Appraisal of Combined Applications of Trichoderma virens and a Biopolymer-Based Biostimulant on Lettuce Agronomical, Physiological, and Qualitative Properties under Variable N Regimes

 ,
,  , ,
, ,  ,
,  , ,
, ,  ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Setup, Design, and Crop Management
2.2. Fungal and Vegetal Biostimulants
2.3. Fungal Colony Forming Units in Soil Rhizosphere and Trichoderma-VBP Compatibility
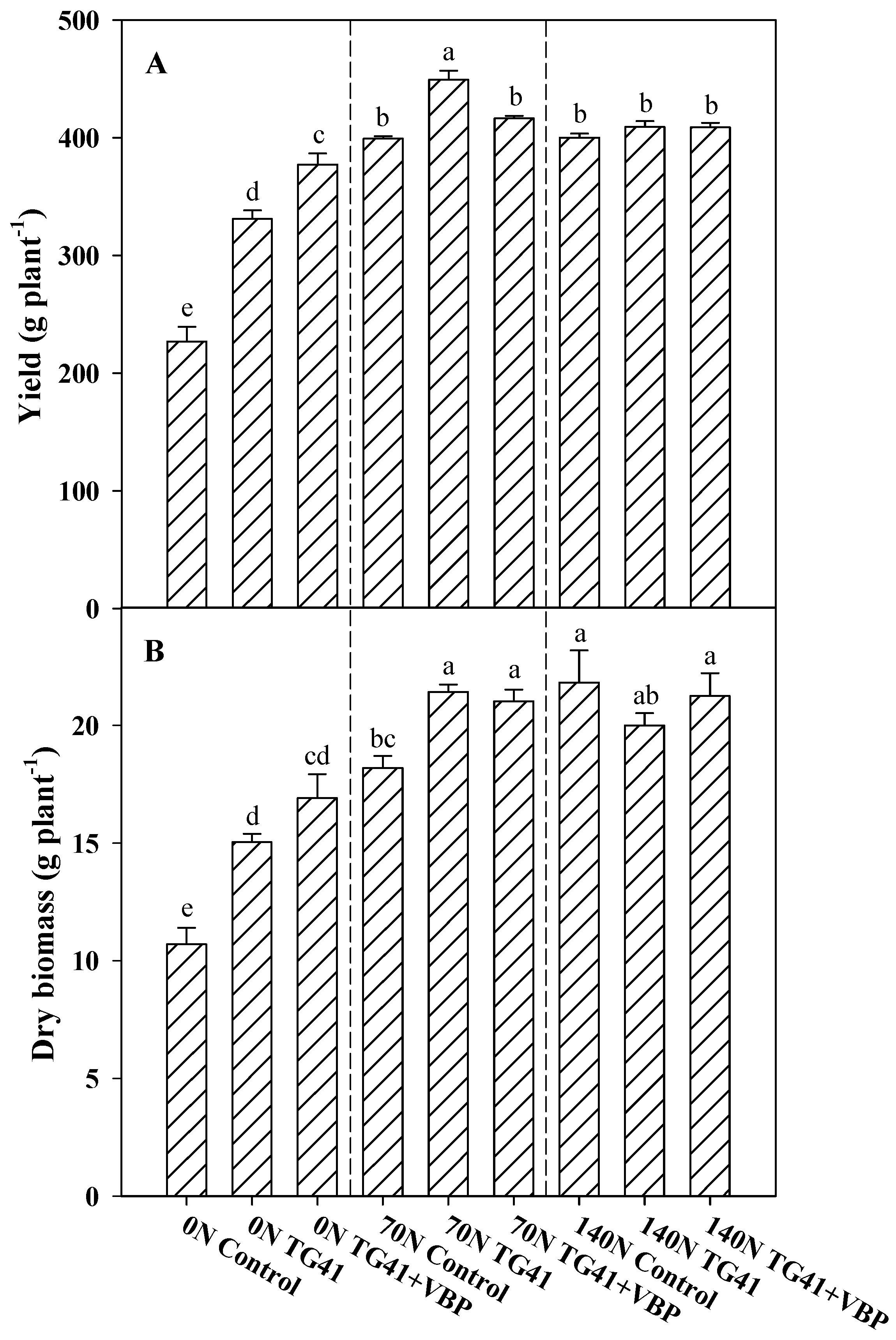
2.4. Fresh and Dry Yield, SPAD index and CIE (lab) Measurements
2.5. Net CO2 Assimilation Rate and Stomatal Resistance Measurements
2.6. Mineral Composition Analysis
2.7. Antioxidant Capacity, Total Phenols, and Total Ascorbic Acid Analysis
2.8. Data Elaboration, Statistical Analysis, Principal Component Analysis, and Heat Map
3. Results
3.1. Fungal Concentration in the Soil
3.2. Growth Responses, SPAD Index and Leaf Colorimetry
3.3. Leaf Gas Exchange: Net CO2 Assimilation Rate and Stomatal Resistance
3.4. Mineral Composition in Leaf Tissue
3.5. Nitrate, Antioxidant Capacity, and Bioactive Content
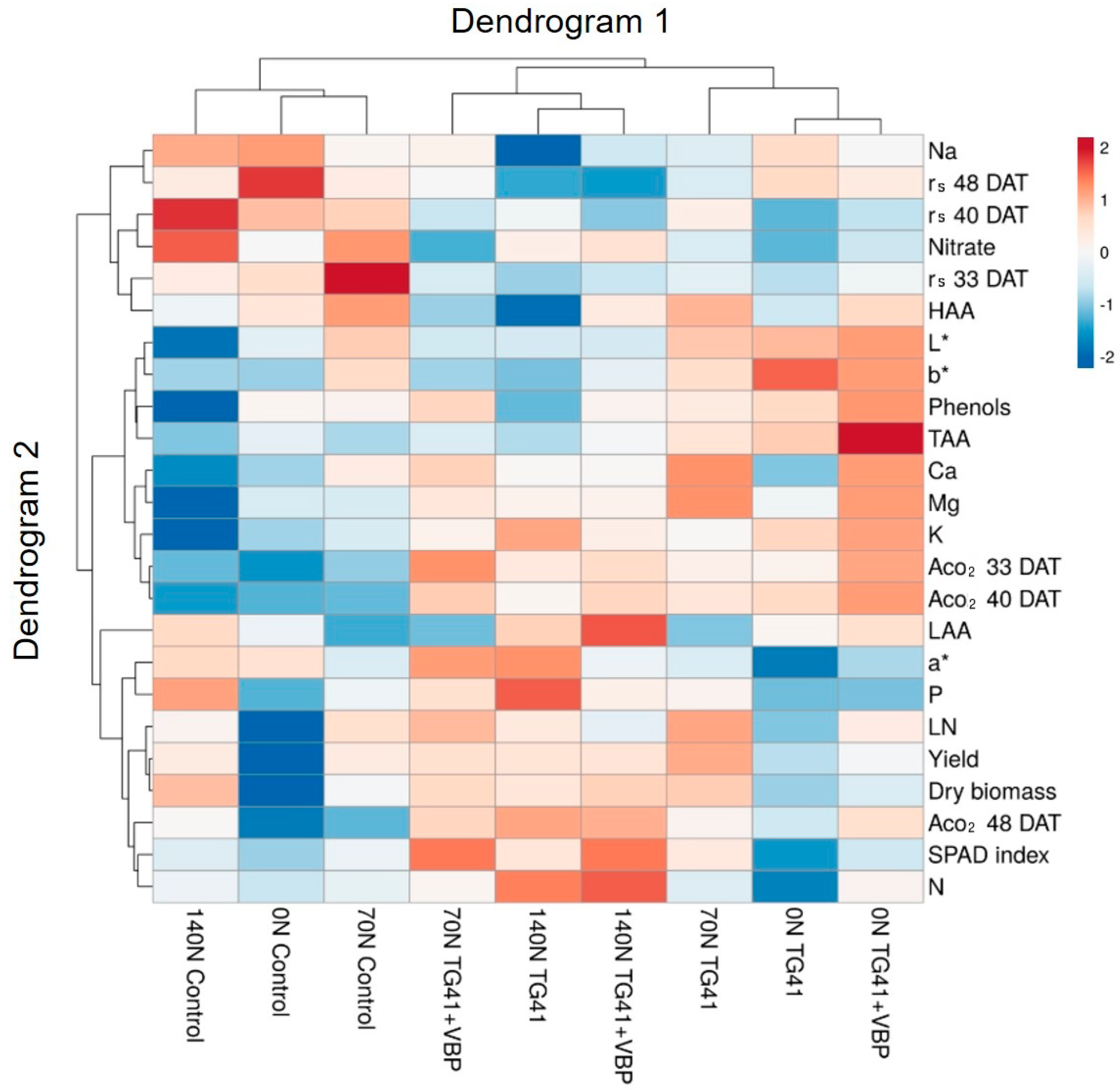
3.6. Heat Map Analysis of all Measured Plant Parameters
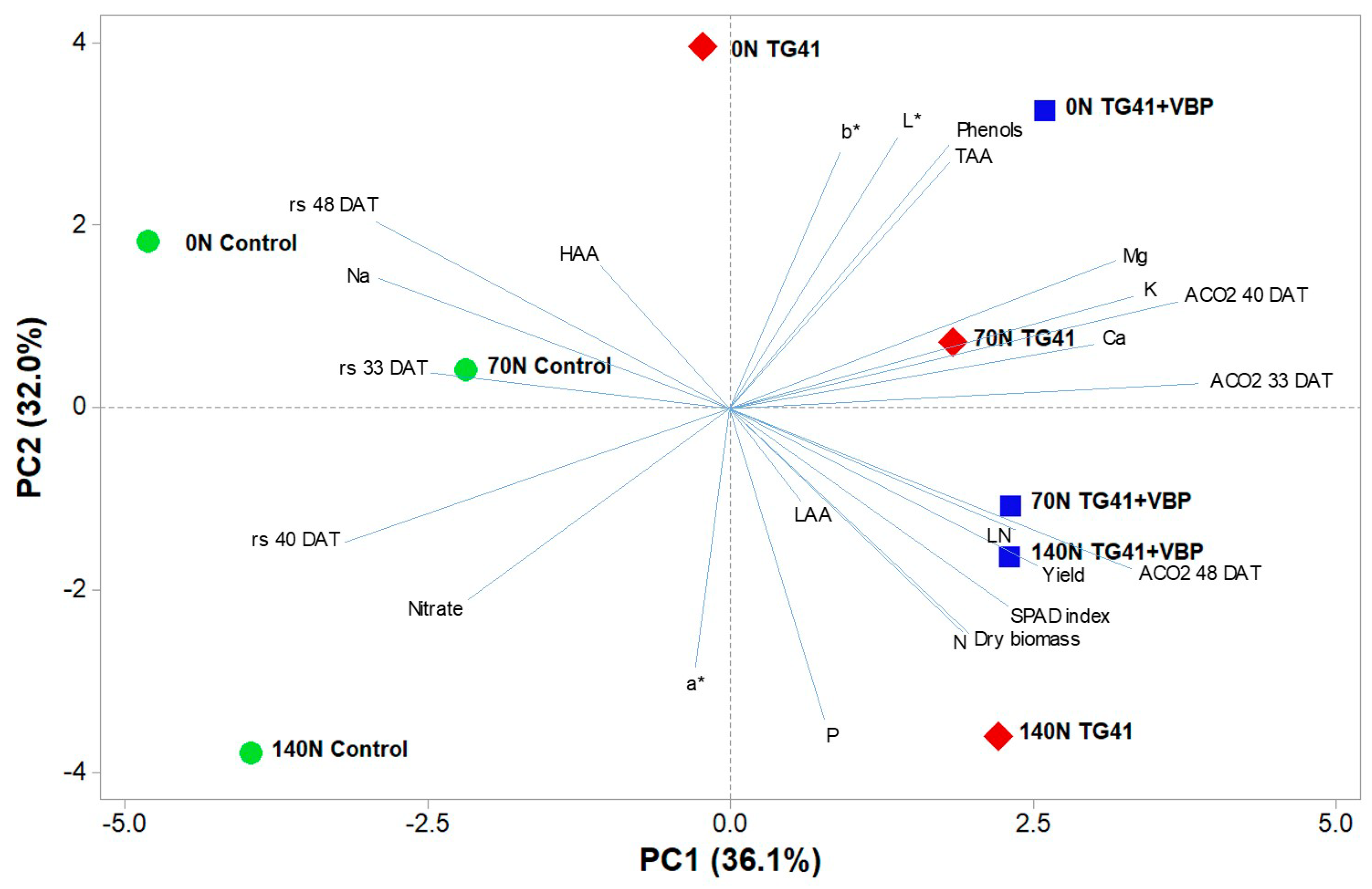
3.7. Principal Component Analysis of all Measured Plant Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAO. Global Agriculture towards 2050, High-Level Expert Forum, How to Feed the World 2050; Food and Agriculture Organization of the United Nations: Rome, Italy, 2009. [Google Scholar]
- D’Antuono, L.F.; Rossini, F. Yield potential and ecophysiological traits of the Altamurano linseed (Linum usitatissimum L.), a landrace of southern Italy. Genet. Resour. Crop Evol. 2006, 53, 65–75. [Google Scholar] [CrossRef]
- Rossini, F.; Loreti, P.; Provenzano, M.E.; De Santis, D.; Ruggeri, R. Agronomic performance and beer quality assessment of twenty hop cultivars grown in central Italy. Ital. J. Agron. 2016, 11, 180–187. [Google Scholar] [CrossRef] [Green Version]
- Lassaletta, L.; Billen, G.; Garnier, J.; Bouwman, L.; Velazquez, E.; Mueller, N.D.; Gerber, J.S. Nitrogen use in the global food system: past trends and future trajectories of agronomic performance, pollution, trade, and dietary demand. Environ. Res. Lett. 2016, 11, 095007. [Google Scholar] [CrossRef]
- FAO. The Future of Food and Agriculture–Alternative Pathways to 2050; Food and Agriculture Organization of the United Nations: Rome, Italy, 2018. [Google Scholar]
- Carillo, P.; Colla, G.; El-Nakhel, C.; Bonini, P.; D’Amelia, L.; Dell’Aversana, E.; Pannico, A.; Giordano, M.; Sifola, M.I.; Kyriacou, M.C.; et al. Biostimulant application with a tropical plant extract enhances Corchorus olitorius adaptation to sub-optimal nutrient regimens by improving physiological parameters. Agronomy 2019, 9, 249. [Google Scholar] [CrossRef] [Green Version]
- Carillo, P.; Colla, G.; Fusco, G.M.; Dell’Aversana, E.; El-Nakhel, C.; Giordano, M.; Pannico, A.; Cozzolino, E.; Mori, M.; Reynaud, H.; et al. Morphological and physiological responses induced by protein hydrolysate-based biostimulant and nitrogen rates in greenhouse spinach. Agronomy 2019, 9, 450. [Google Scholar] [CrossRef] [Green Version]
