
Somatostatin-Dopamine Chimeric Molecules in Neuroendocrine Neoplasms



1
Department of Medical Biotechnologies and Translational Medicine, University of Milan, 20129 Milan, Italy
2
IRCCS, Laboratory of Geriatric and Oncologic Neuroendocrinology Research, Istituto Auxologico Italiano, 20095 Cusano Milanino, Italy
*
Author to whom correspondence should be addressed.
J. Clin. Med. 2021, 10(3), 501; https://doi.org/10.3390/jcm10030501
Submission received: 22 December 2020
/
Revised: 25 January 2021
/
Accepted: 26 January 2021
/
Published: 1 February 2021
(This article belongs to the Section Endocrinology & Metabolism)
Abstract
:Neuroendocrine neoplasms (NENs) are a widely heterogeneous family of neoplasms arising from neuroendocrine cells, which are interspersed throughout the body. Despite NENs are relatively rare, their incidence and prevalence are constantly increasing probably due to the improvement in earlier diagnosis and patients’ management. When surgery is not curative, particularly for patients with metastatic disease, several medical options are available. Somatostatin analogues (SSA) are the first-line medical therapy for well-differentiated NENs. Interestingly, the heterodimerization of somatostatin receptors (SSTs) with dopamine receptors (DRs) has been discovered in NENs. This phenomenon results in hybrid receptors with enhanced functional activity. On these bases, chimeric molecules embracing somatostatin and dopamine features have been recently developed. The aim of this review is to provide a comprehensive overview of the available preclinical and clinical data regarding chimeric somatostatin-dopamine agonists as a new class of “magic bullet” in the therapy of NENs.
1. Introduction
Neuroendocrine neoplasms (NENs) are a rare family of tumors that arise from neuroendocrine cells. These cells are widely dispersed all over the human body and their main function is hormone secretion. NENs can seriously compromise organs homeostasis and quality of life of patients due to the excessive secretion of hormones able to generate disabling symptoms, specific syndromes and other complications [1,2,3]. Somatostatin and dopamine play a relevant role in the regulation of hormone secretion in several neuroendocrine cells. Therefore, both somatostatin and dopamine analogs have been successfully tested for their high anti-secretory activity in symptomatic patients with NENs. Moreover, emerging evidence of antiproliferative activity of somatostatin analogues (SSAs) was provided by two clinical trials PROMID and CLARINET [4,5]. SSAs are the first-line medical therapy for well-differentiated NENs [6]. Due to the robust effect on prolactin (PRL) and growth hormone (GH) secretion inhibition, also dopamine agonists (DAs) were intensively studied in NENs [7,8]. Moreover, the relevant role of both somatostatin and dopamine systems is currently exploited for the diagnosis (68Ga-DOTATATE and 18F-DOPA PET) and peptide receptor radionuclide therapy (177Lu-DOTATE) of metastatic and inoperable NENs [6]. The effects of SSAs are mediated by several somatostatin receptors (SSTs) [9]. Analysis on expression pattern of both SSTs and dopamine receptors (DRs) were performed in several NENs showing their co-expression. Interestingly, formation of heterodimers between SSTs and DRs was found to be associated with an enhanced activity bringing new insight for NENs treatment [10]. Speculating on these data, chimeric molecules harboring somatostatin and dopamine features have been developed [11].
In this review, we provide an overview concerning the complex interplay between somatostatin and dopamine signaling, analyzing the potential clinical application of chimeric somatostatin-dopamine molecules as “magic bullet” in the therapy of NENs.
2. Somatostatin System in NENs
Somatostatin is a small regulatory peptide that can explicit a broad spectrum of effects including maintenance of homeostasis or involvement in pathological events, such as inflammation, through interactions with specific SSTs. Its physiological function is mainly related to inhibition of exocrine and endocrine secretion. Notably, in the central nervous system somatostatin acts as a neurotransmitter/neuromodulator playing an important role in learning and memory. In addition, the inhibitory effects of native somatostatin on hormone release and cell proliferation made this molecule a potential candidate as anticancer drug, particularly in NENs expressing SSTs. SSTs belong to the G-protein coupled receptor superfamily of which five human different receptors subtypes (SST1–SST5) have been identified [12] and they are encoded by five genes. SST1, SST3, and SST4 genes are introneless, consequently they do not undergo to splicing events, while for SST2 and SST5 genes splicing variants are known. Truncated variants, resulting from splicing events of SST2 and SST5 genes, display an abnormal functional response to somatostatin ligands. In particular, the truncated form of SST5 (sst5TMD4), which is mainly expressed in medullary thyroid carcinoma cell lines, non-functioning pituitary adenomas (NFPAs), somatotropinomas, and pancreatic neuroendocrine tumors, is associated with a more aggressive tumor behavior and decreased response to SSAs [13,14,15,16,17,18].
Somatostatin binds its receptor at transmembrane domain level and, based on the receptor subtype, it provides several down-streaming signals through activation of different G-proteins (Figure 1). Notably, SSTs are mainly bound to inhibitory G proteins (Gi/G0), which after ligand binding are able to decrease the activity of adenylyl cyclase, phopholipase-C stimulation and MAPK modulation through phosphotyrosine phosphatase (PTP) activation. SST2 has been reported to be coupled mainly with protein-Gα02/β2/γ3, which can activate inwardly rectifying K+ channels and/or inhibit voltage-dependent Ca2+ channel, providing an inhibition of secretion through hyperpolarization of membrane potential [19]. On the other hand, SST3 is coupled with Gα12 or Gα14 activating pertussis toxin-independent mechanisms. All this evidence underlines that the activated signaling cascade depends on the SST subtype. Not only different subtypes of SSTs but also the nature of the ligand can affect tissue response [20]. Indeed SSAs, due to their various binding affinities, can stabilize specific conformations of the same receptor subtype leading to different cell response [21,22].
The main anti-tumor signaling pathways of SSTs are summarized in Figure 1. The antitumor features of somatostatin system involve direct and indirect mechanisms. Direct mechanisms are associated with cell cycle arrest and/or activation of apoptosis, whereas indirect mechanisms involve the inhibition in secretion of several growth and proangiogenic factors. The first anti-proliferative mechanism, to be associated to somatostatin signaling in NENs, involves PTPs, such as SH2 domain-containing cytosolic tyrosine phosphatase-1/2 [23,24]. PTPs activity increases after both native somatostatin and SSAs treatments, inducing dephosphorylation of several receptor tyrosine kinases leading to inhibition of their signaling [25,26,27]. PTP acts inhibiting MAPK-ERK1/2 signaling, which is one of the most important cell proliferation pathways. In particular, SST1, SST2, SST4, and STT5 have been shown to be associated with cell cycle arrest in G1/S phase, while SST3 induced a block in G2M phase [28]. Finally, PTP interaction with PI3K/Akt pathway results in up-regulation of p21 and p27 cyclin dependent kinase inhibitors as well as the tumor suppression gene Zac1 [29,30]. These anti-tumor mechanisms are mainly triggered by SST2 and SST3, which can also induce apoptosis through p-53/Bax, NFkB and PTPs/caspase8 signaling [29,31,32,33,34,35]. In addition, SST1, SST3, and SST4 can directly mediate antiproliferative effects through inhibition of the Na+/H+ exchanger, especially in enteric endocrine cells.
SSTs signals act not only directly by dephosphorylation of growth factors receptors but also indirectly through down-regulation of their synthesis [36]. After treatment with SSAs, the decreased intracellular cAMP concentration leads to downregulation of several factors and hormones, involved in cell proliferation, such as insulin-like growth factor-1 (IGF-1), epidermal growth factor, transforming growth factors, insulin, GH and PRL [37,38]. Actually, SST2,SST3, and SST5 trigger inhibition of GH synthesis in pituitary somatotropinoma [39,40,41] Moreover, it has been shown in different cell types, that somatostatin induces also inhibition of tumor necrosis factor-α and interleukins secretion also involved in the regulation of cell proliferation [42,43]. Another indirect antitumor effect of SSTs signaling is the inhibition of angiogenesis, which is mainly mediated by SST5. On the other hand, SST3 was indicated as pivotal initiator for downregulation of vascular endothelial growth factor (VEGF), which is crucial for vessel development under tumor hypoxia conditions [44,45].
SSTs activity regulation is also provided through desensitization and internalization of receptors. Interestingly, SSTs show peculiar features for these mechanisms. Internalization process, especially for SST2, is activated by binding of SSAs, while there is no evidence on SSTs internalization induced by somatostatin antagonist. Moreover, time and magnitude of internalization can modify type and susceptibility of the downstream response to SSAs. In particular SST2, which is the most studied SST, is internalized and rapidly recycled back to the plasma membrane without going through degradation [20,46,47,48]. In addition, ligand binding can regulate SSTs status. Indeed, SSTs can form both homo- and hetero-dimers with members of the same or distantly related receptor subtypes [10,36]. Altogether, these aspects might be responsible for the tumor response magnitude to SSAs in NENs with relevant clinical consequences.
Although endogenous somatostatin triggers powerful route for the negative control of hormone secretion and tumor progression, its clinical administration was limited by its short half-life (~2.5 min) [49]. Therefore, in order to overcome this obstacle first synthetic SSAs were developed with an increased half-life of 1.7–2 h. This first generation of SSAs include octreotide (SMS-201-995) and lanreotide (BIM-23014), released in 1982 and 1988, respectively [50,51]. Both SSAs have high binding affinity for SST2 and less affinity for SST5. Over the years, octreotide and lanreotide compounds showed powerful effects, mostly as palliative therapeutic options for treating symptoms related to NENs (flushing, diarrhea, etc.) [52,53,54,55,56,57]. Octreotide action was improved in 1997, when a long acting release (LAR) formulation came out, while lanreotide was also delivered in long acting formulation. These formulations require only one monthly intramuscular (octreotide) or deep subcutaneous (lanreotide) injection, and show better acceptability and patient compliance to therapy. A phase III study evaluated safety and efficacy of octreolin, a novel oral formulation of octreotide. This drug appears to be well tolerated, but the long-term effect is still elusive [58]. Over the years, anti-tumoral effects of both LAR octreotide and lanreotide were tested in two clinical trials PROMID and CLARINET, respectively. They brought positive results concerning anti-proliferative activity in NENs, in addition to efficacy for carcinoid syndrome control in functional tumors. PROMID trial revealed a significant longer time to tumor progression of patients treated with octreotide compared to placebo, while CLARINET trial showed a significant prolonged progression-free survival in patients treated with lanreotide vs. placebo. Unfortunately, neither octreotide nor lanreotide induce relevant tumor shrinkage in patients with NENs, but they are commonly associated to disease stabilization [4,5]. Notably, beside octreotide and lanreotide a novel generation of SSAs (panligands) was developed, such as pasireotide (SOM230). Pasireotide exhibits high binding affinity to SSTs: SST5 > SST2 > SST3 > SST1 [59,60]. These features lead to an enhanced anti-secretory activity in case of octreotide-resistant acromegalic tumors, corticotropinomas and in GH/PRL secreting adenomas [61,62]. Moreover, due to its action on VEGF downregulation and PI3K signaling [63,64], pasireotide showed important anti-proliferative effects in vitro, especially in primary culture of non-functioning pituitary adenomas, human corticotropinomas and pancreatic NENs [65,66,67]. Although in vivo studies have brought promising results, clinical trials have not been as positive as expected [68]. A phase II study of pasireotide LAR has shown an increased progression-free survival rate in a small number of patients with metastatic or unresectable NENs [69], while a phase III study demonstrated that pasireotide LAR and high-dose octreotide LAR have similar efficacy to conventional SSAs for symptom control in patients with functional NENs [70]. In addition, due to its incidence on glucose metabolism, which results in hyperglycemia and diabetes mellitus, pasireotide is not currently used for advanced NEN cases [71]. In 2001, another second generation SSA was released. This novel heptapeptide, named somatropim, was tested at first in vitro showing high affinity for SST2, SST4, and SST5 [72,73,74]. Although somatropim displayed higher anti-secretory effects compared to octreotide, no anti-proliferative and anti-tumor evidence was reported in a clinical trial [75].
