
Pleural Lubrication


Department of Pathophysiology and Transplantation, Section of Human Physiology, University of Milan, Via Mangiagalli 32, 20133 Milan, Italy
*
Author to whom correspondence should be addressed.
Lubricants 2016, 4(2), 15; https://doi.org/10.3390/lubricants4020015
Submission received: 27 January 2016
/
Revised: 21 March 2016
/
Accepted: 28 April 2016
/
Published: 19 May 2016
(This article belongs to the Special Issue Friction and Lubricants Related to Human Bodies)
{kind=link}
{kind=link}
Abstract
:During breathing, the pleural surfaces slide against each other continuously without damage. Pleural liquid and lubricating molecules should provide the lubrication of the sliding surfaces, thus protecting the mesothelium from shear-induced abrasion. D’Angelo et al. (Respir. Physiol. Neurobiol. 2004) measured the coefficient of kinetic friction (μ) of rabbit parietal pleura sliding against visceral pleura in vitro at physiological velocities and under physiological loads; it was ~0.02 and did not change with sliding velocity, consistent with boundary lubrication. μ in boundary lubrication can be influenced by surface molecules like hyaluronan, sialomucin or surface active phospholipidis. Hyaluronan or sialomucin is able to restore good boundary lubrication in damaged mesothelium. Nevertheless, hyaluronidase and neuraminidase treatment of the mesothelium does not increase μ, though neuraminidase cleaves sialic acid from the mesothelium. Short pronase or phospholipase treatment, so as to affect only the mesothelial glycocalyx, increases μ, and this increase is removed by hyaluronan or sialomucin. On the other hand, addition of phospholipids after phospholipase treatment produces a small effect relative to that of hyaluronan or sialomucin, and this effect is similar with unsaturated or saturated phospholipids. In damaged mesothelium, the lubrication regimen becomes mixed, but addition of hyaluronan or sialomucin restores boundary lubrication.
1. Introduction
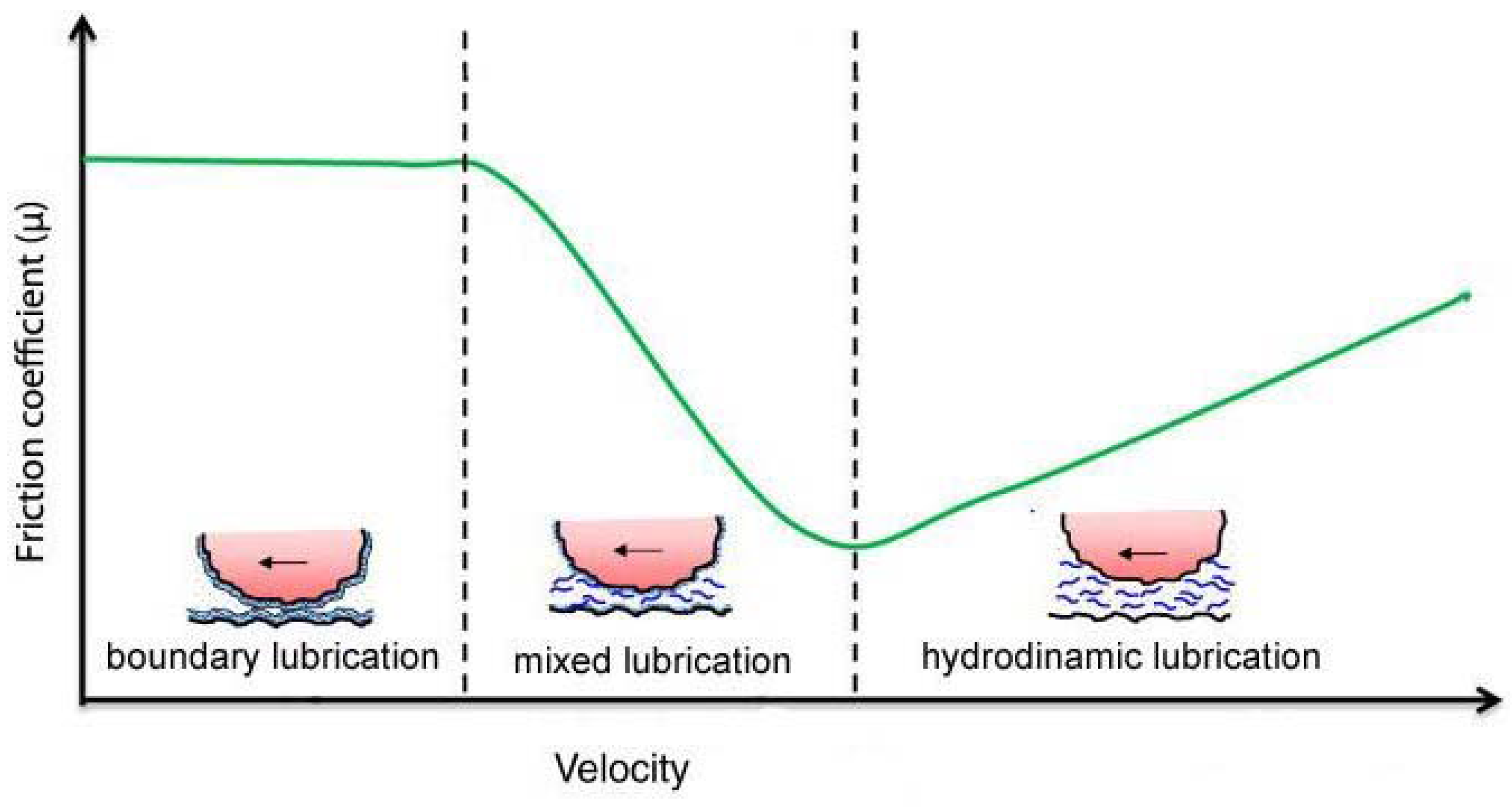
During breathing, the pleural surfaces of the lung and chest wall slide against each other. During a maximal inhalation from low lung volume, the caudal margin of the human lung slides between the rib cage and diaphragm, for a distance of 10 cm or more, though the highest sliding velocity during normal breathing should not exceed 3 cm/s, even during maximal ventilation. The surfaces of the pleural space consist of a single layer of delicate mesothelial cells covered with microvilli up to 6 µm long [1]. Although the mesothelium can also easily be damaged by gentle handling [2], these mesothelial surfaces are able to slide continuously with respect to each other without damage, thereby preventing pathological adhesion. Pleural liquid and lubricating molecules should provide the lubrication of the sliding surfaces, thus protecting the mesothelium from shear-induced abrasion. A central question concerning pleural space mechanics is the nature of the coupling of the lung and chest wall through the pleural mesothelia. Efficient ventilation requires the pleural surfaces to slide with minimal friction while remaining in close apposition to allow immediate and full transmission of the forces generated by the chest wall to the lung. There are two models that at present attempt to explain this coupling. The first [3] assumes that over most of the pleural surface a downward flow of pleural liquid is adjusted in such a way as to match the vertical gradients of pleural liquid and pleural surface pressure: the largest part of the pleural surfaces would be separated by a uniform, relatively thick (20 µm) layer of pleural liquid, under normal load. Recent computational fluid dynamical studies have shown that sliding movements of the pleurae during breathing would promote spatial uniformity of liquid thickness and prevent the surfaces from coming into contact [4,5]. The second model [6] assumes that mechanisms that continuously remove fluid from the pleural space, thus reducing the average thickness of the pleural liquid to less than 10 µm, lower pleural liquid pressure below pleural surface pressure and pull together the visceral and parietal pleura, which eventually contact each other over part of their surface. The preponderance of the experimental evidence indicates that pleural liquid pressure is lower than pleural surface pressure over most of the mesothelial surfaces [6]. Moreover, measurements of the thickness of the pleural space support the hypothesis of contact of the pleural surface, also considering that pleural cells are covered by microvilli. Assessment of the frictional behavior of mesothelial surfaces sliding against each other should contribute to the understanding of pleural space mechanics. The tribological behavior of sliding surface is often described with the Stribeck curve (Figure 1; see [7]) in which the coefficient of friction (µ) varies with velocity. This curve can be divided into three regimens of lubrication: boundary, mixed, and hydrodynamic lubrication. In boundary lubrication, asperities on the sliding surfaces are in contact or separated by extremely thin films of lubricant, and hydrodynamic pressures are irrelevant in supporting the normal load. In this regime, µ does not change with velocity. In hydrodynamic lubrication, fluid thickness is much greater than the amplitude of the surface roughness, hydrodynamic pressure bears the entire load, and µ rises with velocity. In mixed lubrication, asperities are deformed by hydrodynamic pressures and do not come into contact being separated by a continuous layer of lubricant. In mixed lubrication, µ decreases with increasing velocity. Thus, each lubrication regime has its own characteristic frictional behavior. Hence, in exploring the tribological behavior of sliding mesothelial tissues, it can be useful to investigate the occurrence and the importance of contact between the sliding surfaces. In 2004 D’Angelo et al. [8] measured the frictional force of rabbit parietal pleura sliding against visceral pleura in vitro, while oscillating at physiological velocities (from 0.88 to 3.02 cm/s) and amplitudes, under physiological load (~0.8 to ~12.9 cm H2O). In situ, the pleural surfaces are subjected to a compressive force, i.e., a normal load, except at the level of the interlobar fissures and costophrenic sinus where pleural liquid and pleural surface pressure are necessarily the same. According to direct measurements of pleural liquid and pleural surface pressure made with different techniques in several animal species [6], the difference between pleural liquid and pleural surface pressure decreases from top to bottom of the pleural space, and increases with increasing the depth of inspiration. However, maximal spatially averaged pressures holding the opposed mesothelial surfaces together during resting or moderately increased ventilation should not exceed 10 cm H2O. The apparatus used to measure the frictional force consists of a sliding platform connected