
Effect of Agronomic Techniques on Aroma Composition of White Grapevines: A Review


Department of Agriculture, Food, Environment and Forestry (DAGRI), University of Florence, 50019 Sesto Fiorentino (FI), Italy
*
Author to whom correspondence should be addressed.
Agronomy 2021, 11(10), 2027; https://doi.org/10.3390/agronomy11102027
Submission received: 21 August 2021
/
Revised: 28 September 2021
/
Accepted: 4 October 2021
/
Published: 9 October 2021
Abstract
:Climate change with rising temperatures and the unpredictability of rainy events during ripening leads to tough challenges for the winemakers in preserving the quality of white grape varieties. Grape quality is a complex concept that mainly refers to berry chemical composition, including secondary metabolites such as aroma compounds that in white berries play a key role in the identity of the wine. Terpenes, thiols, C13-norisoprenoids, methoxypyrazines, and nonterpenic alcohols are the most important aroma compounds in white grapes and several of them can be found as free volatiles or bound as glycoconjugate molecules. Agronomic practices in vineyards, such as biostimulant application, irrigation, defoliation, training systems, foliar fertilization, and bunch thinning, can have a positive effect on their concentration. This review aims to highlight the aromatic characteristics of the most utilized white grape cultivars and focus on agronomic techniques in the vineyard to enhance, implement, and intensify their aromatic characteristics. The purpose of this review paper is therefore to investigate the state of the art regarding the exaltation of aroma in white grapes in the scientific literature through sustainable agronomic techniques, but further future investigations are considered necessary given the uncertainty concerning some mechanisms of biosynthesis and their correlation with such agronomic practices.
1. Introduction
The aroma is one of the most important factors in determining wine quality, identity, and character [1]. Volatile organic compounds are essential to wine quality, determining their aroma and varietal characteristics; the type and quantity of compounds depend on the cultivar, cultivation methods, weather, the situation and soil of the vineyard, state of plant health, and wine-making practices [2]. Many authors studied the influence of grape aroma on wine quality [3,4,5,6,7,8,9,10,11,12].
Wine’s aroma consists of one thousand compounds [13] and its diversity ranges in concentration from a few ng L−1 to several mg L−1 [14]. Wine flavor can be subdivided in four classes: varietal aroma (typical of white or red grape variety), pre-fermentative aroma (originated during the initial pressed grapes evolution and processing), fermentative aroma (generated by yeast and bacteria during alcoholic and malolactic fermentations), and post-fermentative aroma (developed by transformations that occurred during aging in wood and the wine bottle, and conservation) [15].
Varietal aroma compounds are the greatest contributors to the overall aroma [16]. Varietal and pre-fermentative aromas are substantially influenced by the grape cultivar, berry maturity, microclimatic conditions, viticultural techniques, and soil management [17].
Nevertheless, throughout the years, the conditions for berry ripening are not the same and hence the grapes’ quality is different; in fact, deficiencies are usually expressed in unfavorable final aroma compounds and unappealing color and flavor [18].
In the climate change context, temperatures will presumably rise between 0.3 °C and 1.7 °C during the next 15 years [19]. Weather conditions throughout the seasons have a greater influence on grapevine development and berry composition compared to the past. High temperatures influence the enzymatic activity of primary or secondary metabolism and gene expression, affecting berry ripening and wine flavor characteristics [20]. Currently, temperatures are already increasing and the grapevine physiology is modifying [21]. These changes exert a profound shift in organic acid and sugar balance (primary metabolism) and in aromatic and phenolic berry compounds (secondary metabolism), and, finally, in the resulting wine composition [22,23].
It is in this context that proper agronomic management together with the introduction of innovative and sustainable techniques assume great importance [24,25,26]. The present review paper examines multiple agronomic and canopy management techniques aimed at improving the aromatic quality of some of the most important aromatic white grape varieties grown all over the world. This work also calls for a greater involvement of researchers in biochemical investigations essential to better understand the various mechanisms of synthesis of aromatic compounds in the perspective of climate change.
2. White Grape Aroma Compounds
White wine is a complex mixture where the flavor and aroma compounds have multiple origins [2]. It is possible to divide the aromatic compounds into the following classes: thiols, terpenes, methoxypyrazines, norisoprenoids, C6 alcohols, and aldehydes [27].
- Volatile thiols (or mercaptans) are sulfur compounds that are found in grapes in a bound form; they originate from fatty acids and are normally bound with glutathione or cysteine. Thiols are released during wine fermentation, by the enzyme carbon-sulfur lyase (C-S), from certain yeast strains [28,29]. However, the quantification of their natural precursors in must is important in the determination of the grapes’ aromatic potential. Synthesis of 4-mercapto-4-methylpentan-2-one precursors, S-4-(4-methylpentan-2-one)-L-cysteine and S-4-(4-methylpentan-2-one)-glutathione, was reported as natural and deuterated compounds; accurate quantification of such aroma precursors at trace levels in grapes was achieved by the SIDA (stable isotope dilution assay) that involves labeled analogs. Therefore, the authors demonstrated the affiliation between certain thiols precursors and thiols release; they synthesized deuterated precursors and looked at their corresponding deuterated thiols in the wines. [30].
- Terpenes (or isoprenoids) are present in the grape berries in free and bound (glycosylated) forms [31]. They are frequently conjugated to arabinose, glucose, apiose, and rhamnose [32]. During winemaking, bound terpenes can be released by glycosidase enzymes produced by grapes, bacteria, and yeast, thereby increasing the volatile terpenoid composition. Several wine yeast strains were described as significant producers of extracellular enzymes that are characterized by predominantly β-glucosidase activity [33]. Terpenes are natural hydrocarbon products constructed from connected isoprene units (isopentyl diphosphate—IPP) each containing five carbons. They derive from a rearrangement that involves carbocation intermediates, of acyclic precursors by terpene synthase/cyclase enzymes. Diterpenes (20 carbons) are synthesized from geranylgeranyl diphosphate (GGPP), while sesquiterpenes (15 carbons) from farnesyl diphosphate (FPP), and monoterpenes (10 carbons) from geranyl diphosphate (GPP) [34]. Briefly, the first pathway for the biosynthesis of terpenes is the mevalonic acid pathway (MVA) that starts with the acetyl-CoA located in the cytosol. The second is the methylerythritol phosphate pathway (MEP) that starts in plastids with the pyruvate and gliceraldeide-3-phosphate [35,36].
- Methoxypyrazines (MPs) such as 2-methoxy-3-isobutylpyrazine (IBMP) and 2-isopropyl-3-methoxypyrazine (IPMP) are volatile, grape-derived aroma compounds that give herbaceous characters to the wine [37]. Methoxypyrazines are formed by the catabolism of secondary amino acids, such as valine, glutamate, and methionine present in the grape [38]. However, the full pathway leading to the production of MPs has not yet been clarified, and there is evidence that the final step implicates the methylation of nonvolatile hydroxypyrazine (HP) precursors [39].
- Norisoprenoids originate from the carotenoid biodegradation; firstly, there is the enzymatic conversion to the aroma precursor, and finally the acid-catalyzed conversion to the aroma-active compounds [40]. The most important norisoprenoids for wine flavor are actinidol, β-ionone, β-damascenone, vitispirane, 1,1,6-trimethyl-1,2-dihydronaphthalene (TDN), 4-(2,3,6-trimethylphenyl)buta-1,3-diene (TPB), and 2,2,6-trimethylcyclohexanone (TCH) [41]. Leaves seem to be an important source of reserve for glycosylated C13-norisopreinods, but no evidence has been found that there is a translocation from the leaf to the berry [42].
3. Cultivars
The aromatic element of white wine is a crucial component of its sensory quality; such quality is focused on the perception of aromatic nuances of varying intensity and complexity that can contribute to the recognition of a typical or sensory identity (typicality) [45].
The primary consequence of global warming and increase in average temperature during grape harvest is that the herbaceous vegetal notes of white wines are limited. For instance, 2-methoxy-3-isobutylpyrazine (IBMP) levels in white wines are lower when the temperature is excessively high during the growing season [46,47]. Instead, some C13-norisoprenoid derivatives are higher whenever grapes are more exposed to light. In particular, TDN (1,1,6-trimethyl-1,2-dihydronaphtalene; notes of kerosene), a compound found in Riesling wines and synthesized during aging from nonvolatile precursors, is higher in grapes ripened in warmer microclimates [48]. In contrast, ß-damascenone concentrations, which contribute to fruity notes in white wines, are lower in conditions of increased exposure to light and higher temperatures [49].
All this makes the cultivation and management of white grapes very delicate. It is therefore necessary that the winemaker, following climate change, implement a series of strategies aimed at preserving and enhancing the aromatic potential of white grape varieties to optimize the harvest.
3.1. Sauvignon Blanc
Vitis vinifera L. cv. Sauvignon blanc is one of the traditional varieties of the Sancerre and Sauternes regions (France) and is also planted extensively in several other wine-producing regions of the world such as New Zealand and South Africa [50]. Wines made from this cultivar have a distinctive aroma [51]. Many authors describe the typical aroma of Sauvignon blanc wine as herbaceous, tomato leaf, vegetative, grassy, gooseberry, grapefruit, green pepper, asparagus, capsicum, and passion fruit [52,53,54]. These wines tend to be acidic or crisp to the tasting on the palate and are subdivided into two groups, “tropical” (grapefruit, gooseberry, and passion fruit) and “green” (tomato leaf, vegetative, green pepper, grassy, herbaceous, asparagus, and capsicum) [55]. In Sauvignon blanc wines, some volatile sulfur compounds, called volatile thiols, can contribute to positive fragrances such as passion fruit, tropical, and guava-like scents, [56]. The volatile thiols are thiol compounds with additional functional groups such as alcohols, ketones, and esters [57].
As shown in Table 1, the most important volatile thiols are 4-mercapto-4-methylpentan-2-one (4MMP), 4-mercapto-4-methylpentan-2-ol (4MMPOH), 3-mercaptohexan-1-ol (3MH), and 3-mercapto-hexylacetate (3MHA) [58].
The free thiols and their acetate are practically absent from musts, while they are liberated by the yeasts during fermentation from cysteine conjugate and glutathione conjugate precursors. In fact, 4MMP and 3MH are released during fermentation from glutathionylated precursors [S-3-(hexan-1-ol)-glutathione (Glut-3MH) and S-4-(4-methylpentan-2-one)-glutathione (Glut-4MMP)] and cysteinylated precursors [S-3-(hexan-1-ol)-l-cysteine (Cys-3MH) and S-4-(4-methylpentan-2-one)-l-cysteine (Cys-4MMP)] [63].
For example, S-3-(hexan-1-ol)-l-cysteine (Cys-3MH), S-3-(hexan-1-ol)-glutathione (G3MH), 3-S-glutathionylhexan-1-ol (GSH-3MH), and (E)-2-Hexenal are considered direct precursors of 3MH [64].
Further volatile compounds are important to the flavors of Sauvignon blanc wines, such as the methoxypyrazines and certain C6 alcohols cis-hexen-1-ol and trans-hexen-1-ol, which can impart grassy, capsicum, and leafy aromas to the wines [65,66,67]. Three methoxypyrazine compounds have been identified in Sauvignon blanc, namely 2-isobutyl-3-methoxypyrazine (IBMP, bell peppers), 2-sec-butyl-3-methoxypyrazine (SBMP, vegetal flavor), and 2-isopropyl-3-methoxypyrazine (IPMP, herbaceous flavor) [68].
3.2. Muscat
Muscat is an aromatic variety containing a specific amount of terpene compounds [69]. Muscat wines are distinguished by typical floral aromas that mostly originate from the berries. Monoterpenols, such as nerol, linalool, geraniol, citronellol, and α-terpineol, contribute greatly to the characteristic flavor of the Muscat grape varieties [70,71]. The compounds responsible for varietal aroma are predominantly in the skins [72]; fifteen monoterpenes were actually identified in the berries’ skin, including monoterpene alcohols, aldehydes, and oxides [73]. Gas chromatography–mass spectrometry (GC–MS) was used to identify 32-free and 23-bound compounds in Muscat of Bornova grapes: terpenes were the principal compounds in free and bound fractions; linalool and trans-linalool oxide pyran were the principal compounds in the free fraction. Nerol, geranic acid, and geraniol in the bound fraction were the most abundant terpenes in this type of must [74].
