
Prophylactic Administration of Fucoidan Represses Cancer Metastasis by Inhibiting Vascular Endothelial Growth Factor (VEGF) and Matrix Metalloproteinases (MMPs) in Lewis Tumor-Bearing Mice

 ,
, 
Abstract
:1. Introduction
2. Results
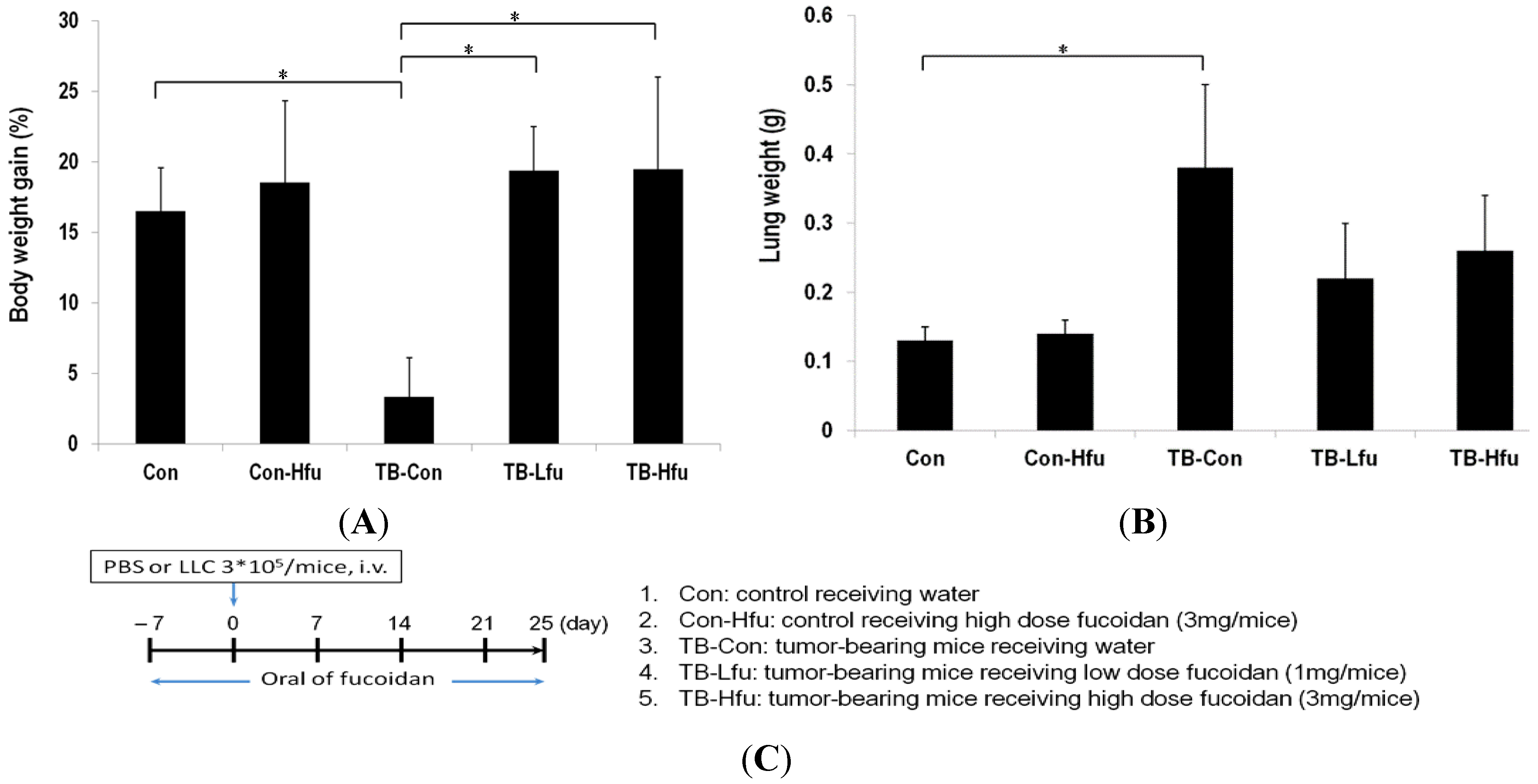
2.1. Fucoidan Mitigates Cachectic Symptoms in LLC-Inoculated C57BL/6 Mice


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Con | Con-Hfu | TB-Con | TB-Lfu | TB-Hfu |
|---|---|---|---|---|---|
| (A) Hematology parameter | |||||
| Total red cell count (1012/L) | 9.7 ± 0.2 | 9.4 ± 0.2 | 6.5 ± 0.2 | 9.2 ± 0.4 b | 9.1 ± 0.8 b |
| Total white cell count (109/L) | 18.9 ± 1.4 | 23 ± 1.0 a | 6.9 ± 0.8 | 14.2 ± 2.0 b | 14.2 ± 2.2 b |
| Absolute neutrophil count (109/L) | 3.8 ± 0.7 | 4.9 ± 0.4 a | 2.1 ± 0.6 | 4.1 ± 0.9 b | 3.4 ± 0.1 b |
| Absolute monocyte count (109/L) | 1.1 ± 0.2 | 1.3 ± 0.1 | 0.6 ± 0.1 | 0.9 ± 0.3 b | 0.9 ± 0.3 b |
| Absolute lymphocyte count (109/L) | 14.1 ± 0.5 | 16.8 ± 1.3 a | 4.4 ± 0.7 | 10.7 ± 1.7 b | 10.6 ± 1.6 b |
| (B) Spleenocyte parameter | |||||
| CD3+ (%) | 37.6 ± 4.2 | 35.8 ± 5.6 | 31.2 ± 2.7 | 35.5 ± 2.5 b | 33.5 ± 1.5 |
| CD4+ (%) | 19.4 ± 1.8 | 15.8 ± 0.8 a | 16.28 ± 2.6 | 15.1 ± 2.0 | 15.4 ± 1.9 |
| CD8+ (%) | 16.8 ± 3.7 | 17.3 ± 1.5 | 14.5 ± 1.7 | 17.1 ± 3.0 | 15.4 ± 1.4 |
| CD19+ (%) | 45.7 ± 5.0 | 43.4 ± 4.4 | 47.5 ± 4.4 | 47.2 ± 3.9 | 45.4 ± 3.5 |
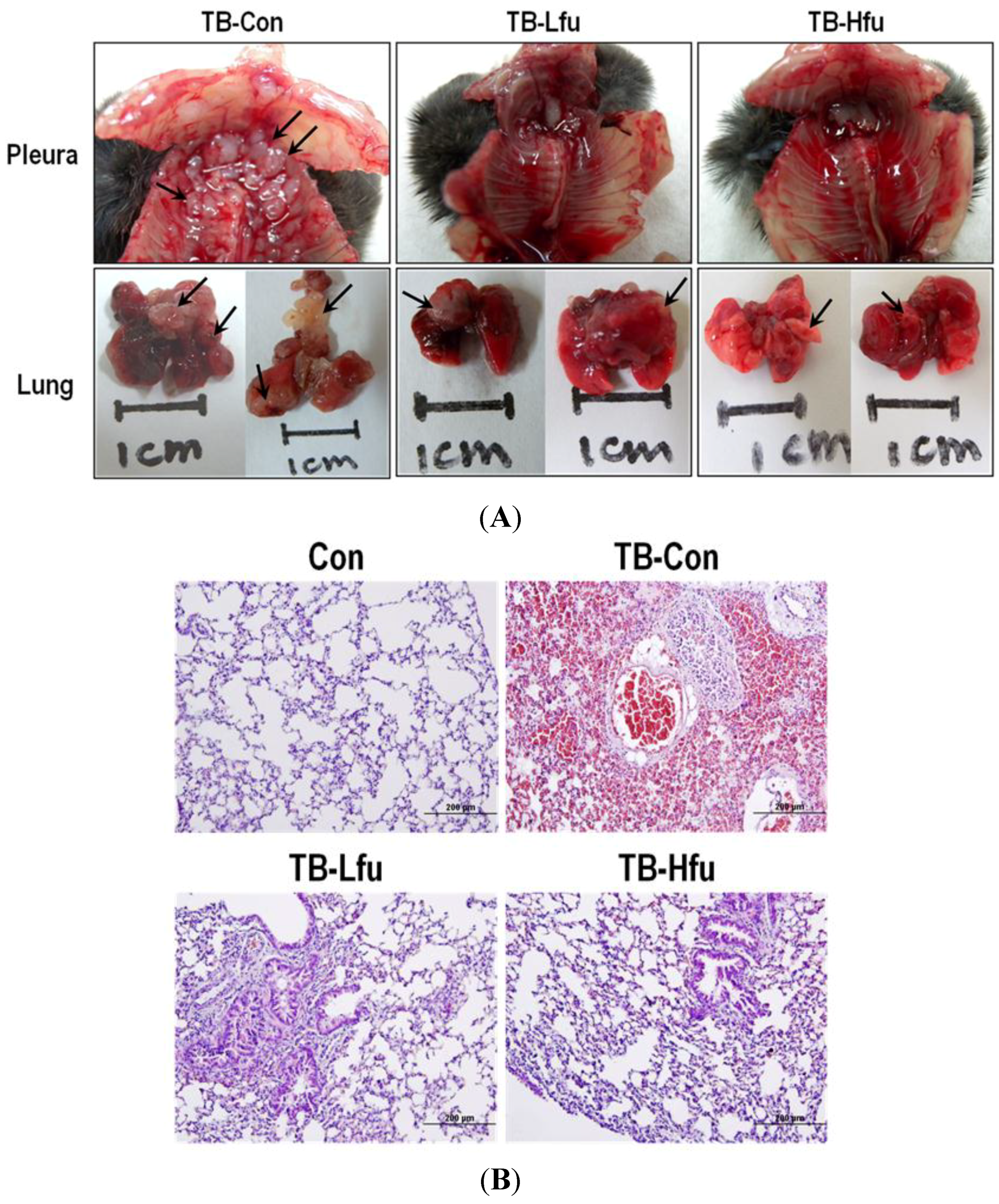
2.2. Fucoidan Inhibits Lung Metastatic Colonization of LLC Cells in C57BL/6 Mice

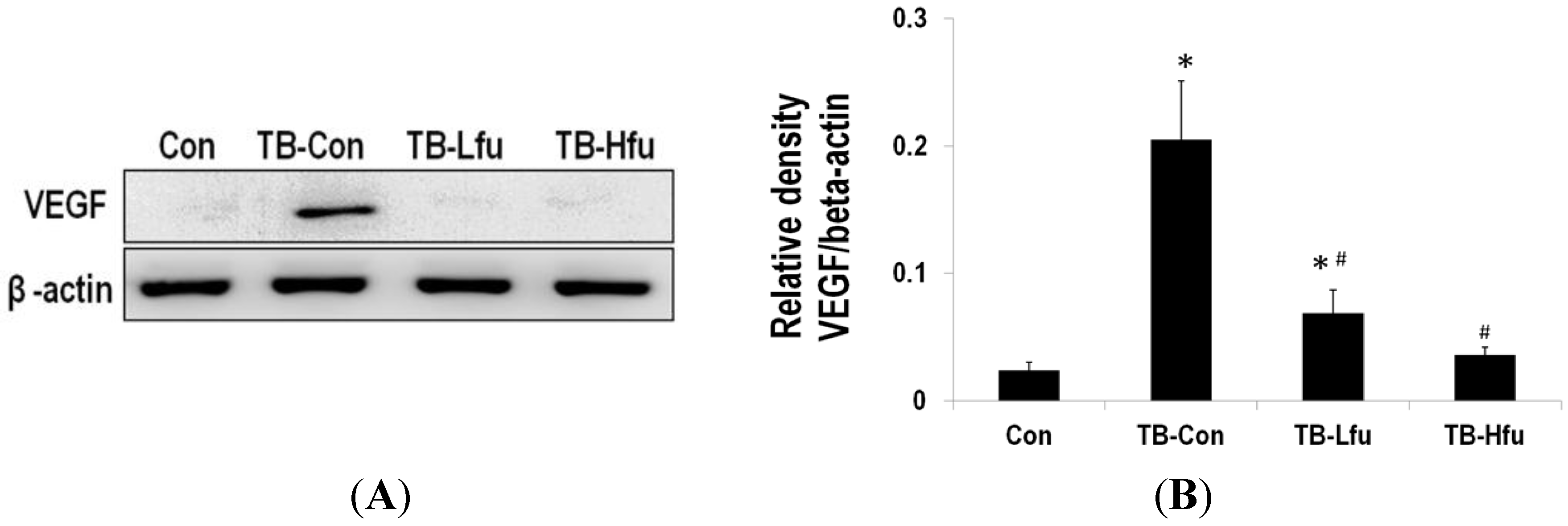
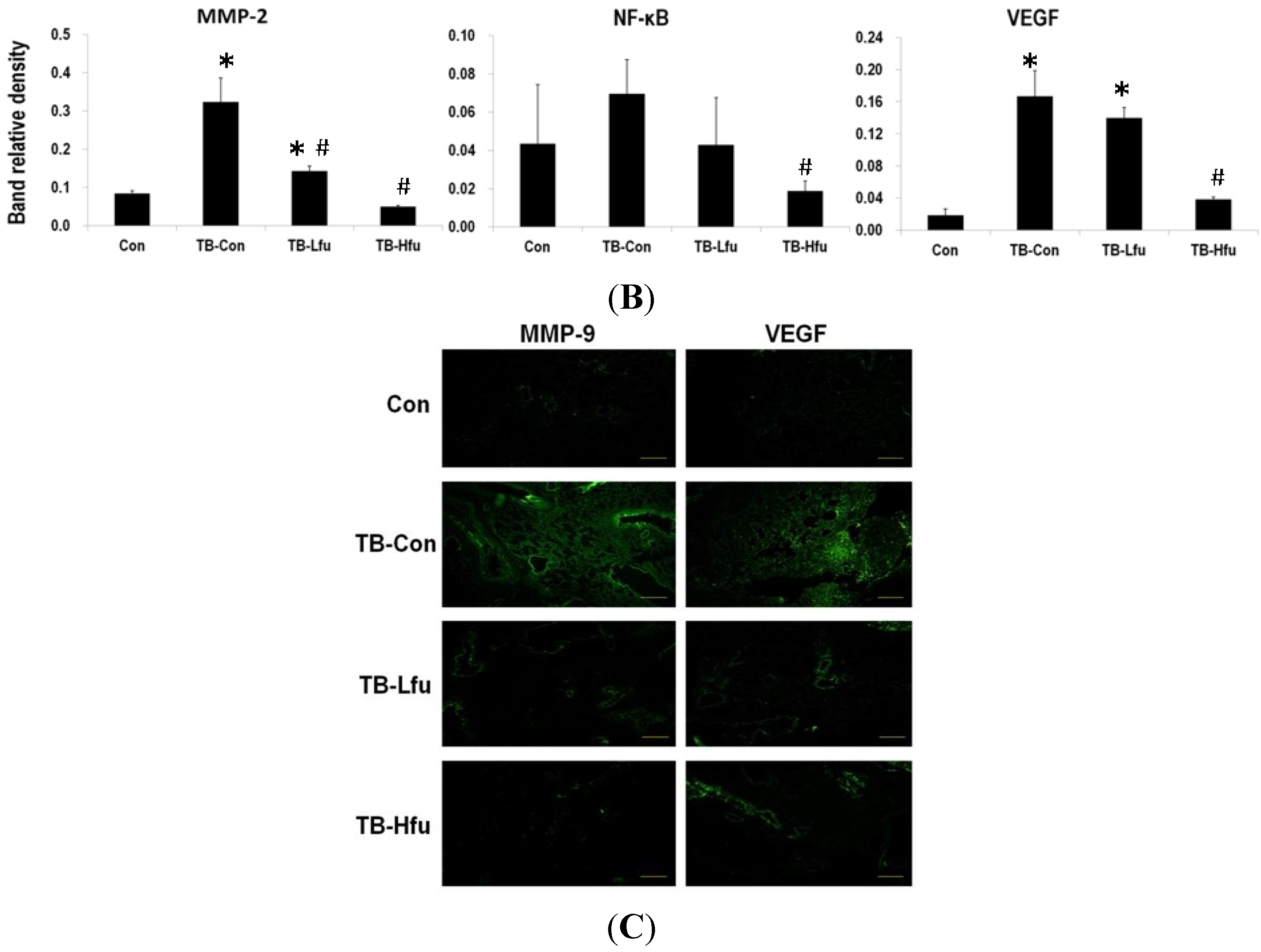