through unextensible threads to the core of a differential transformer, and of a balance arm held stationary at its fulcrum by a force transducer. The specimen of parietal pleura was fixed with the pleural surface facing upwards to the sliding platform, while that of the visceral pleura was fixed with the pleural surface facing downwards to a Perspex rod attached to one end of the balance arm. The balance arm was held horizontal, and counterweights added to its other end enabled to change the normal force applied to the tissue. The frictional force in the direction of motion was measured by the force transducer and μ was computed as the slope of the relationship between the load and the frictional force recorded in the central 50% of the excursion of the sliding platform. The study showed that the frictional force is linearly related to the force normal to the sliding pleural mesothelium, whilst it is largely independent of speed of motion and nominal contact area. Hence, friction between mesothelial surfaces obeys Amonton’s law, i.e., the frictional force is directly proportional to the normal load and independent of the nominal contact area, and Coulomb’s law, i.e., the frictional force is independent of velocity of sliding, the coefficient of kinetic friction being almost constant, consistent with boundary lubrication. Therefore, this result is in line with the model, which assumes contact between pleural surfaces. The values of m for the pleural mesothelium was 0.019 ± 0.02 with pleural liquid between the mesothelial surfaces, and 0.027 (p < 0.01) with Ringer solution. No damage of the mesothelial microvilli in the test specimens was found by transmission electron microscopy. After gentle blotting of the pleural surface for 1–2 min with filter paper, m increased ~6 times; only a part of this increase was due to liquid removal. In fact, if, after blotting, the mesothelium was wetted with Ringer, m was ~3 times greater than that at control.
Recently, Kim et al. [9] determined the stiffness and surface topography of specimens of rat parietal pleura by mean of the atomic force microscopy. The roughness normal to the surface was less than 10 µm (mostly ~5 µm). Considering the softness of the pleura, they concluded that under physiological conditions visceral and parietal pleura could conform to each other through the agency of local hydrodynamic pressure without requiring tissue–tissue contacts, i.e., elastohydrodynamic lubrication should occur over the whole pleural surface. In discussing the view implying contact areas with boundary lubrication, these authors concluded that either the contact pressure should be so high as to damage the contacting tissues or the contact area should be much larger than suggested by microscopic observations. This point deserves a comment. With a coefficient of kinetic friction of 0.02, the largest spatially averaged shear stress applied to the pleural surface in situ should not exceed 0.2 cm H2O, thus ensuring minimal energy dissipation. Both in situ and in vitro, the contact pressure must, however, exceed the spatially averaged contact pressure, depending on the effective area of contact [10]. There are no data on the pleural contact area to allow computation of the effective contact pressure, but the effective contact area could be substantial, due to the presence of a dense carpet of relatively long microvilli [11,12,13], free cells [14], and macromolecules [11,15,16] that behave as load bearing elements [17], thus ensuring a relatively low value for the ratio between effective and nominal contact pressure. From Murphy and Macklem equation [10], which provides the balance of forces within the pleural space, letting Ppl = −3 cm H2O and Pliq = −5 cm H2O, with a contact area of 20%, the contact pressure is 5 cm H2O. At the end of a deep inspiration the difference between Pliq and Ppl increases, but the thickness of the pleural space decreases markedly [18], and, therefore, the percentage of contact area should increase. Consequently, the contact pressure should not increase so much as to damage the mesothelium. Whatever the effective contact area and pressure, the morphological results of D’Angelo et al. [8] show that for physiological values of spatially averaged contact pressure and sliding speeds, shear stresses at the site of contacts are small enough to prevent damage of the mesothelia. The occurrence of contact areas does not prevent the continuity of pleural liquid. Lubrication should be boundary in the contact areas, and elastohydrodynamic elsewhere [6,14,19,20]. This view is compatible with small flows of pleural liquid caused by gravity, breathing and cardiac movements [3,21,22,23,24]. Finally, one has to recall that despite the opposed recoil of the lung and the chest wall (that draw liquid into the pleural space) the volume of the pleural liquid under physiological conditions is kept to a minimum. This implies that there are mechanisms that absorb liquid from the pleural space and others that prevent a complete removal [14,25]. Evidences for mechanisms absorbing liquid from the pleural space have been provided [14,20]. Until there is no evidence for a negative chemical feedback preventing a complete liquid removal, it seems that the only mechanism is provided by the contact between lung and chest wall [14,20] through suitable boundary lubricants [11,12,15,26].
2. Boundary Lubricants
The boundary lubrication of mesothelium can be influenced by the properties of the mesothelium uppermost layer. Several specialized molecules adherent to the surface, like sialomucins, hyaluronan, or surface-active phospholipids have been proposed as boundary lubricants. Anyway, the identification of the lubricating substances has been hindered by the lack of knowledge of the precise composition of the mesothelial coating and its organization.
2.1. Sialomucin Lubricant Effects
The presence of sialomucins between sliding tissues has been known for a long time [27,28]; indeed, sialomucins are cell-membrane associated mucins that have been found on joint cartilage, where they showed lubricant relevance [29]. Their presence has been later suggested also on pleura, peritoneal and pericardial mesothelium [12,30,31,32,33]. Actually, by transmission electron microscopy, Wang [12] and Ohtsuka [32] found that colloidal iron stain of mesothelial glycocalyx is removed by neuraminidase treatment, which cleaves sialic acid from sialomucin. In line with the earlier suggestion of Andrews and Porter [11], that the polyanionic nature of the surface of the mesothelial glycocalyx may protect from friction, Ohtsuka et al. [32] proposed that the negative charges of sialomucin produce repulsive forces between facing serosal surfaces, and may, therefore reduce friction; nevertheless, little information seems available on the lubricant proprieties of sialomucins on mesothelial surfaces as well as on the spatial and functional relationships between mucopolysaccharides coat and phospholipids layers in these sites.
