
Ecological Observations on Hybrid Populations of European Plethodontid Salamanders, Genus Speleomantes

 ,
,  ,
,  ,
,  ,
,  and
and 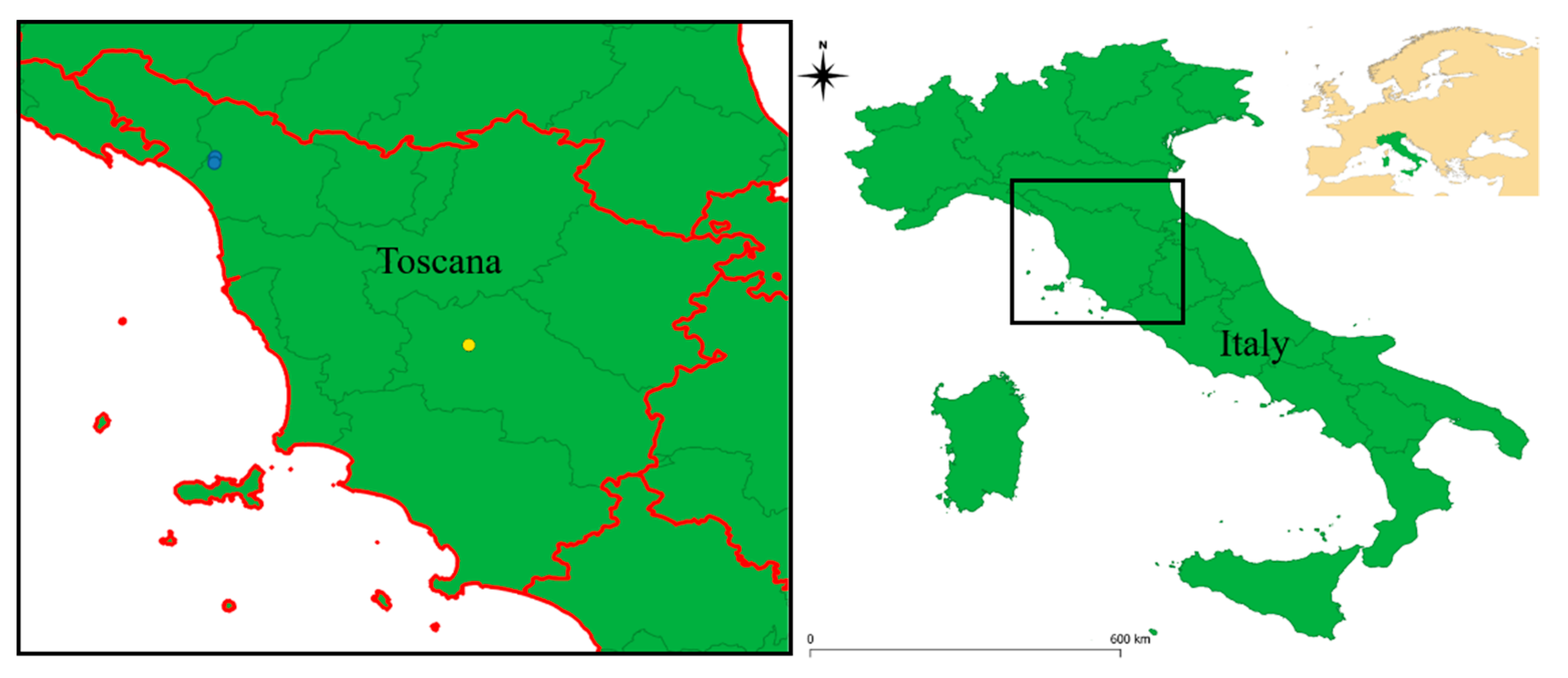{kind=link}
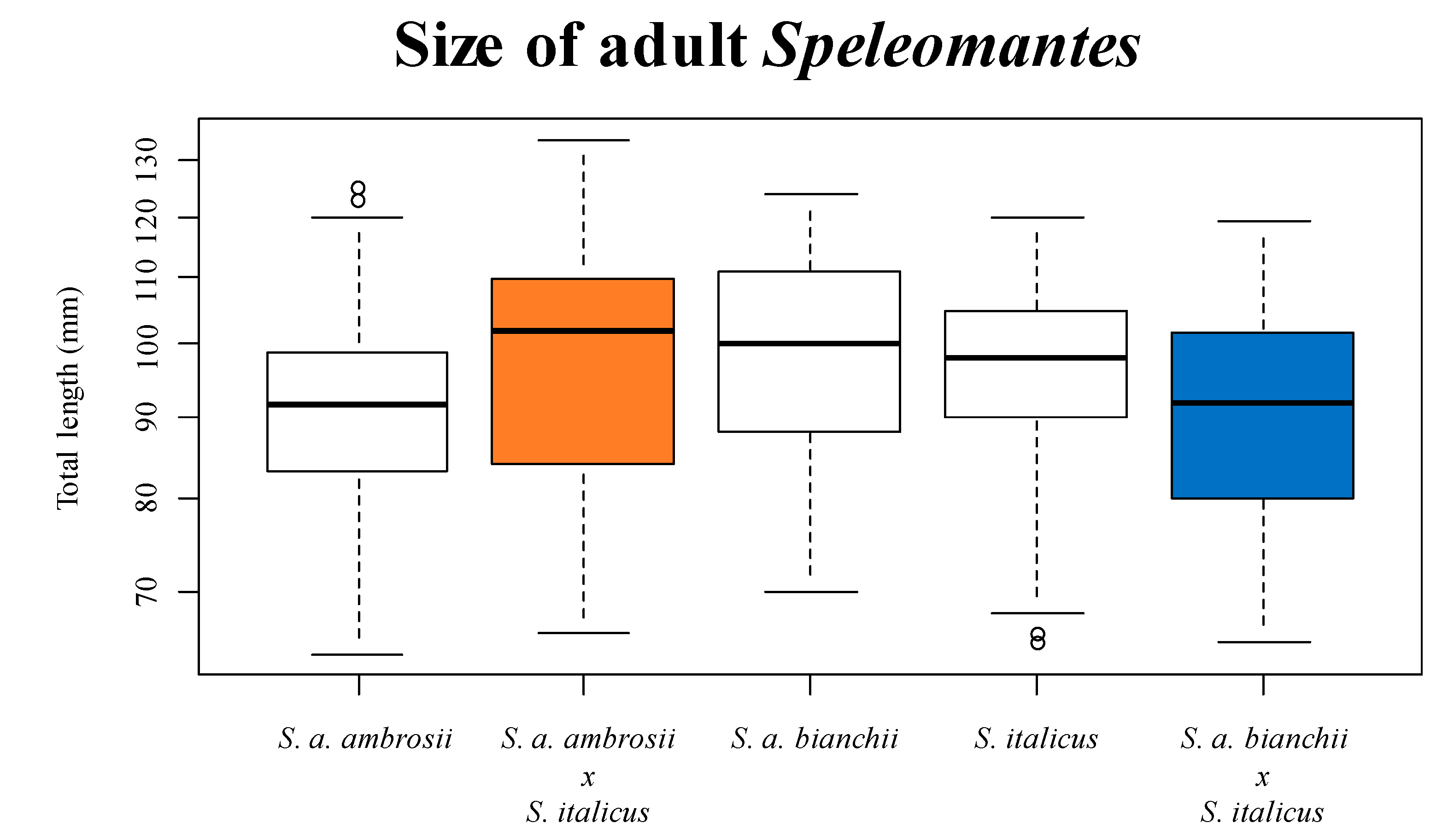{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lanza, B.; Pastorelli, C.; Laghi, P.; Cimmaruta, R. A review of systematics, taxonomy, genetics, biogeography and natural history of the genus Speleomantes Dubois, 1984 (Amphibia Caudata Plethodontidae). Atti. Mus Civ. Stor. Nat. Trieste 2006, 52, 5–135. [Google Scholar]
- Lanza, B.; Andreone, F.; Bologna, M.A.; Corti, C.; Razzetti, E. Fauna d’Italia. Amphibia; Calderini: Bologna, Italy, 2006. [Google Scholar]
- Chiari, Y.; van der Meijden, A.; Mucedda, M.; Lourenço, J.M.; Hochkirch, A.; Veith, M. Phylogeography of Sardinian cave salamanders (genus Hydromantes) is mainly determined by geomorphology. PLoS ONE 2012, 7, e32332. [Google Scholar] [CrossRef] [Green Version]
- Carranza, S.; Romano, A.; Arnold, E.N.; Sotgiu, G. Biogeography and evolution of European cave salamanders, Hydromantes (Urodela: Plethodontidae), inferred from mtDNA sequences. J. Biogeogr. 2008, 35, 724–738. [Google Scholar] [CrossRef]
- Lunghi, E.; Cianferoni, F.; Giachello, S.; Zhao, Y.; Manenti, R.; Corti, C.; Ficetola, G.F. Updating salamander datasets with phenotypic and stomach content information for two mainland Speleomantes. Sci. Data 2021, 8, 150. [Google Scholar] [CrossRef]
- Lunghi, E.; Giachello, S.; Zhao, Y.; Corti, C.; Ficetola, G.F.; Manenti, R. Photographic database of the European cave salamanders, genus Hydromantes. Sci. Data 2020, 7, 171. [Google Scholar] [CrossRef] [PubMed]
- Lanza, B.; Caputo, V.; Nascetti, G.; Bullini, L. Morphologic and genetic studies of the European plethodontid salamanders: Taxonomic inferences (genus Hydromantes). Monogr. Mus. Reg. Sci. Nat. Torino 1995, 16, 1–366. [Google Scholar]
- Ficetola, G.F.; Lunghi, E.; Cimmaruta, R.; Manenti, R. Transgressive niche across a salamander hybrid zone revealed by microhabitat analyses. J. Biogeogr. 2019, 46, 1342–1354. [Google Scholar] [CrossRef]
- Ruggi, A.; Cimmaruta, R.; Forti, G.; Nascetti, G. Preliminary study of a hybrid zone between Speleomantes italicus Dunn 1923 and S. ambrosii Lanza 1955 on the Apuan Alps, using RLFP analysis. Ann. Mus. Civ. Stor. Nat. Giacomo Doria Genova 2005, 97, 135–144. [Google Scholar]
