Biosensor Technology Reveals the Disruption of the Endothelial Barrier Function and the Subsequent Death of Blood Brain Barrier Endothelial Cells to Sodium Azide and Its Gaseous Products

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture of hCMVECs
2.2. Barrier Integrity Measurements Using ECIS ZΘ Technology
2.3. ATP Measurement for Assessment of Cell Viability
2.4. Cell Imaging
2.5. Statistics
3. Results
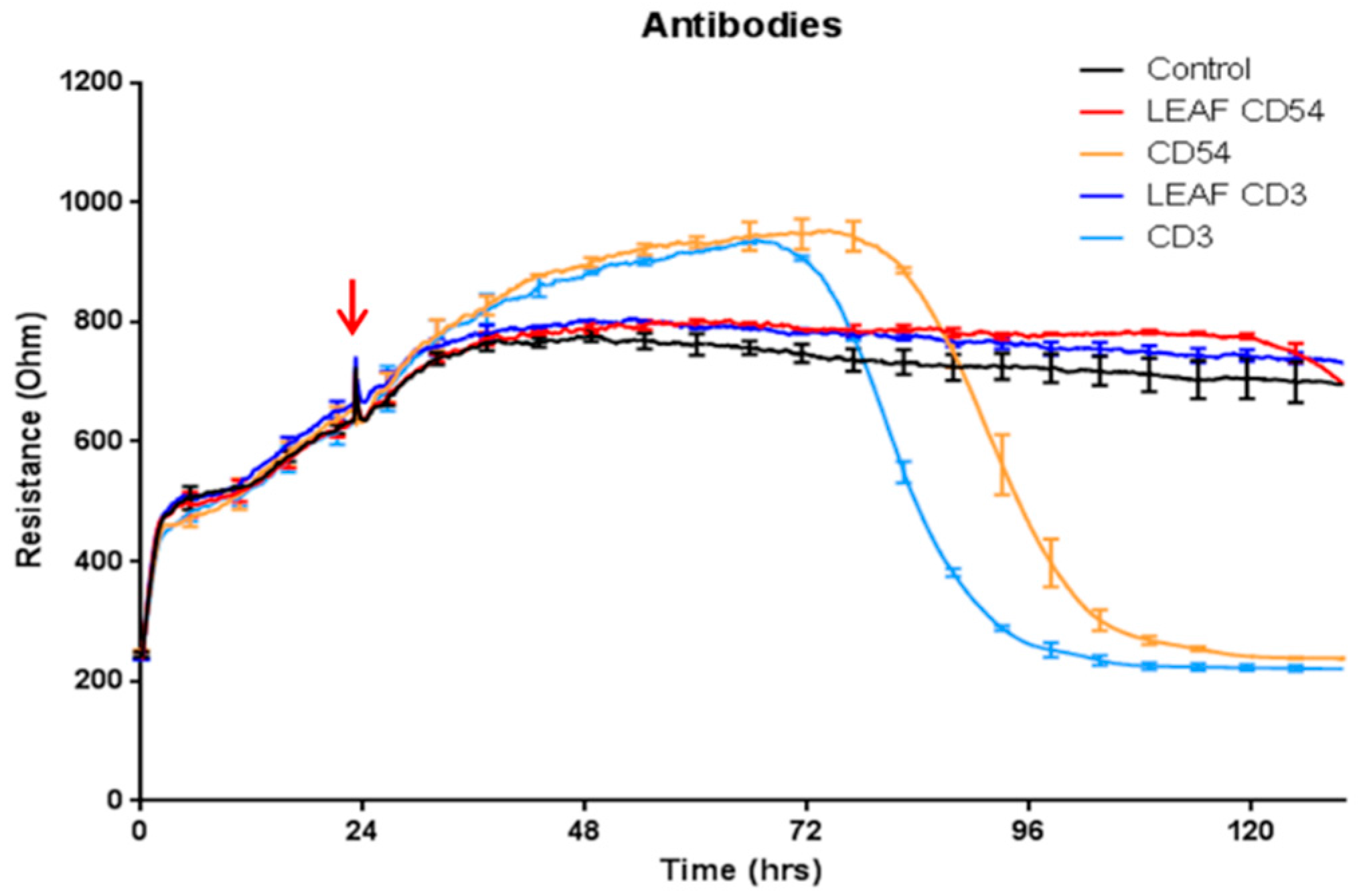
3.1. Antibody Preparations Containing Sodium Azide Mediate Delayed Loss of Endothelial Barrier Resistance, as Measured Using ECIS Technology
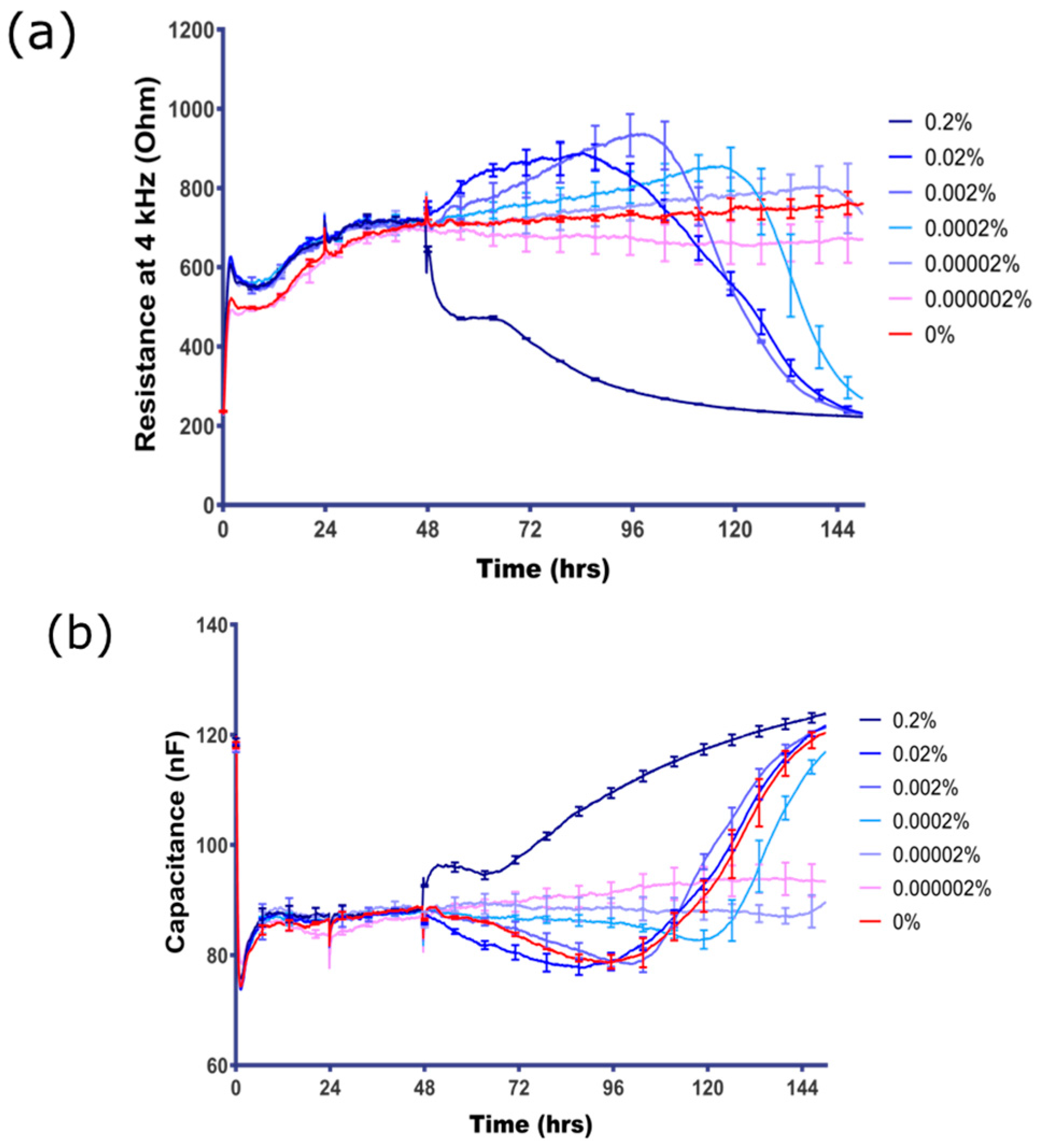
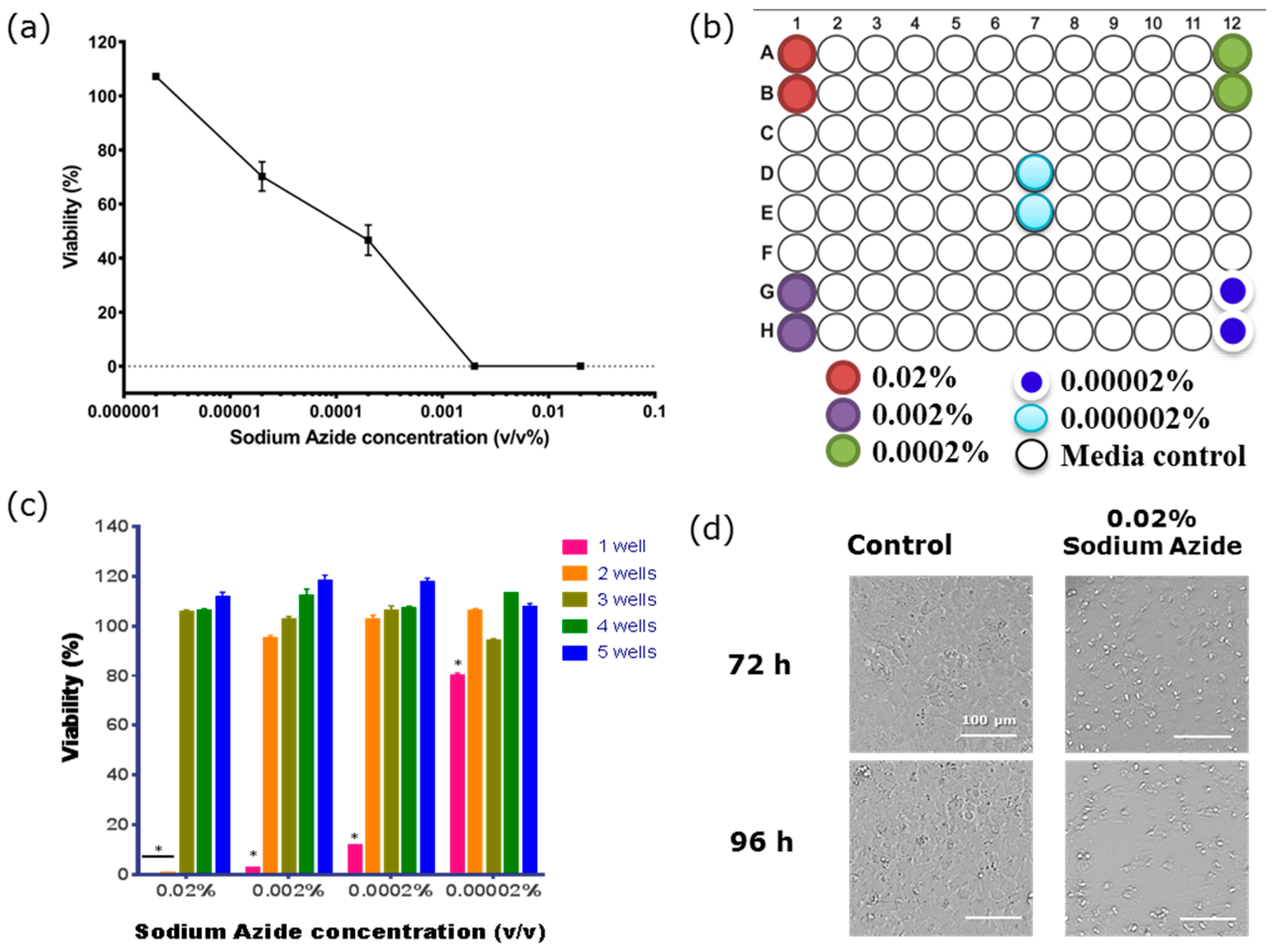
3.2. BBB Endothelial Cells are Highly Sensitive to Sodium Azide
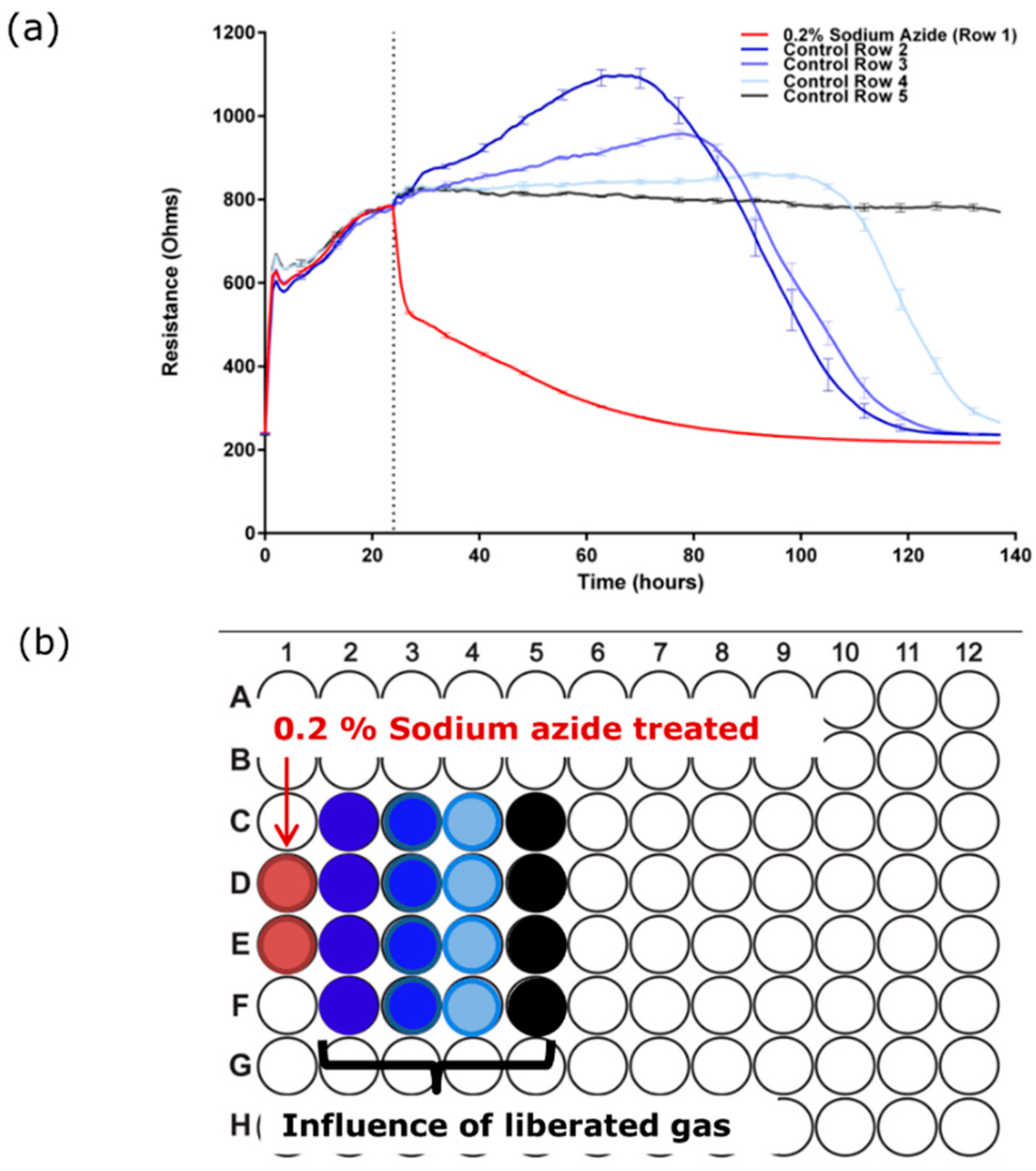
3.3. Sodium Azide Liberates a Gaseous Product that Affects the Endothelial Barrier Integrity
3.4. Determining the Barrier Integrity Loss Due to Direct Effect of the Sodium Azide vs the Gaseous Product
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Zlokovic, B.V. The blood-brain barrier in health and chronic neurodegenerative disorders. Neuron 2008, 57, 178–201. [Google Scholar] [CrossRef] [PubMed]
- Keller, A. Breaking and building the wall: The biology of the blood-brain barrier in health and disease. Swiss Med. Wkly. 2013, 143, w13892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zehendner, C.M.; White, R.; Hedrich, J.; Luhmann, H.J. A neurovascular blood-brain barrier in vitro model. Methods Mol. Biol. 2014, 1135, 403–413. [Google Scholar] [PubMed]