Recently, lubricant effect of sialomucin on pleural mesothelium was investigated by means of the technique previously described [8], in which measurements of the coefficient of kinetic friction (µ) were performed on pleural mesothelium blotted and rewetted while sliding at physiological velocities and loads. Data obtained showed that the addition of sialomucin (25 mg/mL) in Ringer solution after a standard mesothelial blotting with filter paper lowers µ more than the addition of Ringer solution only; this effect is such that after the addition of this macromolecule µ value is brought essentially back to its pre-blotting one [26]. Moreover, after washout with Ringer alone of the solution with sialomucin, µ value increased without reaching its preceding post-blotting value; hence, dilution and partial removal of the added macromolecule reduce the observed effect on µ. On this basis, µ of pleural mesothelium was measured after treatment of the specimens with neuraminidase, which cleaves sialic acid from the peptidic residue of sialomucin and should, therefore, increase µ of pleural mesothelium. After addition of neuraminidase (5 U/mL in Krebs phosphate solution at pH 7 for 60 or 90 min at 37 °C) and its washout (Krebs solution alone) µ of pleural mesothelium was not significantly greater than that of initial Krebs [34]. Despite this, neuraminidase was able to cleave sialic acid from pleural specimens, as shown by fluorimetric measurements of sialic acid in samples of Krebs phosphate solution at pH 7 with 5 U/mL of neuraminidase incubated for 90 min at 37 °C on the pleural surface of diaphragmatic specimens [34]. The amount of sialic acid cleaved by the enzyme was 0.066 mg/cm2 of the mesothelial surface of diaphragmatic specimens [34], while the amount of sialic acid cleaved by neuraminidase from the glycocalyx of rabbit vascular endothelium per unit of macroscopic surface has been found to be only 0.002 mg/cm2 [35]. Only part of this difference may be due to the lower enzymatic concentration used in the vascular endothelium. Moreover, Sironi et al. [34], provided fluorescent microscopy images of sections of pleura and underlying lung parenchima obtained from specimens preincubated for 90 min at 37 °C in Krebs phosphate at pH 7.0 without or with 5 U/mL of neuraminidase [34]. Sialic acid was stained with fluorescein-labeled lectin from Triticum vulgaris. Lectin labeling occurred only on control mesothelial surface: hence, neuraminidase removed most of the sialic acid bound to the sialomucin on the surface of the glycocalyx of pleural mesothelium. This hystochemical finding fits with the morphological findings of Wang [12,30] and Ohtsuka et al. [32] showing that sialomucin occurs on the surface of the mesothelial glycocalyx. On the other hand, it seems to contrast with the lack of increase in μ of the pleural mesothelium after neuraminidase treatment, despite the fact that this enzyme cleaves most of the sialic acid of the sialomucin occurring in the glycocalyx of pleural mesothelium of diaphragm specimens. This observation suggests that other molecules of the glycocalyx could contribute to the good lubrication of the pleural mesothelium.
To further investigate the role of sialomucin protein in pleural lubrication, the value of µ in rabbit pleural mesothelium was also measured after a protease treatment of specimens, in order to hydrolyze sialomucin and, therefore, increase µ of pleural mesothelium. Hence, µ was measured after a short treatment (so as to digest only the proteins of the glycocalyx surface without altering the mesothelial cells) with pronase, a broad spectrum protease that has been used to digest the glycocalyx of mouse capillary endothelium [36,37]. This short pronase treatment (0.1 mg/mL for 5 min) induced an increase of μ, which did not changed after enzyme washout with Ringer alone. If sialomucin (25 mg/mL) was added after pronase treatment, µ decreased to a value not significantly greater than that in initial Ringer [34]. Given the short period of treatment with pronase, its activity should be limited to the glycocalyx without affecting the mesothelial cells (see Section 4; [34,38]). Hence, the marked increase in μ produced by the addition of pronase should be mainly due to the digestion of the proteinic component of the glycoproteins of the glycocalyx.
2.2. Hyaluronan Lubricant Effects
The lubricant relevance of hyaluronan between sliding tissues has been known for a long time [27,29]. This macromolecule might also occur in the coat of the mesothelial surface, because it has been found in small concentration both in the pericardial [39], and pleural [16] liquid of rabbits. However, Ohtsuka et al. [32], like Wang [12], found that while neuraminidase removed colloidal iron staining of their mesothelium specimens, hyaluronidase did not affect this staining. Moreover, D’Angelo et al. [8], showed that addition of hyaluronan (2.5 mg/mL) in Ringer solution after mesothelial blotting with filter paper lowers µ of rabbit pleural mesothelium more than the addition of Ringer solution; this effect is such that after the addition of this macromolecule µ value is brought essentially back to its pre-blotting one [26]. Therefore, hyaluronan is able, like sialomucin, to restore a good boundary lubrication, even though Ohtsuka et al. [32] showed that the main component of pleural glycocalix is sialomucin. The marked decrease in µ obtained by the postblotting addition of lubricant macromolecules requires a concentration of sialomucin 10 times greater than that of hyaluronan. This is in line with the morphological researches of Wang [16,30] and Ohtsuka et al. [32] showing that in the coat covering the mesothelial surface the amount of sialomucin is much greater than that of hyaluronan. Finally, similar effects on blotted mesothelium are obtained by addition of a mixture of these macromolecules at half concentration, i.e., by simultaneous addition of sialomucin 12.5 mg/mL and hyaluronan 1.25 mg/mL [26].
In order to investigate the role of hyaluronan in pleural lubrication, µ of rabbit pleural mesothelium was measured after treatment of specimens with hyaluronidase, which hydrolyze hyaluronan and should, therefore, increase µ of pleural mesothelium [34]. The results obtained showed that µ values after addition of hyaluronidase (30 U/mL in Ringer solution at 37 °C) and after subsequent washout (Ringer solution alone) were not significantly greater than that of initial Ringer. The finding that μ of the pleural mesothelium does not increase after treatment with hyaluronidase is in line with the morphological findings of Wang [12,30] and Ohtsuka et al. [32] showing that hyaluronic acid does not occur on the surface of the glycocalyx of the mesothelium. Though hyaluronic acid does not seem to be located on the surface of the mesothelial glycocalyx, it is intriguing that its addition after pleural mesothelium blotting brings μ back to the control condition [26]. Moreover, the addition of hyaluronan (2.5 mg/mL) brings μ back to a value not significantly different from control even after pronase treatment [40]. The concentration of hyaluronic acid in rabbit pericardial liquid has been found to be 82 μg/mL [39], and that in pleural liquid ~1 μg/mL [16]. Both concentrations are much smaller than that required to bring μ back to its control value after mesothelial blotting and of that found in the synovial fluid (1–4 mg/mL; [41]), where the role of hyaluronan as a lubricant is important. Hence, even though hyaluronic acid acts as a good lubricant when applied to damaged mesothelium, its role as a lubricant of mesothelial surfaces under physiological conditions is not demonstrated. Recently Pecchiari et al. [42] showed that interposition of hyaluronan between blotted or normal mesothelium during 2 min of reciprocating sliding, limited or prevented the damage of the normal surface, as well as the increase of μ. In this model, hyaluronic acid shows therefore very good protective properties. In fact, reciprocating movements of a blotted mesothelium against a normal one wetted with Ringer caused substantial damage to the latter. This protective action could explain the increased secretion of hyaluronic acid into the pleural space that occurs when the reciprocating movements become potentially harmful to the mesothelium, such as high velocities and loads during hyperventilation [16], or localized pleural damages during inflammatory or neoplastic processes [43].