Aroma compounds in the grape are both in the free and glycosylated forms; however, only the free forms contribute directly to the aroma profile of these wines [75]. In addition, volatile compounds formed during alcoholic fermentation also contribute to the aroma profile of wines, and these include short-chain fatty acids (C6, C8, and C10) and their ethyl esters, higher alcohols, and their acetate esters [76]. During the alcoholic fermentation, the most relevant volatile compounds are acetaldehyde, hexanoic acid, 3-methyl-1-butanol, 2-methyl-1-butanol, 2-methyl-1-propanol, 1-propanol, methanol, isobutyric acid, ethyl octanoate, ethyl acetate, isoamyl acetate, ethyl 4-hydroxybutyrate, diethyl succinate, and octanoic acid [69,77].
Another class of volatile compounds, C6 compounds, are derived from the lipoxygenase (LOX) pathway, and they are important for herbaceous characters [78].
3.3. Gewürztraminer
Gewürztraminer is an aromatic grape cultivar, widely grown in cooler wine-producing European regions such as Germany and northern France [79].
Maximizing the development of spicy and floral aromatic characteristics during maturation and ripening is an important objective for vintners. Terpenic components are the main contributors to the typical flavor of this cultivar; they are synthesized in the grapes, extracted into the must during crushing and pressing, and transferred to wine during fermentation and stabilization phases [80]. Terpenes are present as free volatiles and as sugar-bound precursors [81]. Monoterpenes (C10) and sesqui- (C15) are formed in plastids, via the cytosolic mevalonate-dependent metabolic pathway (MVA) and the mevalonate-independent 1-deoxy-d-xylulose 5-phosphate/2-C-methyl-d-erythritol 4-phosphate biosynthesis pathway (DOXP/MEP) [82,83]. The presence of monoterpene glycosides by hydrolytic cleavage of the glycosidic bond was confirmed, identifying both acyclic monoterpene alcohols, such as linalool, nerol, and geraniol, and monocyclic terpenes such as α-terpineol as released aglycones [84]. Twenty-four terpene aglycones and twenty-one monoterpene alcohols were found in hydrolysates of Gewürztraminer berry’s exocarp. More than 80% of all terpene aglycones detected were attributed to six monoterpenols: linalool, nerol, α-terpineol, geraniol, dihydrocarveol, and β-citronellol [82].
3.4. Riesling
Several white varieties are distinguished by their terpenes level:
- -
- Muscat/floral cultivars: high free monoterpene content;
- -
- Non-Muscat aromatic cultivars—medium-free monoterpenes content;
- -
- Neutral cultivars, where monoterpenes do not appear to influence wine aroma [85].
Riesling can be placed in the intermediate level between neutral cultivars and Muscat [86].
The major compounds responsible for Riesling wine varietal character are monoterpenes (such as linalool; floral aroma) and carbon-13 norisoprenoids (especially 1,1,6-trimethyl-1,2-dihydronaphthalene; kerosene aroma) [87]. It is worth noting that in this cv. a large number of volatile phenols exist as glycosides [88].
The aroma compounds found in Riesling grapes are carbon-13 (C13) norisoprenoids (β-damascenone, 1,1,6-trimethyl-1,2-dihydronaphthalene—TDN, and vitispirane), monoterpenes (α-terpineol, linalool oxide), and phenolics (eugenol, 4-vinylguaiacol, and vanillin) [89].
3.5. Chardonnay
Chardonnay is one of the most important white cultivars grown in all parts of the world; it is the most widely planted white cultivar in both Australia and California, and the second most planted in France [90]. The aroma models of Californian Chardonnay wines were evaluated using multivariate statistical procedures to investigate the sensory significance of odor-active compounds after chromatography/olfactometry; partial least squares regression (PLSR) analysis was used to find the relevant combinations of odor-active compounds. Eighty-one aroma compounds were found as odor active in nineteen Californian Chardonnay wines; PLSR revealed that the ”fruity” terms were related to 2-phenylethyl acetate, linalool, and 3-methylbutyl acetate [91,92].
Glycosidic precursors are a diverse group of odorless compounds identified in many grapes and reputed to be the most important group responsible for some of the varietal attributes of wines, such as Chardonnay [93].
Although Chardonnay is an international aromatic cultivar, in Burgundy-specific clones of Chardonnay are used to produce wines differing by their aromatic profile; clone 76 is considered as neutral and clone 809 as aromatic [94].
In Chardonnay grapes, there are several differences based on terroir. In Spanish wines, 3-mercaptohexan-1-ol can be considered unimportant to Chardonnay and 4-mercapto-4-methylpentan-2-one and 3-mercaptohexyl acetate were correlated to the fruity aroma, box hedge, and tropical fruit character [59]. In contrast, in Australian Chardonnay wines, both 4-mercapto-4-methylpentan-2-one and 3-mercaptohexyl acetate were below the sensory threshold, while 3-mercaptohexan-1-ol was generally well above [95]. However, for Spanish and Australian wines, benzyl mercaptan (BM) was present at a sensorily important level [59,95]. BM is considered to be a contributor to smoky, struck match or struck flint characters in white wine, and in a sensory addition study [96], an addition even as low as 0.7 ng/L gave a burnt, toasty, and empyreumatic aroma.
3.6. Ribolla Gialla
The Vitis vinifera L. cv. Ribolla Gialla (famous for its robust berries and plain sensory profile) is an autochthonous white grape variety cultivated in northeastern Italy, parts of Croatia, and southwest Slovenia [97].
Aroma compounds found in the Ribolla Gialla cultivar were 2-methyl propanol, ethyl acetate, 3-methyl butanol, 2-phenyl ethanol, 2-phenylethyl acetate, ethyl octanoate, ethyl butanoate, ethyl hexanoate, ethyl decanoate, and isoamyl acetate [98].
3.7. Malvasia
Istrian Malvasia is a grapevine variety from the north Adriatic area, mainly grown in the regions of Istria (Croatia), Friuli (Italy), and Primorska (Slovenia) [99]. Aroma compounds found in the Istrian Malvasia wines were 2-phenyl ethanol, 3-methyl butanol, ethyl octanoate, ethyl acetate, ethyl butanoate, ethyl hexanoate, ethyl decanoate, linalool, isoamyl acetate, and α-terpineol [98].
3.8. Pinot Gris
The major compounds responsible for Pinot gris varietal character include monoterpenes. All of the monoterpene isomers: S-(−)-limonene, R-(+)-limonene, (2R,4S)-(+)-cis-rose oxide, (2S,4R)-(−)-cis-rose oxide, (2R,4R)-(−)-trans-rose oxide, (2S,4S)-(+)-trans-rose oxide, (2R,5R)-(+)-trans-linalool oxide, (2R,5S)-(−)-cis-linalool oxide, (2S,5S)-(−)-trans-linalool oxide, (2S,5R)-(+)-cis-linalool oxide, R-(−)-linalool, S-(+)-linalool, S-(−)-α-terpineol, R-(+)-α-terpineol, and R-(+)-β-citronellol were identified and quantified in Pinot Gris wines by using mass spectrometry (HS-SPME−MDGC−MS) [100].
4. Agronomic Practices
Agronomic practices such as irrigation, defoliation, foliar fertilization, canopy-training systems, spraying of biostimulants, and bunch thinning can have a great impact on primary and secondary metabolism [101,102]. Under different environmental conditions, choosing the most effective multidisciplinary approach such as agricultural practices, cultivar, and rootstock can optimize the plant’s performance during their entire life cycle and the relationships between phytochemicals compounds (such as glycine betaine, melatonin, proline, polyamines, γ-aminobutyric acid, organic acids, sugars, amino acids, aromas, and phenolic compounds) [103].
4.1. Irrigation
Climate change has a significant impact on rainfall cycles, modifying the vegetative season with the occurrence of droughts or flooding that in turn influence agricultural yields and crop quality in all geographic areas of the world [104,105]. Irrigation is needed to stabilize yield while maintaining white grapes’ quality [106]. The intensity and timing of water deficits or irrigations influence the extent of alterations occurring in berry metabolism and therefore in wine aroma and flavor [107,108].
Des Gachons [109] showed that grape aroma potential was highest in Sauvignon blanc vines under mild water deficit (predawn leaf water potential ΨPD −0.22 MPa) and moderate nitrogen supply, while severe water deficit stress seemed to limit S-Cysteine conjugate precursors. Other scientific works also demonstrated the benefits of irrigation (0 < ΨPD < −0.45 MPa) on precursor thiols (3-S-cysteinylhexan-1-ol, Cys-3MH, and 3-S-glutathionylhexan-1-ol, GSH-3MH) [110,111]. The aroma of Sauvignon blanc at three different locations in Italy (Veneto, Tuscany, and Apulia) under irrigation was monitored. The irrigation treatment improved the quality of must (linalool, α-terpineol, monoterpenes, methoxypyrazine, and norisoprenoids contents) [112].
In northwest Italy, the effects of irrigation on the volatile aroma profile of Muscat blanc grapes was also investigated; free linalool and geraniol concentrations were higher in the standard irrigated plants (pre-dawn water potential levels above −0.2 MPa) than in the drought regime, with 78% more free linalool and 73% more free geraniol than the plants subjected to drought [113].
In Gewürztraminer grapevines, decreasing duration of water stress increased free volatile terpenes (FVT) at harvest, and potentially volatile terpenes (PVT) [114]. Regulated deficit irrigation (RDI), a viticultural practice utilized to improve the concentration of secondary metabolites [115,116], applied after veraison, increased the concentration at harvest of key free terpenes for Gewürztraminer grapes, such as geraniol and citronellol [117]. Water deficit can therefore increase the concentration of terpenes in grapes, via the upregulation of genes of the methylerythritol phosphate pathway (MEP), such as hydroxy-3-methylbut-2-enyl diphosphate reductase and terpene synthases [118].
These findings encourage research and development of climate change adaptation measures, namely the implementation of balance irrigation systems.
4.2. Biostimulant Application
Biostimulants foliar application on vines has become an interesting strategy to improve grape quality and prevent plant diseases on the grapevine (including resistance inducers or elicitors) [119,120]. These products include a wide variety of formulations, such as protein hydrolysates, fulvic and humic acids (or other N-containing compounds), seaweed extracts and botanicals, inorganic compounds, biotic and abiotic elicitors, and beneficial bacteria and fungi, which are able to trigger an innate immune response, leading to the synthesis of secondary metabolites in vines [121,122].
Recent work showed that a low-dose nitrogen biostimulant (foliar phenylalanine application) can improve the synthesis of esters and benzenoids, and decrease the concentrations of terpenoids and C6 compounds; however, its effects on C13 norisoprenoids in must were not clear. The highest dose of phenylalanine increased the content of all except for the C6 compounds [123].
Although the effects of methyl jasmonate on grape amino acid contents are not completely known [124], some data suggest that methyl jasmonate (elicitor biostimulant) promotes activation of terpene metabolism by increasing geranylgeranyl diphosphate synthase, the enzyme that provides the building blocks for terpenes [125].
The effects of foliar oak wood extract applications to Chardonnay grapevines on berry glycosidic aroma precursors were studied [92]; after the foliar applications, alcohols, C6 compounds, phenols, terpenes, and C13-norisoprenoids showed an increase. Moreover, the majority of the compounds coming from the oak extracts were assimilated and stored, in their glycosidic forms, in the berries.
In another study, the response of Riesling grape variety by spraying three different concentrations of humic acid extracted from vermicompost (30, 40, and 50 mL∙L−1) onto leaves was investigated. As a result, it was observed that there was a general increase in fruit quality and aroma in vines [126].
4.3. Foliar or Soil Fertilization
Vine nutrition is an important factor for wine quality and plays an essential role in the winery [127]; a nutritional deficiency of the vine manifests itself for multiple reasons and negatively affects the aromatic profile of a wine, for example, due to slow or blocked fermentation [128]. Foliar treatments are applied mostly in the case when small fertilization corrections are necessary and they are preferred to soil applications when the topsoil is very dry or with decreased root activity [129].
A grapevine’s nutrition can be affected by various factors, as reported in Figure 1 [130,131,132,133,134,135].