3. Dopamine System in NENs
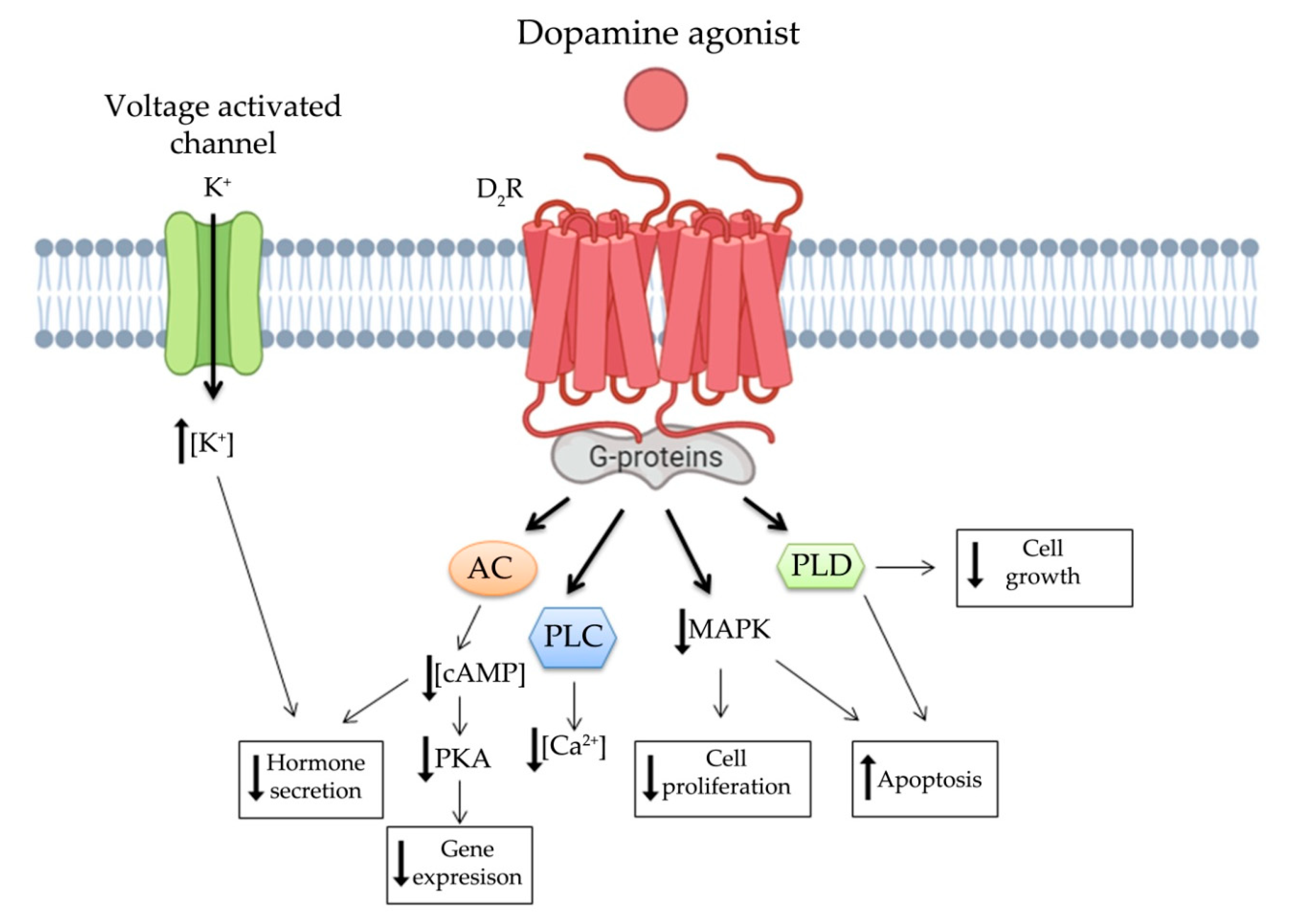
Dopamine is one of the most relevant neurotransmitter belonging to catecholamine family. Its action triggers a broad profile of functions, not only in the central nervous system but also in the periphery. Indeed, it is associated with regulation of cardiovascular and renal function, and it is mainly involved in regulation of hormone secretion [76]. Dopamine cascade signal is modulated by DRs, which are expressed in many cell subtypes of different organs. DRs, as well as SSTs, belong to the G-protein coupled receptor family and five subtypes of DRs were isolated, named D1R to D5R. The third intercellular loop of DRs interacts with G-proteins leading to cAMP regulations, while the hydrophobic transmembrane domain is crucial for DAs and antagonists ligand binding [77]. DRs can be divided into two groups based on their signaling cascades: D1-like group (D1R and D5R) and D2-like group (D2R, D3R, and D4R) [77]. D1-like group is associated to G-stimulatory protein, which leads to PKA activation through an increase in cAMP intracellular concentration. On the contrary, D2-like group inhibits PKA via G-inhibitory protein signaling. Notably, an alternative splicing event generates two isoforms of D2R, which differ for the third cytoplasmic domain, giving them different functional characteristics. The longer form, named D2RL, interacts with Gi-proteins, whereas the shorter one, D2RS, is able to activate phospholipase D providing important anti-proliferative effects [78,79]. D2-like group signaling cascade, including both D2RL and D2RS, is related to inhibition of MAPK signaling together with activation of phospholipase C, phospholipase D and different membrane channels such as calcium and potassium channels, Na+/K+ exchangers and Na+-K+-ATPase [76,77].
Increasing evidence shows that DRs are associated with the regulation of tumor behavior and proliferation, with relevant clinical applications in the therapy of pituitary NENs, particularly prolactinoma. The main anti-secretory and anti-tumor functions of dopamine appear to be modulated through D2R signaling [80]. These pathways are summarized in Figure 2. Anti-secretory functions are mainly associated with modulation of cAMP intracellular concentration and K+ voltage activated channels, while several signaling pathways contribute synergistically to anti-proliferative effects of DAs. Indeed, it has been demonstrated that prolactinomas treated with DAs, able to decrease cAMP intracellular concentration, show not only hormone secretion inhibition but also tumor shrinkage [81]. D2R activated pathway leads to cell death and apoptosis through the well-known p38 and JNK MAPK pathways [82]. D2RS induces stimulation of phospholipase D leading to inhibition of proliferation [79,83]. More recently, the activation of both D2R and D5R has been observed to inhibit pituitary tumor growth by autophagic cell death both in vitro and in vitro [84]. Furthermore, since D2R knockout mice show overexpression of VEGF, which is one of the main factors promoting neoangiogenesis and tumor growth, it was demonstrated that this growth factor is under dopaminergic control indicating a novel potential role of this class of receptors in oncology [66].
Over the years, DAs administration becomes largely used for the treatment of different pituitary NENs. It was demonstrated through immunohistochemistry and in situ hybridization that D2R is extensively expressed in all pituitary tumor types [85]. DAs represent the first-line treatment for PRL-secreting pituitary adenomas. In case of GH-, thyroid-stimulating hormone (TSH)-, and adenocorticotropin (ACTH)-secreting pituitary tumors, DAs are an off-label alternative treatment [80,86]. Cabergoline, which binds with high affinity D2R, is highly used in the therapy of these tumors and shows less side effects and more convenient dosing schedule than the older bromocriptine. Several in vivo experiments were performed to better understand the role of this receptor in cellular development and tumor progression. Indeed, it has been showed that D2R knockout mice develop prolactinomas with age [66]. In hyperprolactinemic patients, PRL reached normal serum level in 74.2% of subjects randomized to cabergoline at 0.3 or 0.6 mg once weekly for 9 weeks [87]. In a large study of patients with prolactinoma, cabergoline treatment normalized prolactin levels in 92% of patients with microprolactinomas and in 77% of patients with macroprolactinomas [88]. After 12–24 months of cabergoline therapy, more than 80% of the patients with macroprolactinomas showed tumor shrinkage (>20% of the baseline tumor size) and complete disappearance of the tumor mass in 26–36% of the cases [89]. On these bases, DA is the first-line medical treatment for prolactinomas. However, a subset of these patients is resistant to DA (20–30% for bromocriptine and 10% for cabergoline) [90]. In acromegalic patients carbergoline monotherapy was shown to achieve a significant decrease of GH and IGF-1 serum levels with doses varied from 0.3 to 7 mg/week (one to seven administrations per week). The mean maximal dose was 2.6 ± 1.5 mg/week, generally administered twice weekly [91,92]. In addition, a meta-analysis showed that, cabergoline single-agent therapy normalizes IGF-I levels in one third of patients with acromegaly, while in resistant acromegaly IGF-1 normalization is achieved in about half of patients when cabergoline is added to SSAs [93]. In Cushing’s disease response rates were 25–50% for cabergoline [94]. Cabergoline effects were also tested in NFPA, showing significant tumor shrinkage in about 30% of patients [95,96,97]. In addition, a decrease in gonadotropin levels was observed both in vitro and in vivo [98,99].
There are also few evidences about the potential antitumor activity of DA in non-pituitary origin NENs. In 1992 Farrell observed promising effect of bromocriptine on regulation of ACTH precursor secretion in small cell lung cancer cell line [100]. Subsequently, both bromocriptine and cabergoline were tested in patients with lung carcinoid showing a significant suppression in ectopic ACTH secretion [76,101]. In addition, it has been demonstrated that cabergoline has an inhibitory effect on cell growth in gastroenteropancreatic (GEP) NENs cells [102]. Indeed, it was reported a reduction of liver metastasis in a patient with polypeptide secreting islet cell tumor during therapy with DA [83].
4. Somatostatin and Dopamine Receptors Interaction in NENs
The well-studied signaling cascade of both somatostatin and dopamine in regulating hormone secretion inhibition has built-up one of the main cornerstones in the treatment of NENs. Several studies evaluated the expression profile of both SSTs and DRs in NENs. Altogether, in vitro autoradiography, real-time PCR and immunohistochemistry analyses have detected a large expression of SSTs and DRs in several types of human NENs. The most prevalent SST in NENs is SST2, expressed in almost all pituitary NENs [103]. NFPAs showed SST3 as the most expressed SSTs [104]. Actually, most of ACTH-secreting adenomas and a significant portion of GH-secreting adenomas express also SST5 [105,106]. Moreover, SST2 is found in 80–100% of GEP NENs, whereas it is expressed in 50–70% of insulinomas [107]. On the other hand, D2R expression has been demonstrated by immunohistochemistry in several pituitary adenomas [108,109,110] and in well-differentiated GEP NENs [111,112]. Following these assumptions, researchers evaluated co-expression of DRs and SSTs in NENs. SSTs and D2R co-expression has been demonstrated in BON-1, a pancreatic neuroendocrine cell line, and in several GEP NENs [113,114]. Recent studies demonstrated co-expression of SST2, SST5 and D2R specifically in a broad variety of NENs including bronchial, GEP NENs and ovarian carcinoid, in particular with higher expression of D2R in low-grade rather than high-grade [115,116,117,118].