2.3. Phospolipids Lubricant Effects
In 1982, Hills et al. [44] showed that surface active phospholipids (phosphatidylcholine, sphingomyelin, and phosphatidylethanolamine) may be obtained from dog pleural space by washing it with Ringer solution. Similar phospholipids were also found on the surface of the pericardium [45]. Recently, Mills et al. [46,47] provided evidence that the percentage of various phosphatidylcolines (PCs) species in pleural lavage of dogs and cats are quite different from those in bronchoalveolar lavage. Moreover, the sample of pleural lavage of Mills et al. [46,47] was obtained quickly (30 s), without contact with air, while in the earlier research by Hills et al. [44], the sampling of pleural lavage was preceded by a pneumothorax. In the liquid from the bronchoalveolar lavage of Mills et al. [46,47], the main component was dipalmitoil-phosphatidylcoline (DPPC, 59% and 68% in dogs and cats, respectively), a saturated PC with strong surface action [48]. Instead, in the liquid from the pleural lavage the main component was stearoyl-linoleoyl-phosphatidylcholine (SLPC, 58% and 46%), an unsaturated PC absent in alveolar liquid; then, there were other unsaturated PCs: PLPC (17% and 31%) and POPC (16% and 13%), and a nearly negligible percentage of DPPC [46,47].
Transmission electron microscopy of samples of sheep visceral pleura, fixed with tannic acid and osmium tetroxide, showed 5–7 osmiophilic layers on the mesothelial surface, thus providing morphological evidence of phospholipid lamellar layers. Moreover, in samples of human and rat peritoneum, Dobbie and Anderson [49], and Michailova [33] provided electron microscopy evidence of several lamellar bodies among the microvilli, and of some superficial lamellar membranes. Unfortunately, no information is available on the spatial and functional relationships between these phospholipids layers and the mucopolysaccharides coat of the mesothelial surface glycocalyx under physiological conditions.
On the whole, the finding that phospholipids have been found in the liquid obtained by Ringer washing of dog or cat pleural space [44,46,47], and in dog pericardial liquid [45] deserve particular attention because they may provide good lubrication [50,51]. Furthermore, the high percentage of SLPC in the pleural lavage, and its lack in the bronchoalveolar lavage, leads to think that SLPC itself plus other unsaturated PCs may have important lubricating properties in pleural mesothelium. Actually, Hills [15,51] showed that the surface active phospholipids, extracted with chloroform-methanol from pleural washing, were good lubricant, particularly when several lamellar layers occur, and therefore may be important for lubrication of the pleural surfaces. Measurements of m were also performed on phospholipids extracted from joint surfaces with chloroform-methanol [51]. These extracts were placed on quartz plates, and after solvent evaporation, m was ~0.005. This extremely low value of m was attributed to the large number of phospholipid lamellar layers probably occurring in these experiments (the computed quantity of phospholipids per unit plate surface was 38 mg/cm2; [51]). Moreover, Hills [17] found that m of phospholipids extracted from the pleural surface with the same procedure used with joint extracts was 0.056, i.e., higher than that obtained with joint phospholipids [51]. This difference is likely due to the thinner layer of phospholipids with pleura extracts (the computed amount of phospholipids per unit plate surface was 5 times smaller than with joint extracts). Anyway, the value of m obtained with pleural extracts shows that surface active phospholipids may provide good lubrication on the mesothelial surfaces.
Later on Bodega et al. [26] showed that the addition for 2.5 h (time required for the formation of lamellar layers) of a mixture of phospholipids without unsaturated PCs [44] was able to reduce the marked increase in μ of the pleural mesothelium specimen occurring after its blotting with filter paper. The same experiment was repeated [38] using two different mixtures of phospholipids, one rich in unsaturated PCs (particularly SLPC) and one without unsaturated PCs; the increase in μ occurring after blotting with filter paper of the rabbit pleural mesothelium specimen was reduced of the same extent by a 2 h (time required for the formation of lamellar layers) addition of each of the two mixtures at the same concentration of 3 mg/mL. The finding that the small effect produced by the phospholipid mixture rich in unsaturated PCs is similar to that of the mixture rich in saturated PCs may be explained by the consideration of Bernhard et al. [52]. Namely, the production of dipalmitoil-phosphatidylcoline (DPPC, the saturated PC with strong surface action) is more complex and, hence, energetically more expensive than that of unsaturated PCs; as a consequence its production is limited to the tissue where a great surface-tension reducing properties is required, as the lung alveoli. In other words, the greater percentage of unsaturated PCs, and the lower percentage of DPPC, found in pleural than in alveolar lavage [46,47], is not due to a greater lubricating properties of the unsaturated PCs. On the other hand, the effect of PCs mixtures was small and does not increase by increasing their concentration from 3 to 6 mg/mL, while at a lower concentration (0.4 mg/mL) these mixtures do not produce an effect, suggesting that lubricating properties of PCs and of other phospholipids are small, likely because of the difficulty in resetting the functional relationships between phospholipids and the other lubricating molecules. Actually, phospholipids beyond a given concentration cannot react with the proteins or the mucopolysaccharides of the mesothelial glycocalyx and, hence, do not become water soluble [50]. In any case, the behavior of phospholipids is opposite to that of sialomucin, which increases its effect on the post-blotting increase in μ with the increase in its concentration until removal of the increase in μ [26,40].
In order to investigate the phospholipids role in lubrication, µ of rabbit pleural mesothelium was measured after treatment of specimens with phospholipase for a short time, so as to digest only the phospholipids occurring on the mesothelial glycocalyx without affecting those of the membranes of the mesothelial cells [38]. After the addition to the specimens of phospholipase C (7.5 U/mL, blocked after 10 min by EDTA and washed with Ringer), μ of the pleural mesothelium increased nearly 3 times. This increase was reduced by a 2 h addition of a phospholipid mixture rich in unsaturated PCs or in saturated PCs, both at a concentration of 3 mg/mL. The increase in µ of rabbit pleural mesothelium induced by a short phospholipase treatment was also reduced to a value not significantly different from that in initial Ringer by the addition of a solution with hyaluronan (2.5 mg/mL) or sialomucin (25 mg/mL). Therefore, in both series of experiments, the behavior of the two macromolecules was similar to that occurring in post-blotting Ringer [26], and after short pronase treatment [34]. The increase in µ after short phospholipase treatment provided the opportunity to check whether hyaluronan protects the phospholipids of the pleural mesothelium surface from the action of phospholipase, in analogy to the following findings obtained in other tissues: (a) hyaluronan [53] decreases alveolar surfactant inactivation by serum; (b) hyaluronan decreases the lysis of synovial phospholipids produced by phospholipase [54]; and (c) hyaluronan decreases alveolar surfactant inhibition caused by phospholipase [55]. Actually, if hyaluronan (2.5 mg/mL) was placed on the specimen before the short treatment with phospholipase, the increase in μ produced by this enzyme was smaller than that occurring without previous addition of hyaluronan. If phospholipase was incubated for 30 min with hyaluronan before its addition to the specimen, μ increased to an extent similar to that occurring with phospholipase not preincubated with hyaluronan. Therefore, the effect of hyaluronan described was not due to an inhibition of the enzyme caused by hyaluronan, but is a protection of the pleural phospholipids from the action of phospholipase, similar to that described for alveolar surface active phospholipids [53,55], and for synovial phospholipids [54]. Moreover, the small increase in μ observed after a short treatment with phospholipase in the presence of hyaluronan pre-treatment was removed by the addition of hyaluronan, similarly to what occurred for the increases induced by blotting [26], short pronase treatment [34], and short phospholipase treatment [38]. Finally, surfactant phospholipids might also occur in other parts of the glycocalyx, as suggested by lamellar layers in the glycocalyx of peritoneal mesothelium [49]. Actually, they may interact with hyaluronan [51,56], and this could occur under physiological conditions. In any case, mucopolisaccharides seem to be the main and more direct lubricating factors of pleural mesothelium under control and damaged conditions [26,32,38,40].