- DePaola, N.; Phelps, J.E.; Florez, L.; Keese, C.R.; Minnear, F.L.; Giaever, I.; Vincent, P. Electrical impedance of cultured endothelium under fluid flow. Ann. Biomed. Eng. 2001, 29, 648–656. [Google Scholar] [CrossRef] [PubMed]
- Arndt, S.; Seebach, J.; Psathaki, K.; Galla, H.J.; Wegener, J. Bioelectrical impedance assay to monitor changes in cell shape during apoptosis. Biosens. Bioelectron. 2004, 19, 583–594. [Google Scholar] [CrossRef]
- Colgan, O.C.; Collins, N.T.; Ferguson, G.; Murphy, R.P.; Birney, Y.A.; Cahill, P.A.; Cummins, P.M. Influence of basolateral condition on the regulation of brain microvascular endothelial tight junction properties and barrier function. Brain Res. 2008, 1193, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Wiltshire, R.; Nelson, V.; Kho, D.T.; Angel, C.E.; O’Carroll, S.J.; Graham, E.S. Regulation of human cerebro-microvascular endothelial baso-lateral adhesion and barrier function by S1P through dual involvement of S1P1 and S1P2 receptors. Sci. Rep. 2016, 6, 19814. [Google Scholar] [CrossRef] [PubMed]
- Thiel, V.E.; Audus, K.L. Nitric oxide and blood-brain barrier integrity. Antioxid. Redox Signal. 2001, 3, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Keese, C.R.; Bhawe, K.; Wegener, J.; Giaever, I. Real-time impedance assay to follow the invasive activities of metastatic cells in culture. Biotechniques 2002, 33, 842–851. [Google Scholar] [PubMed]