2.3. Fucoidan Restrains LLCs Metastasis by Suppressing Expression of VEGF and MMPs



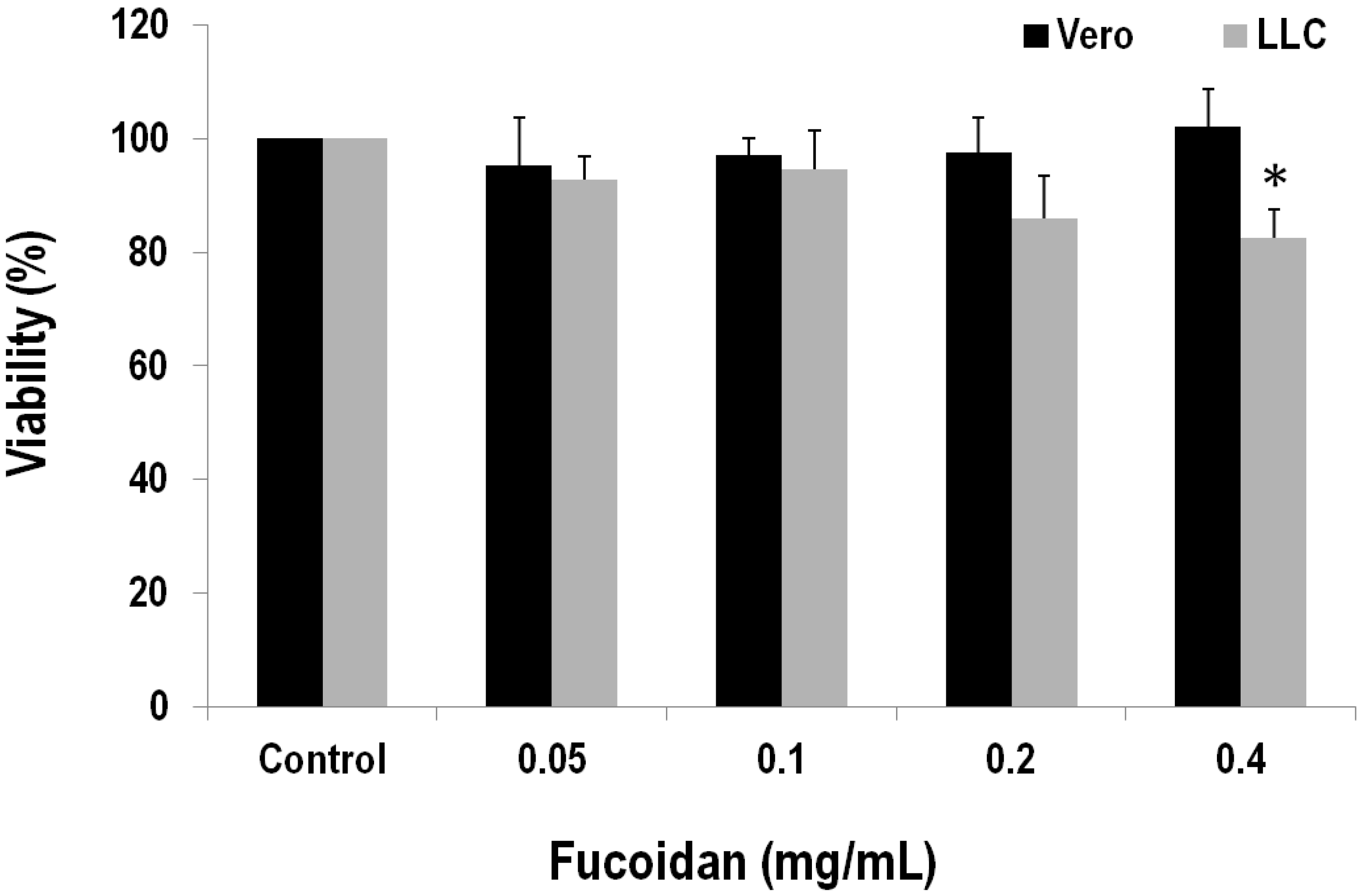
2.4. Fucoidan Has a Cytotoxic Effect on the LLC Cell Line

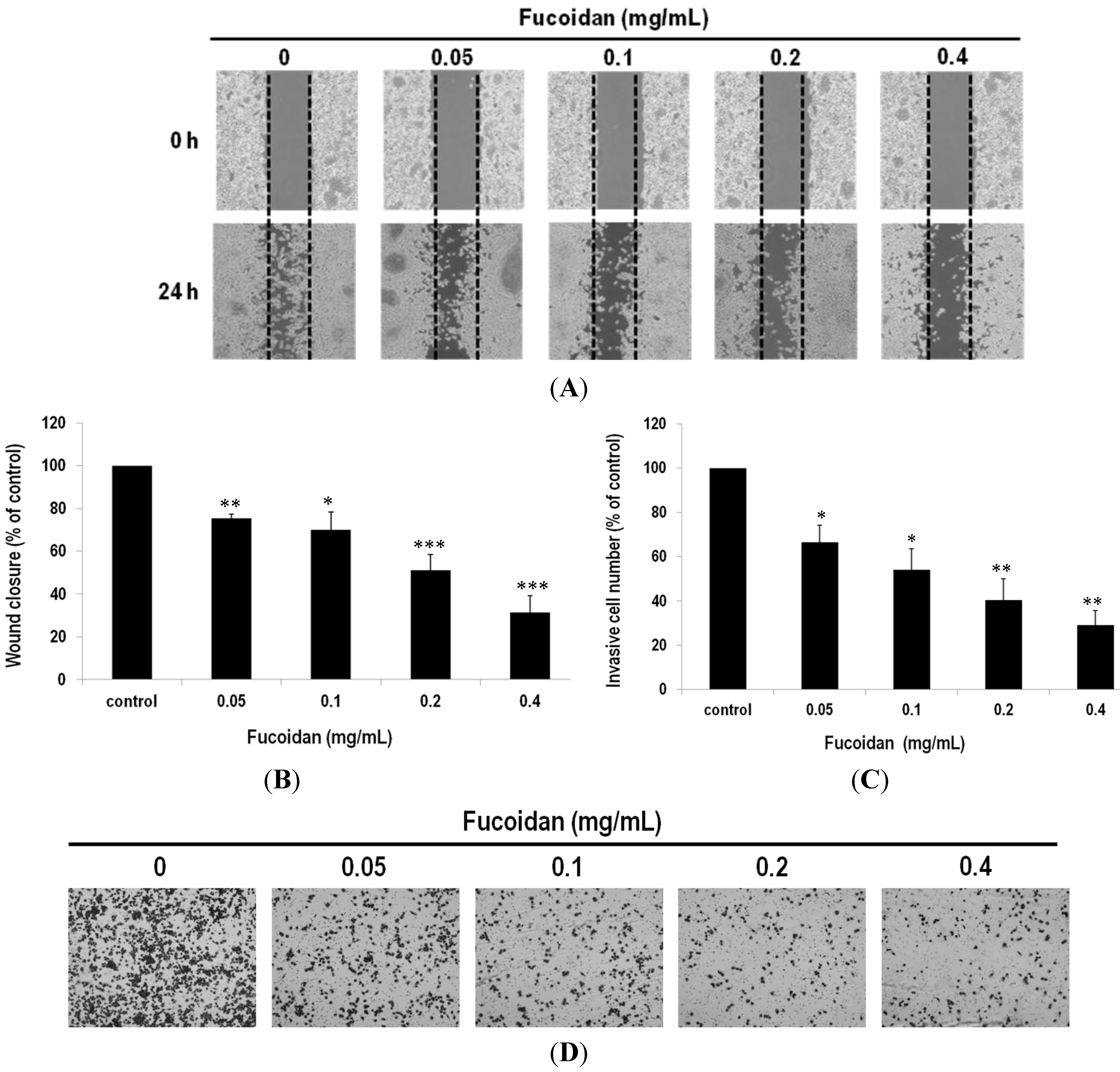
2.5. Fucoidan Prevents Metastasis of Lung Adenocarcinoma Cells

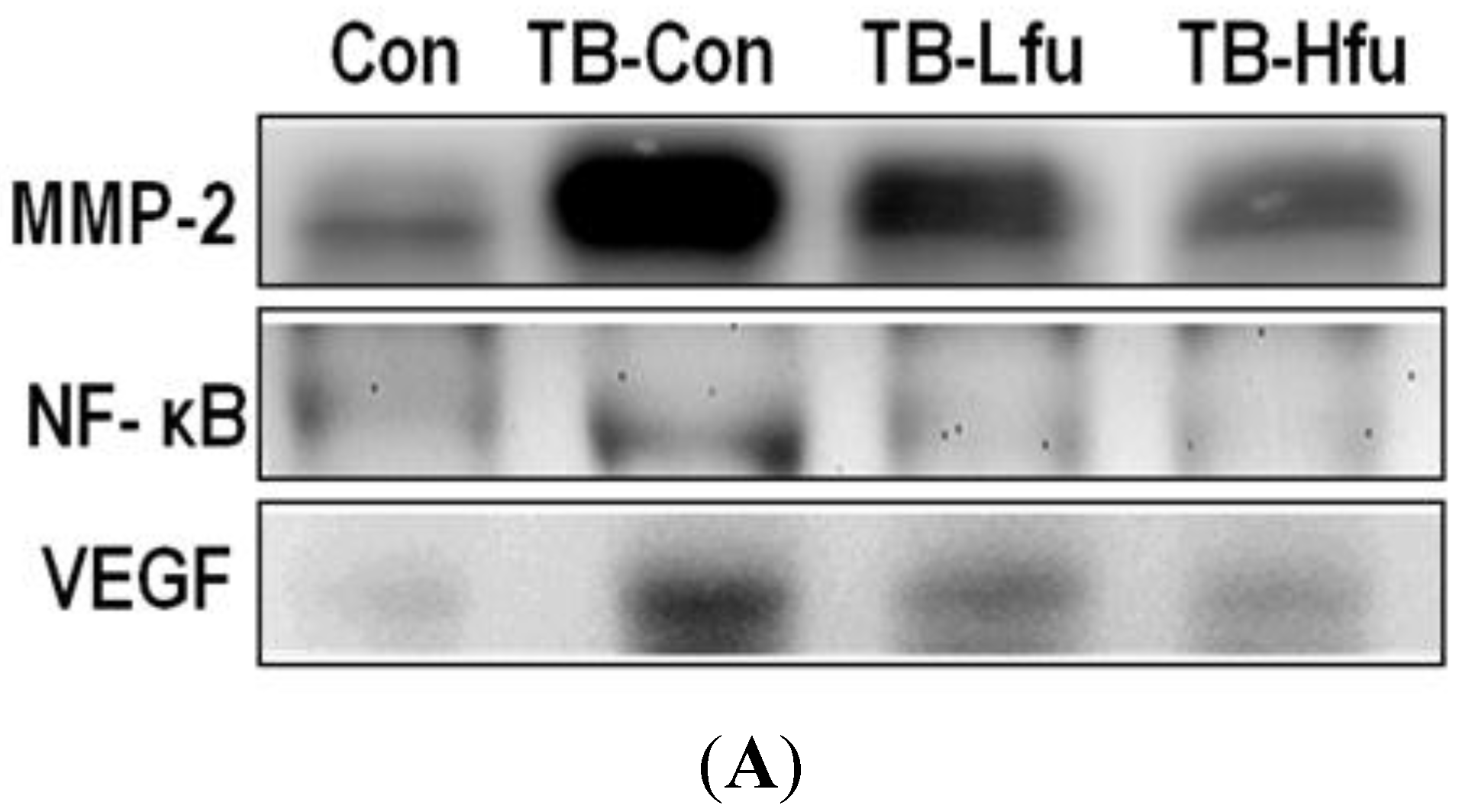
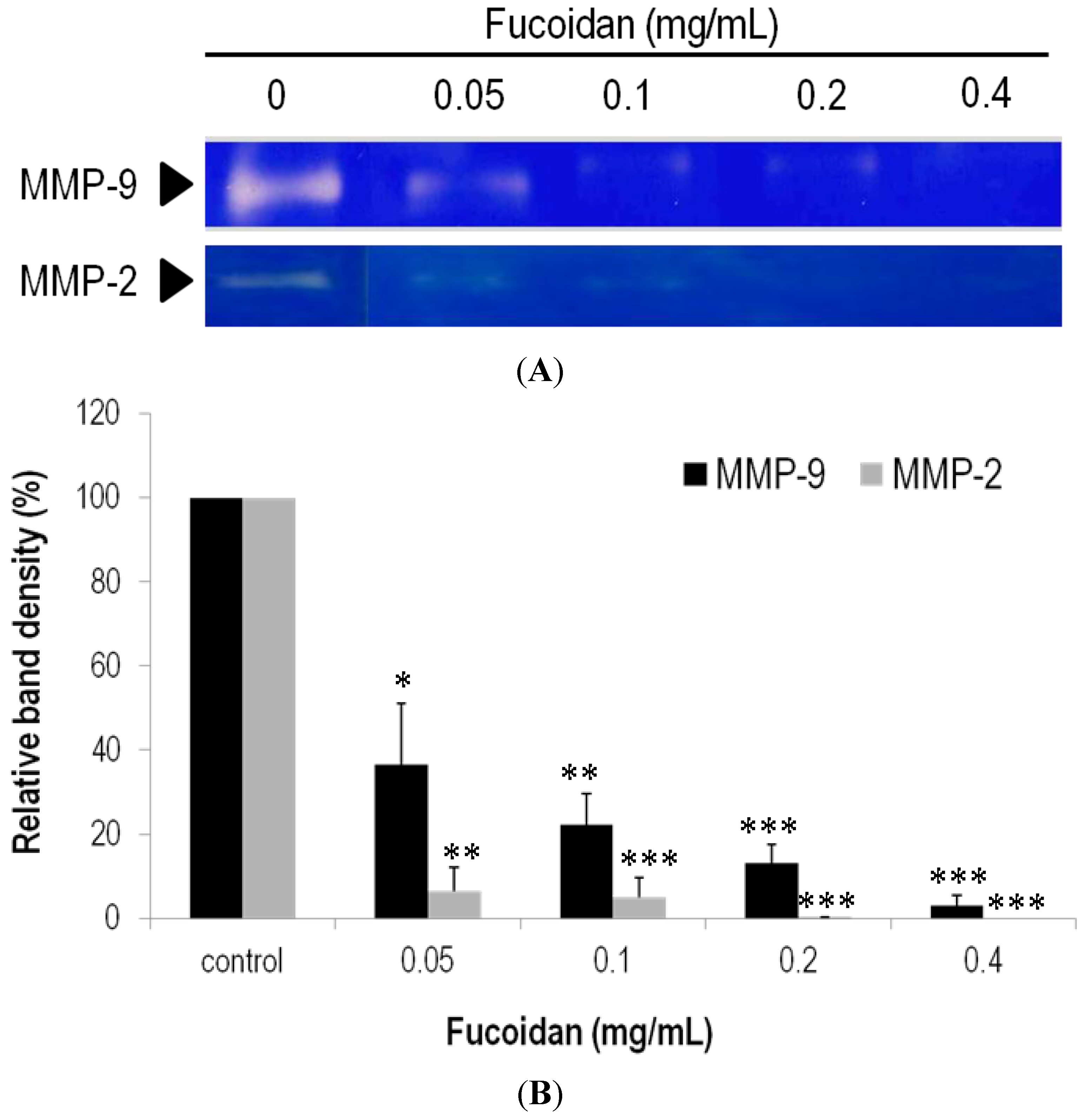
2.6. Effects of Fucoidan on Expression and Activity of MMPs

3. Discussion
4. Materials and Method
4.1. Cells and Cell Culture
4.2. Preparation of Fucoidan Extract
4.3. Ethical Approval and Animals
4.4. Experimental Design
4.5. Blood Sample Analysis
4.6. Flow Cytometry
4.7. Lewis Lung Carcinoma Cells Metastasis Models
4.8. Western Blot Analysis
4.9. Histopathological Analysis
4.10. Immunofluorescence Assay
4.11. Cell Viability Assay
4.12. Cell Migration and Invasion Assays
4.13. Gelatin Zymography
4.14. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ferlay, J.; Soerjomataram, I.I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in GLOBOCAN 2012. Int. J. Cancer 2014. [Google Scholar] [CrossRef]
- Meoni, G.; Cecere, F.L.; Lucherini, E.; di Costanzo, F. Medical treatment of advanced non-small cell lung cancer in elderly patients: A review of the role of chemotherapy and targeted agents. J. Geriatr. Oncol. 2013, 4, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Bremnes, R.M.; Camps, C.; Sirera, R. Angiogenesis in non-small cell lung cancer: The prognostic impact of neoangiogenesis and the cytokines VEGF and bFGF in tumours and blood. Lung Cancer 2006, 51, 143–158. [Google Scholar] [CrossRef] [PubMed]