- Rouphael, Y.; Colla, G. The influence of drip irrigation or subirrigation on zucchini squash grown in closed-loop substrate culture with high and low nutrient solution concentrations. HortScience 2009, 44, 306–311. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.K.; Gupta, A.B.; Gupta, R. Pathophysiology of nitrate toxicity in humans in view of the changing trends of the global nitrogen cycle with special reference to India. In The Indian Nitrogen Assessment; Abrol, Y.P., Adhya, T.K., Aneja, V.P., Eds.; Elsevier: Amsterdam, The Netherland, 2017; pp. 459–468. [Google Scholar] [CrossRef]
- Raun, W.R.; Johnson, G.V. Improving nitrogen use efficiency for cereal production. Agron. J. 1999, 91, 357–363. [Google Scholar] [CrossRef] [Green Version]
- Lassaletta, L.; Billen, G.; Grizzetti, B.; Anglade, J.; Garnier, J. 50 year trends in nitrogen use efficiency of world cropping systems: the relationship between yield and nitrogen input to cropland. Environ. Res. Lett. 2014, 9, 105011. [Google Scholar] [CrossRef]
- Ertani, A.; Cavani, L.; Pizzeghello, D.; Brandellero, E.; Altissimo, A.; Ciavatta, C.; Nardi, S. Biostimulant activity of two protein hydrolyzates in the growth and nitrogen metabolism of maize seedlings. J. Plant Nutr. Soil Sci. 2009, 172, 237–244. [Google Scholar] [CrossRef]
- Colla, G.; Kim, H.J.; Kyriacou, M.; Rouphael, Y. Nitrate in fruits and vegetables. Sci. Hortic. 2018, 237, 221–238. [Google Scholar] [CrossRef]
- Kyriacou, M.C.; Rouphael, Y. Towards a new definition of quality for fresh fruits and vegetables. Sci. Hortic. 2018, 234, 463–469. [Google Scholar] [CrossRef]
- Masclaux-Daubresse, C.; Daniel-Vedele, F.; Dechorgnat, J.; Chardon, F.; Gaufichon, L.; Suzuki, A. Nitrogen uptake, assimilation and remobilization in plants: Challenges for sustainable and productive agriculture. Ann. Bot. 2010, 105, 1141–1157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colla, G.; Rouphael, Y. Biostimulants in horticulture. Sci. Hortic. 2015, 196, 1–2. [Google Scholar] [CrossRef]
- Rouphael, Y.; Spíchal, L.; Panzarová, K.; Casa, R.; Colla, G. High-throughput plant phenotyping for developing novel biostimulants: From lab to field or from field to lab? Front. Plant Sci. 2018, 9, 1197. [Google Scholar] [CrossRef]
- Rouphael, Y.; Kyriacou, M.C.; Petropoulos, S.A.; De Pascale, S.; Colla, G. Improving vegetable quality in controlled environments. Sci. Hortic. 2018, 234, 275–289. [Google Scholar] [CrossRef]
- Du Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef] [Green Version]
- EU. Regulation of the European Parliament and of the council laying down rules on the making available on the market of EU fertilising products and amending Regulations (EC) No 1069/2009 and (EC) No 1107/2009 and repealing Regulation (EC) No 2003/2003. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=OJ:L:2019:170:TOC (accessed on 31 December 2019).