In particular, grapevine nitrogen status can affect vine vigor and grapes; nitrogen (N) fertilization can increase the berry’s N content [129,136]. There is growing interest in the analysis of individual amino acids in must owing to their important role such as precursors to aromas released during fermentation [137]. A relationship between wine aroma composition and must amino acids has been described [138,139]. In fact, isobutyl, isoamyl, and phenylethyl alcohols are derived, respectively, from isoleucine, leucine, and valine; in addition, phenylalanine, threonine, and aspartic acid are the amino acids that most influence the fermentation process [138].
A recent research studying the effect of irrigation and N fertilization, showed that combined soil and foliar nitrogen application on Chardonnay vines, achieved greater increases in amino acid and total concentration of glycosylated precursors [140].
Choné et al. (2006) showed that in Sauvignon blanc, an unlimited nitrogen supply to the vine is a primary factor for a balanced varietal aroma expression; vine nitrogen status increased S-cysteine conjugate precursors, glutathione, and phenolic content in the berries [141]. Moreover, another study showed that vine nitrogen fertilization influenced the Sauvignon blanc vine physiology and the composition of the grapes, enhancing their aroma expression (higher content of nonvolatile and volatile thiols) [142].
Urea applied foliarly to Sauvignon blanc grapevines increased the concentration of amino acids in musts involved in the biosynthesis of precursor thiols [143].
On the other hand, another work investigated the effectiveness of nontoxic calcium phosphate nanoparticles (Ca3(PO4)2∙nH2O) doped with urea (U-ACP) as a nitrogen source for grapevine fertilization, and found that the volatile profile of Pinot gris berries (alkyl- and benzyl- alcohols, aldehydes, ketones, and carboxylic acids) was not significantly affected by the treatments. The following tree fertilization treatments were compared: commercial granular fertilization (45 kg N ha−1), U-ACP applied in fertigation (36 kg N ha−1), and foliar application of U-ACP (36 kg N ha−1) [144].
4.4. Cluster Thinning
The proper balance of reproductive and vegetative growth is fundamental for optimal fruit ripening. In a vineyard, a higher yield per vine may delay ripening, which might result in lower quality of the berries [145]. For this reason, cluster thinning, which promptly removes the reproductive sink, is a standard agronomic practice to prevent overcropping and enhance grape quality used in viticulture [146]; therefore, it can be concluded that canopy management techniques have favorable effects on grapevine growth and grape composition [147].
Several studies showed that cluster thinning helped to improve the terpene concentrations; linalool, the most abundant and active odorant related to Muscat flavor, accumulated in 28.6% and 20.2% higher concentrations from cluster thinning compared to control and girdling at maturity, respectively [148]. In fact, in most cases, change in terpene concentration of berries through cluster thinning was associated with favorable modifications in flavor or aroma characteristics [149]. Another study examined the accumulation of monoterpenes during fruit maturation on a specific group of Chardonnay clones called Chardonnay Musqué, after five different phenological stages of cluster thinning; the results showed that thinning treatments increased monoterpene concentration of Chardonnay Musqué berries [150].
The effects of cluster thinning time at different stages (pre-veraison and post-veraison) of berry development on grape quality and yield components of cv. Sauvignon blanc grapevines were investigated; cluster thinning at various stages of berry development increased monoterpenes [151]. Other research showed that herbaceous aroma scores had a positive correlation with increased pruning weight (cv. Sauvignon blanc) [152].
The effect of 20% cluster thinning on Ribolla Gialla sparkling wine aroma, on lipid compounds, and aromatic amino acid metabolites composition was studied for three seasons in the Friuli Venezia Giulia region of Italy. The thinning of the grape clusters showed a minimal positive effect on the volatile composition, resulting in higher concentration of varietal aroma compounds (citronellol, linalool, and β-myrcene). A similar result was obtained for the C13-norisoprenoid group of substances. In addition, the effect of cluster thinning produced the increase in metabolites associated with aromatic amino acids [153].
4.5. Leaf Removal
A balance between the photosynthetically active leaves and production is an important factor to obtain wines of higher quality [154]. The intensity and composition of sunlight are two of the most important parameters that influence the microclimate of bunches, together with the effects of temperature [155]. In vineyards, leaf removal at the level of the clusters changes the microclimate in the area around the grapes and therefore also berries’ composition [156]. Defoliation directly affects the leaf/fruit ratio, which, depending on the phenological stage in which it is conducted, can produce a different effect on the maturation of the fruit [157]. Many authors examined the influence of light intensity and/or composition of the bunch zone on aroma molecules, focusing on the main aroma molecules: free aroma volatiles, glycosylated aroma precursors, C13-norisoprenoids, methoxypyrazines, and terpenes [158,159].
The technique of leaf removal (removal of five basal leaves from each shoot at veraison) showed the greatest effects on must and wine composition in cv. Riesling italico and Traminer; increasing light exposure increased the flavonoid content of grapes. Nontreated grapes of all cultivars had, for the most part, the lowest levels of hydroxybenzoic acid, catechin, epicatechin, and flavor [160]. Sun-exposed bunches (cv. Riesling) showed an increase in C13-norisoprenoid concentration in berries and wine. β-damascenone concentration did not change between nontreated and defoliated plants, but an increase of 1,1,6-trimethyl-1,2-dihydronaphthalene (TDN) was observed in the defoliated vines [161]. Other research on Riesling showed that bound and free aroma compounds such as linalool, geraniol, hexen-1-ol, α-terpineol, and β-damascenone were positively affected by higher sunlight exposure of berries [89].
In the Isonzo region (Friuli, Italy), the influence of cluster-zone leaf removal, applied after berry set, was evaluated on Sauvignon blanc grapevines; the increased sunlight cluster exposure decreased 2-methoxy-3-isobutylpyrazine (IBMP) and 3-isopro-pyl-2-methoxypyrazine (IPMP) concentrations in early stages of berry development, while at harvest, although light plays a role in the degradation of berry methoxypyrazines, no significant differences between defoliated and nondefoliated grapes were observed [162].
Moreover, leaf removal conducted two weeks before veraison, at veraison, and two weeks after veraison, at two different levels (half basic, 50%; full basic, 100%) increased the concentrations of most free- and bound-form monoterpenes in Sauvignon blanc grapes; the concentrations of total (free + bound) monoterpenes in grapes were significantly higher in plants treated two weeks before veraison-50% (1226.68 μg L−1) and veraison-50% (1255.54 μg L−1) compared to the control (1160.22 μg L−1) [163].
The sun-exposed berries and the berries naturally shaded under foliage were studied in cv. Muscat. Due to the increased amounts of hexanal and E-2-hexenal, the total amounts of C6 compounds were greater in the berries of shaded bunches; bunch shading did not modify the levels of nonterpenic alcohols, but the total amounts of terpenols (nerol, linalool, geraniol and geranic acid) were lower in the shaded berries than in the sun-exposed berries [164]. The cluster temperature could explain these differences [165]: modification of the red/far-red ratio can modify the phytochrome activity that regulates the 3-hydroxy-3-methylglutaryl-CoA reductase (HMG-CoA reductase) involved in monoterpenes biosynthesis (natural shading decreased the red/far-red ratio) [166,167].
Enhancement of Istrian Malvasia wine aroma and higher hydroxycinnamate composition were obtained by hand (HLR) and mechanical leaf removal (MLR), compared with untreated control (UC) [168]; precursors of varietal thiols 3-S-glutathionylhexan-1-ol (G3SH) (HLR 9.5 ng kg−1; MLR 10.6 ng kg−1; UC 7.0 ng kg−1), 3-S-cysteinylhexan-1-ol (Cys3SH) (HLR 0.5 ng kg−1; MLR 0.6 ng kg−1; UC 0.3 ng kg−1), 4-S-glutathionyl-4-methylpentan-2-one (G4MSP) (HLR 16.4 ng kg−1; MLR 17.6 ng kg−1; UC 11.8 ng kg−1), 4-S-cysteinyl-4-methylpentan-2-one (Cys4MSP) (HLR 28.5 ng kg−1; MLR 27 ng kg−1; UC 22.4 ng kg−1) in berries and hydroxycinnamic acids and their esters in wine were higher in defoliated-treated vines [169].
4.6. Shading Nets
Shading net applications over the canopy reduce the photosynthetic photon flux on the leaf surface, and the temperature both of the canopy [170] and the fruit by up to 7 °C [171], and thus, could delay berry ripening. It is demonstrated that sun exposure influences the aromatic composition of grapes [172]. Excessive exposure to sunlight and high berry temperatures reduce the content of methoxpyrazines [173]. However, the glycosidically bound monoterpenes and polyols (potentially volatile terpenes) in Gewürztraminer grapes [165] and the norisoprenoid concentrations in Riesling [161] grapes were maintained at the highest levels in exposed berries during ripening and were considerably higher than in the partially and completely shaded fruit at harvest.
In Sauvignon blanc, partial shading of the canopy (50%) increased iso-butyl methoxypyrazine concentration compared with full sun exposure (4.57 and 1.94 ng/L, respectively) [174].
Different treatments, on Chardonnay vines, using shading nets, were evaluated vis-à-vis an untreated test with no defoliation and shading. Five different treatments were examined: a comparative test without defoliation and shading (ND), a test with total defoliation (east and west side) (TD), and three different systems adopting shading nets applied along the bunch zone. Two of the shaded treatments were defoliated as for TD and covered with one layer of shading net (TD1L) or two layers of shading net (TD2L), while a third treatment was covered by only one layer of shading net but not defoliated (ND1L). Significant results were only seen for Quercetin-3-glucoside, Kaempferol-3-glucuronide, Isorhamnetin-3-glucoside, and Kampferol-3-Glucoside, for which the highest values were recorded for the TD-FD treatment compared to ND1L-FD. Shaded treatments (TD1L, TD2L, and ND1L) led to the highest values in terms of cinnamic acid concentration, while ND1L led to a lower level of flavonols. No significant differences were observed for stilbenes, while a higher level of total thiols was shown for the TD2L treatment compared to ND and TD1L. This was determined by differences identified for S-glutathionylated precursor of 3-Sulfanylhexan-1-ol (GSH-3MH) [175].
The concentrations of free and bound terpenol in shaded Chilean Muscat berries, however, were so low that the characteristic Muscat aroma was lost, which resulted in poor-quality musts [176].
Leaf shading can be a useful strategy to deal with global warming in viticulture [177,178], as it could slow the ripening process and favor the accumulation of herbaceous compounds in grapes;, nevertheless, it is a practice to be applied with caution.
Generally, the treatment tested was able to moderate the effects of excessive radiation and temperature on fruit, without affecting the physiology, and enhancing fruit composition. This result will be particularly relevant in areas in which the effects of climate change increase mean temperatures during the growing season.
4.7. Kaolin
The kaolin sunscreen works by reducing leaf temperature by an average of about 4 °C and up to 6 °C while maintaining a high photosynthetic activity, thus avoiding photoinhibition phenomena and physiological damage such as chlorotic/necrotic leaves, sunburn damage, and dehydrated berries [179]. It was shown that kaolin applications to grapevines significantly reduced berry surface temperatures by 0.7 and 1.5 °C without negatively affecting berry physicochemical parameters at harvest [170]. Kaolin application on overexposed clusters reduces sunburn and influences the grape composition of Sauvignon blanc with the increase of bound secondary grape metabolites [171]. These increases represent, in part, an increase in the pool of potential aroma and flavor components, and may indicate an enhanced potential for high-quality wines [180]. Recent studies have shown different results on kaolin’s impacts on grapevine aroma. The accumulation of volatile phenol glycoconjugates in smoke-exposed grapes (smoke taint can be found in wines made from grapes exposed to smoke from bushfires after a drought) was monitored following grapevine exposure to smoke, with different glycoconjugate profiles observed. Foliar application of kaolin reduced the concentration of volatile phenol glycoconjugates in smoke-exposed fruit, but efficacy depended on the rate of application and extent of coverage (Sauvignon blanc and Chardonnay) [181]. In view of the results, the exogenous foliar application of kaolin, an inert mineral that reflects radiation, has been shown to be an effective short-term climate change mitigation strategy for vineyards.
