 Sara Artigas-Jerónimo1†
Sara Artigas-Jerónimo1† Margarita Villar1†
Margarita Villar1† Alejandro Cabezas-Cruz2
Alejandro Cabezas-Cruz2 James J. Valdés3,4,5
James J. Valdés3,4,5 Agustín Estrada-Peña6
Agustín Estrada-Peña6 Pilar Alberdi1
Pilar Alberdi1 José de la Fuente1,7*
José de la Fuente1,7*- 1SaBio, Instituto de Investigación en Recursos Cinegéticos (IREC), CSIC, Universidad de Castilla-La Mancha (UCLM), Junta de Comunidades de Castilla – La Mancha (JCCM), Ciudad Real, Spain
- 2UMR BIPAR, INRA, ANSES, Ecole Nationale Vétérinaire d’Alfort, Université Paris-Est, Paris, France
- 3Faculty of Science, University of South Bohemia, České Budějovice, Czechia
- 4Institute of Parasitology, Biology Centre, Czech Academy of Sciences, České Budějovice, Czechia
- 5Department of Virology, Veterinary Research Institute, Brno, Czechia
- 6Facultad de Veterinaria, Universidad de Zaragoza, Zaragoza, Spain
- 7Department of Veterinary Pathobiology, Center for Veterinary Health Sciences, Oklahoma State University, Stillwater, OK, United States
The Subolesin/Akirin constitutes a good model for the study of functional evolution because these proteins have been conserved throughout the metazoan and play a role in the regulation of different biological processes. Here, we investigated the evolutionary history of Subolesin/Akirin with recent results on their structure, protein-protein interactions and function in different species to provide insights into the functional evolution of these regulatory proteins, and their potential as vaccine antigens for the control of ectoparasite infestations and pathogen infection. The results suggest that Subolesin/Akirin evolved conserving not only its sequence and structure, but also its function and role in cell interactome and regulome in response to pathogen infection and other biological processes. This functional conservation provides a platform for further characterization of the function of these regulatory proteins, and how their evolution can meet species-specific demands. Furthermore, the conserved functional evolution of Subolesin/Akirin correlates with the protective capacity shown by these proteins in vaccine formulations for the control of different arthropod and pathogen species. These results encourage further research to characterize the structure and function of these proteins, and to develop new vaccine formulations by combining Subolesin/Akirin with interacting proteins for the control of multiple ectoparasite infestations and pathogen infection.
Introduction
Akirin, from the Japanese “akiraka ni suru" meaning “making things clear," was first identified by Goto et al. (2008) as a key component of the immune deficiency (IMD) and Tumor necrosis factor (TNF)/Toll-like receptor (TLR)-nuclear factor-kappa B (NF-kB) (TNF/TLR) signaling pathways in Drosophila melanogaster and Mus musculus, respectively. However, previous reports identified akirin as a gene involved in developmental processes in flies (Peña-Rangel et al., 2002; DasGupta et al., 2005). Subolesin, from the Latin “suboles" meaning “progeny," was first reported in 2003 with its discovery as the candidate protective antigen 4D8 by expression library immunization in Ixodes scapularis (Almazán et al., 2003). Gene orthology is a key concept in functional evolution (Koonin, 2005). Orthologs genes, defined as derived from a single ancestral gene that diverged during speciation, usually perform equivalent or identical functions, while paralogs that originated after gene duplication are considered to have more divergent functions (Koonin, 2005; Adipietro et al., 2012). Studies at genome level have identified many orthologs genes between divergent species, but the functional equivalency of the proteins encoded by these genes has not been fully characterized (Koonin, 2005).
Subolesin/Akirin are encoded by orthologs evolutionarily conserved throughout the metazoan that play a role in the regulation of different biological processes including immune response (de la Fuente et al., 2006a; Beutler and Moresco, 2008; Goto et al., 2008; Galindo et al., 2009; Macqueen, 2009; Macqueen and Johnston, 2009; Wan and Lenardo, 2010). Two akirin paralogs encoding Akirin1 and Akirin2 have been identified in vertebrates, but the functional homolog for invertebrate Subolesin/Akirin appears to be Akirin2 ortholog (Beutler and Moresco, 2008; Goto et al., 2008; Galindo et al., 2009; Macqueen and Johnston, 2009).