- Canellas, L.P.; Olivares, F.L.; Aguiar, N.O.; Jones, D.L.; Nebbioso, A.; Mazzei, P.; Piccolo, A. Humic and fulvic acids as biostimulants in horticulture. Sci. Hortic. 2015, 196, 15–27. [Google Scholar] [CrossRef]
- Ruzzi, M.; Aroca, R. Plant growth-promoting rhizobacteria act as biostimulants in horticulture. Sci. Hortic. 2015, 196, 124–134. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G. Synergistic Biostimulatory action: designing the next generation of plant biostimulants for sustainable agriculture. Front. Plant Sci. 2018, 9, 1655. [Google Scholar] [CrossRef] [Green Version]
- Colla, G.; Hoagland, L.; Ruzzi, M.; Cardarelli, M.; Bonini, P.; Canaguier, R.; Rouphael, Y. Biostimulant action of protein hydrolysates: unraveling their effects on plant physiology and microbiome. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Fiorentino, N.; Ventorino, V.; Woo, S.L.; Pepe, O.; De Rosa, A.; Gioia, L.; Romano, I.; Lombardi, N.; Napolitano, M.; Colla, G.; et al. Trichoderma-based biostimulants modulate rhizosphere microbial populations and improve N uptake efficiency, yield, and nutritional quality of leafy vegetables. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Bucio, J.; Pelagio-Flores, R.; Herrera-Estrella, A. Trichoderma as biostimulant: exploiting the multilevel properties of a plant beneficial fungus. Sci. Hortic. 2015, 196, 109–123. [Google Scholar] [CrossRef]
- Woo, S.; Ruocco, M.; Vinale, F.; Nigro, M.; Marra, R.; Lombardi, N.; Pascale, A.; Lanzuise, S.; Manganiello, G.; Lorito, M. Trichoderma-based products and their widespread use in agriculture. Open Mycol. J. 2014, 8. [Google Scholar] [CrossRef] [Green Version]
- Vinale, F.; Sivasithamparam, K.; Ghisalberti, E.L.; Marra, R.; Woo, S.L.; Lorito, M. Trichoderma–plant–pathogen interactions. Soil Biol. Biochem. 2008, 40, 1–10. [Google Scholar] [CrossRef]
- Stewart, A.; Hill, R. Applications of Trichoderma in plant growth promotion. In Biotechnology and Biology of Trichoderma; Gupta, V.K., Schmoll, M., Herrera-Estrella, A., Upadhyay, R.S., Druzhinina, I., Tuohy, M.G., Eds.; Elsevier: Amsterdam, The Netherland, 2014; Chapter 31; pp. 415–428. [Google Scholar] [CrossRef]
- Harman, G.E. Overview of new insights into mechanisms and uses of Trichoderma based products. Phytopathology 2004, 94, 138. [Google Scholar]
- Hermosa, R.; Viterbo, A.; Chet, I.; Monte, E. Plant-beneficial effects of Trichoderma and of its genes. Microbiol. 2012, 158, 17–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species–opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2004, 2, 43–56. [Google Scholar] [CrossRef]
- Lorito, M.; Woo, S.L. Trichoderma: A multi-purpose tool for integrated pest management. In Principles of Plant-Microbe Interactions; Lugtenberg, B., Ed.; Springer International Publishing: Manhattan, NY, USA, 2015; pp. 345–353. [Google Scholar] [CrossRef]
- Baker, R. Trichoderma spp. as plant-growth stimulants. CRC Crit. Rev. Biotechnol. 1988, 7, 97–106. [Google Scholar] [CrossRef]