3. Mixed Lubrication in Damaged Mesothelium
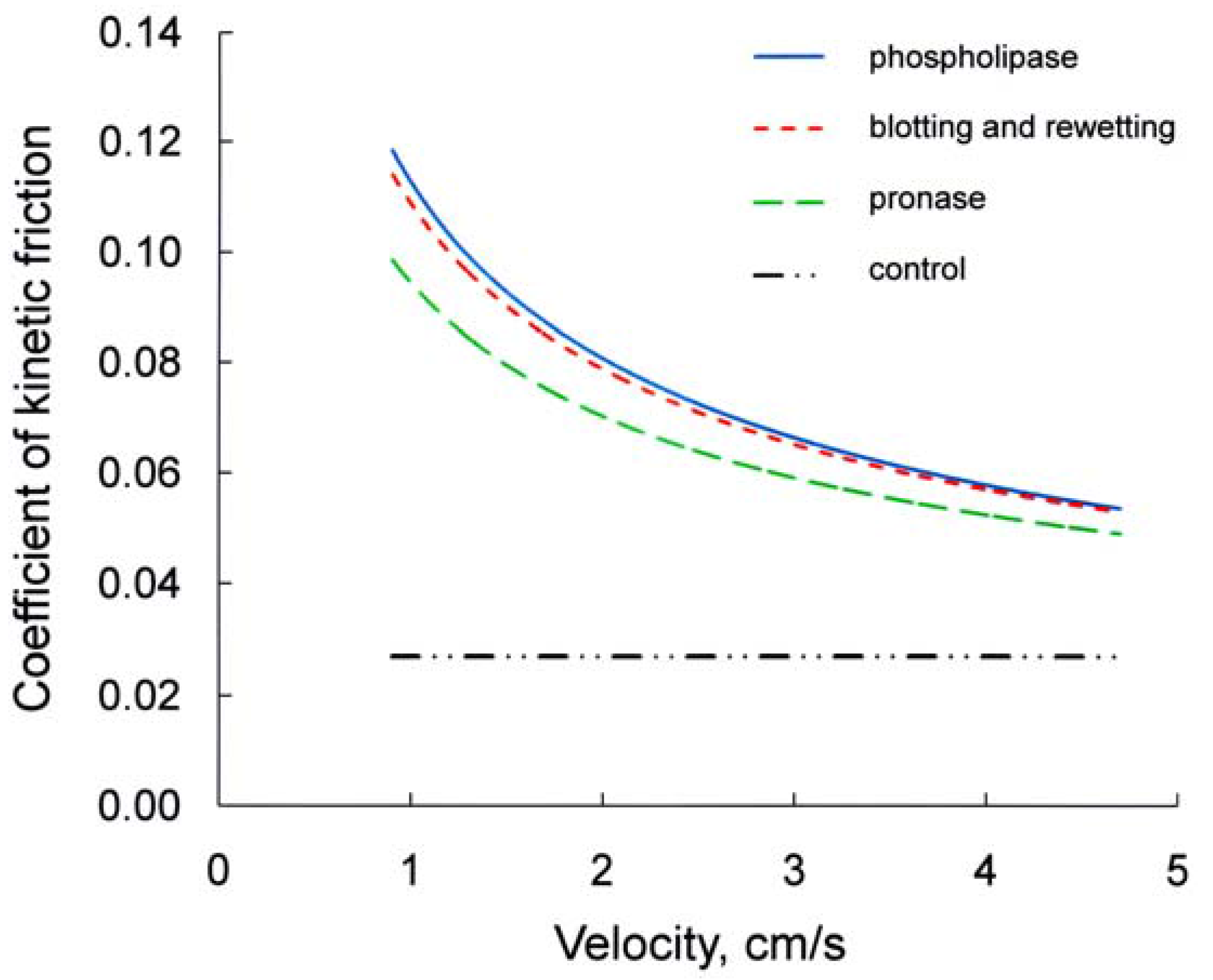
D’Angelo et al. [8], when measuring the coefficient of kinetics friction (µ) between specimens of rabbit visceral and parietal pleura during oscillatory sliding in vitro at physiological velocities and loads, found that µ is directly proportional to the load and independent of sliding velocity and contact area (Figure 2). The finding that µ does not change with changes in sliding velocity is consistent with boundary lubrication (see Introduction). Instead, in specimens that have been blotted and wetted with Ringer solution µ is much higher and decreases markedly with the increase in velocity (Figure 2) [40]. Moreover, despite the marked decrease occurred with the increase in sliding velocity, µ in post-blotting Ringer at highest velocity (4.7 cm/s) is still greater than in pre-blotting Ringer, which is independent from sliding velocity [8,26]. The addition of sialomucin (25 mg/mL) or hyaluronan (2.5 mg/mL) in Ringer after a standard blotting of the mesothelium brings µ essentially back to its pre-blotting value, which does not change when the sliding velocity is increased [8,26]. After the addition of smaller concentrations of sialomucin (8 mg/mL) or hyaluronan (0.8 mg/mL), µ at each velocity is markedly lower than that without the addition of these macromolecules, and this difference is greater al low velocity. This finding indicates a regime of mixed lubrication in post-blotting Ringer, at variance with boundary lubrication occurring before blotting or postblotting with sufficient macromolecule addition. Actually, after mesothelial blotting the pleural surface appears damaged (see Section 4; [26,42]), consequently the mesothelial surface is likely much more rough than it was under control conditions and, after the addition of Ringer, the amount of liquid between the opposed mesothelial surfaces in several points should be greater than that before blotting. Therefore, when the visceral and parietal pleura are made to slide under physiological load, the shear induced hydrodynamic pressure, besides reducing the roughness of the mesothelial surface [5], generates some areas of elastohydrodynamic lubrication, which increase with the increase in sliding velocity, while those of boundary lubrication decrease [40]. Hence, in post-blotting Ringer a regime of mixed lubrication might occur. Furthermore, as a result of loss of part of the boundary lubricant, the friction in the contact area should be markedly increased, but this component should decrease with the increase in sliding velocity, owing to the decrease in contact area. Since contact area between the opposed mesothelia should still occur in post-blotting Ringer, the addition of a small concentration of lubricant macromolecules restores boundary lubrication.
Similar considerations could be applied to phospoliphase and pronase treatment (Figure 2). After short phospholipase C treatment of the specimens, μ increases ~3 times. This increase diminishes with the increase in sliding velocity (from 0.9 to 4.7 cm/s), and at the highest velocity it is still greater than that in initial Ringer [40]. Hence, the lubrication shifts from boundary to mixed. The addition of sialomucin (25 mg/mL) or hyaluronan (2.5 mg/mL) brings μ back to a value not significantly different from the control one and, hence, to a condition of boundary lubrication which is independent from sliding velocity [8,26,40]. After short pronase treatment of the specimens, µ doubles. This increase diminishes markedly with the increase in sliding velocity [34], and at the highest velocity it is still greater than that in initial Ringer, which is independent of sliding velocity [8,26,40]. Hence, also in this case there is a regimen of mixed lubrication (boundary and elasto-hydrodynamic). The addition of sialomucin or hyaluronic acid after phospholipase or pronase treatment bring μ back to control value and, hence, to a condition of boundary lubrication [8,26,40].