- Kho, D.T.; Johnson, R.; Robilliard, L.; Mez, E.D.; Mcintosh, J.; O’Carroll, S.J.; Angel, C.E.; Graham, E.S. ECIS technology reveals that monocytes isolated by CD14+ ve selection mediate greater loss of BBB integrity than untouched monocytes, which occurs to a greater extent with IL-1β activated endothelium in comparison to TNFα. PLoS ONE 2017, 12, e0180267. [Google Scholar] [CrossRef] [PubMed]
- O’Carroll, S.J.; Dan, T.K.; Wiltshire, R.; Nelson, V.; Rotimi, O.; Johnson, R.; Angel, C.E.; Graham, E.S. Pro-inflammatory TNF and IL-1 differentially regulate the inflammatory phenotype of brain microvascular endothelial cells. J. Neuroinflamm. 2015, 12, 131. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Li, L.; Ban, L.; An, W.; Liu, S.; Li, X.; Xue, B.; Xu, Y. Effect of sodium azide on mitochondrial membrane potential in SH-SY5Y human neuroblastoma cells. Zhongguo Yi Xue Ke Xue Yuan Xue Bao 2000, 22, 436–439. [Google Scholar] [PubMed]
- Shityakov, S.; Salmas, R.E.; Salvador, E.; Roewer, N.; Broscheit, J.; Förster, C. Evaluation of the potential toxicity of unmodified and modified cyclodextrins on murine blood-brain barrier endothelial cells. J. Toxicol. Sci. 2016, 41, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Balbuena, P.; Li, W.; Magnin-Bissel, G.; Meldrum, J.B.; Ehrich, M. Comparison of Two Blood-Brain Barrier In Vitro Systems: Cytotoxicity and Transfer Assessments of Malathion/Oxon and Lead Acetate. Toxicol. Sci. 2010, 114, 260–271. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, H.; Hamid, S.S.; Ali, S.S.; Anwar, J.; Siddiqui, A.A.; Khan, N.A. Cytotoxic effects of aflatoxin B1 on human brain microvascular endothelial cells of the blood-brain barrier. Med. Mycol. 