- Altomare, C.; Norvell, W.A.; Bjorkman, T.; Harman, G.E. Solubilization of phosphates and micronutrients by the plant-growth-promoting and biocontrol fungus Trichoderma harzianum Rifai 1295-22. Appl Environ Microbiol. 1999, 65, 2926–2933. [Google Scholar] [CrossRef] [Green Version]
- De Santiago, A.; Quintero, J.M.; Avilés, M.; Delgado, A. Effect of Trichoderma asperellum strain T34 on iron, copper, manganese, and zinc uptake by wheat grown on a calcareous medium. Plant Soil 2011, 342, 97–104. [Google Scholar] [CrossRef]
- Mastouri, F.; Björkman, T.; Harman, G.E. Trichoderma harzianum enhances antioxidant defense of tomato seedlings and resistance to water deficit. Mol. Plant-Microbe Interact. 2012, 25, 1264–1271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colla, G.; Rouphael, Y.; Di Mattia, E.; El-Nakhel, C.; Cardarelli, M. Co-inoculation of Glomus intraradices and Trichoderma atroviride acts as a biostimulant to promote growth, yield and nutrient uptake of vegetable crops. J. Sci. Food Agric. 2015, 95, 1706–1715. [Google Scholar] [CrossRef] [PubMed]
- Woo, S.L.; Scala, F.; Ruocco, M.; Lorito, M. The molecular biology of the interactions between Trichoderma spp., phytopathogenic fungi, and plants. Phytopathology 2006, 96, 181–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manganiello, G.; Sacco, A.; Ercolano, M.R.; Vinale, F.; Lanzuise, S.; Pascale, A.; Napolitano, M.; Lombardi, N.; Lorito, M.; Woo, S.L. Modulation of tomato response to Rhizoctonia solani by Trichoderma harzianum and its secondary metabolite harzianic acid. Front. Microbiol. 2018, 9, 1966. [Google Scholar] [CrossRef] [PubMed]
- Contreras-Cornejo, H.A.; Macías-Rodríguez, L.; Cortés-Penagos, C.; López-Bucio, J. Trichoderma virens, a plant beneficial fungus, enhances biomass production and promotes lateral root growth through an auxin-dependent mechanism in Arabidopsis. Plant Physiol. 2009, 149, 1579–1592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prisa, D.; Sarrocco, S.; Vannacci, G. Microrganismi biostimolanti: Trichoderma sp. in substrati innovativi e alternativi alla torba per piante ornamentali. Italus Hort. 2014, 17–28. [Google Scholar]
- Salwan, R.; Kumari, N.; Sharma, V. Bioactive Volatile Metabolites of Trichoderma: An overview. In Secondary Metabolites Plant Growth Promoting Rhizomicroorganisms; Singh, H., Keswani, C., Reddy, M., Sansinenea, E., García-Estrada, C., Eds.; Springer: Singapore, 2019; pp. 87–111. [Google Scholar] [CrossRef]
- Coppola, M.; Diretto, G.; Digilio, M.C.; Woo, S.L.; Giuliano, G.; Molisso, D.; Pennacchio, F.; Lorito, M.; Rao, R. Transcriptome and metabolome reprogramming in tomato plants by Trichoderma harzianum strain T22 primes and enhances defense responses against aphids. Front. Physiol. 2019, 10. [Google Scholar] [CrossRef]
- Ertani, A.; Francioso, O.; Tugnoli, V.; Righi, V.; Nardi, S. Effect of commercial lignosulfonate-humate on Zea mays L. metabolism. J. Agric. Food Chem. 2011, 59, 11940–11948. [Google Scholar] [CrossRef]