4. Hystological Remarks
Andrews and Porter [11] by transmission electron microscopy showed a coat, probably acid mucopolysaccharide in nature, entrapped among the microvilli of rat pleura and peritoneal mesothelium. They pointed out that this coat should be important for lubrication of the mesothelial surfaces. Moreover, staining with ruthenium red, they showed many fine polyanionic strands radiating from the sides of microvilli, interconnected with each other and adjacent microvilli. Finally, evidence has been provided by transmission electron microscopy of strip-like structures covering the granulo-filamentous material surrounding the microvilli of rat pericardium [33]. Wang [12] by transmission electron microscopy noticed that colloidal iron stains the surface of rabbit pleural mesothelium, and that this staining persists after hyaluronidase treatment, but is abolished by neuroaminidase treatment. Later on, he pointed out that this finding indicates the presence of sialomucin and other mucoproteins in the glycocalyx of the pleural mesothelium [30]. More recently, Ohtsuka et al. [32] with either cationic colloidal iron or Limax flavus lectin showed a coat of sialomucin with many negative charges on the mesothelial surface of mouse pleura, pericardium, and peritoneum. This coat is much thicker than the usual glycocalyx occurring on cell plasma membrane. In line with the proposal of Andrews and Porter [11], Ohtsuka et al. [32] concluded that the negative charges of sialomucin produce repulsive forces between facing serosal surfaces, and may, therefore, reduce friction. In line with these observations, Sironi et al. [34] showed with Triticum vulgaris lectin a coat of sialomucin on the mesothelial surface of rabbit pleura (see Section 2). Moreover, D’Angelo et al. [8] and Bodega et al. [26] by transmission electron microscopy (TEM) did not find differences between control specimens and specimens used to measure the coefficient of kinetic friction in physiological solution. Conversely, TEM micrographs show that, after mesothelial blotting with filter paper, pleural surface was damaged to various extent, showing areas where microvilli were partially or largely removed from the mesothelium surface [26] or even areas with substantial mesothelial damage with the disappearance of the cellular layer [42]. Anyway, the increase in µ after blotting should not be related to the removal of microvilli, because Pecchiari et al. [42] showed that the superficial density of microvilli and their characteristics are not determinants of the frictional forces which oppose sliding of normal mesothelial surfaces under physiological conditions, nor of the lubricant effect of hyaluronic acid. On the other hand, the mesothelium damage is consistent with a substantial loss of part of the coat of macromolecules normally entrapped among them at the mesothelial surface [11].
After, mesothelial blotting with filter paper µ increases markedly [8,26]. This finding demonstrates the relevance for pleural lubrication of the macromolecules of the thick coat covering mesothelial surface, and make even more remarkable the finding that postblotting addition of sialomucin or hyaluronan is able to restore µ to its preblotting value and, hence, to restore good lubrication in damaged mesothelium [26]. Moreover, by staining pleural specimens with silver nitrate under control condition or after short treatment with phospholipase or pronase, Bodega et al. [38] investigated whether mesothelial cells were disrupted by these enzymatic treatments. Cells were not disrupted by short phospholipase treatment: hence, the action of the enzyme during this treatment should involve only the phospholipids occurring among the mucopolysaccharides of the mesothelial glycocalyx, without affecting those of the membranes of the mesothelial cells. Instead, after pronase treatment a few cells were disrupted. It is noteworthy that hyaluronan or sialomucin are able to restore μ to its control value, despite the disruption of ~60% of the mesothelium [38]. Considering that sialomucin is normally occurring in the glycocalyx of the mesothelial cells, the above finding suggests that it might be worthwhile to investigate whether sialomucin may be useful in the cases of thoracotomy that require prevention of pleural adhesion. Experiments in this field have been done in rats by using hyaluronate based bioresorbable membranes [57], hemostatic fleeces with a collagen matrix coated with fibrinogen and thrombin [58] or poly-ethylene glycol membranes [59]. Furthermore, peritoneal adhesions have been reduced by a peritoneal application of phospholipids in rats [60] or a peritoneal spray of phospholipids in rabbits [61]. Hence, a mixture of sialomucin and phospholipids injected into the pleural space might provide a more efficient and less invasive method to prevent pleural adhesion.
5. Conclusions
Efficient ventilation requires that pleural surfaces slide against each other with minimal friction, thereby preventing mesothelial damage. The finding that, in physiological conditions, the coefficient of friction (µ) does not change with changes in sliding velocity is consistent with a regime of boundary lubrication, and therefore is in line with the model which assumes contact between pleural surfaces [8].
Pleural boundary lubrication can be influenced by the presence in the pleural liquid and on the mesothelial surface of several specialized molecules. Well known lubricating molecules like sialomucins, hyaluronan, or phospholipids have been proposed as boundary lubricants in pleural mesothelium. Anyway, the identification of the lubricating substances has been hindered by the lack of knowledge of the composition of the mesothelial coating and its organization. Actually, the presence of mucoproteins like sialomucins has been shown on the surface of the pleural mesothelial glycocalyx [30,32,34], while phospholipids have been obtained from pleural space washing [44,46,47]. Unfortunately, no information is available on the spatial and functional relationships between these phospholipids layers and the mucopolysaccharides coat of the mesothelial surface glycocalyx under physiological conditions. Finally, hyaluronic acid has been found in small concentration in pleural liquid [16], but it does not occur on the surface of the glycocalyx of the mesothelium [12,30,32]. This is in line with the finding that μ of the pleural mesothelium does not increase after treatment with hyaluronidase, while a marked increase in μ is induced by a short treatment with pronase, that should mainly digest the proteinic component of the glycoproteins of the glycocalyx. An increase in μ is also observed after a short treatment with phospholipase, so as to digest only the phospholipids occurring on the mesothelial glycocalyx without affecting those of the membranes of the mesothelial cells [38].
When pleural surface is damaged to various extent by blotting or short treatment with pronase or phospholipase [26,38,42], with consequent substantial loss of part of the coat of macromolecules present at the mesothelial surface [11] and even of the mesothelial cells [42], µ increases markedly [8,26,42]. Moreover, this increase decreases with increasing velocity, suggesting that in damaged mesothelium lubrication regimen turns from boundary to mixed (boundary and elasto-hydrodynamic). In these conditions, even with a disruption of ~60% of the mesothelium [38], addition of sialomucin or hyaluronan brings μ back to its control value and, hence, restores a condition of boundary lubrication which is independent from sliding velocity [8,26,40]. On the other hand, addition of phospholipids (either saturated or unsaturated) produces a small effect relative to that of hyaluronan or sialomucin.
These findings demonstrate the relevance for pleural lubrication of the macromolecules of the thick coat covering mesothelial surface. In particular, considering that sialomucin is normally occurring at the surface of the pleural mesothelial cells, where is the main component of the glycocalyx [32], mucopolysaccharides seem to be the main lubricating factors of pleural mesothelium under physiological and damaged conditions [26,32,38,40]. On the other hand, the small lubricating properties of added phospholipids are likely due to the difficulty in resetting the functional relationships with the other lubricating molecules on mesothelial surface. Finally, even though hyaluronic acid acts as a good lubricant when applied to damaged mesothelium [26], it does not seem to be located on the surface of the mesothelial glycocalyx and its role as a lubricant of mesothelial surfaces under physiological conditions is not demonstrated.
On this basis, it might be worthwhile to investigate whether sialomucin may be useful in the cases of thoracotomy that require prevention of pleural adhesion, as a mixture of sialomucin and phospholipids injected into the pleural space might provide an efficient method to prevent pleural adhesion.
Acknowledgments
The authors thank Roberto Galli for his skillful technical assistance.