- Juhász, E.; Kim, J.H.; Klingelschmitt, G.; Walzer, S. Effects of erlotinib first-line maintenance therapy versus placebo on the health-related quality of life of patients with metastatic non-small-cell lung cancer. Eur. J. Cancer 2013, 49, 1205–1215. [Google Scholar] [CrossRef] [PubMed]
- Folkman, J. Role of angiogenesis in tumor growth and metastasis. Semin. Oncol. 2002, 29, 15–18. [Google Scholar] [CrossRef] [PubMed]
- Husmann, K.; Arlt, M.J.; Muff, R.; Langsam, B.; Bertz, J.; Born, W.; Fuchs, B. Matrix Metalloproteinase 1 promotes tumor formation and lung metastasis in an intratibial injectionosteosarcoma mouse model. Biochim. Biophys. Acta 2013, 1832, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Safranek, J.; Pesta, M.; Holubec, L.; Kulda, V.; Dreslerova, J.; Vrzalova, J.; Topolcan, O.; Pesek, M.; Finek, J.; Treska, V. Expression of MMP-7, MMP-9, TIMP-1 and TIMP-2 mRNA in lung tissue of patients with non-small cell lung cancer (NSCLC) and benign pulmonary disease. Anticancer Res. 2009, 29, 2513–2517. [Google Scholar] [PubMed]
- Zhou, H.; Wu, A.; Fu, W.; Lv, Z.; Zhang, Z. Significance of semaphorin-3A and MMP-14 protein expression in non-small cell lung cancer. Oncol. Lett. 2014, 7, 1395–1400. [Google Scholar] [PubMed]
- Wimberger, P.; Chebouti, I.; Kasimir-Bauer, S.; Lachmann, R.; Kuhlisch, E.; Kimmig, R.; Süleyman, E.; Kuhlmann, J.D. Explorative investigation of vascular endothelial growth factor receptor expression in primary ovarian cancer and its clinical relevance. Gynecol. Oncol. 2014, 133, 467–472. [Google Scholar] [CrossRef] [PubMed]
