devaux, C.; Costello, R.; Kroemer, G.; Mege, J.L.; Raoult, D. A transcriptional signature associated with non-Hodgkin lymphoma in the blood of patients with Q fever. PLoS ONE 2019, 14, e0217542. [Google Scholar] [CrossRef]
- Hezaveh, K.; Kloetgen, A.; Bernhart, S.H.; Mahapatra, K.D.; Lenze, D.; Richter, J.; Haake, A.; Bergmann, A.K.; Brors, B.; Burkhardt, B.; et al. Alterations of microRNA and microRNA-regulated messenger RNA expression in germinal center B-cell lymphomas determined by integrative sequencing analysis. Haematologica 2016, 101, 1380–1389. [Google Scholar] [CrossRef] [PubMed]
- Krijgsman, O.; Gonzalez, P.; Ponz, O.B.; Roemer, M.G.; Slot, S.; Broeks, A.; Braaf, L.; Kerkhoven, R.M.; Bot, F.; van Groningen, K.; et al. Dissecting the gray zone between follicular lymphoma and marginal zone lymphoma using morphological and genetic features. Haematologica 2013, 98, 1921–1929. [Google Scholar] [CrossRef]
- Bates, M.; Spillane, C.D.; Gallagher, M.F.; McCann, A.; Martin, C.; Blackshields, G.; Keegan, H.; Gubbins, L.; Brooks, R.; Brooks, D.; et al. The role of the MAD2-TLR4-MyD88 axis in paclitaxel resistance in ovarian cancer. PLoS ONE 2020, 15, e0243715. [Google Scholar] [CrossRef]
- Stemmler, M.P. Cadherins in development and cancer. Mol. Biosyst. 2008, 4, 835–850. [Google Scholar] [CrossRef]
- Vieira, A.F.; Paredes, J. P-cadherin and the journey to cancer metastasis. Mol. Cancer 2015, 14, 178. [Google Scholar] [CrossRef]
- Yang, H.; Green, M.R. Harnessing lymphoma epigenetics to improve therapies. Hematology Am. Soc. Hematol. Educ. Program. 2020, 2020, 95–100. [Google Scholar] [CrossRef]
- Thermo Fisher Scientific. Axiom Genotyping Solution—Data Analysis Guide; Technical Report; Thermo Fisher Scientific: Waltham, MA, USA, 2011. [Google Scholar]
- GENEVA Coordinating Center. GWASTools Data Cleaning; Technical Report; GENEVA Coordinating Center: Genève, Switzerland, 2020. [Google Scholar]
- Wiggs, J. Quality Control Report for Genotypic Data; Technical Report; NIH: Bethesda, MD, USA, 2016. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | ||
|---|---|---|
| No. | 185 | |
| Age (mean (SD)) | 59.17 (13.56) | |
| Sex | Male | 93 (50.3) |
| Female | 92 (49.7) | |
| Disease stage (%) | I | 20 (10.8) |
| II | 64 (34.6) | |
| III | 39 (21.1) | |
| IV | 62 (33.5) | |
| R-IPI (%) | Very good (0) | 26 (14.1) |
| Good (1–2) | 105 (56.8) | |
| Poor (3–5) | 54 (29.2) | |
| “B” symptoms (%) | Yes | 41 (22.2) |
| No | 139 (75.1) | |
| Missing | 5 (2.7) | |
| Bulky disease (%) | Yes | 58 (31.4) |
| No | 122 (65.9) | |
| Missing | 5 (2.7) | |
| Bone marrow involvement (%) | Yes | 23 (12.4) |
| No | 140 (75.7) | |
| Missing | 22 (11.9) | |
| Performance status (ECOG) (%) | 0 | 115 (62.2) |
| 1 | 58 (31.4) | |
| 2 | 12 (6.5) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perrone, G.; Rigacci, L.; Urru, S.; Kovalchuk, S.; Brugia, M.; Fabbri, A.; Iovino, L.; Puccini, B.; Cencini, E.; Orciuolo, E.; et al. Exploratory Genome-Wide Association Analysis to Identify Pharmacogenetic Determinants of Response to R-CHOP in Diffuse Large B-Cell Lymphoma. Cancers 2023, 15, 2753. https://doi.org/10.3390/cancers15102753
Perrone G, Rigacci L, Urru S, Kovalchuk S, Brugia M, Fabbri A, Iovino L, Puccini B, Cencini E, Orciuolo E, et al. Exploratory Genome-Wide Association Analysis to Identify Pharmacogenetic Determinants of Response to R-CHOP in Diffuse Large B-Cell Lymphoma. Cancers. 2023; 15(10):2753. https://doi.org/10.3390/cancers15102753
Chicago/Turabian StylePerrone, Gabriele, Luigi Rigacci, Sara Urru, Sofya Kovalchuk, Marco Brugia, Alberto Fabbri, Lorenzo Iovino, Benedetta Puccini, Emanuele Cencini, Enrico Orciuolo, and et al. 2023. "Exploratory Genome-Wide Association Analysis to Identify Pharmacogenetic Determinants of Response to R-CHOP in Diffuse Large B-Cell Lymphoma" Cancers 15, no. 10: 2753. https://doi.org/10.3390/cancers15102753