2015, 53, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Inomata, K.; Tanaka, H. Protective effect of benidipine against sodium azide-induced cell death in cultured neonatal rat cardiac myocytes. J. Pharmacol. Sci. 2003, 93, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Sato, E.; Suzuki, T.; Hoshi, N.; Sugino, T.; Hasegawa, H. Sodium azide induces necrotic cell death in rat squamous cell carcinoma SCC131. Med. Mol. Morphol. 2008, 41, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Pardridge, W.M. Blood-brain barrier biology and methodology. J. Neurovirol. 1999, 5, 556–569. [Google Scholar] [CrossRef] [PubMed]
- Rubin, L.L.; Staddon, J.M. The cell biology of the blood-brain barrier. Annu. Rev. Neurosci. 1999, 22, 11–28. [Google Scholar] [CrossRef] [PubMed]
- Nicolazzo, J.A.; Katneni, K. Drug transport across the blood-brain barrier and the impact of breast cancer resistance protein (ABCG2). Curr. Top. Med. Chem. 2009, 9, 130–147. [Google Scholar] [CrossRef] [PubMed]
- Cioni, C.; Turlizzi, E.; Zanelli, U.; Oliveri, G.; Annunziata, P. Expression of Tight Junction and Drug Efflux Transporter Proteins in an in vitro Model of Human Blood-Brain Barrier. Front Psychiatry 2012, 3, 47. [Google Scholar] [CrossRef] [PubMed]
- Jacob, A.; Decleves, X.; Scherrmann, J.M. Recent advances in quantitative proteomics as a sensitive tool to quantify drug transporters and drug metabolizing enzymes at the human blood-brain barrier. Biol. Aujourdhui 2012, 206, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.; Lamm, S.H. Human health effects of sodium azide exposure: A literature review and analysis. Int. J. Toxicol. 