- Forti, G.; Lanza, B.; Cimmaruta, R.; Nascetti, G. An experiment of artificial syntopy ex situ between Speleomantes italicus (Dunn, 1923) and S. ambrosii ambrosii (Lanza, 1955) (Amphibia, Plethodontidae). Ann. Mus. Civ. Stor. Nat. Giacomo Doria Genova 2005, 97, 123–133. [Google Scholar]
- Cimmaruta, R.; Forti, G.; Lucente, D.; Nascetti, G. Thirty years of artificial syntopy between Hydromantes italicus and H. ambrosii ambrosii (Amphibia, Plethodontidae). Amphib.-Reptil. 2013, 34, 413–420. [Google Scholar] [CrossRef]
- Lunghi, E.; Guillaume, O.; Blaimont, P.; Manenti, R. The first ecological study on the oldest allochthonous population of European cave salamanders (Hydromantes sp.). Amphib.-Reptil. 2018, 39, 113–119. [Google Scholar] [CrossRef] [Green Version]
- Lunghi, E.; Corti, C.; Manenti, R.; Ficetola, G.F. Consider species specialism when publishing datasets. Nat. Ecol. Evol. 2019, 3, 319. [Google Scholar] [CrossRef] [PubMed]
- Lunghi, E.; Bruni, G. Long-term reliability of visual implant elastomers in the Italian cave salamander (Hydromantes italicus). Salamandra 2018, 54, 283–286. [Google Scholar]
- Crovetto, F.; Romano, A.; Salvidio, S. Comparison of two non-lethal methods for dietary studies in terrestrial salamanders. Wildl. Res. 2012, 39, 266–270. [Google Scholar] [CrossRef]
- Lunghi, E.; Giachello, S.; Manenti, R.; Zhao, Y.; Corti, C.; Ficetola, G.F.; Bradley, J.G. The post hoc measurement as a safe and reliable method to age and size plethodontid salamanders. Ecol. Evol. 2020, 10, 11111–11116. [Google Scholar] [CrossRef] [PubMed]
- Lunghi, E.; Cianferoni, F.; Ceccolini, F.; Mulargia, M.; Cogoni, R.; Barzaghi, B.; Cornago, L.; Avitabile, D.; Veith, M.; Manenti, R.; et al. Field-recorded data on the diet of six species of European Hydromantes cave salamanders. Sci. Data 2018, 5, 180083. [Google Scholar] [CrossRef] [Green Version]
- Lunghi, E.; Romeo, D.; Mulargia, M.; Cogoni, R.; Manenti, R.; Corti, C.; Ficetola, G.F.; Veith, M. On the stability of the dorsal pattern of European cave salamanders (genus Hydromantes). Herpetozoa 2019, 32, 249–253. [Google Scholar] [CrossRef] [Green Version]
- Lunghi, E.; Cianferoni, F.; Ceccolini, F.; Veith, M.; Manenti, R.; Mancinelli, G.; Corti, C.; Ficetola, G.F. What shapes the trophic niche of European plethodontid salamanders? PLoS ONE 2018, 13, e0205672. [Google Scholar] [CrossRef] [PubMed]