- Shinkaruk, S.; Bayle, M.; Laïn, G.; Déléris, G. Vascular endothelial cell growth factor (VEGF), an emerging target for cancer chemotherapy. Curr. Med. Chem. Anticancer Agents 2003, 3, 95–117. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Zhu, J.; Liu, D.Y.; Li, H.Y.; Xu, N.; Hou, M. Over-expression of survivin and VEGF in small-cell lung cancer may predict the poorer prognosis. Med. Oncol. 2014, 31, 775. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Xu, S.; Jiao, F.; Ren, T.; Li, Q. Vascular endothelial growth factor B coordinates metastasis of non-small cell lung cancer. Tumour. Biol. 2014. [Google Scholar] [CrossRef]
- Sanmartín, E.; Sirera, R.; Usó, M.; Blasco, A.; Gallach, S.; Figueroa, S.; Martínez, N.; Hernando, C.; Honguero, A.; Martorell, M.; et al. A gene signature combining the tissue expression of three angiogenic factors is a prognostic marker in early-stage non-small cell lung cancer. Ann. Surg. Oncol. 2014, 21, 612–620. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wu, Q.; Rowley, D.C.; Al-Kareef, A.M.; Wang, H. Anticancer agent-based marine natural products and related compounds. J. Asian Nat. Prod. Res. 2015, 3, 1–18. [Google Scholar]
- Nagamine, T.; Nakazato, K.; Tomioka, S.; Iha, M.; Nakajima, K. Intestinal absorption of fucoidan extracted from the brown seaweed, Cladosiphon okamuranus. Mar. Drugs 2014, 13, 48–64. [Google Scholar] [CrossRef] [PubMed]
- Silva, T.H.; Moreira-Silva, J.; Marques, A.L.; Domingues, A.; Bayon, Y.; Reis, R.L. Marine origin collagens and its potential applications. Mar. Drugs 2014, 12, 5881–5901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nair, D.G.; Weiskirchen, R.; Al-Musharafi, S.K. The use of marine-derived bioactive compounds as potential hepatoprotective agents. Acta Pharmacol. Sin. 2014. [Google Scholar] [CrossRef]