2003, 22, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Mooradian, A.D. Effect of aging on the blood-brain barrier. Neurobiol. Aging 1988, 9, 31–39. [Google Scholar] [CrossRef]
- Elovaara, I.; Nykyri, E.; Poutianinen, E.; Hokkanen, L.; Raininko, R.; Suni, J. CSF follow-up in HIV-1 infection: Intrathecal production of HIV-specific and unspecific IGG, and beta-2-microglobulin increase with duration of HIV-1 infection. Acta Neurol. Scand. 1993, 87, 388–396. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.Y.; Matsui, M.; Araya, S.; Onai, N.; Matsushima, K.; Saida, T. Chemokine receptors associated with immunity within and outside the central nervous system in early relapsing-remitting multiple sclerosis. J. Neuroimmunol. 2002, 133, 184–192. [Google Scholar] [CrossRef]
- Cucullo, L.; Aumayr, B.; Rapp, E.; Janigro, D. Drug delivery and in vitro models of the blood-brain barrier. Curr. Opin. Drug Discov. Devel. 2005, 8, 89–99. [Google Scholar] [PubMed]
- Taheri, S.; Gasparovic, C.; Shah, N.J.; Rosenberg, G.A. Quantitative measurement of blood-brain barrier permeability in human using dynamic contrast-enhanced MRI with fast T1 mapping. Magn. Reson. Med. 2011, 65, 1036–1042. [Google Scholar] [CrossRef] [PubMed]
- Abdul Muneer, P.M.; Alikunju, S.; Szlachetka, A.M.; Haorah, J. The mechanisms of cerebral vascular dysfunction and neuroinflammation by MMP-mediated degradation of VEGFR-2 in alcohol ingestion. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 1167–1177. [Google Scholar] [CrossRef] [PubMed]