Author Contributions
All authors equally contributed to the overall organization, the associated research, the writing and the editing of this review article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mariassy, A.T.; Wheeldon, E.B. The pleura: A combined light microscopic, scanning and trasmission electron microscopic study in the sheep. I. Normal pleura. Exp. Lung Res. 1983, 4, 293–314. [Google Scholar] [CrossRef] [PubMed]
- Zocchi, L. Physiology and pathophysiology of pleural fluid turnover. Eur. Respir. J. 2002, 20, 1545–1558. [Google Scholar] [CrossRef] [PubMed]
- Lai-Fook, S.J.; Rodarte, J.R. Pleural pressure distribution and its relationship to lung volume and interstitial pressure. J. Appl. Physiol. 1991, 70, 967–978. [Google Scholar] [PubMed]
- Lai, J.; Gouldstone, A.; Butler, J.P.; Federspiel, W.J.; Loring, S.H. Relative motion of the lung and chest wall promotes uniform pleural space thickness. Respir. Physiol. Neurobiol. 2002, 131, 233–243. [Google Scholar] [CrossRef]
- Gouldstone, A.; Brown, R.E.; Butler, J.P.; Loring, S.H. Elastohydrodynamic separation of pleural surfaces during breathing. Respir. Physiol. Neurobiol. 2003, 137, 97–106. [Google Scholar] [CrossRef]
- Agostoni, E.; D’Angelo, E. Pleural liquid pressure. J. Appl. Physiol. 1991, 71, 393–403. [Google Scholar] [PubMed]
- Loring, S.H.; Brown, R.E.; Gouldsone, A.; Butler, J.P. Lubrication regimes in mesothelial sliding. J. Biomech. 2005, 38, 2390–2396. [Google Scholar] [CrossRef] [PubMed]
- D’Angelo, E.; Loring, S.H.; Gioia, M.E.; Pecchiari, M.; Moscheni, C. Friction and lubrication of pleural tissues. Respir. Physiol. Neurobiol. 2004, 142, 55–68. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Butler, J.P.; Loring, SH. Influence of the softness of the parietal pleura on respiratory sliding mechanisms. Respir. Physiol. Neurobiol. 2011, 177, 114–119. [Google Scholar] [CrossRef] [PubMed]
- Murphy, B.G.; Macklem, P.T. Stress at the pleural surface. Respir. Physiol. 1976, 28, 65–74. [Google Scholar] [CrossRef]
- Andrews, P.M.; Porter, K.R. The ultrastructural morphology and possible functional significance of mesothelial microvilli. Anat. Rec. 1973, 177, 409–426. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.S. The regional difference of pleural mesothelial cells in rabbits. Am. Rev. Respir. Dis. 1974, 110, 623–633. [Google Scholar] [PubMed]
- Michailova, K.N. Ultrastructural observations on the human visceral pleura. Eur. J. Morphol. 1997, 35, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Agostoni, E. Mechanics of the pleural space. Physiol. Rev. 1972, 53, 57–128. [Google Scholar]
- Hills, B.A. Graphite-like lubrication of mesothelium by oligolamellar pleural surfactant. J. Appl. Physiol. 1992, 73, 1034–1039. [Google Scholar] [PubMed]
- Wang, P.M.; Lai-Fook, S.J. Effects of ventilation on hyaluronan and protein concentration in pleural liquid in anesthetized and conscious rabbits. Lung 1998, 176, 309–324. [Google Scholar] [CrossRef] [PubMed]
- Ogston, A.G.; Stainer, J.E. The physiological function of hyaluronic acid in synovial fluid; viscous, elastic and lubricant properties. J. Physiol. 1953, 119, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Agostoni, E.; D’Angelo, E.; Roncoroni, G. The thickness of the pleural liquid. Respir. Physiol. 1968, 5, 1–13. [Google Scholar] [CrossRef]
- Agostoni, E. Mechanics of the pleural space. In Handbook of Physiology. The Respiratory System. Mechanics of Breathing; Macklem, P.T., Mead, J., Eds.; American Physiological Society: Bethesda, MD, USA, 1986; Volume 3, pp. 531–559. [Google Scholar]
- Agostoni, E.; Zocchi, L. Pleural liquid and its exchange. Respir. Physiol. Neurobiol. 2007, 159, 311–323. [Google Scholar] [CrossRef] [PubMed]
- Lai-Fook, S.J.; Beck, K.C.; Southorn, P.A. Pleural liquid pressure measured by micropipettes in rabbits. J. Appl. Physiol. 1984, 56, 1633–1639. [Google Scholar] [PubMed]
- Agostoni, E.; Agostoni, P.G.; Zocchi, L. Pleural liquid pressure in the zone of apposition and in the lung zone. Respir. Physiol. 1989, 75, 357–370. [Google Scholar] [CrossRef]
- Lai-Fook, S.J.; Wang, P.M. Dynamics of pleural liquid. In Complexity in Structure and Function of the Lung; Hlastala, M.P., Robertson, H.T., Eds.; Dekker: New York, NY, USA, 1998; pp. 123–149. [Google Scholar]
- Lai-Fook, S.J. Pleural mechanics and fluid exchange. Physiol. Rev. 2004, 84, 385–410. [Google Scholar] [CrossRef] [PubMed]
- Von Neergaard, K. Zur Frage des Druckes im Pleuraspalt. Beitr. Klin. Erforsch. Tuberk. Lungenkr. 1927, 65, 476–485. [Google Scholar] [CrossRef]
- Bodega, F.; Pecchiari, M.; Sironi, C.; Porta, C.; Arnaboldi, F.; Barajon, I.; Agostoni, E. Lubricating effect of sialomucin and hyaluronan on pleural mesothelium. Respir. Physiol. Neurobiol. 2012, 180, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Frazer, J.R.; Laurent, T.C. Hyaluronan. In Extracellular Matrix; Comper, W.D., Ed.; Harwood Academic Publishers: Amsterdam, The Netherlands, 1996; Volume 2, pp. 141–169. [Google Scholar]
- Schmidt, T.A.; Gastelum, N.S.; Nguyen, Q.T.; Schumacher, B.L.; Sah, R.L. Boundary lubrication of articular cartilage: Role of synovial fluid constituents. Arthritis Rheum. 2007, 56, 882–891. [Google Scholar] [CrossRef] [PubMed]
- Linn, F.C. Lubrication of animal joints. II. The mechanism. J. Biomech. 1968, 1, 193–205. [Google Scholar] [CrossRef]
- Wang, N.S. Mesothelial cells in situ. In The Pleura in Health and Disease; Chrétien, J., Bignon, J., Hirsh, A., Eds.; Dekker: New York, NY, USA, 1985; pp. 23–42. [Google Scholar]
- Hilkens, J.; Ligtenberg, M.J.L.; Vos, H.L.; Litvinov, S.V. Cell membraneassociated mucins and their adhesion-modulating property. Trends Biochem. Sci. 1992, 17, 359–363. [Google Scholar] [CrossRef]
- Ohtsuka, A.; Yamana, S.; Murakami, T. Localization of membrane-associated sialomucin on the free surface of mesothelial cells of the pleura, pericardium and peritoneum. Histochem. Cell Biol. 1997, 107, 441–447. [Google Scholar] [CrossRef] [PubMed]
- Michailova, K.N. Mesothelial lamellar bodies in norm and experimental conditions. Transmission and scanning electron microscopic observations on the peritoneum, pleura and pericardium. Anat. Embryol. 2004, 208, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Sironi, C.; Bodega, F.; Porta, C.; Agostoni, E. Pleural mesothelium lubrication after hyaluronidase, neuraminidase or pronase treatment. Respir. Physiol. Neurobiol. 2013, 188, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Born, G.V.; Palinski, W. Unusually high concentrations of sialic acids on the surface of vascular endothelia. Br. J. Exp. Pathol. 1985, 66, 543–549. [Google Scholar] [PubMed]