- Kwak, J.Y. Fucoidan as a marine anticancer agent in preclinical development. Mar. Drugs 2014, 12, 851–870. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Zhang, Q.; Chen, L.; Ren, S.; Xu, P.; Tang, Y.; Luo, D. Higher specificity of the activity of low molecular weight fucoidan for thrombin-induced platelet aggregation. Thrombo. Res. 2010, 125, 419–426. [Google Scholar] [CrossRef]
- Semenov, A.V.; Mazurov, A.V.; Preobrazhenskaia, M.E.; Ushakova, N.A.; Mikhaĭlov, V.I.; Berman, A.E.; Usov, A.I.; Nifant’ev, N.E.; Bovin, N.V. Sulfated polysaccharides as inhibitors of receptor activity of P-selectin and P-selectin-dependent inflammation. Vopr. Med. Khim. 1998, 44, 135–144. [Google Scholar] [PubMed]
- Wang, J.; Zhang, Q.; Zhang, Z.; Song, H.; Li, P. Potential antioxidant and anticoagulant capacity of low molecular weight fucoidan fractions extracted from Laminaria japonica. Int. J. Biol. Macromol. 2010, 46, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Veena, C.K.; Josephine, A.; Preetha, S.P.; Varalakshmi, P.; Sundarapandiyan, R. Renal peroxidative changes mediated by oxalate: The protective role of fucoidan. Life Sci. 2006, 79, 1789–1795. [Google Scholar] [CrossRef] [PubMed]
- Hayakawa, K.; Nagamine, T. Effect of fucoidan on the biotinidase kinetics in human hepatocellular carcinoma. Anticancer Res. 2009, 29, 1211–1217. [Google Scholar] [PubMed]
- Min, E.Y.; Kim, I.H.; Lee, J.; Kim, E.Y.; Choi, Y.H.; Nam, T.J. The effects of fucodian on senescence are controlled by the p16INK4a-pRb and p14Arf-p53 pathways in hepatocellular carcinoma and hepatic cell lines. Int. J. Oncol. 2014, 45, 47–56. [Google Scholar] [PubMed]