- Ficetola, G.F.; Lunghi, E.; Canedoli, C.; Padoa-Schioppa, E.; Pennati, R.; Manenti, R. Differences between microhabitat and broad-scale patterns of niche evolution in terrestrial salamanders. Sci. Rep. 2018, 8, 10575. [Google Scholar] [CrossRef]
- Schielzeth, H.; Dingemanse, N.J.; Nakagawa, S.; Westneat, D.F.; Allegue, H.; Teplitsky, C.; Réale, D.; Dochtermann, N.A.; Garamszegi, L.Z.; Araya-Ajoy, Y.G. Robustness of linear mixed-effects models to violations of distributional assumptions. Methods Ecol. Evol. 2020, 11, 1141–1152. [Google Scholar] [CrossRef]
- Labocha, M.K.; Schutz, H.; Hayes, J.P. Which body condition index is best? Oikos 2014, 123, 111–119. [Google Scholar] [CrossRef]
- Băncilă, R.I.; Hartel, T.; Băncilă, R.I.; Smets, J.; Cogălniceanu, D. Comparing three body condition indices in amphibians: A case study of yellow-bellied toad Bombina variegata. Amphib.-Reptil. 2010, 31, 558–562. [Google Scholar] [CrossRef]
- Lunghi, E.; Manenti, R.; Mulargia, M.; Veith, M.; Corti, C.; Ficetola, G.F. Environmental suitability models predict population density, performance and body condition for microendemic salamanders. Sci. Rep. 2018, 8, 7527. [Google Scholar] [CrossRef] [PubMed]
- Lunghi, E.; Manenti, R.; Ficetola, G.F. Seasonal variation in microhabitat of salamanders: Environmental variation or shift of habitat selection? PeerJ 2015, 3, e1122. [Google Scholar] [CrossRef] [Green Version]
- Salvidio, S.; Romano, A.; Oneto, F.; Ottonello, D.; Michelon, R. Different season, different strategies: Feeding ecology of two syntopic forest-dwelling salamanders. Acta Oecol. 2012, 43, 42–50. [Google Scholar]
- Lunghi, E.; Manenti, R.; Cianferoni, F.; Ceccolini, F.; Veith, M.; Corti, C.; Ficetola, G.F.; Mancinelli, G. Interspecific and inter-population variation in individual diet specialization: Do environmental factors have a role? Ecology 2020, 101, e03088. [Google Scholar] [CrossRef]
- Manenti, R.; Lunghi, E.; Ficetola, G.F. Distribution of spiders in cave twilight zone depends on microclimatic features and trophic supply. Invertebr. Biol. 2015, 134, 242–251. [Google Scholar] [CrossRef]
- Lunghi, E.; Ficetola, G.F.; Zhao, Y.; Manenti, R. Are the neglected Tipuloidea crane flies (Diptera) an important component for subterranean environments? Diversity 2020, 12, 333. [Google Scholar] [CrossRef]