- Simionescu, M.; Simionescu, N.; Silbert, J.E.; Palade, G.E. Differentiated microdomains on the luminal surface of the capillary endothelium. II. Partial characterization of their anionic sites. J. Cell Biol. 1981, 90, 614–621. [Google Scholar] [CrossRef] [PubMed]
- Simionescu, M.; Simionescu, N.; Santoro, F.; Palade, G.E. Differentiated microdomains of the luminal plasmalemma of murine muscle capillaries: Segmental variations in young and old animals. J. Cell Biol. 1985, 100, 1396–1407. [Google Scholar] [CrossRef] [PubMed]
- Bodega, F.; Sironi, C.; Porta, C.; Zocchi, L.; Agostoni, E. Pleural mesothelium lubrication after phospholipase treatment. Respir. Physiol. Neurobiol. 2014, 194, 49–53. [Google Scholar] [CrossRef] [PubMed]
- Honda, A.; Ohashi, Y.; Mori, Y. Hyaluronic acid in rabbit pericardial fluid and its production by pericardium. FEBS Lett. 1986, 203, 273–278. [Google Scholar] [CrossRef]
- Bodega, F.; Sironi, C.; Porta, C.; Pecchiari, M.; Zocchi, L.; Agostoni, E. Mixed lubrication after rewetting blotted mesothelium. Respir. Physiol. Neurobiol. 2013, 185, 369–373. [Google Scholar] [CrossRef] [PubMed]
- Castor, C.W.; Prince, R.K.; Hazelton, M.J. Hyaluronic acid in human synovial effusions; A sensitive indicator of altered connective tissue cell function during inflammation. Arthritis Rheum. 1966, 9, 783–794. [Google Scholar] [CrossRef] [PubMed]
- Pecchiari, M.; Sartori, P.; Conte, V.; D’Angelo, E.; Moscheni, C. Friction and morphology of pleural mesothelia. Respir Physiol Neurobiol. 2016, 220, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Pettersson, T.; Fröseth, B.; Riska, H.; Klockars, M. Concentration of hyaluronic acid in pleural fluid as a diagnostic aid for malignant mesothelioma. Chest 1988, 94, 1037–1039. [Google Scholar] [CrossRef] [PubMed]
- Hills, B.A.; Butler, B.D.; Barrow, R.E. Boundary lubrication imparted by pleural surfactants and their identification. J. Appl. Physiol. 1982, 53, 463–469. [Google Scholar] [PubMed]
- Hills, B.A.; Butler, B.D. Phospholipids identified on the pericardium and their ability to impart boundary lubrication. Ann. Biomed. Eng. 1985, 13, 573–586. [Google Scholar] [CrossRef] [PubMed]
- Mills, P.C.; Chen, Y.; Hills, Y.C.; Hills, B.A. Differences in surfactant lipids collected from pleural and pulmonary lining fluids. Pharm. Res. 2005, 22, 1926–1930. [Google Scholar] [CrossRef] [PubMed]
- Mills, P.C.; Chen, Y.; Hills, Y.C.; Hills, B.A. Comparison of surfactant lipids between pleural and pulmonary lining fluids. Pulm. Pharmacol. Ther. 2006, 19, 292–296. [Google Scholar] [CrossRef] [PubMed]
- Brown, E.S. Isolation and assay of dipalmitoyl lecithin in lung extracts. Am. J. Physiol. 1964, 207, 402–406. [Google Scholar] [PubMed]
- Dobbie, J.W.; Anderson, J.D. Ultrastructure, distribution, and density of lamellar bodies in human peritoneum. Perit. Dial. Int. 1996, 16, 482–487. [Google Scholar] [PubMed]
- Hills, B.A. The Biology of Surfactant; Cambridge University Press: Cambridge, UK, 1988. [Google Scholar]
- Hills, B.A. Oligolamellar lubrication of joints by surface active phospholipid. J. Rheumatol. 1989, 16, 82–91. [Google Scholar] [PubMed]
- Bernhard, W.; Postle, A.D.; Rau, G.A.; Freihorst, J. Pulmonary and gastric surfactants. A comparison of the effect of surface requirements on function and phospholipid composition. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2001, 129, 173–182. [Google Scholar] [CrossRef]
- Lu, K.W.; Goerke, J.; Clements, J.A.; Taeusch, H.W. Hyaluronan decreases surfactant inactivation in vitro. Pediat. Res. 2005, 57, 237–241. [Google Scholar] [CrossRef] [PubMed]
- Nitzan, D.W.; Nitzan, U.; Dan, P.; Yedgar, S. The role of hyaluronic acid in protecting surface-active phospholipids from lysis by exogenous phospholipase A(2). Rheumatology 2001, 40, 336–340. [Google Scholar] [CrossRef] [PubMed]
- Iwanicki, J.L.; Lu, K.W.; Taeusch, H.W. Reductions of phospholipase A(2) inhibition of pulmonary surfactant with hyaluronan. Exp. Lung Res. 2010, 36, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Pasquali-Ronchetti, I.; Quaglino, D.; Mori, G.; Bacchelli, B.; Ghosh, P. Hyaluronan-phospholipid interactions. J. Struct. Biol. 1997, 120, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, A.; Abe, T.; Matsuura, A. Prevention of postoperative intrapleural adhesion of the thoracotomy incision by a bioresorbable membrane in the rat adhesion model. Ann. Thorac. Cardiovasc. Surg. 2000, 6, 151–160. [Google Scholar] [PubMed]
- Getman, V.; Devyatko, E.; Wolner, E.; Aharinejad, S.; Mueller, M.R. Fleece bound sealing prevents pleural adhesions. Interact. Cardiovasc. Thorac. Surg. 2006, 5, 243–246. [Google Scholar] [CrossRef] [PubMed]
- Karacam, V.; Onen, A.; Sanli, A.; Gurel, D.; Kargi, A.; Karapolat, S.; Ozdemir, N. Prevention of pleural adhesions using a membrane containing polyethylene glycol in rats. Int. J. Med. Sci. 2011, 8, 380–386. [Google Scholar] [CrossRef] [PubMed]
- Ar’Rajab, A.; Snoj, M.; Larsson, K.; Bengmark, S. Exogenous phospholipid reduces postoperative peritoneal adhesions in rat. Eur. J. Surg. 1995, 161, 341–344. [Google Scholar] [PubMed]
- Bhandarkar, D.S.; Nathanson, L.K.; Hills, B.A. Spray of phospholipid powder reduces peritoneal adhesions in rabbits. Aust. N. Z. J. Surg. 1999, 69, 388–390. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic Stribeck curve showing coefficient of kinetic friction (µ) as a function of sliding velocity in the three different lubrication regimens.
Figure 1.
Schematic Stribeck curve showing coefficient of kinetic friction (µ) as a function of sliding velocity in the three different lubrication regimens.

Figure 2.
Relationship between coefficient of kinetic friction and sliding velocity obtained under the following conditions: after phospholipase C treatment, after mesothelial blotting and rewetting with Ringer, after short pronase treatment, and at control.
Figure 2.
Relationship between coefficient of kinetic friction and sliding velocity obtained under the following conditions: after phospholipase C treatment, after mesothelial blotting and rewetting with Ringer, after short pronase treatment, and at control.

© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Porta, C.; Sironi, C.; Bodega, F.; Agostoni, E. Pleural Lubrication. Lubricants 2016, 4, 15. https://doi.org/10.3390/lubricants4020015
AMA Style
Porta C, Sironi C, Bodega F, Agostoni E. Pleural Lubrication. Lubricants. 2016; 4(2):15. https://doi.org/10.3390/lubricants4020015
Chicago/Turabian StylePorta, Cristina, Chiara Sironi, Francesca Bodega, and Emilio Agostoni. 2016. "Pleural Lubrication" Lubricants 4, no. 2: 15. https://doi.org/10.3390/lubricants4020015
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.