- Lucini, L.; Rouphael, Y.; Cardarelli, M.; Bonini, P.; Baffi, C.; Colla, G. A vegetal biopolymer-based biostimulant promoted root growth in melon while triggering brassinosteroids and stress-related compounds. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Broadley, M.R.; Escobar-Gutièrrez, A.J.; Burns, A.; Burns, G. What are the effects of nitrogen deficiency on growth components of lettuce? New Phytol. 2000, 147, 519–526. [Google Scholar] [CrossRef]
- Bettoni, M.M.; Mogor, A.F.; Pauletti, V.; Goicoechea, N. Growth and metabolism of onion seedlings as affected by the application of humic substances, mycorrhizal inoculation and elevated CO2. Sci. Hortic. 2014, 180, 227–235. [Google Scholar] [CrossRef]
- Nikbakht, A.; Pessarakli, M.; Daneshvar-Hakimi-Maibodi, N.; Kafi, M. Perennial ryegrass growth responses to mycorrhizal infection and humic acid treatments. Agron. J. 2014, 106, 585–595. [Google Scholar] [CrossRef]
- Rouphael, Y.; Cardarelli, M.; Bonini, P.; Colla, G. Synergistic action of a microbial-based biostimulant and a plant derived-protein hydrolysate enhances lettuce tolerance to alkalinity and salinity. Front. Plant Sci. 2017, 8, 131. [Google Scholar] [CrossRef] [Green Version]
- Kumar, P.; Lucini, L.; Rouphael, Y.; Cardarelli, C.; Kalunke, R.M.; Colla, G. Insight into the role of grafting and arbuscular mycorrhiza on cadmium stress tolerance in tomato. Front. Plant Sci. 2015, 6, 477. [Google Scholar] [CrossRef] [Green Version]
- Colonna, E.; Rouphael, Y.; Barbieri, G.; De Pascale, S. Nutritional quality of leafy vegetables harvested at two light intensities. Food Chem. 2016, 199, 702–710. [Google Scholar] [CrossRef]
- Bremner, J.M. Total nitrogen. In Methods of Soil Analysis; Black, C.A., Evans, D.D., White, D.D., Ensminger, E., Clark, F.E., Eds.; American Society of Agronomy: Madison, WI, USA, 1965; pp. 149–1178. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved Abts radical cation decolorization assay. Free Rad. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Fogliano, V.; Verde, V.; Randazzo, G.; Ritieni, A. Method for measuring antioxidant activity and its application to monitoring the antioxidant capacity of wines. J. Agric. Food Chem. 1999, 47, 1035–1040. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventos, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteu reagent. Oxidants Antioxidants Part A. Meth. Enzymol. 1999, 299, 152–178. [Google Scholar] [CrossRef]
- Kampfenkel, K.; Van Montagu, M.; Inzé, D. Extraction and determination of ascorbate and dehydroascorbate from plant tissue. Anal. Biochem. 1995, 225, 165–167. [Google Scholar] [CrossRef]
- Rouphael, Y.; Kyriacou, M.C.; Carillo, P.; Pizzolongo, F.; Romano, R.; Sifola, M.I. Chemical eustress elicits tailored responses and enhances the functional quality of novel food Perilla Frutescens. Molecules 2019, 24, 185. [Google Scholar] [CrossRef] [Green Version]