5. Conclusions
Due to climate change and rising temperatures, winemakers are increasingly forced to find winning strategies so as not to jeopardize the quality of white grapes in the vineyard. In a genotype-dependent way, several agronomic practices can positively influence primary and secondary metabolite concentrations in the berries. In white wines, the aroma, originating from different chemical and biochemical compounds, plays a key role in the identity of the product. It is the task of the winemaker to enhance the synthesis of these molecules through different approaches in the vineyard and in the cellar. However, currently, given the increasingly high and sophisticated demand from consumers for a healthy and high quality product, the choices to increase the aromatic quality of white wines will have to be oriented toward sustainable agronomic management practices. Agronomic techniques such as defoliation, balancing of the sink/source ratio, thinning of bunches, foliar and soil fertilization have all proved to be excellent sustainable strategies to increase the aromatic potential of white grapes. Above all, however, with a view toward climate change, the following agronomic practices are considered valid aids to not deplete aromatic compounds: balanced irrigation, shading nets, and the application of kaolin and/or biostimulants. Nevertheless, in most studies, the data were only reported in terms of metabolite concentrations with no discussion about dynamics in the berries’ growth, so physiological mechanisms, plant/agronomic technical interactions, and synthesis mechanisms still require further study and investigation. Moreover, if these viticultural strategies were to be used together, their cumulative effect could lead to a more pronounced improvement in grape ripening and aroma.
Author Contributions
E.C. wrote the original draft manuscript, which was edited and improved by G.B.M.; investigation, L.S., F.P., and M.F. All authors have read and accepted the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Rocha, S.; Coutinho, P.; Barros, A.; Coimbra, M.A.; Delgadillo, I.; Dias Cardoso, A. Aroma potential of two Bairrada white grape varieties: Maria Gomes and Bical. J. Agric. Food Chem. 2000, 48, 4802–4807. [Google Scholar] [CrossRef]
- González-Barreiro, C.; Rial-Otero, R.; Cancho-Grande, B.; Simal-Gándara, J. Wine aroma compounds in grapes: A critical review. Crit. Rev. Food Sci. Nutr. 2015, 55, 202–218. [Google Scholar] [CrossRef] [PubMed]
- Gil, J.V.; Mateo, J.J.; Jiménez, M.; Pastor, A.; Huerta, T. Aroma compounds in wine as influenced by apiculate yeasts. J. Food Sci. 1996, 61, 1247–1250. [Google Scholar] [CrossRef]
- Kovačević Ganić, K.; Staver, M.; Peršurić, Đ.; Banović, M.; Komes, D.; Gracin, L. Influence of blending on the aroma of Malvasia istriana wine. Food Technol. Biotechnol. 2003, 41, 305–314. [Google Scholar]
- Francis, I.L.; Newton, J.L. Determining wine aroma from compositional data. Aust. J. Grape Wine Res. 2005, 11, 114–126. [Google Scholar] [CrossRef]
- Koundouras, S.; Marinos, V.; Gkoulioti, A.; Kotseridis, Y.; van Leeuwen, C. Influence of vineyard location and vine water status on fruit maturation of nonirrigated cv. Agiorgitiko (Vitis vinifera L.). Effects on wine phenolic and aroma components. J. Agric. Food Chem. 2006, 54, 5077–5086. [Google Scholar] [CrossRef] [PubMed]
- Molina, A.M.; Swiegers, J.H.; Varela, C.; Pretorius, I.S.; Agosin, E. Influence of wine fermentation temperature on the synthesis of yeast-derived volatile aroma compounds. Appl. Microbiol. 2007, 77, 675–687. [Google Scholar] [CrossRef]
- Hernandez-Orte, P.; Concejero, B.; Astrain, J.; Lacau, B.; Cacho, J.; Ferreira, V. Influence of viticulture practices on grape aroma precursors and their relation with wine aroma. J. Sci. Food Agric. 2015, 95, 688–701. [Google Scholar] [CrossRef] [PubMed]
- Condurso, C.; Cincotta, F.; Tripodi, G.; Sparacio, A.; Giglio, D.M.L.; Sparla, S.; Verzera, A. Effects of cluster thinning on wine quality of Syrah cultivar (Vitis vinifera L.). Eur. Food Res. Technol. 2016, 242, 1719–1726. [Google Scholar] [CrossRef]
- Ilc, T.; Werck-Reichhart, D.; Navrot, N. Meta-analysis of the core aroma components of grape and wine aroma. Front. Plant. Sci. 2016, 7, 1472. [Google Scholar] [CrossRef] [Green Version]
- Song, C.Z.; Liu, M.Y.; Meng, J.F.; Shi, P.B.; Zhang, Z.W.; Xi, Z.M. Influence of foliage-sprayed zinc sulfate on grape quality and wine aroma characteristics of Merlot. Eur. Food Res. Technol. 2016, 242, 609–623. [Google Scholar] [CrossRef]
- Belda, I.; Ruiz, J.; Esteban-Fernández, A.; Navascués, E.; Marquina, D.; Santos, A.; Moreno-Arribas, M. Microbial contribution to wine aroma and its intended use for wine quality improvement. Molecules 2017, 22, 189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, F.; Du, B.; Li, J. Aroma compounds in wine. In Grape Wine Biotechnology; InTechOpen: London, UK, 2016; pp. 273–283. [Google Scholar] [CrossRef] [Green Version]
- Polášková, P.; Herszage, J.; Ebeler, S.E. Wine flavor: Chemistry in a glass. Chem. Soc. Rev. 2008, 37, 2478–2489. [Google Scholar] [CrossRef]
- Rubio-Bretón, P.; Salinas, M.R.; Nevares, I.; Pérez-Álvarez, E.P.; del Álamo-Sanza, M.; Marín-San Román, S.; Alonso, G.L.; Garde-Cerdán, T. Recent advances in the study of grape and wine volatile composition: Varietal, fermentative, and aging aroma compounds. Food Aroma Evol. 2019, 22, 439–463. [Google Scholar]
- Ruiz, J.; Kiene, F.; Belda, I.; Fracassetti, D.; Marquina, D.; Navascués, E.; Calderon, F.; Benito, A.; Rauhut, D.; Santos, A.; et al. Effects on varietal aromas during wine making: A review of the impact of varietal aromas on the flavor of wine. Appl. Microbiol. 2019, 103, 7425–7450. [Google Scholar] [CrossRef]
- Bueno, J.E.; Peinado, R.; Moreno, J.; Medina, M.; Moyano, L.; Zea, L. Selection of volatile aroma compounds by statistical and enological criteria for analytical differentiation of musts and wines of two grape varieties. J. Food Sci. 2003, 68, 158–163. [Google Scholar] [CrossRef]
- Boulton, R.B.; Singleton, V.L.; Bisson, L.F.; Kunkee, R.E. Principles and Practices of Winemaking; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Brovkin, V.; Sitch, S.; Von Bloh, W.; Claussen, M.; Bauer, E.; Cramer, W. Role of land cover changes for atmospheric CO2 increase and climate change during the last 150 years. Glob. Chang. Biol. 2004, 10, 1253–1266. [Google Scholar] [CrossRef] [Green Version]
- Drappier, J.; Thibon, C.; Rabot, A.; Geny-Denis, L. Relationship between wine composition and temperature: Impact on Bordeaux wine typicity in the context of global warming. Crit Rev. Food Sci Nutr. 2019, 59, 14–30. [Google Scholar] [CrossRef]
- Rienth, M.; Torregrosa, L.; Luchaire, N.; Chatbanyong, R.; Lecourieux, D.; Kelly, M.T.; Romieu, C. Day and night heat stress trigger different transcriptomic responses in green and ripening grapevine (Vitis vinifera) fruit. BMC Plant Biol. 2014, 14, 108. [Google Scholar] [CrossRef] [Green Version]
- Jordão, A.M.; Vilela, A.; Cosme, F. From sugar of grape to alcohol of wine: Sensorial impact of alcohol in wine. Beverages 2015, 1, 292–310. [Google Scholar] [CrossRef] [Green Version]
- Cataldo, E.; Salvi, L.; Sbraci, S.; Storchi, P.; Mattii, G.B. Sustainable viticulture: Effects of soil management in Vitis vinifera. Agronomy 2020, 10, 1949. [Google Scholar] [CrossRef]
- Smart, R.E.; Dick, J.K.; Gravett, I.M.; Fisher, B. M Canopy management to improve grape yield and wine quality-principles and practices. S. Afr. J. Enol. Vitic. 1990, 11, 3–17. [Google Scholar] [CrossRef] [Green Version]
- Lock, P.; Mounter, S.; Fleming, E.; Moss, J. Wineries and wine quality: The influence of location and archetype in the Hunter Valley region in Australia. Wine Econ. Policy 2019, 8, 180–190. [Google Scholar] [CrossRef]
- Bernardo, S.; Dinis, L.T.; Machado, N.; Barros, A.; Pitarch-Bielsa, M.; Malheiro, A.C.; Gomez-Cadenas, A.; Moutinho-Pereira, J. Uncovering the effects of kaolin on balancing berry phytohormones and quality attributes of Vitis vinifera grown in warm-temperate climate regions. J. Sci. Food Agric. 2021. [Google Scholar] [CrossRef]
- Alem, H.; Rigou, P.; Schneider, R.; Ojeda, H.; Torregrosa, L. Impact of agronomic practices on grape aroma composition: A review. J. Sci. Food Agric. 2019, 99, 975–985. [Google Scholar] [CrossRef] [PubMed]
- Santiago, M.; Gardner, R.C. Yeast genes required for conversion of grape precursors to varietal thiols in wine. FEMS Yeast Res. 2015, 15, fov034. [Google Scholar] [CrossRef] [Green Version]
- Capone, D.L.; Barker, A.; Williamson, P.O.; Francis, I.L. The role of potent thiols in Chardonnay wine aroma. Aust. J. Grape Wine Res. 2018, 24, 38–50. [Google Scholar] [CrossRef]
- Roland, A.; Schneider, R.; Razungles, A.; Le Guerneve, C.; Cavelier, F. Straightforward synthesis of deuterated precursors to demonstrate the biogenesis of aromatic thiols in wine. J. Agric. Food Chem. 2010, 58, 10684–10689. [Google Scholar] [CrossRef]
- Mele, M.A.; Kang, H.M.; Lee, Y.T.; Islam, M.Z. Grape terpenoids: Flavor importance, genetic regulation, and future potential. Crit. Rev. Food Sci. Nutr. 2021, 61, 1429–1447. [Google Scholar] [CrossRef]
- Voirin, S.G.; Baumes, R.L.; Sapis, J.C.; Bayonove, C.L. Analytical methods for monoterpene glycosides in grape and wine: II. Qualitative and quantitative determination of monoterpene glycosides in grape. J. Chromatogr. A 1992, 595, 269–281. [Google Scholar] [CrossRef]
- Vilanova, M.; Siebert, T.E.; Varela, C.; Pretorius, I.S.; Henschke, P.A. Effect of ammonium nitrogen supplementation of grape juice on wine volatiles and non-volatiles composition of the aromatic grape variety Albariño. Food Chem. 2012, 133, 124–131. [Google Scholar] [CrossRef]