- Chen, S.; Zhao, Y.; Zhang, Y.; Zhang, D. Fucoidan induces cancer cell apoptosis by modulating the endoplasmic reticulum stress cascades. PLoS ONE 2014, 9, e108157. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Kim, J.S.; Kim, E. Fucoidan from seaweed fucus vesiculosus inhibits migration and invasion of human lung cancer cell via PI3K-Akt-mTOR pathways. PLoS ONE 2012, 7, e50624. [Google Scholar] [CrossRef] [PubMed]
- Riou, D.; Colliec-Jouault, S.; Pinczon du Sel, D.; Bosch, S.; Siavoshian, S.; le Bert, V.; Tomasoni, C.; Sinquin, C.; Durand, P.; Roussakis, C. Antitumor and antiproliferative effects of a fucan extracted from ascophyllum nodosum against a non-small-cell bronchopulmonary carcinoma line. Anticancer Res. 1996, 16, 1213–1218. [Google Scholar] [PubMed]
- Koyanagi, S.; Tanigawa, N.; Nakagawa, H.; Soeda, S.; Shimeno, H. Oversulfation of fucoidan enhances its anti-angiogenic and antitumor activities. Biochem. Pharmacol. 2003, 65, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Xue, M.; Ge, Y.; Zhang, J.; Wang, Q.; Hou, L.; Liu, Y.; Sun, L.; Li, Q. Anticancer properties and mechanisms of fucoidan on mouse breast cancer in vitro and in vivo. PLoS ONE 2012, 7, e43483. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; Peng, Z.; Chen, L.; Si, J.; Dong, Z.; Xia, Y. Nuclear factor-κB modulates cellular glutathione and prevents oxidative stress in cancer cells. Cancer Lett. 2010, 299, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Willett, C.G.; Boucher, Y.; Duda, D.G.; di