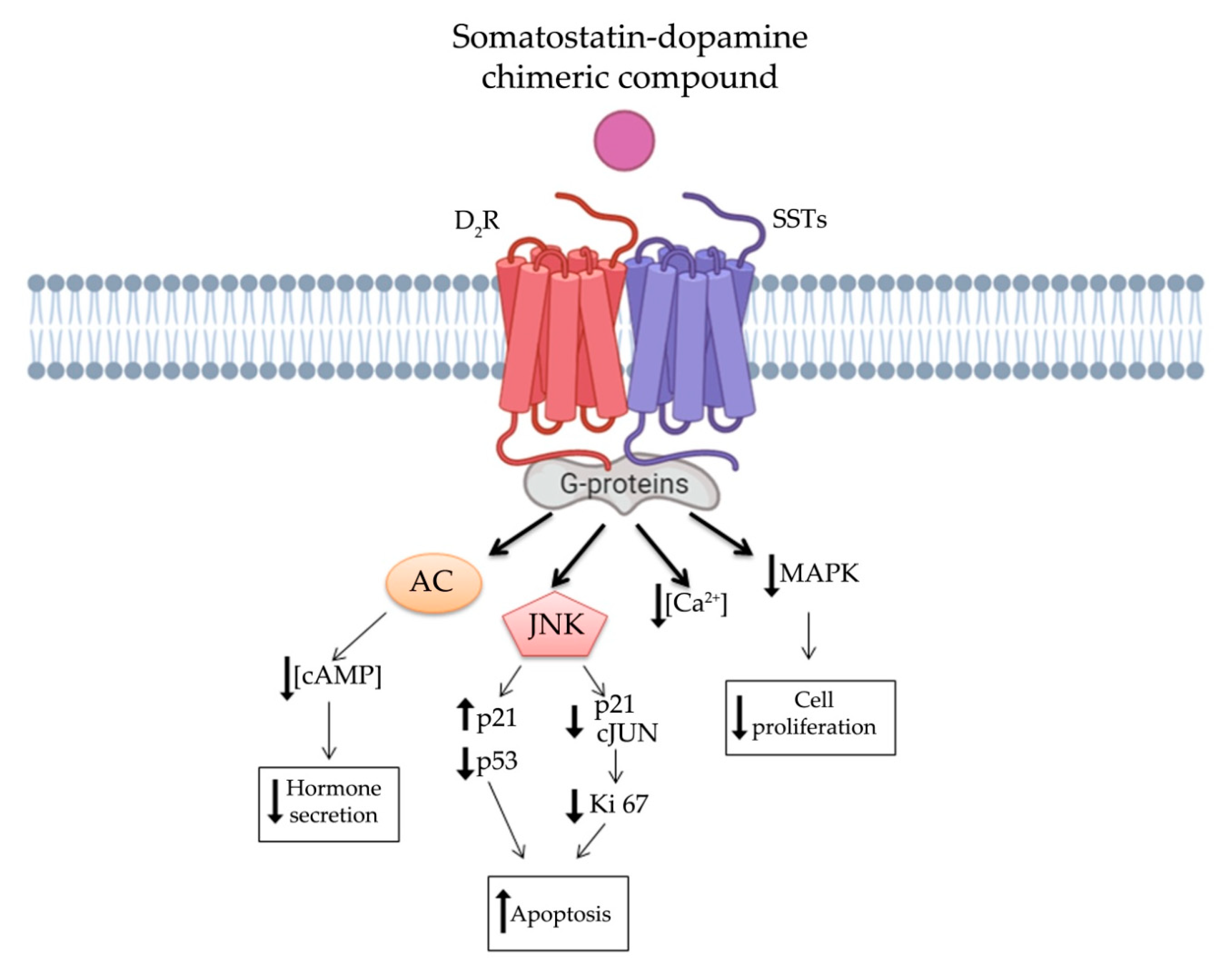
It is worthy of note that SSTs and DRs have been showed to interact via hetero- or homo- oligomerization. It has been demonstrated that heterodimerization of D2R and SST5 resulted in an enhanced functional activity and, more recently, also heterodimerization of D2R with SST2 has been reported [10,119]. Actually, it has been shown that D2R and SSTs (SST2 and SST5) heterodimers display a reciprocal influence on their effectors and an increased ability to their response [10]. These heterodimers may constitute a novel receptor, adding another level of complexity to the understanding of post-receptor events in NENs, but also the opportunity to discover a more specific and efficient target therapy. However, these two signaling cascades have several common down-streaming pathways, such as intracellular cAMP concentration, phospholipase C modulation and channels modifications leading to a comprehensive inhibition of hormone secretion (Figure 3). Additionally, they share the β-arrestin mechanism regulating receptors internalization and desensitization [120]. Speculating on these assumptions, several therapeutic combinations of SSAs and DAs were tested in patients with NENs. Indeed, therapy based on SSAs together with DAs, showed improvement for acromegaly and Cushing’s disease, as well as for resistant prolactinoma [121,122,123,124].
5. Somatostatin-Dopamine Chimeric Compounds
The growing knowledge of receptor profile expression in NENs has led to synthesis of several chimeric molecules targeting multi-receptors, with a particular interest for SSTs and DRs due to the functional interactions between these receptors. On these assumptions, novel chimeric compounds (Table 1), embracing somatostatin and dopamine features, were developed opening new routes for NENs treatments [125].
5.1. First Generation Chimeras: Advantages and Limitations
The first chimeric compound was synthetized by Ipsen Biomeasure, Inc. (Milford, MA, USA). This molecule, named BIM-23A387, combines structural elements of both SSAs and DAs. Based on above mentioned studies on receptor expression profile, BIM-23A387 retains affinity for SST2 and D2R (full agonist for both receptor). Savenau et al. [11] demonstrated that BIM-23A387 was more effective than either BIM-23023 (full agonist for SST2) or BIM-53097 (full agonist for D2R), either alone or in combination, in suppressing both GH- and PRL secretion in primary pituitary adenoma cells from patients previously treated with octreotide and/or quinagolide (a selective D2R agonist). The reason for the enhanced potency seems to be related to the heterodimerization of SSTs and DRs creating a novel receptor with distinct functionality. In 2003, another study analyzed mechanisms of action of BIM-23A387 in both human and rat pituitary adenoma primary cells. Ren et al. [126], showed that D2R antagonist, but not SST2 antagonist, abolished the SST2-active component of BIM-23A387 to inhibit GH secretion. In addition, the structure of the D2R/SST2 heterodimer might be different from the individual receptor forms, since it showed peculiar features: the heterodimer binding domain had a higher affinity to DA-selective ligand and the intracellular signaling occurred mainly via SST2 pathways [126]. Interestingly, BIM-23A387 was more potent in GH and PRL suppression compared to BIM-23023, BIM-53097, cabergoline, octreotide, and lanreotide. Subsequently, Jaquet et al. [127] tested several somatostatin-dopamine chimeric molecules (BIM-23A758, BIM-23A760 and BIM-23A761) in cells from 18 human partially responsive GH adenomas. The best results in suppressing GH and PRL secretion have been observed with BIM-23A760. Its effect was higher also than the first chimeric BIM-23A387 molecule and octreotide [127]. These results were also confirmed by Savenau et al. [128]. In this last study the inhibitory effects of octreotide, cabergoline and other chimeric molecules, including BIM-23A760 were tested on GH secretion in a large series of primary cultures from human GH-secreting pituitary tumors. BIM-23A760 showed the highest inhibitory effect [128]. Florio et al. demonstrated that BIM-23A760 has a higher efficiency in suppressing primary NFPAs cell proliferation than octreotide and cabergoline alone or in combination [129]. Moreover, BIM-23A760 molecule showed promising results in PRL- and TSH-secreting adenomas, when its antisecretory effect was compared with BIM-23A387, cabergoline or octreotide in primary cell lines [130,131]. In contrast, no improved effect on chromogranin A (CgA) secretion in GEP NENs was found using any chimeric molecules when compared with cabergoline or lanreotide [132]. Nevertheless, BIM-23A760 showed a decreased activation of IGF-induced insulin-like receptor-A in BON-1 cell line [133]. BIM-23A760 showed a predominant D2R signaling in primary cultures of DA-resistant prolactinomas, since its action did not provide an enhanced response when SST2 was successfully overexpressed [134]. In primary cultures of pituitary acromegaly, BIM-23A758 showed less efficiency in suppressing GH [127], while Zitzmann et al. observed that this chimeric compound displayed a greater antitumor activity than BIM-23A760 through Akt and MAPK signaling inhibition in human midgut carcinoid cells (GOT-1) [135]. Nevertheless, BIM-23A760 seems to modulate ERK1/2 pathway, showing a higher antiproliferative action compared to bromocriptine on primary NFPA cell culture [136,137]. Subsequently, both BIM-23A760 and BIM-23A761 antiproliferative effects were tested on bronchopulmonary cell lines (NCI-H720 and NCI-H727), small intestine cell line (KRJ-I) and in primary NFPA cells [137,138]. BIM-23A761 showed antiproliferative effect in bronchopulmonary cell lines through c-Jun N-terminal phosphorylation, leading to Ki67 down-regulation and an increase expression of p21 cell cycle inhibitor. Interestingly, KRJ cell lines did not respond to any chimeric compound [138]. Very recently, Halem and colleagues tested the anti-tumor effects of BIM-23A760 through subcutaneous injection in a novel pro-opiomelanocortin knock out mouse model, which is able to spontaneously develop aggressive NFPAs. BIM-23A760 was able to nearly complete arrest tumor growth within eight weeks and its action was significantly higher than octreotide and cabergoline alone and in combination [139].
Unfortunately, this evidence about BIM-23A760 failed to bring positive results in clinical human trials. Although the compound showed a clean safety profile in phase I and a significant therapeutic efficacy in a phase IIa single-dose study in patients with acromegaly, after repeated administration, BIM-23A760 produced a long-lasting and highly potent dopaminergic metabolite. Circulating accumulation of this metabolite gradually reduced the effects of the parent compound BIM-23A760, compromising its GH suppression activity [140,141].
5.2. Second Generation Chimeras: Advantages and Limitations
In order to overcome clinical limitations, recently, a novel chimeric compound named TBR-065 (formerly BIM-23B065), was synthetized by IPSEN Pharma (now in development by Tiburio Therapeutics, Cambridge, MA, USA). It is a full agonist at D2R and SST2 and a partial agonist at SST5 (Table 1). Testing its effects in vitro and in vivo TBR-065 showed a higher potency and efficacy in suppressing GH secretion compared with both octreotide and cabergoline (alone or in combination) in primary human pituitary adenoma cells from patients with partially octreotide-responsive acromegaly [142]. Furthermore, TBR-065 was shown to trigger apoptosis and to suppress hormones secretion and Ca2+ intracellular concentration in primary cell cultures from different pituitary tumors (ACTH-secreting adenomas, GH-secreting adenomas and NFPAs) [143] (Figure 3). In 2019, an interesting comparison between SSAs, DAs and novel chimeric compounds (BIM-23A760 and TBR-065) effects were performed in 2D and 3D of QGP-1 and BON-1 cell cultures models [144]. In this study, the authors showed that SST-D2R multi-receptor targeting drugs were able to inhibit CgA and serotonin secretion, but not cell growth [144]. Recently, we performed a preclinical study evaluating the antitumor activity of TBR-065 and lanreotide through both in vitro and in vivo models of medullary thyroid carcinoma. TBR-065 exerted a more relevant anti-tumor activity on human medullary thyroid carcinoma cell lines, when compared with lanreotide, through modulation of cell cycle, induction of apoptosis, and reduction in migration and calcitonin secretion. The zebrafish/tumor xenograft model, exploited in this study, did not show any effects on tumor-induced angiogenesis with both drugs [145]. Newly, the ability of TBR-065 to inhibit GH secretion from primary human GH or GH/PRLoma cells was compared with BIM-23A760, octreotide and cabergoline either alone or in combination. The authors showed that TBR-065 was the most efficient on GH suppression among them [146]. In another in vitro study, it was demonstrated that its main metabolite, BIM-23B133, has no binding to any SSTs and no interference with the chimera (TBR-065) activity [147]. Due to this analysis, TBR-065 appeared to be able to overcome one of the main limitations of the previous chimeric compound (BIM-23A760). Therefore, pharmacokinetics and pharmacodynamics, as well as safety and tolerability of this compound, were studied. The first human clinical trial was performed in 63 healthy male volunteers through subcutaneous injection of TBR-065. This phase I trial showed a significant reduction in GH, IGF-1 and PRL serum levels at doses upward of 0.4 mg. The main metabolite, BIM-23B133, did not interfere with the activity of TBR-065. Indeed, the inhibitory effects on secretion of GH, IGF-1, and PRL remained unaltered after 13 days of treatment with the new chimera [148]. Furthermore, the treatment was well tolerated with side effects mainly at high dosing regimens, specifically orthostatic hypotension (rarely vasovagal syncope), gastrointestinal disorders (abdominal discomfort, nausea and vomiting) and injection side reactions. An up-titration period of 6 days prevented the orthostatic effects, indeed side effects in the multiple ascending dose were less severe and no syncope was observed [148,149].
In the view of these findings, further research is needed for TBR-065, since it appears to be the most promising somatostatin-dopamine chimera with potential clinical applications in NENs.

{kind=link}
{kind=link}
{kind=link}
Table 1.
Comprehensive overview of somatostatin-dopamine chimeric compounds, their receptor binding affinity and antitumor activity in NENs. Human SSTs and D2R binding affinities are expressed in IC50 in nM, binding affinities marked with * are expressed with Ki (dissociation constant (nM)). ND: Not detected.
Table 1.