- Wedler, H.B.; Pemberton, R.P.; Tantillo, D.J. Carbocations and the complex flavor and bouquet of wine: Mechanistic aspects of terpene biosynthesis in wine grapes. Molecules 2015, 20, 10781–10792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrıguez-Concepción, M.; Boronat, A. Elucidation of the methylerythritol phosphate pathway for isoprenoid biosynthesis in bacteria and plastids. A metabolic milestone achieved through genomics. Plant Physiol. 2002, 130, 1079–1089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chizzola, R. Regular monoterpenes and sesquiterpenes (essential oils). Nat. Prod. 2013, 10, 978–983. [Google Scholar]
- Zhao, X.; Ju, Y.; Wei, X.; Dong, S.; Sun, X.; Fang, Y. Significance and transformation of 3-alkyl-2-methoxypyrazines through grapes to wine: Olfactory properties, metabolism, biochemical regulation, and the HP–MP cycle. Molecules 2019, 24, 4598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, T.; Reineccius, G.; Bjorklund, J.; Leete, E. Biosynthesis of 2-methoxy-3-isopropylpyrazine in Pseudomonas perolens. J. Agric. Food Chem. 1991, 39, 1009–1012. [Google Scholar] [CrossRef]
- Dunlevy, J.D.; Soole, K.L.; Perkins, M.V.; Dennis, E.G.; Keyzers, R.A.; Kalua, C.M.; Boss, P.K. Two O-methyltransferases involved in the biosynthesis of methoxypyrazines: Grape-derived aroma compounds important to wine flavour. Plant Mol. Biol. 2010, 74, 77–89. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Massonnet, M.; Cantu, D. The genetic basis of grape and wine aroma. Hortic. Res. 2019, 6, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Mendes-Pinto, M.M. Carotenoid breakdown products the—Norisoprenoids—In wine aroma. Arch. Biochem. Biophys. 2009, 483, 236–245. [Google Scholar] [CrossRef]
- Hjelmeland, A.K.; Ebeler, S.E. Glycosidically bound volatile aroma compounds in grapes and wine: A review. Am. J. Enol. Vitic. 2015, 66, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Dennis, E.G.; Keyzers, R.A.; Kalua, C.M.; Maffei, S.M.; Nicholson, E.L.; Boss, P.K. Grape contribution to wine aroma: Production of hexyl acetate, octyl acetate, and benzyl acetate during yeast fermentation is dependent upon precursors in the must. J. Agric. Food Chem. 2012, 60, 2638–2646. [Google Scholar] [CrossRef]
- Bureau, S.M.; Baumes, R.L.; Razungles, A.J. Effects of vine or bunch shading on the glycosylated flavor precursors in grapes of Vitis vinifera L. cv. Syrah. J. Agric. Food Chem. 2000, 48, 1290–1297. [Google Scholar] [CrossRef]
- Pons, A.; Allamy, L.; Schüttler, A.; Rauhut, D.; Thibon, C.; Darriet, P. What is the expected impact of climate change on wine aroma compounds and their precursors in grape? OENO One 2017, 51, 141–146. [Google Scholar] [CrossRef] [Green Version]
- Allen, M.S.; Lacey, M.J. Methoxypyrazine grape flavour: Influence of climate, cultivar and viticulture. Wein-Wissenschaft 1993, 48, 211–213. [Google Scholar]
- Falcão, L.D.; de Revel, G.; Perello, M.C.; Moutsiou, A.; Zanus, M.C.; Bordignon-Luiz, M.T. A survey of seasonal temperatures and vineyard altitude influences on 2-methoxy-3-isobutylpyrazine, C13-norisoprenoids, and the sensory profile of Brazilian Cabernet Sauvignon wines. J. Agric. Food Chem. 2007, 55, 3605–3612. [Google Scholar] [CrossRef] [Green Version]
- Rienth, M.; Vigneron, N.; Darriet, P.; Sweetman, C.; Burbidge, C.; Bonghi, C.; Walker, R.P.; Famiani, F.; Castellarin, S.D. Grape Berry Secondary Metabolites and Their Modulation by Abiotic Factors in a Climate Change Scenario–A Review. Front. Plant Sci. 2021, 12, 262. [Google Scholar] [CrossRef]
- Kwasniewski, M.T.; Vanden Heuvel, J.E.; Pan, B.S.; Sacks, G.L. Timing of cluster light environment manipulation during grape development affects C13 norisoprenoid and carotenoid concentrations in Riesling. J. Agric. Food Chem. 2010, 58, 6841–6849. [Google Scholar] [CrossRef] [PubMed]
- Augustyn, P.H.; Rapp, A.; Van Wyk, C.J. Some volatile aroma components of Vitis vinifera L. cv. Sauvignon blanc. S. Afr. J. Enol. Vitic. 1982, 3, 52–60. [Google Scholar] [CrossRef] [Green Version]
- Deed, R.C.; Fedrizzi, B.; Gardner, R.C. Influence of fermentation temperature, yeast strain, and grape juice on the aroma chemistry and sensory profile of Sauvignon blanc wines. J. Agric. Food Chem. 2017, 65, 8902–8912. [Google Scholar] [CrossRef]
- Tominaga, T.; Murat, M.L.; Dubourdieu, D. Development of a method for analyzing the volatile thiols involved in the characteristic aroma of wines made from Vitis vinifera L. cv. Sauvignon blanc. J. Agric. Food Chem. 1998, 46, 1044–1048. [Google Scholar] [CrossRef]
- Swiegers, J.H.; Francis, I.L.; Herderich, M.J.; Pretorius, I.S. Meeting consumer expectations through management in vineyard and winery. Wine Ind. J. 2006, 21, 34–43. [Google Scholar]
- Lezaeta, A.; Bordeu, E.; Næs, T.; Varela, P. Exploration of consumer perception of Sauvignon Blanc wines with enhanced aroma properties using two different descriptive methods. Food Res. Int. 2017, 99, 186–197. [Google Scholar] [CrossRef]
- Lyu, X.; Del Prado, D.R.; Araujo, L.D.; Quek, S.Y.; Kilmartin, P.A. Effect of glutathione addition at harvest on Sauvignon Blanc wines. Aust. J. Grape Wine Res. 2021. [Google Scholar] [CrossRef]
- Tominaga, T.; Masneuf, I.; Dubourdieu, D. Powerful aromatic volatile thiols in wines made from several Vitis vinifera L.; Cv. Sauvignon Blanc ACS Symposium Series; ACS Publications: Washington, DC, USA, 2004; Volume 871, pp. 314–337. [Google Scholar]
- Vermeulen, C.; Gijs, L.; Collin, S. Sensorial contribution and formation pathways of thiols in foods: A review. Food Rev. Int. 2005, 21, 69–137. [Google Scholar] [CrossRef]
- Coetzee, C.; du Toit, W.J. A comprehensive review on Sauvignon blanc aroma with a focus on certain positive volatile thiols. Food Res. Int. 2012, 45, 287–298. [Google Scholar] [CrossRef]
- Mateo-Vivaracho, L.; Zapata, J.; Cacho, J.; Ferreira, V. Analysis, occurrence, and potential sensory significance of five polyfunctional mercaptans in white wines. J. Agric. Food Chem. 2010, 58, 10184–10194. [Google Scholar] [CrossRef]
- Tominaga, T.; Niclass, Y.; Frérot, E.; Dubourdieu, D. Stereoisomeric distribution of 3-mercaptohexan-1-ol and 3-mercaptohexyl acetate in dry and sweet white wines made from Vitis vinifera (Var. Sauvignon Blanc and Semillon). J. Agric. Food Chem. 2006, 54, 7251–7255. [Google Scholar] [CrossRef]
- Capone, D.L.; Sefton, M.A.; Jeffery, D.W. Application of a modified method for 3-mercaptohexan-1-ol determination to investigate the relationship between free thiol and related conjugates in grape juice and wine. J. Agric. Food Chem. 2011, 59, 4649–4658. [Google Scholar] [CrossRef]
- Tominaga, T.; Baltenweck-Guyot, R.; Des Gachons, C.P.; Dubourdieu, D. Contribution of volatile thiols to the aromas of white wines made from several Vitis vinifera grape varieties. Am. J. Enol. Vitic. 2000, 51, 178–181. [Google Scholar]
- Peyrot des Gachons, C.; Tominaga, T.; Dubourdieu, D. Sulfur aroma precursor present in S-glutathione conjugate form: Identification of S-3-(hexan-1-ol)-glutathione in must from Vitis vinifera L. cv. Sauvignon blanc. J. Agric. Food Chem. 2002, 50, 4076–4079. [Google Scholar] [CrossRef] [PubMed]
- Subileau, M.; Schneider, R.; Salmon, J.M.; Degryse, E. New insights on 3-mercaptohexanol (3MH) biogenesis in Sauvignon Blanc wines: Cys-3MH and (E)-hexen-2-al are not the major precursors. J. Agric. Food Chem. 2008, 56, 9230–9235. [Google Scholar] [CrossRef] [PubMed]
- Moyano, L.; Zea, L.; Moreno, J.; Medina, M. Analytical study of aromatic series in sherry wines subjected to biological aging. J. Agric. Food Chem. 2002, 50, 7356–7361. [Google Scholar] [CrossRef] [PubMed]
- Escudero, A.; Campo, E.; Ugliano, M. Analytical characterisation of the aroma of five premium red wines. Insights into the role of odor families and the concept of fruitiness of wines. J. Agric. Food Chem. 2007, 55, 4501–4510. [Google Scholar] [CrossRef]
- Parish-Virtue, K.; Herbst-Johnstone, M.; Bouda, F.; Fedrizzi, B.; Deed, R.C.; Kilmartin, P.A. Aroma and Sensory Profiles of Sauvignon Blanc Wines from Commercially Produced Free Run and Pressed Juices. Beverages 2021, 7, 29. [Google Scholar] [CrossRef]
- Lacey, M.J.; Allen, M.S.; Harris, R.L.N.; Brown, W.V. Methoxypyrazines in Sauvignon blanc grapes and wines. Am. J. Enol. Vitic. 1991, 42, 103–138. [Google Scholar]
- Palomo, E.S.; Díaz-Maroto, M.C.; Viñas, M.G.; Soriano-Pérez, A.; Pérez-Coello, M.S. Aroma profile of wines from Albillo and Muscat grape varieties at different stages of ripening. Food Control. 2007, 18, 398–403. [Google Scholar] [CrossRef]
- Sánchez-Palomo, E.; Alañón, M.; Díaz-Maroto, M.; González-Viñas, M.; Pérez-Coello, M. Comparison of extraction methods for volatile compounds of Muscat grape juice. Talanta 2009, 79, 871–876. [Google Scholar] [CrossRef]
- Bordiga, M.; Rinaldi, M.; Locatelli, M.; Piana, G.; Travaglia, F.; Coïsson, J.D.; Arlorio, M. Characterization of Muscat wines aroma evolution using comprehensive gas chromatography followed by a post-analytic approach to 2D contour plots comparison. Food Chem. 2013, 140, 57–67. [Google Scholar] [CrossRef]
- Selli, S.; Canbas, A.; Cabaroglu, T.; Erten, H.; Günata, Z. Aroma components of cv. Muscat of Bornova wines and influence of skin contact treatment. Food Chem. 2006, 94, 319–326. [Google Scholar] [CrossRef]
- Park, S.K.; Morrison, J.C.; Adams, D.O.; Noble, A.C. Distribution of free and glycosidically bound monoterpenes in the skin and mesocarp of Muscat of Alexandria grapes during development. J. Agric. Food Chem. 1991, 39, 514–518. [Google Scholar] [CrossRef]