- El-Nakhel, C.; Petropoulos, S.A.; Pannico, A.; Kyriacou, M.C.; Giordano, M.; Colla, G.; Troise, A.D.; Vitaglione, P.; De Pascale, S.; Rouphael, Y. The bioactive profile of lettuce produced in a closed soilless system as configured by combinatorial effects of genotype and macrocation supply composition. Food Chem. 2020, in press. [Google Scholar] [CrossRef] [PubMed]
- El-Nakhel, C.; Pannico, A.; Kyriacou, M.C.; Giordano, M.; De Pascale, S.; Rouphael, Y. Macronutrient deprivation eustress elicits differential secondary metabolites in red and green-pigmented butterhead lettuce grown in closed soilless system. J. Sci. Food Agric. 2019, 99, 6962–6972. [Google Scholar] [CrossRef] [PubMed]
- Kyriacou, M.C.; Rouphael, Y.; Colla, G.; Zrenner, R.M.; Schwarz, D. Vegetable grafting: The implications of a growing agronomic imperative for vegetable fruit quality and nutritive value. Front. Plant Sci. 2017, 8, 741. [Google Scholar] [CrossRef] [PubMed]
- Bassi, D.; Menossi, M.; Mattiello, L. Nitrogen supply influences photosynthesis establishment along the sugarcane leaf. Sci. Rep. 2018, 8, 2327. [Google Scholar] [CrossRef] [Green Version]
- Van Oosten, M.J.; Dell’Aversana, E.; Ruggiero, A.; Cirillo, V.; Gibon, Y.; Woodrow, P.; Maggio, A.; Carillo, P. Omeprazole treatment enhances nitrogen use efficiency through increased nitrogen uptake and assimilation in corn. Front. Plant Sci. 2019, 10, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.H.; Li, X.P.; Razeghifard, R.; Anderson, J.M.; Niyogi, K.K.; Pogson, B.J.; Chow, W.S. The multiple roles of light-harvesting chlorophyll a/b-protein complexes define structure and optimize function of Arabidopsis chloroplasts: A study using two chlorophyll b-less mutants. Biochim. Biophys. Acta 2009, 1787, 973–984. [Google Scholar] [CrossRef] [Green Version]
- Di Mola, I.; Ottaiano, L.; Cozzolino, E.; Senatore, M.; Giordano, M.; El-Nakhel, C.; Sacco, A.; Rouphael, Y.; Colla, G.; Mori, M. Plant-based biostimulants influence the agronomical, physiological, and qualitative responses of baby rocket leaves under diverse nitrogen conditions. Plants 2019, 8, 522. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.H.; Li, S.X.; Malhi, S. Effects of fertilization and other agronomic measures on nutritional quality of crops. J. Sci. Food Agric. 2008, 88, 7–323. [Google Scholar] [CrossRef]
- Hermosa, R.; Rubio, B.; Cardoza, R.E.; Nicolas, C.; Monte, E.; Gutièrrez, S. The contribution of Trichoderma to balancing the costs of plant growth and defense. Int. Microbio. 2013, 69–80. [Google Scholar] [CrossRef]
- Colla, G.; Rouphael, Y.; Canaguier, R.; Svecova, E.; Cardarelli, M. Biostimulant action of a plant-derived protein hydrolysateproduced through enzymatic hydrolysis. Front. Plant Sci. 2014, 5. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Treatments | SPAD Index | L | a* | b* | |
|---|---|---|---|---|---|
| 45 DAT | 75 DAT | ||||
| Nitrogen rate (N) | NS | *** | ** | NS | * |
| Biostimulant (B) | * | ** | NS | NS | NS |
| N × B | NS | NS | NS | NS | * |
| Nitrogen rate (kg ha−1) | |||||