Comprehensive overview of somatostatin-dopamine chimeric compounds, their receptor binding affinity and antitumor activity in NENs. Human SSTs and D2R binding affinities are expressed in IC50 in nM, binding affinities marked with * are expressed with Ki (dissociation constant (nM)). ND: Not detected.
| Compound | Binding Affinity [125,127,134,137] | NEN Model | Anti-Tumor Activity | Ref. | |||||
|---|---|---|---|---|---|---|---|---|---|
| SST1 | SST2 | SST3 | SST4 | SST5 | D2R | ||||
| BIM-23A387 | 293 | 0.1 | 77.4 | ND | >1000 | 25 | Human and rat pituitary adenoma primary cells | ↓GH | [126] |
| Primary cultures of acromegaly | ↓GH | [11,127,128] | |||||||
| BIM-23A760 | 853 | 0.03 | 52 | 1000 | 3.1 | 15 | Primary cultures of acromegaly | ↓GH | [127,128] |
| Primary cultures of NFPA | ↓proliferation | [123,136,137] | |||||||
| Primary cultures of resistant prolactinoma | ↓PRL | [130,134] | |||||||
| Primary cultures of TSH-oma | ↓TSH | [131] | |||||||
| GEP NEN cell lines (BON-1 and QGP1) | ↓IGF-2 | [133] | |||||||
| Acromegaly patients (phase IIa study) | ↓GH serum levels | [140,141] | |||||||
| Broncopulmonary cell lines (NCI-H720 and NCI-H727) | ↓proliferation | [138] | |||||||
| In vivo NFPA mouse model | ↓proliferation | [139] | |||||||
| BIM-23A761 | 602 | 0.128 | 196 | >1000 | 8.7 | 25 | GEP NEN cell line (KRJ-I cell) | ↓proliferation | [138] |
| Primary cultures of acromegaly | ↓GH | [127] | |||||||
| BIM-23A758 | 549 * | 0.2 * | >1000 * | >1000 * | 43 * | 14 * | Human pancreatic NEN cells (BON-1) | ↑apoptosis, ↓Akt | [135] |
| Human midgut carcinoid cell (GOT-1) | ↑apoptosis, ↓Akt | [135] | |||||||
| Bronchopulmonary cell line (NCI-H727) | ↑apoptosis, ↓Akt | [135] | |||||||
| Primary cultures of acromegaly | ↓GH | [127] | |||||||
| BIM-23A781 | 93 | 0.8 | 11 | 42 | 4.5 | 29 | Primary cultures of acromegaly | ↓GH | [128] |
| Primary cultures of GEP NEN | ↓CgA | [132] | |||||||
| TBR-065 (BIM-23B065) | ND | 0.03 | ND | ND | 0.5 | 27.2 | Primary cultures of acromegaly | ↓cell viability, ↑apoptosis, ↓GH | [142,143,146] |
| Primary cultures of corticotropinoma | ↓cell viability, ↑apoptosis, ↓ATCH | [143] | |||||||
| Primary cultures of NFPA | ↑apoptosis, ↓CgA | [143] | |||||||
| AtT-20 cell line | ↑apoptosis | [143] | |||||||
| 3D cell models of QPG-1 and BON-1 cell | ↓ CgA, ↓Serotonin | [144] | |||||||
| MTC cell line (TT and MZ-CRC-1) | ↓cell viability, ↑apoptosis, ↓migration, ↓calcitonin | [145] | |||||||
| In vivo rat model | ↓GH | [147] | |||||||
| Healthy male volunteers (Phase I study) | ↓GH, ↓PRL and ↓IGF-I serum levels | [148,149] | |||||||
6. Conclusions
After almost twenty years from the development of the first somatostatin-dopamine chimeric molecule, prominent positive conclusions in the neuroendocrinology field can be drawn. First, it became clear the central role of SSAs and DAs in the therapy of well-differentiated NENs. Second, it has been observed that both SSTs and DRs displayed a large co-expression in several types of NENs. The most abundant receptors subtypes were SST2, SST5, and D2R. Interestingly, the formation of heterodimers between these receptors is associated with an enhanced antitumor activity. This evidence promoted the research for the development of new chimeric multi-target molecules. TBR-065 targeting SST2, SST5 and D2R appear to be a new molecule with relevant perspectives in the therapy of well-differentiated NENs, as shown by several preclinical and preliminary clinical studies. The great capability of TBR-065 to decrease in vivo GH, IGF-1 and PRL serum levels is encouraging.
Nevertheless, the knowledge of these chimeric molecules is leaving behind some outstanding questions which may be worth elucidating in further studies. For instance, both the hypothetical signal transduction pathway, that we have described in Figure 3, as well as a detailed binding affinity study for TBR-065 need to be well defined. Additionally, the binding affinity to D2R of available somatostatin-dopamine chimeric molecules is lower compared to that reported for the SST2. The development of new molecules with higher affinity for D2R could enhance the antitumor activity of these compounds.
In conclusion, despite the encouraging read-out about TBR-065, additional clinical studies are required to clearly define the safety, antitumor activity and the place of this molecule in the therapeutic algorithm of patients with NFPA, acromegaly, and advanced well-differentiated NENs.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- DeLellis, R.A. The Neuroendocrine System and Its Tumors an Overview. Rev. Am. J. Clin. Pathol. 2001, 115 (Suppl. S5–S16). [Google Scholar] [CrossRef]
- Alexandraki, K.I.; Tsoli, M.; Kyriakopoulos, G.; Angelousi, A.; Nikolopoulos, G.; Kolomodi, D.; Kaltsas, G.A. Current concepts in the diagnosis and management of neuroendocrine neoplasms of unknown primary origin. Minerva Endocrinol. 2019, 44, 378–386. [Google Scholar] [CrossRef] [PubMed]
- Tamagno, G.; Bennett, A.; Ivanovski, I. Lights and darks of neuroendocrine tumors of the appendix. Minerva Endocrinol. 2020, 45, 381–392. [Google Scholar] [CrossRef] [PubMed]
- Rinke, A.; Wittenberg, M.; Schade-Brittinger, C.; Aminossadati, B.; Ronicke, E.; Gress, T.M.; Müller, H.H.; Arnold, R.; PROMID Study Group. Placebo-Controlled, Double-Blind, Prospective, Randomized Study on the Effect of Octreotide LAR in the Control of Tumor Growth in Patients with Metastatic Neuroendocrine Midgut Tumors (PROMID): Results of Long-Term Survival. Neuroendocrinology 2017, 104, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Caplin, M.E.; Pavel, M.; Ćwikła, J.B.; Phan, A.T.; Raderer, M.; Sedláčková, E.; Cadiot, G.; Wolin, E.M.; Capdevila, J.; Wall, L.; et al. CLARINET Investigators. Anti-tumour effects of lanreotide for pancreatic and intestinal neuroendocrine tumours: The CLARINET open-label extension study. Endocr. Relat. Cancer 2016, 23, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Mazziotti, G.; Mosca, A.; Frara, S.; Vitale, G.; Giustina, A. Somatostatin analogs in the treatment of neuroendocrine tumors: Current and emerging aspects. Expert Opin. Pharmacother. 2017, 18, 1679–1689. [Google Scholar] [CrossRef]
- Peillon, F.; Cesselin, F.; Bression, D.; Zygelman, N.; Brandi, A.M.; Nousbaum, A.; Mauborgne, A. In Vitro Effect of Dopamine and L-Dopa on Prolactin and Growth Hormone Release from Human Pituitary Adenomas. Clin. Endocrinol. Metab. 1979, 49, 737–741. [Google Scholar] [CrossRef]
- Ferrari, C.; Paracchi, A.; Romano, C.; Gerevini, G.; Boghen, M.; Barreca, A.; Fortini, P.; Dubini, A. Long-lasting lowering of serum growth hormone and prolactin levels by single and repetitive cabergoline administration in dopamine-responsive acromegalic patients. Clinical. Trial. Clin. Endocrinol. 1988, 29, 467–476. [Google Scholar] [CrossRef]
- Günther, T.; Tulipano, G.; Dournaud, P.; Bousquet, C.; Csaba, Z.; Kreienkamp, H.J.; Lupp, A.; Korbonits, M.; Castaño, J.P.; Wester, H.J.; et al. International Union of Basic and Clinical Pharmacology. CV. Somatostatin Receptors: Structure, Function, Ligands, and New Nomenclature. Pharmacol. Rev. 2018, 70, 763–835. [Google Scholar] [CrossRef] [Green Version]
- Rocheville, M.; Lange, D.C.; Kumar, U.; Patel, S.C.; Patel, R.C.; Patel, Y.C. Receptors for dopamine and somatostatin: Formation of heterooligomers with enhanced functional activity. Science 2000, 288, 154–157. [Google Scholar] [CrossRef] [PubMed]
- Saveanu, A.; Lavaque, E.; Gunz, G.; Barlier, A.; Kim, S.; Taylor, J.E.; Culler, M.D.; Enjalbert, A.; Jaquet, P. Demonstration of Enhanced Potency of a Chimeric Somatostatin-Dopamine Molecule, BIM-23A387, in Suppressing Growth Hormone and Prolactin Secretion from Human Pituitary Somatotroph Adenoma Cells. J. Clin. Endocrinol. Metab. 2002, 87, 554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoyer, D. Somatostatin Receptors in the lUPHAR Compendium of Receptor Characterization and Classification; Nightingale Press: Royston, UK, 2000; pp. 354–364. [Google Scholar]
- Puig-Domingo, M.; Marazuela, M. Precision Medicine in the Treatment of Acromegaly. Minerva Endocrinol. 2019, 44, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Fuentes-Fayos, A.C.; García-Martínez, A.; Herrera-Martínez, A.D.; Jiménez-Vacas, J.M.; Vázquez-Borrego, M.C.; Castaño, J.P.; Picó, A.; Gahete, M.D.; Luque, R.M. Molecular Determinants of the Response to Medical Treatment of Growth Hormone Secreting Pituitary Neuroendocrine Tumors. Minerva Endocrinol. 2019, 44, 109–128. [Google Scholar] [CrossRef] [PubMed]
- Molè, D.; Gentilin, E.; Ibañez-Costa, A.; Gagliano, T.; Gahete, M.D.; Tagliati, F.; Rossi, R.; Pelizzo, M.R.; Pansini, G.; Luque, R.M.; et al. The expression of the truncated isoform of somatostatin receptor subtype 5 associates with aggressiveness in medullary thyroid carcinoma cells. Endocrine 2015, 50, 442–452. [Google Scholar] [CrossRef]
- Luque, R.M.; Ibáñez-Costa, A.; Neto, L.V.; Taboada, G.F.; Hormaechea-Agulla, D.; Kasuki, L.; Venegas-Moreno, E.; Moreno-Carazo, A.; Gálvez, M.Á.; Soto-Moreno, A.; et al. Truncated somatostatin receptor variant sst5TMD4 confers aggressive features (proliferation, invasion and reduced octreotide response) to somatotropinomas. Cancer Lett. 2015, 359, 299–306. [Google Scholar] [CrossRef] [Green Version]
- Durán-Prado, M.; Saveanu, A.; Luque, R.M.; Gahete, M.D.; Gracia-Navarro, F.; Jaquet, P.; Dufour, H.; Malagón, M.M.; Culler, M.D.; Barlier, A.; et al. A potential inhibitory role for the new truncated variant of somatostatin receptor 5, sst5TMD4, in pituitary adenomas poorly responsive to somatostatin analogs. J. Clin. Endocrinol. Metab. 2010, 95, 2497–2502. [Google Scholar] [CrossRef] [Green Version]
- Sampedro-Núñez, M.; Luque, R.M.; Ramos-Levi, A.M.; Gahete, M.D.; Serrano-Somavilla, A.; Villa-Osaba, A.; Adrados, M.; Ibáñez-Costa, A.; Martín-Pérez, E.; Culler, M.D.; et al. Presence of sst5TMD4, a Truncated Splice Variant of the Somatostatin Receptor Subtype 5, Is Associated to Features of Increased Aggressiveness in Pancreatic Neuroendocrine Tumors. Oncotarget 2016, 7, 6593–6608. [Google Scholar] [CrossRef] [Green Version]
- Martinez, V. Somatostatin. In Handbook of Biologically Active Peptides: Gastrointestinal Peptides, Copyright © 2021; Elsevier Inc.: Amsterdam, The Netherlands, 2013; pp. 1320–1329, Section XIII, Chapter 180. [Google Scholar]