Understanding the function of the cell interactome (protein-protein physical and functional interactions) and regulome (transcription factors-target genes interactions) in response to infection is critical toward a better understanding of host-pathogen interactions and the identification of potential targets for new interventions for the prevention and control of tick infestations and tick-borne diseases (Rioualen et al., 2017; de la Fuente, 2018). Subolesin/Akirin are involved in both cell interactome and regulome, and constitute a good model for the study of the functional evolution of these processes in response to infection. In this review, we integrated the evolutionary history of Subolesin/Akirin with recent results on their structure and function in different species to provide insights into the functional evolution of these regulatory proteins, and their potential as vaccine antigens for the control of ectoparasite infestations and pathogen infection.
Evolution of Subolesin/Akirin
The phylogenetic analysis of subolesin/akirin coding sequences using an updated sequence database (Figure 1A and Supplementary Figure S1) expanded the information on the evolution of these genes, and supported the results reported previously by Macqueen and Johnston (2009) that akirin1 and akirin2 are vertebrate-specific paralogs that form a separate clade from invertebrate subolesin/akirin. In some vertebrate species, Akirins constitute a family of paralog proteins that probably originated as a result of whole-genome duplications (Macqueen and Johnston, 2009; Macqueen et al., 2010a,b; Liu et al., 2015). After akirin duplication, akirin1 evolved faster than akirin2, the ortholog of tick subolesin (Figure 1A; Macqueen and Johnston, 2009). Furthermore, the loss of some akirin paralogs may have also occurred after genome duplications (Macqueen et al., 2010b; Liu et al., 2015). For example, the subolesin/akirin gene family consists of a single member in invertebrates (subolesin/akirin), birds and reptiles (subolesin/akirin2), two members in amphibians and mammals (akirin1 and akirin2), two to three members in teleosts, and more than three members in Salmonidae (Macqueen et al., 2010a,b; Liu et al., 2015).
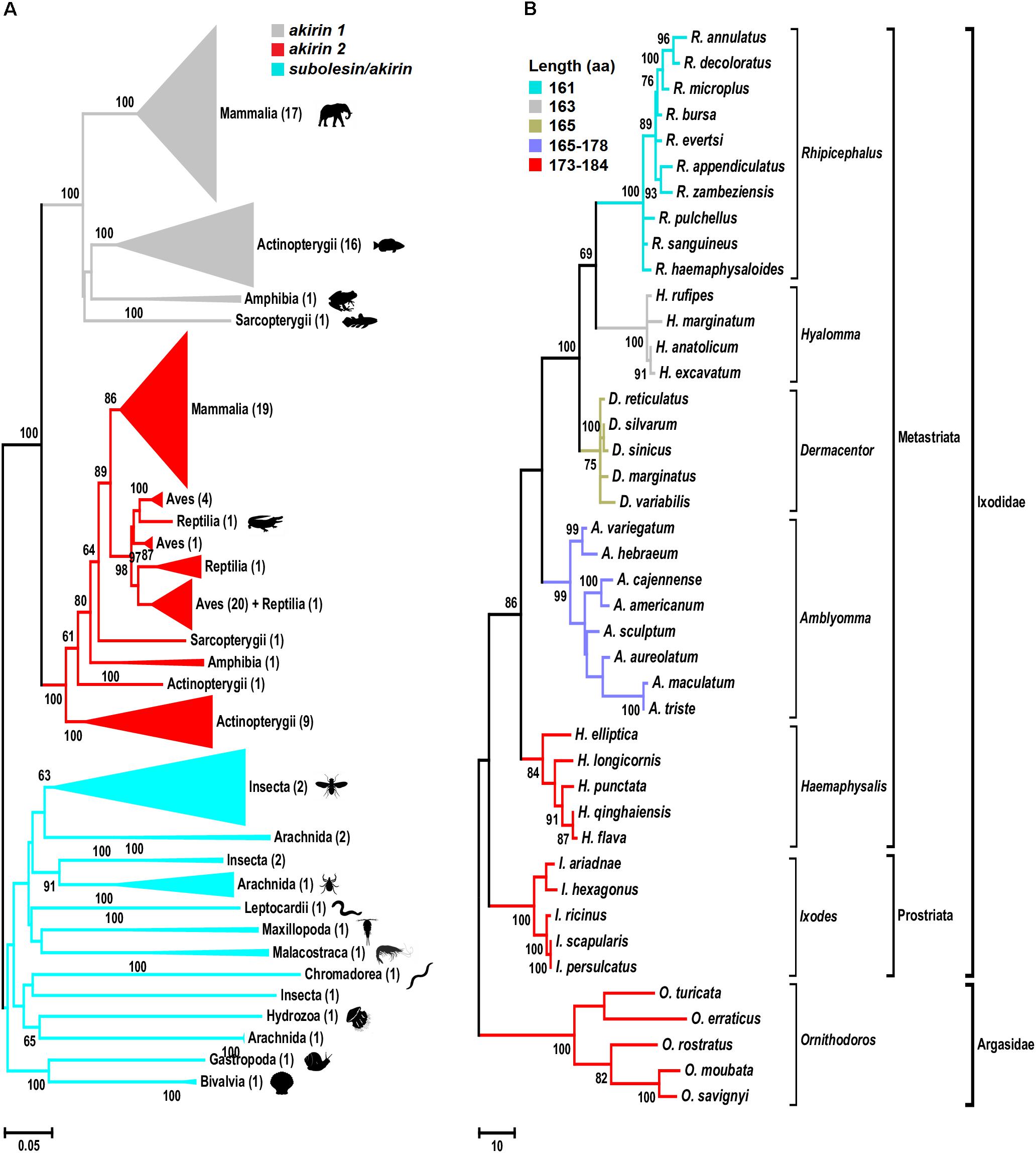