| 0 | 38.27 | 37.92 b | 42.81 a | −16.61 | 24.53 a |
| 70 | 38.94 | 39.35 a | 42.60 a | −16.37 | 24.20 ab |
| 140 | 38.47 | 39.24 a | 41.54 b | −16.23 | 23.62 b |
| Biostimulant | |||||
| Control | 37.54 b | 38.39 b | 41.94 | −16.33 | 23.86 |
| TG41 | 39.50 a | 38.56 b | 42.65 | −16.49 | 24.37 |
| TG41+VBP | 38.63 a | 39.50 a | 42.36 | −16.38 | 24.13 |
| N × B | |||||
| 0N Control | 37.23 | 38.00 | 42.08 | −16.25 | 23.50 c |
| 0N TG41 | 39.30 | 37.45 | 43.08 | −16.93 | 25.18 a |
| 0N TG41+VBP | 38.27 | 38.30 | 43.28 | −16.65 | 24.93 ab |
| 70N Control | 37.50 | 38.69 | 42.95 | −16.53 | 24.55 abc |
| 70N TG41 | 40.60 | 39.15 | 42.98 | −16.53 | 24.53 abc |
| 70N TG41+VBP | 38.73 | 40.15 | 41.88 | −16.05 | 23.53 bc |
| 140N Control | 37.90 | 38.48 | 40.80 | −16.20 | 23.53 bc |
| 140N TG41 | 38.60 | 39.23 | 41.90 | −16.03 | 23.40 c |
| 140N TG41+VBP | 38.90 | 40.14 | 41.93 | −16.45 | 23.93 abc |
| Treatments | Net CO2 Assimilation Rate (μmol CO2 m−2 s−1) | Stomatal Resistance (m2 s1 mol−1) | ||||
|---|---|---|---|---|---|---|
| 33 DAT | 40 DAT | 48 DAT | 33 DAT | 40 DAT | 48 DAT | |
| Nitrogen rate (N) | NS | NS | *** | NS | NS | NS |
| Biostimulant (B) | *** | *** | *** | NS | * | NS |
| N × B | NS | NS | NS | NS | NS | NS |
| Nitrogen rate (kg ha−1) | ||||||
| 0 | 14.51 | 21.26 | 13.20 c | 3.32 | 4.06 | 4.35 |
| 70 | 15.16 | 20.93 | 14.12 b | 3.94 | 4.40 | 3.80 |
| 140 | 14.61 | 20.45 | 15.78 a | 2.96 | 4.52 | 3.36 |
| Biostimulant | ||||||
| Control | 11.74 b | 18.49 b | 12.48 c | 4.59 | 5.25 a | 4.27 |
| TG41 | 15.25 a | 21.60 a | 14.76 b | 2.77 | 4.03 b | 3.63 |
| TG41+BVP | 17.34 a | 22.55 a | 15.86 a | 3.02 | 3.70 b | 3.61 |
| N × B | ||||||
| 0N Control | 10.92 | 18.59 | 10.93 | 4.01 | 5.05 | 4.83 |
| 0N TG41 | 15.08 | 22.12 | 13.23 | 2.64 | 3.37 | 4.20 |
| 0N TG41+VBP | 17.52 | 23.08 | 15.44 | 3.29 | 3.77 | 4.01 |
| 70N Control | 12.44 | 18.73 | 12.06 | 5.80 | 4.91 | 3.98 |
| 70N TG41 | 15.17 | 21.67 | 14.54 | 3.11 | 4.49 | 3.61 |
| 70N TG41+VBP | 17.86 | 22.38 | 15.76 | 2.93 | 3.79 | 3.82 |
| 140N Control | 11.94 | 18.15 | 14.44 | 3.67 | 5.79 | 4.01 |
| 140N TG41 | 15.60 | 21.01 | 16.52 | 2.46 | 4.23 | 3.08 |
| 140N TG41+VBP | 16.29 | 22.20 | 16.37 | 2.74 | 3.53 | 3.01 |
| Treatments | N | P | K | Ca | Mg | Na |
|---|---|---|---|---|---|---|
| (mg g−1 dw) | (mg g−1 dw) | (mg g−1 dw) | (mg g−1 dw) | (mg g−1 dw) | (mg g−1 dw) | |
| Nitrogen rate (N) | ** | *** | NS | NS | NS | NS |
| Biostimulant (B) | NS | NS | ** | NS | * | NS |
| N × B | NS | NS | NS | NS | NS | NS |
| Nitrogen rate (kg ha−1) | ||||||
| 0 | 36.90 b | 2.02 b | 55.02 | 6.83 | 3.63 | 1.52 |
| 70 | 37.63 b | 2.58 a | 53.71 | 7.53 | 3.69 | 1.42 |
| 140 | 39.04 a | 2.93 a | 53.40 | 6.62 | 3.42 | 1.35 |
| Biostimulant | ||||||
| Control | 37.43 | 2.48 | 50.65 b | 6.47 | 3.31 b | 1.54 |
| TG41 | 37.57 | 2.59 | 55.90 a | 7.06 | 3.70 a | 1.34 |
| TG41+VBP | 38.58 | 2.46 | 55.57 a | 7.46 | 3.73 a | 1.41 |
| N × B | ||||||
| 0N Control | 37.03 | 1.98 | 51.36 | 6.39 | 3.46 | 1.60 |
| 0N TG41 | 35.70 | 2.03 | 56.17 | 6.27 | 3.55 | 1.52 |