- Lunghi, E.; Cianferoni, F.; Ceccolini, F.; Zhao, Y.; Manenti, R.; Corti, C.; Ficetola, G.F.; Mancinelli, G. Same diet, different strategies: Variability of individual feeding habits across three populations of Ambrosi’s cave salamander (Hydromantes ambrosii). Diversity 2020, 12, 180. [Google Scholar] [CrossRef]
- Pierson, T.W.; Fitzpatrick, B.M.; Camp, C.D. Genetic data reveal fine-scale ecological segregation between larval plethodontid salamanders in replicate contact zones. Evol. Ecol. 2021, 35, 309–322. [Google Scholar] [CrossRef]
- Gargiulo, R.; Kull, T.; Fay, M.F. Effective double-digest RAD sequencing and genotyping despite large genome size. Mol. Ecol. Resour. 2021, 21, 1037–1055. [Google Scholar] [CrossRef] [PubMed]
- Lunghi, E.; Manenti, R.; Canciani, G.; Scarì, G.; Pennati, R.; Ficetola, G.F. Thermal equilibrium and temperature differences among body regions in European plethodontid salamanders. J. Therm. Biol. 2016, 60, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Markle, T.M.; Kozak, K.H. Low acclimation capacity of narrow-ranging thermal specialists exposes susceptibility to global climate change. Ecol. Evol. 2018, 8, 4644–4656. [Google Scholar] [CrossRef]
- Vieites, D.R.; Min, M.-S.; Wake, D.B. Rapid diversification and dispersal during periods of global warming by plethodontid salamanders. Proc. Natl. Acad. Sci. USA 2007, 104, 19903–19907. [Google Scholar] [CrossRef] [PubMed] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lunghi, E.; Cianferoni, F.; Merilli, S.; Zhao, Y.; Manenti, R.; Ficetola, G.F.; Corti, C. Ecological Observations on Hybrid Populations of European Plethodontid Salamanders, Genus Speleomantes. Diversity 2021, 13, 285. https://doi.org/10.3390/d13070285
Lunghi E, Cianferoni F, Merilli S, Zhao Y, Manenti R, Ficetola GF, Corti C. Ecological Observations on Hybrid Populations of European Plethodontid Salamanders, Genus Speleomantes. Diversity. 2021; 13(7):285. https://doi.org/10.3390/d13070285
Chicago/Turabian StyleLunghi, Enrico, Fabio Cianferoni, Stefano Merilli, Yahui Zhao, Raoul Manenti, Gentile Francesco Ficetola, and Claudia Corti. 2021. "Ecological Observations on Hybrid Populations of European Plethodontid Salamanders, Genus Speleomantes" Diversity 13, no. 7: 285. https://doi.org/10.3390/d13070285