- Selli, S.; Cabaroglu, T.; Canbas, A.; Erten, H.; Nurgel, C. Effect of skin contact on the aroma composition of the musts of Vitis vinifera L. cv. Muscat of Bornova and Narince grown in Turkey. Food Chem. 2003, 81, 341–347. [Google Scholar] [CrossRef]
- Wu, Y.; Zhang, W.; Song, S.; Xu, W.; Zhang, C.; Ma, C.; Wang, L.; Wang, S. Evolution of volatile compounds during the development of Muscat grape ‘Shine Muscat’ (Vitis labrusca x V. vinifera). Food Chem. 2020, 309, 125778. [Google Scholar] [CrossRef] [PubMed]
- Vilanova, M.; Genisheva, Z.; Graña, M.; Oliveira, J.M. Determination of odorants in varietal wines from international grape cultivars (Vitis vinÃfera) grown in NW Spain. S. Afr. J. Enol. Vitic. 2013, 34, 212–222. [Google Scholar]
- Jesus, D.; Campos, F.M.; Ferreira, M.; Couto, J.A. Characterization of the aroma and colour profiles of fortified Muscat wines: Comparison of Muscat Blanc “à petit grains” grape variety with Red Muscat. Eur. Food Res. Technol. 2017, 243, 1277–1285. [Google Scholar] [CrossRef] [Green Version]
- Capone, S.; Tufariello, M.; Siciliano, P. Analytical characterisation of Negroamaro red wines by “Aroma Wheels”. Food Chem. 2013, 141, 2906–2915. [Google Scholar] [CrossRef]
- Girard, B.; Fukumoto, L.; Mazza, G.; Delaquis, P.; Ewert, B. Volatile terpene constituents in maturing Gewürztraminer grapes from British Columbia. Am. J. Enol. Vitic. 2002, 53, 99–109. [Google Scholar]
- Marais, J. Terpene concentrations and wine quality of Vitis vinifera L. cv. Gewurztraminer as affected by grape maturity and cellar practices. Vitis 1987, 26, 231–245. [Google Scholar]
- Wilson, B.; Strauss, C.R.; Williams, P.J. The distribution of free and glycosidically-bound monoterpenes among skin, juice, and pulp fractions of some white grape varieties. Am. J. Enol. Vitic. 1986, 37, 107–111. [Google Scholar]
- Li, Z.; Howell, K.; Fang, Z.; Zhang, P. Sesquiterpenes in grapes and wines: Occurrence, biosynthesis, functionality, and influence of winemaking processes. Compr. Rev. Food Sci. Food Saf. 2020, 19, 247–281. [Google Scholar] [CrossRef]
- Könen, P.P.; Stötzel, I.; Schwab, W.; Wüst, M. Qualitative profiling of mono-and sesquiterpenols in aglycon libraries from Vitis vinifera L. Gewürztraminer using multidimensional gas chromatography–mass spectrometry. Eur. Food Res. Technol. 2021, 247, 1117–1124. [Google Scholar] [CrossRef]
- Williams, P.J.; Strauss, C.R.; Wilson, B.; Massy-Westropp, R.A. Studies on the hydrolysis of Vitis vinifera monoterpene precursor compounds and model monoterpene β-d-glucosides rationalizing the monoterpene composition of grapes. J. Agric. Food Chem. 1982, 30, 1219–1223. [Google Scholar] [CrossRef]
- Kalua, C.M.; Boss, P.K. Comparison of major volatile compounds from Riesling and Cabernet Sauvignon grapes (Vitis vinifera L.) from fruitset to harvest. Aust. J. Grape Wine Res. 2010, 16, 337–348. [Google Scholar] [CrossRef]
- Rapp, A. Volatile flavour of wine: Correlation between instrumental analysis and sensory perception. Food/Nahrung 1998, 42, 351–363. [Google Scholar] [CrossRef]
- Lee, S.H.; Seo, M.J.; Riu, M.ì; Cotta, J.P.; Block, D.E.; Dokoozlian, N.K.; Ebeler, S.E. Vine microclimate and norisoprenoid concentration in Cabernet Sauvignon grapes and wines. Am. J. Enol. Vitic. 2007, 58, 291–301. [Google Scholar]
- Ryona, I.; Sacks, G.L. Behavior of glycosylated monoterpenes, C13-norisoprenoids, and benzenoids in Vitis vinifera cv. Riesling during ripening and following hedging. In Carotenoid Cleavage Products; American Chemical Society: Washington, DC, USA, 2013; pp. 109–124. [Google Scholar]
- Meyers, J.M.; Sacks, G.L.; Heuvel, J.E.V. Glycosylated aroma compound responses in ‘Riesling’ wine grapes to cluster exposure and vine yield. Hort. Technol. 2013, 23, 581–588. [Google Scholar] [CrossRef] [Green Version]
- Robinson, J.; Harding, J.; Vouillamoz, J. Wine Grapes: A Complete Guide to 1368 Vine Varieties, Including Their Origins and Flavours; Penguin UK: London, UK, 2013. [Google Scholar]
- Lee, S.J.; Noble, A.C. Characterization of odor-active compounds in Californian Chardonnay wines using GC-olfactometry and GC-mass spectrometry. J. Agric. Food Chem. 2003, 51, 8036–8044. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Noble, A.C. Use of partial least squares regression and multidimensional scaling on aroma models of California Chardonnay wines. Am. J. Enol. Vitic. 2006, 57, 363–370. [Google Scholar]
- Martínez-Gil, A.M.; Angenieux, M.; Pardo-García, A.I.; Alonso, G.L.; Ojeda, H.; Salinas, M.R. Glycosidic aroma precursors of Syrah and Chardonnay grapes after an oak extract application to the grapevines. Food Chem. 2013, 138, 956–965. [Google Scholar] [CrossRef] [PubMed]
- Duchêne, E.; Legras, J.L.; Karst, F.; Merdinoglu, D.; Claudel, P.; Jaegli, N.; Pelsy, F. Variation of linalool and geraniol content within two pairs of aromatic and non-aromatic grapevine clones. Aust. J. Grape Wine Res. 2009, 15, 120–130. [Google Scholar] [CrossRef]
- Capone, D.L.; Ristic, R.; Pardon, K.H.; Jeffery, D.W. Simple quantitative determination of potent thiols at ultratrace levels in wine by derivatization and high-performance liquid chromatography–tandem mass spectrometry (HPLC-MS/MS) analysis. Anal. Chem. 2015, 87, 1226–1231. [Google Scholar] [CrossRef] [PubMed]
- Tominaga, T.; Guimbertau, G.; Dubourdieu, D. Contribution of benzenemethanethiol to smoky aroma of certain Vitis vinifera L. wines. J. Agric. Food Chem. 2003, 51, 1373–1376. [Google Scholar] [CrossRef] [PubMed]
- Dashko, S.; Zhou, N.; Tinta, T.; Sivilotti, P.; Lemut, M.S.; Trost, K.; Gamero, A.; Boekhout, T.; Butinar, L.; Vrhovsek, U.; et al. Use of non-conventional yeast improves the wine aroma profile of Ribolla Gialla. J. Ind. Microbiol. Biotechnol. 2015, 42, 997–1010. [Google Scholar] [CrossRef] [PubMed]
- Bavčar, D.; Baša Česnik, H.; Čuš, F.; Košmerl, T. The influence of skin contact during alcoholic fermentation on the aroma composition of Ribolla Gialla and Malvasia Istriana Vitis vinifera (L.) grape wines. IJFST 2011, 46, 1801–1808. [Google Scholar] [CrossRef]
- Maletić, E.; Pejić, I.; Kontić, J.K.; Zdunić, D.; Preiner, D.; Šimon, S.; Andabaka, Z.; Zulimihaljevic, M.; Bubola, M.; Markovic, D.; et al. Ampelographic and genetic characterization of Croatian grapevine varieties. Vitis J. Grapevine Res. 2015, 54, 93–98. [Google Scholar]
- Tomasino, E.; Song, M.; Fuentes, C. Odor perception interactions between free monoterpene isomers and wine composition of Pinot Gris wines. J. Agric. Food Chem. 2020, 68, 3220–3227. [Google Scholar] [CrossRef]
- García-Pastor, M.E.; Serrano, M.; Guillén, F.; Castillo, S.; Martínez-Romero, D.; Valero, D.; Zapata, P.J. Methyl jasmonate effects on table grape ripening, vine yield, berry quality and bioactive compounds depend on applied concentration. Sci. Hortic. 2019, 247, 380–389. [Google Scholar] [CrossRef]
- Mancha, L.A.; Uriarte, D.; Valdés, E.; Moreno, D.; Prieto, M.D.H. Effects of Regulated Deficit Irrigation and Early Cluster Thinning on Production and Quality Parameters in a Vineyard cv. Tempranillo Under Semi-Arid Conditions in Southwestern Spain. Agronomy 2021, 11, 34. [Google Scholar] [CrossRef]
- Askari-Khorasgani, O.; Pessarakli, M. Improving plant yield and quality under normal and stressful conditions by modifying the interactive signaling and metabolic pathways and metabolic interaction networks. In Handbook of Plant and Crop Stress, 4th ed.; CRC Press: Boca Raton, FL, USA, 2019; pp. 893–906. [Google Scholar]
- Liu, S.; Mo, X.; Lin, Z.; Xu, Y.; Ji, J.; Wen, G.; Richey, J. Crop yield responses to climate change in the Huang-Huai-Hai Plain of China. Agric. Water Manag. 2010, 97, 1195–1209. [Google Scholar] [CrossRef]
- Heumesser, C.; Fuss, S.; Szolgayová, J.; Strauss, F.; Schmid, E. Investment in irrigation systems under precipitation uncertainty. Water Resour. Manag. 2012, 26, 3113–3137. [Google Scholar] [CrossRef]
- Chaves, M.M.; Zarrouk, O.; Francisco, R.; Costa, J.M.; Santos, T.; Regalado, A.P.; Rodrigues, M.L.; Lopes, C.M. Grapevine under deficit irrigation: Hints from physiological and molecular data. Ann. Bot. 2010, 105, 661–676. [Google Scholar] [CrossRef] [Green Version]
- Savoi, S.; Wong, D.C.; Arapitsas, P.; Miculan, M.; Bucchetti, B.; Peterlunger, E.; Fait, A.; Mattivi, F.; Castellarin, S.D. Transcriptome and metabolite profiling reveals that prolonged drought modulates the phenylpropanoid and terpenoid pathway in white grapes (Vitis vinifera L.). BMC Plant Biol. 2016, 16, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Savoi, S.; Herrera, J.C.; Carlin, S.; Lotti, C.; Bucchetti, B.; Peterlunger, E.; Castellarin, S.D.; Mattivi, F. From grape berries to wines: Drought impacts on key secondary metabolites. OENO One 2020, 54, 569–582. [Google Scholar] [CrossRef]
- Des Gachons, C.P.; Leeuwen, C.V.; Tominaga, T.; Soyer, J.P.; Gaudillère, J.P.; Dubourdieu, D. Influence of water and nitrogen deficit on fruit ripening and aroma potential of Vitis vinifera L cv Sauvignon blanc in field conditions. J. Sci. Food Agric. 2005, 85, 73–85. [Google Scholar] [CrossRef]
- Myburgh, P.A. Juice and wine quality responses of Vitis vinifera L. cvs. Sauvignon blanc and Chenin blanc to timing of irrigation during berry ripening in the coastal region of South Africa. S. Afr. J. Enol. Vitic. 2006, 27, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Cataldo, E.; Salvi, L.; Mattii, G.B. Effects of irrigation on ecophysiology, sugar content and thiol precursors (3-S-cysteinylhexan-1-ol and 3-S-glutathionylhexan-1-ol) on Vitis vinifera cv. Sauvignon Blanc. Plant Physiol. Biochem. 2021, 164, 247–259. [Google Scholar] [CrossRef]