- Miller, F.; Afonso, P.V.; Gessain, A.; Ceccaldi, P.-E. Blood-brain barrier and retroviral infections. Virulence 2012, 3, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Cristante, E.; McArthur, S.; Mauro, C.; Maggioli, E.; Romero, I.A.; Wylezinska-Arridge, M.; Couraud, P.O.; Lopez-Tremoleda, J.; Christian, H.C.; Weksler, B.B.; et al. Identification of an essential endogenous regulator of blood-brain barrier integrity, and its pathological and therapeutic implications. Proc. Natl. Acad. Sci. USA 2013, 110, 832–841. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sodium Azide (%) | Sodium Azide Molarity |
|---|---|
| 0.2% | 30 mM |
| 0.02% | 3 mM |
| 0.002% | 300 µM |
| 0.0002% | 30 µM |
| 0.00002% | 3 µM |
| 0.000002% | 300 nM |
| 0.0000002% | 30 nM |
| Product | Azide Present | Comments |
|---|---|---|
| Primary antibodies | 0.05 to 0.1% 7.5 mM to 15 mM | Majority of primary antibodies where storage is at 4 °C will contain azide |
| Secondary antibodies | 0.05 to 0.1% | Majority of antibodies where storage is at 4 °C will contain azide |
| Flow cytometry antibodies | 0.05 to 0.1% | Some LEAF versions are now available from certain vendors |
| Cell isolation kits (e.g., Miltenyi) | 0.05 to 0.08% | Antibody cocktails and some buffers contain azide |
| ELISA kits | 0.05–0.1% | Storage buffers containing azide; Disposal issues |