- Schonbrunn, A. Selective agonism in somatostatin receptor signaling and regulation. Mol. Cell Endocrinol. 2008, 286, 35–39. [Google Scholar] [CrossRef] [Green Version]
- Siehler, S.; Hoyer, D. Characterisation of human recombinant somatostatin receptors. 2. Modulation of GTPgammaS binding. Naunyn Schmiedebergs Arch. Pharmacol. 1999, 360, 500–509. [Google Scholar] [CrossRef]
- Liu, Q.; Cescato, R.; Dewi, D.A.; Rivier, J.; Reubi, J.C.; Schonbrunn, A. Receptor signaling and endocytosis are differentially regulated by somatostatin analogs. Mol. Pharmacol. 2005, 68, 90–101. [Google Scholar] [CrossRef] [Green Version]
- Florio, T.; Rim, C.; Hershberger, R.E.; Loda, M.; Stork, P.J. The Somatostatin Receptor SSTR1 Is Coupled to Phosphotyrosine Phosphatase Activity in CHO-K1 Cells. Mol. Endocrinol. 1994, 8, 1289–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuzaki, S.; Moses, A. Somatostatin Inhibits Deoxyribonucleic Acid Synthesis Induced by Both Thyrotropin and Insulin-Like Growth factor-I in FRTL5 Cells. Endocrinology 1990, 126, 3131–3138. [Google Scholar] [CrossRef] [PubMed]
- Florio, T.; Scorizello, A.; Fattore, M.; D’Alto, V.; Salzano, S.; Rossi, G.; Berlingieri, M.T.; Fusco, A.; Schettini, G. Somatostatin Inhibits PC Cl3 Thyroid Cell Proliferation through the Modulation of Phosphotyrosine Activity. Impairment of the Somatostatinergic Effects by Stable Expression of E1A. J. Biol. Chem. 1996, 271, 6129–6136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reardon, D.B.; Wood, S.L.; Brautigan, D.L.; Bell, G.I.; Dent, P.; Sturgill, T.W. Activation of a Protein Tyrosine Phosphatase and Inactivation of Raf-1 by Somatostatin. Biochem. J. 1996, 314, 401–404. [Google Scholar] [CrossRef] [Green Version]
- Pan, M.G.; Florio, T.; Stork, P.J. G-protein activation of a hormone-stimulated phosphatase in human tumor-cells. Science 1992, 256, 1215–1217. [Google Scholar] [CrossRef]
- Cheung, N.W.; Boyages, S.C. Somatostatin-14 and its analog octreotide exert a cytostatic effect on GH3 rat pituitary tumor cell proliferation via a transient G0/ G1 cell cycle block. Endocrinology 1995, 136, 4174–4181. [Google Scholar] [CrossRef]
- Guillermet-Guibert, J.; Saint-Laurent, N.; Davenne, L.; Rochaix, P.; Cuvillier, O.; Culler, M.D.; Pradayrol, L.; Buscail, L.; Susini, C.; Bousquet, C. Novel synergistic mechanism for sst2 somatostatin and TNFalpha receptors to induce apoptosis: Crosstalk between NF-kappaB and JNK pathways. Cell Death Differ. 2007, 14, 197–208. [Google Scholar] [CrossRef]
- Theodoropoulou, M.; Zhang, J.; Laupheimer, S.; Paez-Pereda, M.; Erneux, C.; Florio, T.; Pagotto, U.; Stalla, G.K. Octreotide, a somatostatin analogue, mediates its antiproliferative action in pituitary tumor cells by altering phosphatidylinositol 3-kinase signaling and inducing Zac1 expression. Cancer Res. 2006, 66, 1576–1582. [Google Scholar] [CrossRef] [Green Version]
- Lopez, F.; Estève, J.P.; Buscail, L.; Delesque, N.; Saint-Laurent, N.; Théveniau, M.; Nahmias, C.; Vaysse, N.; Susini, C. The tyrosine phosphatase SHP-1 associates with the sst2 somatostatin receptor and is an essential component of sst2-mediated inhibitory growth signaling. J. Biol. Chem. 1997, 272, 24448–24454. [Google Scholar] [CrossRef] [Green Version]
- Sharma, K.; Srikant, C.B. Induction of wild-type p53, Bax, and acidic endonuclease during somatostatin-signaled apoptosis in MCF-7 human breast cancer cells. Int. J. Cancer 1998, 76, 259–266. [Google Scholar] [CrossRef]
- Liu, D.; Martino, G.; Thangaraju, M.; Sharma, M.; Halwani, F.; Shen, S.H.; Patel, Y.C.; Srikant, C.B. Caspase-8-mediated intracellular acidification precedes mitochondrial dysfunction in somatostatin-induced apoptosis. J. Biol. Chem. 2000, 275, 9244–9250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thangaraju, M.; Sharma, K.; Leber, B.; Andrews, D.W.; Shen, S.H.; Srikant, C.B. Regulation of acidification and apoptosis by SHP-1 and Bcl-2. J. Biol. Chem. 1999, 274, 29549–29557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teijeiro, R.; Rios, R.; Costoya, J.A.; Castro, R.; Bello, J.L.; Devesa, J.; Arce, V.M. Activation of human somatostatin receptor 2 promotes apoptosis through a mechanism that is independent from induction of p53. Cell Physiol. Biochem. 2002, 12, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Florio, T. Molecular mechanisms of the antiproliferative activity of somatostatin receptors (SSTRs) in neuroendocrine tumors. Front. Biosci. 2008, 13, 822–840. [Google Scholar] [CrossRef] [Green Version]
- Ambler, G.R.; Butler, A.A.; Padmanabhan, J.; Breier, B.H.; Gluckman, P.D. The effects of octreotide on GH receptor and IGF-I expression in the GH-deficient rat. J. Endocrinol. 1996, 149, 223–231. [Google Scholar] [CrossRef]
- Häyry, P.; Räisänen, A.; Ustinov, J.; Mennander, A.; Paavonen, T. Somatostatin analog lanreotide inhibits myocyte replication and several growth factors in allograft arteriosclerosis. FASEB J. 1993, 7, 1055–1060. [Google Scholar] [CrossRef]
- Trouillas, J.; Vasiljevic, A.; Lapoirie, M.; Chinezu, L.; Jouanneau, E.; Raverot, G. Pathological Markers of Somatotroph Pituitary Neuroendocrine Tumors Predicting the Response to Medical Treatment. Minerva Endocrinol. 2019, 44, 129–136. [Google Scholar] [CrossRef]
- Shimon, I.; Taylor, J.E.; Dong, J.Z.; Bitonte, R.A.; Kim, S.; Morgan, B.; Coy, D.H.; Culler, M.D.; Melmed, S. Somatostatin receptor subtype specificity in human fetal pituitary cultures. Differential role of SSTR2 and SSTR5 for growth hormone, thyroid-stimulating hormone, and prolactin regulation. J. Clin. Investig. 1997, 99, 789–798. [Google Scholar] [CrossRef]
- Murray, R.D.; Kim, K.; Ren, S.G.; Chelly, M.; Umehara, Y.; Melmed, S. Central and peripheral actions of somatostatin on the growth hormone-IGF-I axis. J. Clin. Invest. 2004, 114, 349–356. [Google Scholar] [CrossRef]
- Chowers, Y.; Cahalon, L.; Lahav, M.; Schor, H.; Tal, R.; Bar-Meir, S.; Levite, M. Somatostatin through its specific receptor inhibits spontaneous and TNF-alpha- and bacteria-induced IL-8 and IL-1 beta secretion from intestinal epithelial cells. J. Immunol. 2000, 165, 2955–2961. [Google Scholar] [CrossRef] [Green Version]
- Grimaldi, M.; Florio, T.; Schettini, G. Somatostatin inhibits interleukin 6 release from rat cortical type I astrocytes via the inhibition of adenylyl cyclase. Biochem. Biophys. Res. Commun. 1997, 235, 242–248. [Google Scholar] [CrossRef] [PubMed]
- Florio, T.; Morini, M.; Villa, V.; Arena, S.; Corsaro, A.; Thellung, S.; Culler, M.D.; Pfeffer, U.; Noonan, D.M.; Schettini, G.; et al. Somatostatin Inhibits Tumor Angiogenesis and Growth via Somatostatin receptor-3-mediated Regulation of Endothelial Nitric Oxide Synthase and Mitogen-Activated Protein Kinase Activities. Endocrinology 2003, 144, 1574–1584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theodoropoulou, M.; Stalla, G.K. Somatostatin receptors: From signaling to clinical practice. Front. Neuroendocrinol. 2013, 34, 228–252. [Google Scholar] [CrossRef] [PubMed]
- Hukovic, N.; Panetta, R.; Kumar, U.; Patel, Y.C. Agonist-dependent regulation of cloned human somatostatin receptor types 1-5 (hSSTR1-5): Subtype selective internalization or upregulation. Endocrinology 1996, 137, 4046–4049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Csaba, Z.; Peineau, S.; Dournaud, P. Molecular mechanisms of somatostatin receptor trafficking. J. Mol. Endocrinol. 2012, 48, R1–R12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hipkin, R.W.; Friedman, J.; Clark, R.B.; Eppler, C.M.; Schonbrunn, A. Agonist-induced desensitization, internalization, and phosphorylation of the sst2A somatostatin receptor. J. Biol. Chem. 1997, 272, 13869–13876. [Google Scholar] [CrossRef] [Green Version]
- Patel, Y.C.; Wheatley, T. In Vivo and In Vitro Plasma Disappearance and Metabolism of somatostatin-28 and somatostatin-14 in the Rat. Endocrinology 1983, 112, 220–225. [Google Scholar] [CrossRef]
- Bauer, W.; Briner, U.; Doepfner, W.; Haller, R.; Huguenin, R.; Marbach, P.; Petcher, T.J.; Pless, J. SMS 201-995: A Very Potent and Selective Octapeptide Analogue of Somatostatin with Prolonged Action. Life Sci. 1982, 31, 1133–1140. [Google Scholar] [CrossRef]
- Taylor, J.E.; Bogden, A.E.; Moreau, J.P.; Coy, D.H. In Vitro and In Vivo Inhibition of Human Small Cell Lung Carcinoma (NCI-H69) Growth by a Somatostatin Analogue. Biochem. Biophys. Res. Commun. 1988, 153, 81–86. [Google Scholar] [CrossRef]
- Vitale, G.; Tagliaferri, P.; Caraglia, M.; Rampone, E.; Ciccarelli, A.; Bianco, A.R.; Abbruzzese, A.; Lupoli, G. Slow Release Lanreotide in Combination with interferon-alpha2b in the Treatment of Symptomatic Advanced Medullary Thyroid Carcinoma. J. Clin. Endocrinol. Metab. 2000, 85, 983–988. [Google Scholar] [CrossRef]
- Lupoli, G.; Cascone, E.; Arlotta, F.; Vitale, G.; Celentano, L.; Salvatore, M.; Lombardi, G. Treatment of Advanced Medullary Thyroid Carcinoma with a Combination of Recombinant Interferon alpha-2b and Octreotide. Cancer 1996, 78, 1114–1118. [Google Scholar] [CrossRef]
- Hofland, J.; Herrera-Martínez, A.D.; Zandee, W.T.; de Herder, W.W. Management of carcinoid syndrome: A systematic review and meta-analysis. Endocr. Relat. Cancer 2019, 26, R145–R156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Efraij, K.; Aljama, M.A.; Kennecke, H.F. Association of Dose Escalation of Octreotide Long-Acting Release on Clinical Symptoms and Tumor Markers and Response among Patients with Neuroendocrine Tumors. Cancer Med. 2015, 4, 864–870. [Google Scholar] [CrossRef] [PubMed]
- O’Toole, D.; Ducreux, M.; Bommelaer, G.; Wemeau, J.L.; Bouché, O.; Catus, F.; Blumberg, J.; Ruszniewski, P. Treatment of carcinoid syndrome: A prospective crossover evaluation of lanreotide versus octreotide in terms of efficacy, patient acceptability, and tolerance. Cancer 2000, 88, 770–776. [Google Scholar] [CrossRef]
- Cella, D.; Beaumont, J.L.; Hudgens, S.; Marteau, F.; Feuilly, M.; Houchard, A.; Lapuerta, P.; Ramage, J.; Pavel, M.; Hörsch, D.; et al. Relationship Between Symptoms and Health-related Quality-of-life Benefits in Patients with Carcinoid Syndrome: Post Hoc Analyses from TELESTAR. Clin. Ther. 2018, 40, 2006–2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuvia, S.; Salama, P.; Weinstein, I.; Marom, K.; Neumark, E.; Arama, M.L.; Mishli, N.; Levy, S.; Lapidot, T.; Kadoshi, R.; et al. Octreolin™, a safe oral alternative for parenteral octreotide treatment. In Proceedings of the Fifth International Congress of the GRS and the IGF Society, New York, NY, USA, 3–7 October 2010. [Google Scholar]