- Storchi, P.; Giorgessi, F.; Valentini, P.; Tarricone, L.; Bonello, F.; Tamborra, P. Effect of irrigation on vegetative and reproductive behavior of ‘Sauvignon blanc’in Italy. Acta Hortic. 2005, 689, 349–356. [Google Scholar] [CrossRef]
- Giordano, M.; Zecca, O.; Belviso, S.; Reinotti, M.; Gerbi, V.; Rolle, L. Volatile fingerprint and physico-mechanical properties of 'Muscat blanc' grapes grown in mountain area: A first evidence of the influence of water regimes. IJFS 2013, 25, 329. [Google Scholar]
- Reynolds, A.G.; Parchomchuk, P.; Berard, R.; Naylor, A.P.; Hogue, E. Gewurztraminer grapevines respond to length of water stress duration. Int. J. Fruit Sci. 2005, 5, 75–94. [Google Scholar] [CrossRef]
- Romero, P.; Botía, P.; del Amor, F.M.; Gil-Muñoz, R.; Flores, P.; Navarro, J.M. Interactive effects of the rootstock and the deficit irrigation technique on wine composition, nutraceutical potential, aromatic profile, and sensory attributes under semiarid and water limiting conditions. Agric. Water Manag. 2019, 225, 105733. [Google Scholar] [CrossRef]
- Vilanova, M.; Fandiño, M.; Frutos-Puerto, S.; Cancela, J.J. Assessment fertigation effects on chemical composition of Vitis vinifera L. cv. Albariño. Food Chem. 2019, 278, 636–643. [Google Scholar] [CrossRef]
- Kovalenko, Y.; Tindjau, R.; Madilao, L.L.; Castellarin, S.D. Regulated deficit irrigation strategies affect the terpene accumulation in Gewürztraminer (Vitis vinifera L.) grapes grown in the Okanagan valley. Food Chem. 2021, 341, 128172. [Google Scholar] [CrossRef]
- Wang, J.; Abbey, T.; Kozak, B.; Madilao, L.L.; Tindjau, R.; Del Nin, J.; Castellarin, S.D. Evolution over the growing season of volatile organic compounds in Viognier (Vitis vinifera L.) grapes under three irrigation regimes. Food Res. Int. 2019, 125, 108512. [Google Scholar] [CrossRef] [PubMed]
- Frioni, T.; Tombesi, S.; Quaglia, M.; Calderini, O.; Moretti, C.; Poni, S.; Gatti, M.; Moncalvo, A.; Sabbatini, P.; Berrìos, J.G.; et al. Metabolic and transcriptional changes associated with the use of Ascophyllum nodosum extracts as tools to improve the quality of wine grapes (Vitis vinifera cv. Sangiovese) and their tolerance to biotic stress. J. Sci. Food Agric. 2019, 99, 6350–6363. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Gamboa, G.; Romanazzi, G.; Garde-Cerdán, T.; Pérez-Álvarez, E.P. A review of the use of biostimulants in the vineyard for improved grape and wine quality: Effects on prevention of grapevine diseases. J. Sci. Food Agric. 2019, 99, 1001–1009. [Google Scholar] [CrossRef]
- Du Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez-Gamboa, G.; Garde-Cerdán, T.; Rubio-Bretón, P.; Pérez-Álvarez, E.P. Seaweed foliar applications at two dosages to Tempranillo blanco (Vitis vinifera L.) grapevines in two seasons: Effects on grape and wine volatile composition. Food Res. Int. 2020, 130, 108918. [Google Scholar] [CrossRef] [PubMed]
- Garde-Cerdán, T.; Gutiérrez-Gamboa, G.; López, R.; Rubio-Bretón, P.; Pérez-Álvarez, E.P. Influence of foliar application of phenylalanine and urea at two doses to vineyards on grape volatile composition and amino acids content. Vitis 2018, 57, 137–141. [Google Scholar]
- Garde-Cerdán, T.; Portu, J.; López, R.; Santamaría, P. Effect of methyl jasmonate application to grapevine leaves on grape amino acids content. Food Chem. 2016, 203, 536–539. [Google Scholar] [CrossRef]
- Martin, D.; Gershenzon, J.; Bohlmann, J. Induction of volatile terpene biosynthesis and diurnal emission by methyl jasmonate in foliage of Norway spruce (Picea abies). Plant Physiol. 2003, 132, 1586–1599. [Google Scholar] [CrossRef] [Green Version]
- Popescu, G.C.; Popescu, M. Yield, berry quality and physiological response of grapevine to foliar humic acid application. Bragantia 2018, 77, 273–282. [Google Scholar] [CrossRef]
- Bell, S.J.; Henschke, P.A. Implications of nitrogen nutrition for grapes, fermentation and wine. Aust. J. Grape Wine Res. 2005, 11, 242–295. [Google Scholar] [CrossRef]
- Monteiro, F.F.; Bisson, L.F. Biological assay of nitrogen content of grape juice and prediction of sluggish fermentations. Am. J. Enol. Vitic. 1991, 42, 47–57. [Google Scholar]
- Bruwer, F.A.; Du Toit, W.; Buica, A. Nitrogen and sulphur foliar fertilisation. S. Afr. J. Enol. Vitic. 2019, 40, 1. [Google Scholar] [CrossRef]
- Huang, Z.; Ough, C. Effect of vineyard locations, varieties and rootstocks on the juice amino acid composition of several cultivars. Am. J. Enol. Vitic. 1989, 40, 135–139. [Google Scholar]
- Ewart, A.; Kliewer, W. Effects of controlled day and night temperatures and nitrogen on fruit-set, ovule fertility, and fruit composition of several wine grape cultivars. Am. J. Enol. Vitic. 1977, 28, 88–95. [Google Scholar]
- Collins, C.; Wang, X.; Lesefko, S.; De Bei, R.; Fuentes, S. Effects of canopy management practices on grapevine bud fruitfulness. OENO One 2020, 54, 313–325. [Google Scholar] [CrossRef]
- Bell, S.J.; Robson, A. Effect of nitrogen fertilisation on growth, canopy density and yield of Vitis vinifera L. cv. Cabernet Sauvignon. Am. J. Enol. Vitic. 1999, 50, 351–358. [Google Scholar]
- Bordelon, B.P.; Skinkis, P.A.; Howard, P.H. Impact of training system on vine performance and fruit composition of Traminette. Am. J. Enol. Vitic. 2008, 59, 39–46. [Google Scholar]
- Jreij, R.; Kelly, M.T.; Deloire, A.; Brenon, E.; Blaise, A. Combined effects of soil-applied and foliar-applied nitrogen on the nitrogen composition and distribution in water stressed” Vitis vinifera L.” cv Sauvignon blanc grapes. OENO One 2009, 43, 179–187. [Google Scholar] [CrossRef]
- Le Menn, N.; Van Leeuwen, C.; Picard, M.; Riquier, L.; de Revel, G.; Marchand, S. Effect of vine water and nitrogen status, as well as temperature, on some aroma compounds of aged red Bordeaux wines. J. Agric. Food Chem. 2019, 67, 7098–7109. [Google Scholar] [CrossRef]
- Bouzas-Cid, Y.; Falqué, E.; Orriols, I.; Trigo-Córdoba, E.; Díaz-Losada, E.; Fornos-Rivas, D.; Mirás-Avalos, J.M. Amino acids profile of two Galician white grapevine cultivars (Godello and Treixadura). Ciência E Técnica Vitivinícola 2015, 30, 84–93. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Orte, P.; Cacho, J.F.; Ferreira, V. Relationship between varietal amino acid profile of grapes and wine aromatic composition. Experiments with model solutions and chemometric study. J. Agric. Food Chem. 2002, 50, 2891–2899. [Google Scholar] [CrossRef]
- Lytra, G.; Miot-Sertier, C.; Moine, V.; Coulon, J.; Barbe, J.C. Influence of must yeast-assimilable nitrogen content on fruity aroma variation during malolactic fermentation in red wine. Food Res. Int. 2020, 135, 109294. [Google Scholar] [CrossRef]
- Canoura, C.; Kelly, M.T.; Ojeda, H. Effect of irrigation and timing and type of nitrogen application on the biochemical composition of Vitis vinifera L. cv. Chardonnay and Syrah grapeberries. Food Chem. 2018, 241, 171–181. [Google Scholar] [CrossRef]
- Choné, X.; Lavigne-Cruège, V.; Tominaga, T.; van Leeuwen, C.; Castagnède, C.; Saucier, C.; Dubourdieu, D. Effect of vine nitrogen status on grape aromatic potential: Flavor precursors (S-cysteine conjugates), glutathione and phenolic content in Vitis vinifera L. Cv Sauvignon blanc grape juice. OENO One 2006, 40, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Bruwer, F.A. Effect of foliar Nitrogen and Sulphur Spraying on White Wine Composition (Vitis vinifera L. cv. Chenin Blanc and Sauvignon Blanc). Doctoral Dissertation, Stellenbosch University, Stellenbosch, South Africa, 2018. [Google Scholar]
- Lasa, B.; Menendez, S.; Sagastizabal, K.; Cervantes, M.E.C.; Irigoyen, I.; Muro, J.; Aparicio-Tejo, P.M.; Ariz, I. Foliar application of urea to “Sauvignon Blanc” and “Merlot” vines: Doses and time of application. Plant Growth Regul. 2012, 67, 73–81. [Google Scholar] [CrossRef]
- Gaiotti, F.; Lucchetta, M.; Rodegher, G.; Lorenzoni, D.; Longo, E.; Boselli, E.; Cesco, S.; Belfiore, N.; Lovat, N.; Delgado-López, J.M.; et al. Urea-doped calcium phosphate nanoparticles as sustainable nitrogen nanofertilizers for viticulture: Implications on yield and quality of Pinot gris grapevines. Agronomy 2021, 11, 1026. [Google Scholar] [CrossRef]
- Wang, Y.; He, Y.N.; Chen, W.K.; He, F.; Chen, W.; Cai, X.D.; Duan, C.Q.; Wang, J. Effects of cluster thinning on vine photosynthesis, berry ripeness and flavonoid composition of Cabernet Sauvignon. Food Chem. 2018, 248, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; He, Y.N.; He, L.; He, F.; Chen, W.; Duan, C.Q.; Wang, J. Changes in global aroma profiles of cabernet sauvignon in response to cluster thinning. Food Res. Int. 2019, 122, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Moreno, D.; Vilanova, M.; Gamero, E.; Intrigliolo, D.S.; Talaverano, M.I.; Uriarte, D.; Valdés, M.E. Effects of preflowering leaf removal on phenolic composition of Tempranillo in the semiarid terroir of western Spain. Am. J. Enol. Vitic. 2015, 66, 204–211. [Google Scholar] [CrossRef] [Green Version]
- Xi, X.; Zha, Q.; He, Y.; Tian, Y.; Jiang, A. Influence of cluster thinning and girdling on aroma composition in ‘Jumeigui’table grape. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Alem, H.; Ojeda, H.; Rigou, P.; Schneider, R.; Torregrosa, L. The reduction of plant sink/source does not systematically improve the metabolic composition of Vitis vinifera white fruit. Food Chem. 2021, 345, 128825. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, A.G.; Schlosser, J.; Sorokowsky, D.; Roberts, R.; Willwerth, J.; De Savigny, C. Magnitude of viticultural and enological effects. II. Relative impacts of cluster thinning and yeast strain on composition and sensory attributes of Chardonnay Musqué. Am. J. Enol. Vitic. 2007, 58, 25–41. [Google Scholar]