Tomaso, E.; Munn, L.L.; Tong, R.T.; Kozin, S.V.; Petit, L.; Jain, R.K.; Chung, D.C.; et al. Surrogate markers for antiangiogenic therapy and dose-limiting toxicities for bevacizumab with radiation and chemotherapy: Continued experience of a phase I trial in rectal cancer patients. J. Clin. Oncol. 2005, 23, 8136–8139. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.D.; Chap, L.I.; Holmes, F.A.; Cobleigh, M.A.; Marcom, P.K.; Fehrenbacher, L.; Dickler, M.; Overmoyer, B.A.; Reimann, J.D.; Sing, A.P.; et al. Randomized phase III trial of capecitabine compared with bevacizumab plus capecitabine in patients with previously treated metastatic breast cancer. J. Clin. Oncol. 2005, 23, 792–799. [Google Scholar] [CrossRef] [PubMed]
- Lester, J.F.; MacBeth, F.R.; Coles, B. Prophylactic cranial irradiation for preventing brain metastases in patients undergoingradical treatment for non-small- cell lung cancer: A Cochrane Review. Int. J. Radiat. Oncol. Biol. Phys. 2005, 63, 690–694. [Google Scholar] [CrossRef] [PubMed]
- Hicklin, D.J.; Ellis, L.M. Role of the vascular endothelial growth factor pathway in tumor growth and angiogenesis. J. Clin. Oncol. 2005, 23, 1011–1027. [Google Scholar] [CrossRef] [PubMed]
- Ale, M.T.; Maruyama, H.; Tamauchi, H.; Mikkelsen, J.D.; Meyer, A.S. Fucoidan from Sargassum. sp. and Fucus. vesiculosus reduces cell viability of lung carcinoma and melanoma cells in vitro and activates natural killer cells in mice in vivo. Int. J. Biol. Macromol. 2011, 49, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.R.; Lee, J.S.; Song, J.E.; Ha, S.J.; Hong, E.K. Inonotus obliquus-derived polysaccharide inhibits the migration and invasion of human non-small cell lung carcinoma cells via suppression of MMP-2 and MMP-9. Int. J. Oncol. 2014, 45, 2533–2540. [Google Scholar] [PubMed]