| Other assays kits involving buffers and antibody storage | 0.05–0.1% | Storage buffers containing azide; Disposal issues |
| Vehicle airbags | kg quantities | Issues with exposure of the azide resulting in hydrazoic gas formation.Long term environmental effects not known |
| Aircraft escape shuts | >100 kg quantities |
| Sodium Azide Concentration | Direct Toxicity | Gaseous Toxicity |
|---|---|---|
| 0.02% | Yes | Yes |
| 0.002% | Yes | Yes |
| 0.0002% | Yes | Yes |
| 0.00002% | Yes | Minimal |
| 0.000002% | No | No |
| 0.0000002% | No | No |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kho, D.T.; Johnson, R.H.; O’Carroll, S.J.; Angel, C.E.; Graham, E.S. Biosensor Technology Reveals the Disruption of the Endothelial Barrier Function and the Subsequent Death of Blood Brain Barrier Endothelial Cells to Sodium Azide and Its Gaseous Products. Biosensors 2017, 7, 41. https://doi.org/10.3390/bios7040041
Kho DT, Johnson RH, O’Carroll SJ, Angel CE, Graham ES. Biosensor Technology Reveals the Disruption of the Endothelial Barrier Function and the Subsequent Death of Blood Brain Barrier Endothelial Cells to Sodium Azide and Its Gaseous Products. Biosensors. 2017; 7(4):41. https://doi.org/10.3390/bios7040041
Chicago/Turabian StyleKho, Dan T., Rebecca H. Johnson, Simon J. O’Carroll, Catherine E. Angel, and E. Scott Graham. 2017. "Biosensor Technology Reveals the Disruption of the Endothelial Barrier Function and the Subsequent Death of Blood Brain Barrier Endothelial Cells to Sodium Azide and Its Gaseous Products" Biosensors 7, no. 4: 41. https://doi.org/10.3390/bios7040041