- Kok, D. Influences of pre-and post-veraison cluster thinning treatments on grape composition variables and monoterpene levels of Vitis vinifera L. cv. Sauvignon Blanc. J. Food Agric. Environ. 2011, 9, 22–26. [Google Scholar]
- Naor, A.; Gal, Y.; Bravdo, B. Shoot and cluster thinning influence vegetative growth, fruit yield, and wine quality of Sauvignon blanc'grapevines. J. Am. Soc. Hortic. 2002, 127, 628–634. [Google Scholar] [CrossRef] [Green Version]
- Škrab, D.; Sivilotti, P.; Comuzzo, P.; Voce, S.; Degano, F.; Carlin, S.; Arapitsas, P.; Masuero, D.; Vrhovšek, U. Cluster Thinning and Vineyard Site Modulate the Metabolomic Profile of Ribolla Gialla Base and Sparkling Wines. Metabolites 2021, 11, 331. [Google Scholar] [CrossRef]
- Kliewer, W.M.; Dokoozlian, N.K. Leaf area/crop weight ratios of grapevines: Influence on fruit composition and wine quality. Am. J. Enol. Vitic. 2005, 56, 170–181. [Google Scholar]
- Pereira, G.E.; Gaudillere, J.P.; Pieri, P.; Hilbert, G.; Maucourt, M.; Deborde, C.; Moing, A.; Rolin, D. Microclimate influence on mineral and metabolic profiles of grape berries. J. Agric. Food Chem. 2006, 54, 6765–6775. [Google Scholar] [CrossRef]
- Feng., H.; Yuan, F.; Skinkis, P.; Qian, M.C. Effect of cluster zone leaf removal on grape sugar, acids, carotenoids, and volatile composition. Am. J. Enol. Vitic. 2012, 63, 458A. [Google Scholar]
- Falchi, R.; Bonghi, C.; Drincovich, M.F.; Famiani, F.; Lara, M.V.; Walker, R.P.; Vizzotto, G. Sugar metabolism in stone fruit: Source-sink relationships and environmental and agronomical effects. Front. Plant Sci. 2020, 11, 1820. [Google Scholar] [CrossRef]
- Gerdes, S.; Winterhalter, P.; Ebeler, S. Effect of sunlight exposure on norisoprenoid formation in white Riesling grapes. In Carotenoid-Derived Aroma Compounds; Chapter 19; Winterhalter, P., Rouseff, R.L., Eds.; Symposium Series; American Chemical Society: Washington, DC, USA, 2002; Volume 802, pp. 262–272. [Google Scholar] [CrossRef]
- Feng, H.; Yuan, F.; Skinkis, P.A.; Qian, M.C. Influence of cluster zone leaf removal on Pinot noir grape chemical and volatile composition. Food Chem. 2015, 173, 414–423. [Google Scholar] [CrossRef] [PubMed]
- Osrečak, M.; Karoglan, M.; Kozina, B.; Preiner, D. Influence of leaf removal and reflective mulch on phenolic composition of white wines. OENO One 2015, 49, 183–193. [Google Scholar] [CrossRef]
- Marais, J.; van Wyk, C.J.; Rapp, A. Effect of sunlight and shade on norisoprenoid levels in maturing Weisser Riesling and Chenin Blanc grapes and Weisser Riesling wines. S. Afr. J. Enol. Vitic. 1992, 13, 23–32. [Google Scholar]
- Mosetti, D.; Herrera, J.C.; Sabbatini, P.; Green, A.; Alberti, G.; Peterlunger, E.; Lisjak, K.; Castellarin, S.D. Impact of leaf removal after berry set on fruit composition and bunch rot in ‘Sauvignon blanc’. Vitis J. Grapevine Res. 2016, 55, 57–64. [Google Scholar]
- Yue, X.; Ma, X.; Tang, Y.; Wang, Y.; Wu, B.; Jiao, X.; Zhang, Z.; Ju, Y. Effect of cluster zone leaf removal on monoterpene profiles of Sauvignon Blanc grapes and wines. Food Res. Int. 2020, 131, 109028. [Google Scholar] [CrossRef]
- Bureau, S.M.; Razungles, A.J.; Baumes, R.L. The aroma of Muscat of Frontignan grapes: Effect of the light environment of vine or bunch on volatiles and glycoconjugates. J. Sci. Food Agric. 2000, 80, 2012–2020. [Google Scholar] [CrossRef]
- Reynolds, A.G.; Wardle, D.A. Influence of fruit microclimate on monoterpene levels of Gewürztraminer. Am. J. Enol. Vitic. 1989, 40, 149–154. [Google Scholar]
- Russell, D.W. 3-Hydroxy-3-methylglutaryl-CoA reductase from pea seedlings. Plastid HMG-CoA reductase: Assay, isolation and properties. Methods Enzymol. 1985, 110, 36–40. [Google Scholar]
- Smart, R.E.; Smith, S.M.; Winchester, R.V. Light quality and quantity effects on fruit ripening for Cabernet Sauvignon. Am. J. Enol. Vitic. 1988, 39, 250–258. [Google Scholar]
- Bubola, M.; Rusjan, D.; Lukić, I. Crop level vs. leaf removal: Effects on Istrian Malvasia wine aroma and phenolic acids composition. Food Chem. 2020, 312, 126046. [Google Scholar] [CrossRef]
- Bubola, M.; Lukić, I.; Radeka, S.; Sivilotti, P.; Grozić, K.; Vanzo, A.; Bavcar, D.; Lisjak, K. Enhancement of Istrian Malvasia wine aroma and hydroxycinnamate composition by hand and mechanical leaf removal. J. Sci. Food Agric. 2019, 99, 904–914. [Google Scholar] [CrossRef] [PubMed]
- Lobos, G.A.; Acevedo-Opazo, C.; Guajardo-Moreno, A.; Valdés-Gómez, H.; Taylor, J.A.; Laurie, V.F. Effects of kaolin-based particle film and fruit zone netting on Cabernet Sauvignon grapevine physiology and fruit quality. OENO One 2015, 49, 137–144. [Google Scholar] [CrossRef]
- Coniberti, A.; Ferrari, V.; Dellacassa, E.; Boido, E.; Carrau, F.; Gepp, V.; Disegna, E. Kaolin over sun-exposed fruit affects berry temperature, must composition and wine sensory attributes of Sauvignon blanc. Eur. J. Agron. 2013, 50, 75–81. [Google Scholar] [CrossRef]
- Scafidi, P.; Pisciotta, A.; Patti, D.; Tamborra, P.; Di Lorenzo, R.; Barbagallo, M.G. Effect of artificial shading on the tannin accumulation and aromatic composition of the Grillo cultivar (Vitis vinifera L.). BMC Plant Biol. 2013, 13, 175. [Google Scholar] [CrossRef] [Green Version]
- Marais, J.; Calitz, F.; Haasbroek, P.D. Relationship between microclimatic data, aroma component concentrations and wine quality parameters in the prediction of Sauvignon Blanc wine quality. S. Afr. J. Enol. Vitic. 2001, 22, 47–51. [Google Scholar] [CrossRef] [Green Version]
- Ford, R.J. The Effect of Shading and Crop Load on Flavour and Aroma Compounds in Sauvignon Blanc Grapes and Wine. Doctoral Dissertation, Lincoln University, Lincoln, UK, 2007. [Google Scholar]
- Ghiglieno, I.; Mattivi, F.; Cola, G.; Trionfini, D.; Perenzoni, D.; Simonetto, A.; Gilioli, G.; Valenti, L. The effects of leaf removal and artificial shading on the composition of Chardonnay and Pinot noir grapes. OENO One 2020, 54, 761–777. [Google Scholar] [CrossRef]
- Belancic, A.; Agosin, E.; Ibacache, A.; Bordeu, E.; Baumes, R.; Razungles, A.; Bayonove, C. Influence of Sun exposure on the aromatic composition of Chilean Muscat grape cultivars Moscatel de alejandria and Moscatel rosada. Am. J. Enol. Vitic. 1997, 48, 181–186. [Google Scholar]
- Gutiérrez-Gamboa, G.; Zheng, W.; de Toda, F.M. Current viticultural techniques to mitigate the effects of global warming on grape and wine quality: A comprehensive review. Food Res. Int. 2020, 10, 9946. [Google Scholar]
- Gutiérrez-Gamboa, G.; Zheng, W.; de Toda, F.M. Strategies in vineyard establishment to face global warming in viticulture: A mini review. J. Sci. Food Agric. 2021, 101, 1261–1269. [Google Scholar] [CrossRef] [PubMed]
- Frioni, T.; Saracino, S.; Squeri, C.; Tombesi, S.; Palliotti, A.; Sabbatini, P.; Magnanini, E.; Poni, S. Understanding kaolin effects on grapevine leaf and whole-canopy physiology during water stress and re-watering. J. Plant Physiol. 2019, 242, 153020. [Google Scholar] [CrossRef] [PubMed]
- Zoecklein, B.W.; Wolf, T.K.; Duncan, S.E.; Marcy, J.E.; Jasinski, Y. Effect of fruit zone leaf removal on total glycoconjugates and conjugate fraction concentration of Riesling and Chardonnay (Vitis vinifera L.) grapes. Am. J. Enol. Vitic. 1998, 49, 259–265. [Google Scholar]
- van der Hulst, L.; Munguia, P.; Culbert, J.A.; Ford, C.M.; Burton, R.A.; Wilkinson, K.L. Accumulation of volatile phenol glycoconjugates in grapes following grapevine exposure to smoke and potential mitigation of smoke taint by foliar application of kaolin. Planta 2019, 249, 941–952. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Interaction among factors that can affect the nutrition of the vineyard and call for a need to calibrate agronomic practice. The factors illustrated in the figure interact with each other and influence the nutritional and health status of the plant, which in turn affects the aromas of the berries.
Figure 1.
Interaction among factors that can affect the nutrition of the vineyard and call for a need to calibrate agronomic practice. The factors illustrated in the figure interact with each other and influence the nutritional and health status of the plant, which in turn affects the aromas of the berries.


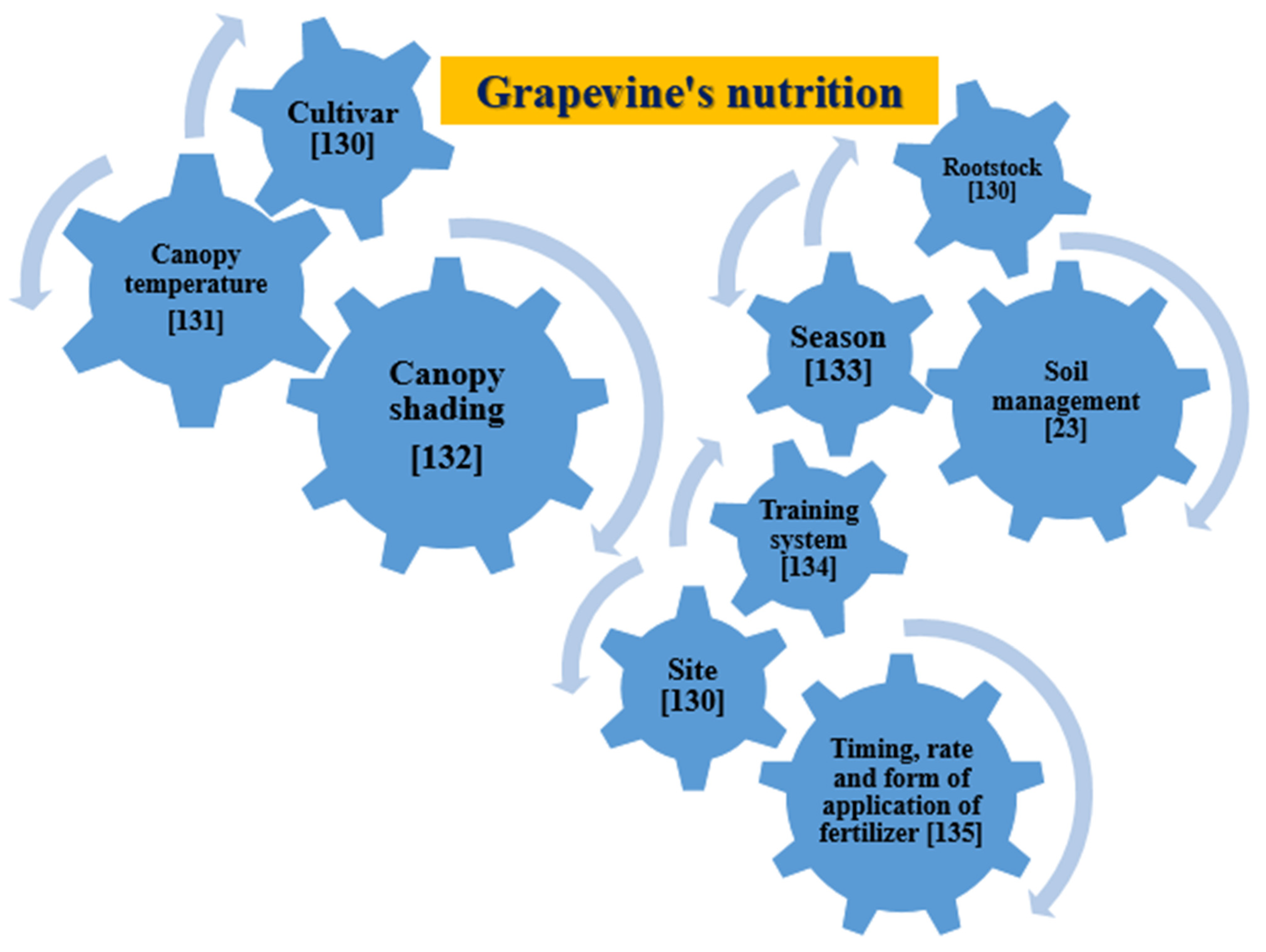{kind=link}
Table 1.
Volatile thiols contributing to Sauvignon blanc’s aroma.
| Thiol | Abbreviation | Olfactory Description | Perception Threshold (ng/L) | Range in Wines Repored in Literature (ng/L) | Bibliography |
|---|---|---|---|---|---|
| 4-mercapto-4-methylpentan-2-one | 4MMP | passionfruit and boxtree | 0.8 | 4–40 | [59] |
| 4-mercapto-4-methylpentan-2-ol | 4MMPOH | citrus zest | 55 | 90 | [58] |
| 3-mercaptohexan-1-ol | 3MH | gooseberry and guava | 50–100 | 26–18,000 | [59,60,61] |
| 3-mercapto-hexylacetate | 3MHA | grapefruit and boxtree | 2.5–4.2 | 0–2500 | [59,60] |
| 3-mercapto-3-methylbutan-l-ol | 3MMB | cooked leeks | 1300 | - | [62] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Cataldo, E.; Salvi, L.; Paoli, F.; Fucile, M.; Mattii, G.B. Effect of Agronomic Techniques on Aroma Composition of White Grapevines: A Review. Agronomy 2021, 11, 2027. https://doi.org/10.3390/agronomy11102027
AMA Style
Cataldo E, Salvi L, Paoli F, Fucile M, Mattii GB. Effect of Agronomic Techniques on Aroma Composition of White Grapevines: A Review. Agronomy. 2021; 11(10):2027. https://doi.org/10.3390/agronomy11102027
Chicago/Turabian StyleCataldo, Eleonora, Linda Salvi, Francesca Paoli, Maddalena Fucile, and Giovan Battista Mattii. 2021. "Effect of Agronomic Techniques on Aroma Composition of White Grapevines: A Review" Agronomy 11, no. 10: 2027. https://doi.org/10.3390/agronomy11102027
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.