- Bruns, C.; Lewis, I.; Briner, U.; Meno-Tetang, G.; Weckbecker, G. SOM230: A Novel Somatostatin Peptidomimetic with Broad Somatotropin Release Inhibiting Factor (SRIF) Receptor Binding and a Unique Antisecretory Profile. Eur. J. Endocrinol. 2002, 146, 707–716. [Google Scholar] [CrossRef]
- Schmid, H.A. Pasireotide (SOM230): Development, Mechanism of Action and Potential Applications. Mol. Cell Endocrinol. 2008, 286, 69–74. [Google Scholar] [CrossRef]
- Murray, R.D.; Kim, K.; Ren, S.G.; Lewis, I.; Weckbecker, G.; Bruns, C.; Melmed, S. The novel somatostatin ligand (SOM230) Regulates Human and Rat Anterior Pituitary Hormone Secretion. J. Clin. Endocrinol. Metab. 2004, 89, 3027–3032. [Google Scholar] [CrossRef] [Green Version]
- Van der Hoek, J.; de Herder, W.W.; Feelders, R.A.; van der Lely, A.J.; Uitterlinden, P.; Boerlin, V.; Bruns, C.; Poon, K.W.; Lewis, I.; Weckbecker, G.; et al. A Single-Dose Comparison of the Acute Effects between the New Somatostatin Analog SOM230 and Octreotide in Acromegalic Patients. J. Clin. Endocrinol. Metab. 2004, 89, 638–645. [Google Scholar] [CrossRef] [Green Version]
- Onofri, C.; Theodoropoulou, M.; Losa, M.; Uhl, E.; Lange, M.; Arzt, E.; Stalla, G.K.; Renner, U. Localization of Vascular Endothelial Growth Factor (VEGF) Receptors in Normal and Adenomatous Pituitaries: Detection of a Non-Endothelial Function of VEGF in Pituitary Tumours. J. Endocrinol. 2006, 191, 249–261. [Google Scholar] [CrossRef]
- Vidal, S.; Oliveira, M.C.; Kovacs, K.; Scheithauer, B.W.; Lloyd, R. Immunolocalization of Vascular Endothelial Growth Factor in the GH3 Cell Line. Cell Tissue Res. 2000, 300, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, A.; Blanchard, M.P.; Albertelli, M.; Barbieri, F.; Brue, T.; Niccoli, P.; Delpero, J.R.; Monges, G.; Garcia, S.; Ferone, D.; et al. Pasireotide and octreotide antiproliferative effects and sst2 trafficking in human pancreatic neuroendocrine tumor cultures. Endocr. Relat. Cancer 2014, 21, 691–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zatelli, M.C.; Piccin, D.; Vignali, C.; Tagliati, F.; Ambrosio, M.R.; Bondanelli, M.; Cimino, V.; Bianchi, A.; Schmid, H.A.; Scanarini, M.; et al. Pasireotide, a Multiple Somatostatin Receptor Subtypes Ligand, Reduces Cell Viability in Non-Functioning Pituitary Adenomas by Inhibiting Vascular Endothelial Growth Factor Secretion. Endocr. Relat. Cancer 2007, 14, 91–102. [Google Scholar] [CrossRef] [Green Version]
- Batista, D.L.; Zhang, X.; Gejman, R.; Ansell, P.J.; Zhou, Y.; Johnson, S.A.; Swearingen, B.; Hedley-Whyte, E.T.; Stratakis, C.A.; Klibanski, A. The Effects of SOM230 on Cell Proliferation and Adrenocorticotropin Secretion in Human Corticotroph Pituitary Adenomas. J. Clin. Endocrinol. Metab. 2006, 91, 4482–4488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stueven, A.K.; Kayser, A.; Wetz, C.; Amthauer, H.; Wree, A.; Tacke, F.; Wiedenmann, B.; Roderburg, C.; Jann, H. Somatostatin Analogues in the Treatment of Neuroendocrine Tumors: Past, Present and Future. Int. J. Mol. Sci. 2019, 20, 3049. [Google Scholar] [CrossRef] [Green Version]
- Cives, M.; Kunz, P.L.; Morse, B.; Coppola, D.; Schell, M.J.; Campos, T.; Nguyen, P.T.; Nandoskar, P.; Khandelwal, V.; Strosberg, J.R. Phase II clinical trial of pasireotide long-acting repeatable in patients with metastatic neuroendocrine tumors. Endocr. Relat. Cancer 2015, 22, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Wolin, E.M.; Jarzab, B.; Eriksson, B.; Walter, T.; Toumpanakis, C.; Morse, M.A.; Tomassetti, P.; Weber, M.M.; Fogelman, D.R.; Ramage, J.; et al. Phase III study of pasireotide long-acting release. Drug Des. Devel. Ther. 2015, 9, 5075–5086. [Google Scholar] [CrossRef] [Green Version]
- Vitale, G.; Dicitore, A.; Sciammarella, C.; Di Molfetta, S.; Rubino, M.; Faggiano, A.; Colao, A. Pasireotide in the treatment of neuroendocrine tumors: A review of the literature. Endocr. Relat. Cancer 2018, 25, R351–R364. [Google Scholar] [CrossRef] [Green Version]
- Afargan, M.; Janson, E.T.; Gelerman, G.; Rosenfeld, R.; Ziv, O.; Karpov, O.; Wolf, A.; Bracha, M.; Shohat, D.; Liapakis, G.; et al. Novel Long-Acting Somatostatin Analog with Endocrine Selectivity: Potent Suppression of Growth Hormone but Not of Insulin. Endocrinology 2001, 142, 477–486. [Google Scholar] [CrossRef]
- Gazal, S.; Gelerman, G.; Ziv, O.; Karpov, O.; Litman, P.; Bracha, M.; Afargan, M.; Gilon, C. Human Somatostatin Receptor Specificity of Backbone-Cyclic Analogues Containing Novel Sulfur Building Units. J. Med. Chem. 2002, 45, 1665–1671. [Google Scholar] [CrossRef]
- Shimon, I.; Rubinek, T.; Hadani, M.; Alhadef, N. PTR-3173 (Somatoprim), a Novel Somatostatin Analog with Affinity for Somatostatin Receptors 2, 4 and 5 Is a Potent Inhibitor of Human GH Secretion. J. Endocrinol. Investig. 2004, 27, 721–727. [Google Scholar] [CrossRef] [PubMed]
- Plöckinger, U.; Hoffmann, U.; Geese, M.; Lupp, A.; Buchfelder, M.; Flitsch, J.; Vajkoczy, P.; Jakob, W.; Saeger, W.; Schulz, S.; et al. DG3173 (Somatoprim), a Unique Somatostatin Receptor Subtypes 2-, 4- And 5-selective Analogue, Effectively Reduces GH Secretion in Human GH-secreting Pituitary Adenomas Even in Octreotide Non-Responsive Tumours. Eur. J. Endocrinol. 2012, 166, 223–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pivonello, R.; Ferone, D.; Lombardi, G.; Colao, A.; Lamberts, S.; Hofland, L.J. Novel insights in dopamine receptor physiology. Eur. J. Endocrinol. 2007, 156, S13–S21. [Google Scholar] [CrossRef] [PubMed]
- Missale, C.; Nash, S.R.; Robinson, S.W.; Jaber, M.; Caron, M.G. Dopamine Receptors: From Structure to Function. Physiol. Rev. 1998, 78, 189–225. [Google Scholar] [CrossRef] [Green Version]
- Ben-Shlomo, A.; Liu, N.A.; Melmed, S. Somatostatin and dopamine receptor regulation of pituitary somatotroph adenomas. Pituitary 2017, 20, 93–99. [Google Scholar] [CrossRef]
- Senogles, S.E. D2S dopamine receptor mediates phospholipase D and antiproliferation. Mol. Cell Endocrinol. 2003, 2209, 61–69. [Google Scholar] [CrossRef]
- De Camilli, P.; Macconi, D.; Spada, A. Dopamine inhibits adenylate cyclase in human prolactin-secreting pituitary adenomas. Nature 1979, 278, 252–254. [Google Scholar] [CrossRef]
- Bevan, J.S.; Webster, J.; Burke, C.W.; Scanlon, M.F. Dopamine Agonists and Pituitary Tumor Shrinkage. Endocr. Rev. 1992, 13, 220–240. [Google Scholar] [CrossRef]
- Choi, E.Y.; Jeong, D.; Park, K.W.; Baik, J.H. G protein-mediated mitogen-activated protein kinase activation by two dopamine D2 receptors. Biochem. Biophys. Res. Commun. 1999, 256, 33–40. [Google Scholar] [CrossRef]
- Pathak, R.D.; Tran, T.H.; Burshell, A.L. A case of dopamine agonists inhibiting pancreatic polypeptide secretion from an islet cell tumor. J. Clin. Endocrinol. Metab. 2004, 89, 581–584. [Google Scholar] [CrossRef] [Green Version]
- Leng, Z.G.; Lin, S.J.; Wu, Z.R.; Guo, Y.H.; Cai, L.; Shang, H.B.; Tang, H.; Xue, Y.J.; Lou, M.Q.; Zhao, W.; et al. Activation of DRD5 (dopamine receptor D5) inhibits tumor growth by autophagic cell death. Autophagy 2017, 13, 1404–1419. [Google Scholar] [CrossRef]
- Stefaneanu, L.; Kovacs, K.; Horvath, E.; Buchfelder, M.; Fahlbusch, R.; Lancranjan, L. Dopamine D2 receptor gene expression in human adenohypophysial adenomas. Endocrine 2001, 14, 329–336. [Google Scholar] [CrossRef]
- Pivonello, C.; Rousaki, P.; Negri, M.; Sarnataro, M.; Napolitano, M.; Marino, F.Z.; Patalano, R.; De Martino, M.C.; Sciammarella, C.; Faggiano, A.; et al. Effects of the single and combined treatment with dopamine agonist, somatostatin analog and mTOR inhibitors in a human lung carcinoidcell line: An in vitro study. Endocrine 2017, 56, 603–620. [Google Scholar] [CrossRef] [PubMed]
- Mattei, A.M.; Ferrari, C.; Baroldi, P.; Cavioni, V.; Paracchi, A.; Galparoli, C.; Romano, C.; Spellecchia, D.; Gerevini, G.; Crosignani, P.G. Prolactin-lowering Effect of Acute and Once Weekly Repetitive Oral Administration of Cabergoline at Two Dose Levels in Hyperprolactinemic Patients. J. Clin. Endocrinol. Metab. 1988, 66, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Verhelst, J.; Abs, R.; Maiter, D.; van den Bruel, A.; Vandeweghe, M.; Velkeniers, B.; Mockel, J.; Lamberigts, G.; Petrossians, P.; Coremans, P.; et al. Cabergoline in the treatment of hyperprolactinemia: A study in 455 patients. J. Clin. Endocrinol. Metab. 1999, 84, 2518–2522. [Google Scholar] [CrossRef] [PubMed]
- Colao, A.; Di Sarno, A.; Landi, M.L.; Scavuzzo, F.; Cappabianca, P.; Pivonello, R.; Volpe, R.; Di Salle, F.; Cirillo, S.; Annunziato, L.; et al. Macroprolactinoma shrinkage during cabergoline treatment is greater in naive patients than in patients pretreated with other dopamine agonists: A prospective study in 110 patients. J. Clin. Endocrinol. Metab. 2000, 85, 2247–2252. [Google Scholar] [CrossRef] [PubMed]
- Maiter, D. Management of Dopamine Agonist-Resistant Prolactinoma. Neuroendocrinology 2019, 109, 42–50. [Google Scholar] [CrossRef]
- Abs, J.R.; Verhelst, D.; Maiter, K.; Van Acker, F.; Nobels, J.L.; Coolens, C.; Beckers, M.e.A. Cabergoline in the Treatment of Acromegaly: A Study in 64 Patients. J. Clin. Endocrinol. Metab. 1998, 83, 374–378. [Google Scholar] [CrossRef]
- Kuhn, E.; Chanson, P. Cabergoline in acromegaly. Pituitary 2017, 20, 121–128. [Google Scholar] [CrossRef]
- Sandret, L.; Maison, P.; Chanson, P. Place of cabergoline in acromegaly: A meta-analysis. J. Clin. Endocrinol. Metab. 2011, 96, 1327–1335. [Google Scholar] [CrossRef] [Green Version]
- Gadelha, M.R.; Vieira Neto, L. Efficacy of medical treatment in Cushing’s disease: A systematic review. Clin. Endocrinol. 2014, 80, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Lohmann, T.; Trantakis, C.; Biesold, M.; Prothmann, S.; Guenzel, S.; Schober, R.; Paschke, R. Minor tumour shrinkage in nonfunctioning pituitary adenomas by long-term treatment with the dopamine agonist cabergoline. Pituitary 2001, 4, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Garcia, E.C.; Naves, L.A.; Silva, A.O.; de Castro, L.F.; Casulari, L.A.; Azevedo, M.F. Short-term treatment with cabergoline can lead to tumor shrinkage in patients with nonfunctioning pituitary adenomas. Pituitary 2013, 16, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Batista, R.L.; Musolino, N.; Cescato, V.; da Silva, G.O.; Medeiros, R.; Herkenhoff, C.; Trarbach, E.B.; Cunha-Neto, M.B. Nonfunctioning Pituitary Adenoma: A Single-Center, Open-Label, 2-Year Randomized Clinical Trial. Am. J. Clin. Oncol. 2019, 42, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Kwekkeboom, D.J.; Hofland, L.J.; van Koetsveld, P.M.; Singh, R.; van den Berge, J.H.; Lamberts, S.W. Bromocriptine increasingly suppresses the in vitro gonadotropin and alpha-subunit release from pituitary adenomas during long term culture. J. Clin. Endocrinol. Metab. 1990, 71, 718–724. [Google Scholar] [CrossRef] [PubMed]