- Rask, L.; Fregil, M.; Høgdall, E.; Mitchelmore, C.; Eriksen, J. Development of a metastatic fluorescent Lewis Lung carcinoma mouse model: Identification of mRNAs and microRNAs involved in tumor invasion. Gene 2013, 517, 72–81. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, T.-H.; Chiu, Y.-H.; Chan, Y.-L.; Chiu, Y.-H.; Wang, H.; Huang, K.-C.; Li, T.-L.; Hsu, K.-H.; Wu, C.-J. Prophylactic Administration of Fucoidan Represses Cancer Metastasis by Inhibiting Vascular Endothelial Growth Factor (VEGF) and Matrix Metalloproteinases (MMPs) in Lewis Tumor-Bearing Mice. Mar. Drugs 2015, 13, 1882-1900. https://doi.org/10.3390/md13041882
Huang T-H, Chiu Y-H, Chan Y-L, Chiu Y-H, Wang H, Huang K-C, Li T-L, Hsu K-H, Wu C-J. Prophylactic Administration of Fucoidan Represses Cancer Metastasis by Inhibiting Vascular Endothelial Growth Factor (VEGF) and Matrix Metalloproteinases (MMPs) in Lewis Tumor-Bearing Mice. Marine Drugs. 2015; 13(4):1882-1900. https://doi.org/10.3390/md13041882
Chicago/Turabian StyleHuang, Tse-Hung, Yi-Han Chiu, Yi-Lin Chan, Ya-Huang Chiu, Hang Wang, Kuo-Chin Huang, Tsung-Lin Li, Kuang-Hung Hsu, and Chang-Jer Wu. 2015. "Prophylactic Administration of Fucoidan Represses Cancer Metastasis by Inhibiting Vascular Endothelial Growth Factor (VEGF) and Matrix Metalloproteinases (MMPs) in Lewis Tumor-Bearing Mice" Marine Drugs 13, no. 4: 1882-1900. https://doi.org/10.3390/md13041882