| 0N TG41+VBP | 37.98 | 2.05 | 57.53 | 7.84 | 3.90 | 1.43 |
| 70N Control | 37.60 | 2.45 | 52.59 | 7.18 | 3.46 | 1.44 |
| 70N TG41 | 37.38 | 2.55 | 54.09 | 7.88 | 3.92 | 1.38 |
| 70N TG41+VBP | 37.93 | 2.75 | 54.45 | 7.53 | 3.69 | 1.45 |
| 140N Control | 37.65 | 3.00 | 48.01 | 5.85 | 3.01 | 1.58 |
| 140N TG41 | 39.63 | 3.20 | 57.45 | 7.02 | 3.62 | 1.13 |
| 140N TG41+VBP | 39.85 | 2.58 | 54.75 | 7.00 | 3.62 | 1.35 |
| Treatments | Nitrate | HAA | LAA | Phenols | TAA |
|---|---|---|---|---|---|
| (mg kg−1 fw) | (mmol eq. ascorbic acid 100g−1 dw) | (mmol eq. trolox 100g−1 dw) | (mg eq. gallic acid g−1 dw) | (mg 100g−1 fw) | |
| Nitrogen rate (N) | * | NS | *** | *** | *** |
| Biostimulant (B) | ** | NS | *** | * | *** |
| N × B | NS | NS | NS | NS | *** |
| Nitrogen rate (kg ha−1) | |||||
| 0 | 1019.09 b | 1.54 | 4.06 b | 55.94 a | 22.66 a |
| 70 | 1119.46 ab | 1.56 | 2.84 c | 54.44 a | 13.81 b |
| 140 | 1319.38 a | 1.47 | 4.88 a | 47.38 b | 11.20 c |
| Biostimulant | |||||
| Control | 1356.88 a | 1.56 | 3.66 b | 49.39 b | 10.53 c |
| TG41 | 1052.41 b | 1.47 | 3.85 b | 52.28 ab | 16.97 b |
| TG41+VBP | 1048.64 b | 1.53 | 4.26 a | 56.10 a | 20.17 a |
| N × B | |||||
| 0N Control | 1152.55 | 1.56 | 3.74 | 52.84 | 13.88 de |
| 0N TG41 | 890.63 | 1.47 | 3.99 | 56.07 | 21.73 b |
| 0N TG41+VBP | 1014.10 | 1.58 | 4.44 | 58.91 | 32.36 a |
| 70N Control | 1422.03 | 1.63 | 2.69 | 53.09 | 9.54 ef |
| 70N TG41 | 1063.18 | 1.61 | 2.95 | 54.06 | 19.27 bc |
| 70N TG41+VBP | 873.18 | 1.44 | 2.88 | 56.18 | 12.61 def |
| 140N Control | 1496.08 | 1.51 | 4.56 | 42.23 | 8.15 f |
| 140N TG41 | 1203.43 | 1.35 | 4.62 | 46.71 | 9.91 ef |
| 140N TG41+VBP | 1258.65 | 1.55 | 5.45 | 53.19 | 15.54 cd |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rouphael, Y.; Carillo, P.; Colla, G.; Fiorentino, N.; Sabatino, L.; El-Nakhel, C.; Giordano, M.; Pannico, A.; Cirillo, V.; Shabani, E.; et al. Appraisal of Combined Applications of Trichoderma virens and a Biopolymer-Based Biostimulant on Lettuce Agronomical, Physiological, and Qualitative Properties under Variable N Regimes. Agronomy 2020, 10, 196. https://doi.org/10.3390/agronomy10020196
Rouphael Y, Carillo P, Colla G, Fiorentino N, Sabatino L, El-Nakhel C, Giordano M, Pannico A, Cirillo V, Shabani E, et al. Appraisal of Combined Applications of Trichoderma virens and a Biopolymer-Based Biostimulant on Lettuce Agronomical, Physiological, and Qualitative Properties under Variable N Regimes. Agronomy. 2020; 10(2):196. https://doi.org/10.3390/agronomy10020196
Chicago/Turabian StyleRouphael, Youssef, Petronia Carillo, Giuseppe Colla, Nunzio Fiorentino, Leo Sabatino, Christophe El-Nakhel, Maria Giordano, Antonio Pannico, Valerio Cirillo, Edris Shabani, and et al. 2020. "Appraisal of Combined Applications of Trichoderma virens and a Biopolymer-Based Biostimulant on Lettuce Agronomical, Physiological, and Qualitative Properties under Variable N Regimes" Agronomy 10, no. 2: 196. https://doi.org/10.3390/agronomy10020196