- Kwekkeboom, D.J.; Lamberts, S.W. Long-term treatment with the dopamine agonist CV 205-502 of patients with a clinically non-functioning, gonadotroph, or alpha-subunit secreting pituitary adenoma. Clin. Endocrinol. 1992, 36, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Farrell, W.E.; Clark, A.J.; Stewart, M.F.; Crosby, S.R.; White, A. Bromocriptine inhibits pro-opiomelanocortin mRNA and ACTH precursor secretion in small cell lung cancer cell lines. J. Clin. Investig. 1992, 90, 705–710. [Google Scholar] [CrossRef]
- Reith, P.; Monnot, E.A.; Bathija, P.J. Prolonged suppression of a corticotropin-producing bronchial Carcinoid by Oral Bromocriptine. Arch. Intern. Med. 1987, 147, 989–991. [Google Scholar] [CrossRef]
- Pivonello, R.; Ceresol, E.; Albertelli, M.; Faggiano, A.; Torre, G.; De Martino, M.C.; Lombardi, G.; Minuto, F.; Colao, A.; Ferone, D. Expression of somatostatin and dopamine receptors and effect of chimeric somatostatin–dopamine molecules on cell proliferation in pancreatic neuroendocrine tumors. In Proceedings of the 5th Annual Conference, European Neuroendocrine Tumour Society, Paris, France, 6–8 March2008. [Google Scholar]
- Reubi, J.C.; Waser, B.; Schaer, J.C.; Laissue, J.A. Somatostatin receptor sst 1 -sst 5 expression in normal and neoplastic human tissues using receptor autoradiography with subtype-selective ligands. Eur. J. Nucl. Med. 2001, 28, 836–846. [Google Scholar] [CrossRef]
- Vitale, G.; Pivonello, R.; Ferone, D.; De Martino, M.C.; Auriemma, R.S.; Caraglia, M.; Abbruzzese, A.; Lombardi, G.; Colao, A. The role of somatostatin receptors in the medical treatment of acromegaly. Dig. Liver Dis. 1995, 36 (Suppl. S1), S55–S59. [Google Scholar] [CrossRef]
- Miller, G.M.; Alexander, J.M.; Bikkal, H.A.; Katznelson, L.; Zervas, N.T.; Klibanski, A. Somatostatin receptor subtype gene expression in pituitary adenomas. J. Clin. Endocrinol. Metab. 1995, 80, 1386–1392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hofland, L.J. Somatostatin and somatostatin receptors in Cushing’s disease. Mol. Cell Endocrinol. 2008, 286, 199–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reubi, J.C.; Waser, B. Concomitant expression of several peptide receptors in neuroendocrine tumours: Molecular basis for in vivo multireceptor tumour targeting. Eur. J. Nucl. Med. Mol. Imaging 2003, 30, 781–793. [Google Scholar] [CrossRef] [PubMed]
- Pivonello, R.; Ferone, D.; de Herder, W.W.; Kros, J.M.; De Caro, M.L.; Arvigo, M.; Annunziato, L.; Lombardi, G.; Colao, A.; Hofland, L.J.; et al. Dopamine receptor expression and function in corticotroph pituitary tumors. J. Clin. Endocrinol. Metab. 2004, 89, 2452–2462. [Google Scholar] [CrossRef] [Green Version]
- Ferone, D.; de Herder, W.W.; Pivonello, R.; Kros, J.M.; van Koetsveld, P.M.; de Jong, T.; Minuto, F.; Colao, A.; Lamberts, S.W.; Hofland, L.J. Correlation of in vitro and in vivo somatotropic adenoma responsiveness to somatostatin analogs and dopamine agonists with immunohistochemical evaluation of somatostatin and dopamine receptors and electron microscopy. J. Clin. Endocrinol. Metabol. 2008, 93, 1412–1417. [Google Scholar] [CrossRef] [Green Version]
- Saveanu, A.; Jaquet, P.; Brue, T.; Barlier, A. Relevance of coexpression of somatostatin and dopamine D2 receptors in pituitary adenomas. Mol. Cell Endocrinol. 2008, 286, 206–213. [Google Scholar] [CrossRef] [Green Version]
- Diakatou, E.; Kaltsas, G.; Tzivras, M.; Kanakis, G.; Papaliodi, E.; Kontogeorgos, G. Somatostatin and dopamine receptor profile of gastroenteropancreatic neuroendocrine tumors: An immunohistochemical study. Endocr. Pathol. 2011, 22, 24–30. [Google Scholar] [CrossRef]
- Grossrubatscher, E.; Veronese, S.; Ciaramella, P.D.; Pugliese, R.; Boniardi, M.; De Carlis, L.; Torre, M.; Ravini, M.; Gambacorta, M.; Loli, P. High expression of dopamine receptor subtype 2 in a large series of neuroendocrine tumors. Cancer Biol. Ther. 2008, 7, 1970–1978. [Google Scholar] [CrossRef] [Green Version]
- Lemmer, K.; Ahnert-Hilger, G.; Höpfner, M.; Hoegerle, S.; Faiss, S.; Grabowski, P.; Jockers-Scherübl, M.; Riecken, E.O.; Zeitz, M.; Scherübl, H. Expression of dopamine receptors and transporter in neuroendocrine gastrointestinal tumor cells. Life Sci. 2002, 71, 667–678. [Google Scholar] [CrossRef]
- O’Toole, D.; Saveanu, A.; Couvelard, A.; Gunz, G.; Enjalbert, A.; Jaquet, P.; Ruszniewski, P.; Barlier, A. The analysis of quantitative expression of somatostatin and dopamine receptors in gastro-entero-pancreatic tumours opens new therapeutic strategies. Eur. J. Endocrinol. 2006, 155, 849–857. [Google Scholar] [CrossRef]
- Srirajaskanthan, R.; Watkins, J.; Marelli, L.; Khan, K.; Caplin, M.E. Expression of Somatostatin and Dopamine 2 Receptors in Neuroendocrine Tumours and the Potential Role for New Biotherapies. Neuroendocrinology 2009, 89, 308–314n. [Google Scholar] [CrossRef] [PubMed]
- Ferone, D.; Arvigo, M.; Semino, C.; Jaquet, P.; Saveanu, A.; Taylor, J.E.; Moreau, J.P.; Culler, M.D.; Albertelli, M.; Minuto, F.; et al. Somatostatin and Dopamine Receptor Expression in Lung Carcinoma Cells and Effects of Chimeric Somatostatin-Dopamine Molecules on Cell Proliferation. Am. J. Physiol. Endocrinol. Metab. 2005, 289, E1044–E1050. [Google Scholar] [CrossRef] [PubMed]
- Kidd, M.; Modlin, I.M.; Pfragner, R.; Eick, G.N.; Champaneria, M.C.; Chan, A.K.; Camp, R.L.; Mane, S.M. Small bowel carcinoid (enterochromaffin cell) neoplasia exhibits transforming growth factor-beta1-mediated regulatory abnormalities including up-regulation of C-Myc and MTA1. Cancer 2007, 109, 2420–2431. [Google Scholar] [CrossRef] [PubMed]
- De Bruin, C.; Feelders, R.A.; Waaijers, A.M.; van Koetsveld, P.M.; Sprij-Mooij, D.M.; Lamberts, S.W.; Hofland, L.J. Differential regulation of human dopamine D2 and somatostatin receptor subtype expression by glucocorticoids in vitro. J. Mol. Endocrinol. 2009, 42, 47–56. [Google Scholar] [CrossRef]
- Baragli, A.; Alturaihi, H.; Watt, H.L.; Abdallah, A.; Kumar, U. Heterooligomerization of human dopamine receptor 2 and somatostatin receptor 2: Co-immunoprecipitation and fluorescence resonance energy transfer analysis. Cell Signal. 2007, 19, 2304–2316. [Google Scholar] [CrossRef]
- Gatto, F.; Hofland, L.J. The role of somatostatin and dopamine D2 receptors in endocrine tumors. Rev. Endocr. Relat. Cancer 2011, 18, R233–R251. [Google Scholar] [CrossRef] [Green Version]
- Lasolle, H.; Ferriere, A.; Vasiljevic, A.; Eimer, S.; Nunes, M.L.; Tabarin, A. Pasireotide-LAR in Acromegaly Patients Treated with a Combination Therapy: A Real-Life Study; Endocr Connect. Endocr. Connect. 2019, 8, 1383–1394. [Google Scholar] [CrossRef] [Green Version]
- Pivonello, R.; Ferone, D.; Lamberts, S.W.; Colao, A. Cabergoline plus Lanreotide for Ectopic Cushing’s Syndrome. N. Engl. J. Med. 2005, 352, 2457–2458. [Google Scholar] [CrossRef]
- Pivonello, R.; De Leo, M.; Cozzolino, A.; Colao, A. The Treatment of Cushing’s Disease. Endocr. Rev. 2015, 36, 385–486. [Google Scholar] [CrossRef]
- Sosa-Eroza, E.; Espinosa, E.; Ramírez-Rentería, C.; Mendoza, V.; Arreola, R.; Mercado, M. Treatment of Multiresistant Prolactinomas with a Combination of Cabergoline and Octreotide LAR. Endocrine 2018, 61, 343–348. [Google Scholar] [CrossRef]
- Hofland, L.J.; Vandamme, T.; Albertelli, M.; Ferone, D. Hormone and Receptor Candidates for Target and Biotherapy of Neuroendocrine Tumors. Front. Horm Res. 2015, 44, 216–238. [Google Scholar] [CrossRef] [PubMed]
- Ren, S.G.; Kim, S.; Taylor, J.; Dong, J.; Moreau, J.P.; Culler, M.D.; Melmed, S. Suppression of rat and human growth hormone and prolactin secretion by a novel somatostatin/dopaminergic chimeric ligand. J. Clin. Endocrinol. Metab. 2003, 88, 5414–5421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaquet, P.; Gunz, G.; Saveanu, A.; Dufour, H.; Taylor, J.; Dong, J.; Kim, S.; Moreau, J.P.; Enjalbert, A.; Culler, M.D. Efficacy of Chimeric Molecules Directed Towards Multiple Somatostatin and Dopamine Receptors on Inhibition of GH and Prolactin Secretion From GH-secreting Pituitary Adenomas Classified as Partially Responsive to Somatostatin Analog Therapy. Eur. J. Endocrinol. 2005, 153, 135–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saveanu, A.; Gunz, G.; Guillen, S.; Dufour, H.; Culler, M.D.; Jaquet, P. Somatostatin and dopamine-somatostatin multiple ligands directed towards somatostatin and dopamine receptors in pituitary adenomas. Neuroendocrinology 2006, 283, 258–263. [Google Scholar] [CrossRef] [PubMed]
- Florio, T.; Barbieri, F.; Spaziante, R.; Zona, G.; Hofland, L.J.; van Koetsveld, P.M.; Feelders, R.A.; Stalla, G.K.; Theodoropoulou, M.; Culler, M.D.; et al. Efficacy of a Dopamine-Somatostatin Chimeric Molecule, BIM-23A760, in the Control of Cell Growth from Primary Cultures of Human Non-Functioning Pituitary Adenomas: A Multi-Center Study. Multicent. Study Endocr. Relat. Cancer 2008, 15, 583–596. [Google Scholar] [CrossRef] [Green Version]
- Fusco, A.; Gunz, G.; Jaquet, P.; Dufour, H.; Germanetti, A.L.; Culler, M.D.; Barlier, A.; Saveanu, A. Somatostatinergic Ligands in Dopamine-Sensitive and -Resistant Prolactinomas. Eur. J. Endocrinol. 2008, 158, 595–603. [Google Scholar] [CrossRef] [Green Version]
- Gatto, F.; Barbieri, F.; Gatti, M.; Wurth, R.; Schulz, S.; Ravetti, J.L.; Zona, G.; Culler, M.D.; Saveanu, A.; Giusti, M.; et al. Balance Between Somatostatin and D2 Receptor Expression Drives TSH-secreting Adenoma Response to Somatostatin Analogues and Dopastatins. Clin. Endocrinol. 2012, 76, 407–414. [Google Scholar] [CrossRef]
- Couvelard, A.; Pélaprat, D.; Dokmak, S.; Sauvanet, A.; Voisin, T.; Couvineau, A.; Ruszniewski, P. Antisecretory Effects of Chimeric Somatostatin/Dopamine Receptor Ligands on Gastroenteropancreatic Neuroendocrine Tumors. Pancreas 2017, 46, 631–638. [Google Scholar] [CrossRef]
- Van Adrichem, R.C.; de Herder, W.W.; Kamp, K.; Brugts, M.P.; de Krijger, R.R.; Sprij-Mooij, D.M.; Lamberts, S.W.; van Koetsveld, P.M.; Janssen, J.A.; Hofland, L.J. Effects of Somatostatin Analogs and Dopamine Agonists on Insulin-Like Growth Factor 2-Induced Insulin Receptor Isoform A Activation by Gastroenteropancreatic Neuroendocrine Tumor Cells. Neuroendocrinology 2016, 103, 815–825. [Google Scholar] [CrossRef] [Green Version]
- Cuny, T.; Mohamed, A.; Graillon, T.; Roche, C.; Defilles, C.; Germanetti, A.L.; Couderc, B.; Figarella-Branger, D.; Enjalbert, A.; Barlier, A.; et al. Somatostatin Receptor sst2 Gene Transfer in Human Prolactinomas in Vitro: Impact on Sensitivity to Dopamine, Somatostatin and Dopastatin, in the Control of Prolactin Secretion. Mol. Cell Endocrinol. 2012, 355, 106–113. [Google Scholar] [CrossRef]
- Zitzmann, K.; Andersen, S.; Vlotides, G.; Spöttl, G.; Zhang, S.; Datta, R.; Culler, M.; Göke, B.; Auernhammer, C.J. The Novel Somatostatin Receptor 2/dopamine Type 2 Receptor Chimeric Compound BIM-23A758 Decreases the Viability of Human GOT1 Midgut Carcinoid Cells. Neuroendocrinology 2013, 98, 128–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gruszka, A.; Kunert-Radek, J.; Radek, A.; Pisarek, H.; Taylor, J.; Dong, J.Z.; Culler, M.D.; Pawlikowski, M. The Effect of Selective sst1, sst2, sst5 Somatostatin Receptors Agonists, a Somatostatin/Dopamine (SST/DA) Chimera and Bromocriptine on the “Clinically Non-Functioning” Pituitary Adenomas In Vitro. Life Sci. 2006, 78, 689–693. [Google Scholar] [CrossRef] [PubMed]
- Peverelli, E.; Olgiati, L.; Locatelli, M.; Magni, P.; Fustini, M.F.; Frank, G.; Mantovani, G.; Beck-Peccoz, P.; Spada, A.; Lania, A. The Dopamine-Somatostatin Chimeric Compound BIM-23A760 Exerts Antiproliferative and Cytotoxic Effects in Human Non-Functioning Pituitary Tumors by Activating ERK1/2 and p38 Pathways. Cancer Lett. 2010, 288, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Kidd, M.; Drozdov, I.; Joseph, R.; Pfragner, R.; Culler, M.; Modlin, I. Differential Cytotoxicity of Novel Somatostatin and dopamine Chimeric compounds on Bronchopulmonary and Small intestinal Neuroendocrine tumor cell lines Cancer. Cancer 2008, 113, 690–700. [Google Scholar] [CrossRef] [Green Version]
- Halem, H.A.; Hochgeschwender, U.; Rih, J.K.; Nelson, R.; Johnson, G.A.; Thiagalingam, A.; Culler, M.D. TBR-760, a Dopamine-Somatostatin Compound, Arrests Growth of Aggressive Nonfunctioning Pituitary Adenomas in Mice. Endocrinology 2020, 161, bqaa101. [Google Scholar] [CrossRef]
- Froehlich, J.; Ramis, J.; Lesage, C.; Obach, R. Safety, Pharmacokinetics (PK) and Pharmacodynamics (PD) after Subcutaneous (s. c.) Administration (adm) of BIM23A760, a Chimeric Compound Combining Dopaminergic Agonist and Somatostatin Analogue, in Healthy Male Volunteers. In Proceedings of the 91st Annual Meeting of Endocrine Society, Washington, DC, USA, 10–13 June 2009; pp. 3–685. [Google Scholar]
- Lesage, C.; Seymour, C.; Urbanavivius, G.; Colao, A.M. A Phase II Exploratory Study of BIM23A760 in Acromegalic Patients: Preliminary Results of Safety and Efficacy after a Single-Dose Administration. In Proceedings of the 91st Annual Meeting of the Endocrine Society, Washington, DC, USA, 10–13 June 2009; pp. 3–673. [Google Scholar]
- Saveanu, A.; Datta, R.; Zhang, S.; Shen, Y.; Dong, J.Z.; Graillon, T.; Desfilles, C.; Landsman, T.; Halem, H.; Enjalbert, A.; et al. Novel somatostatin-dopamine chimeric compound demonstrates superior efficacy in suppressing growth hormone secretion from human acromegalic tumors partially responsive to current somatostatin and dopamine therapies. In Proceedings of the Endocrine Society’s 98th Annual Meeting and Expo, Boston, MA, USA, 1–4 April 2016. [Google Scholar]
- Vázquez-Borrego, M.C.; L-López, F.; Gálvez-Moreno, M.A.; Fuentes-Fayos, A.C.; Venegas-Moreno, E.; Herrera-Martínez, A.D.; Blanco-Acevedo, C.; Solivera, J.; Landsman, T.; Gahete, M.D.; et al. A New Generation Somatostatin-Dopamine Analogue Exerts Potent Antitumoral Actions on Pituitary Neuroendocrine Tumor Cells. Neuroendocrinology 2020, 110, 70–82. [Google Scholar] [CrossRef]
- Herrera-Martínez, A.D.; van den Dungen, R.; Dogan-Oruc, F.; van Koetsveld, P.M.; Culler, M.D.; de Herder, W.W.; Luque, R.M.; Feelders, R.A.; Hofland, L.J. Effects of Novel Somatostatin-Dopamine Chimeric Drugs in 2D and 3D Cell Culture Models of Neuroendocrine Tumors. Endocr. Relat. Cancer 2019, 26, 585–599. [Google Scholar] [CrossRef]
- Dicitore, A.; Cantone, M.C.; Gaudenzi, G.; Saronni, D.; Carra, S.; Borghi, M.O.; Albertelli, M.; Ferone, D.; Hofland, L.J.; Persani, L.; et al. Efficacy of a novel second-generation somatostatin-dopamine chimera (TBR-065) in human medullary thyroid cancer: A preclinical study. Neuroendocrinology 2020. online ahead of print. [Google Scholar] [CrossRef]
- Cuny, T.; Graillon, T.; Defilles, C.; Datta, R.; Zhang, S.; Figarella-Branger, D.; Dufour, H.; Mougel, G.; Brue, T.; Landsman, T.; et al. Characterization of the ability of a, second-generation SST-DA chimeric molecule, TBR-065, to suppress GH secretion from human GH-secreting adenoma cells. Pituitary 2021. advance online publication. [Google Scholar] [CrossRef]
- Halem, H.; Zhang, S.; Datta, R.; Bastille, A.; Beech, J.; Marques, M.; Patel, S.; Shen, Y.; Dong, J.; Culler, M.D. A novel somatostatin-dopamine chimeric compound induces dose-related suppression of GHRH-stimulated growth hormone secretion and increases insulin sensitivity in normal rats. In Proceedings of the Endocrine Society’s 98th Annual Meeting and Expo, Boston, MA, USA, 1–4 April 2016. [Google Scholar]
- De Boon, W.; van Esdonk, M.J.; Stuurman, F.E.; Biermasz, N.R.; Pons, L.; Paty, I.; Burggraaf, J. A Novel Somatostatin-Dopamine Chimera (BIM23B065) Reduced GH Secretion in a First-in-Human Clinical Trial. J. Clin. Endocrinol. Metab. 2019, 104, 883–891. [Google Scholar] [CrossRef] [Green Version]
- Van Esdonk, M.J.; Stevens, J.; Stuurman, F.E.; de Boon, W.; Dehez, M.; van der Graaf, P.H.; Burggraaf, J. The Pharmacodynamic Effects of a Dopamine–Somatostatin Chimera Agonist on the Cardiovascular System Reduced GH Secretion in a First in-Human Clinical Trial. J. Cardiovasc. Pharmacol. 2019, 74, 128–136. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Simplified representation of SSTs signaling: Somatostatin and SSAs binding to SSTs activate G proteins and inhibit AC activity, inhibit Na+/H+ exchanger, activate inwardly rectifying K+ channels and Ca2+ channels. These pathways modulate the antitumor activity of native somatostatin and SSAs through inhibition in secretion of several hormones and proangiogenic factors, induction of apoptosis and cell cycle arrest. GFR, growth factor receptor; PLC, phospholipase C; AC, adenylyl cyclase; cAMP, cyclic adenosine monophosphate; VEGF, vascular endothelial growth factor; PTP, phosphotyrosine phosphatase; MAPK, mitogen-activated protein kinase and Akt protein kinase B.
Figure 1.
Simplified representation of SSTs signaling: Somatostatin and SSAs binding to SSTs activate G proteins and inhibit AC activity, inhibit Na+/H+ exchanger, activate inwardly rectifying K+ channels and Ca2+ channels. These pathways modulate the antitumor activity of native somatostatin and SSAs through inhibition in secretion of several hormones and proangiogenic factors, induction of apoptosis and cell cycle arrest. GFR, growth factor receptor; PLC, phospholipase C; AC, adenylyl cyclase; cAMP, cyclic adenosine monophosphate; VEGF, vascular endothelial growth factor; PTP, phosphotyrosine phosphatase; MAPK, mitogen-activated protein kinase and Akt protein kinase B.

Figure 2.
Simplified representation of D2 signaling. Dopamine and DA binding to D2R, via interaction with G proteins, inhibit AC activity and phosphatidylinositol metabolism, activate voltage-activated potassium channels and decrease voltage-calcium currents, and modulates the activity of PLC, PLD, and MAPKs. These processes result in decreasing hormone secretion and cell proliferation. AC, adenylyl cyclase; PLC, phospholipase C; MAPK, mitogen-activated protein kinase, PLD, phospholipase D; cAMP, cyclic adenosine monophosphate and PKA, protein kinase A.
Figure 2.
Simplified representation of D2 signaling. Dopamine and DA binding to D2R, via interaction with G proteins, inhibit AC activity and phosphatidylinositol metabolism, activate voltage-activated potassium channels and decrease voltage-calcium currents, and modulates the activity of PLC, PLD, and MAPKs. These processes result in decreasing hormone secretion and cell proliferation. AC, adenylyl cyclase; PLC, phospholipase C; MAPK, mitogen-activated protein kinase, PLD, phospholipase D; cAMP, cyclic adenosine monophosphate and PKA, protein kinase A.

Figure 3.
Somatostatin-dopamine chimeric compound induced dimerization of somatostatin and dopamine receptors. Potential intracellular signal pathways linked to receptors heterodimerization and involved in inhibition of hormone secretion, increased apoptosis, and decreased cell proliferation. AC, adenylyl cyclase; cAMP, cyclic adenosine monophosphate; JNK, c-Jun N-terminal kinase and MAPK, mitogen-activated protein kinase.
Figure 3.
Somatostatin-dopamine chimeric compound induced dimerization of somatostatin and dopamine receptors. Potential intracellular signal pathways linked to receptors heterodimerization and involved in inhibition of hormone secretion, increased apoptosis, and decreased cell proliferation. AC, adenylyl cyclase; cAMP, cyclic adenosine monophosphate; JNK, c-Jun N-terminal kinase and MAPK, mitogen-activated protein kinase.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Cantone, M.C.; Dicitore, A.; Vitale, G. Somatostatin-Dopamine Chimeric Molecules in Neuroendocrine Neoplasms. J. Clin. Med. 2021, 10, 501. https://doi.org/10.3390/jcm10030501
AMA Style
Cantone MC, Dicitore A, Vitale G. Somatostatin-Dopamine Chimeric Molecules in Neuroendocrine Neoplasms. Journal of Clinical Medicine. 2021; 10(3):501. https://doi.org/10.3390/jcm10030501
Chicago/Turabian StyleCantone, Maria Celeste, Alessandra Dicitore, and Giovanni Vitale. 2021. "Somatostatin-Dopamine Chimeric Molecules in Neuroendocrine Neoplasms" Journal of Clinical Medicine 10, no. 3: 501. https://doi.org/10.3390/jcm10030501
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.