FIGURE 1. Phylogenetic analysis of akirin and subolesin nucleotide sequences. (A) A Neighbor Joining (NJ) phylogenetic tree was constructed with 361 nucleotide sequences belonging to 152 families, 73 orders and 15 classes (Mammalia, Actinopterygii, Amphibia, Sarcopterygii, Aves, Reptilia, Arachnida, Malacostraca, Insecta, Leptocardii, Maxillopoda, Chromadorea, Hydrozoa, Gastropoda and Bivalvia) of animals. All branches were collapsed at the class level and the number of orders per cluster is shown inside brackets. GenBank accession numbers and species names are provided in Supplementary Figure S1. Sequences were aligned using MAFFT configured for the maximum accuracy (Katoh and Standley, 2013). The final alignment contained 303 gap-free sites. All ambiguous positions were removed for each sequence pair. The best-fit model of the sequence evolution was selected based on Corrected Akaike Information Criterion (cAIC) and Bayesian Information Criterion (BIC) implemented in Molecular Evolutionary Genetics Analysis (MEGA) version 7. The Kimura 2-parameter model, which showed the lowest values of cAIC and BIC, was chosen for tree reconstruction. The evolutionary history was inferred using the NJ method implemented in MEGA 7 (Kumar et al., 2016). The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test (500 replicates) is shown next to the branches (Felsenstein, 1985). (B) Phylogenetic tree of tick subolesin sequences. A Maximum Parsimony (MP) phylogenetic tree was constructed with 42 nucleotide sequences belonging to 6 and 1 genera of hard (family Ixodidae) and soft (family Argasidae) ticks, respectively. Because the evolution of subolesin in ticks has been less studied when compared to akirins, MP was used to generate a robust hypothesis on the evolution of this molecule in ticks. Sequences were aligned using MAFFT configured for the maximum accuracy (Katoh and Standley, 2013). Then, using the MAFFT alignment as template, a condon aligment was build (HIV database; www.hiv.lanl.gov accessed on 29-12-2017). The final alignment contained 576 total sites of which 329 were gap-free. The evolutionary history was inferred using the MP method (implemented in Molecular Evolutionary Genetics Analysis (MEGA) version 7 (Kumar et al., 2016). The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test (500 replicates) is shown next to the branches (Felsenstein, 1985). The MP tree was obtained using the Subtree-Pruning-Regrafting (SPR) algorithm with search level 1 in which the initial trees were obtained by the random addition of sequences (10 replicates). Sequences were collected from Genbank and transcriptome projects and accession numbers are as follow: Ixodes scapularis (AY652654), I. persulcatus (KM888876), I. ricinus (JX193817), I. ariadnae (KM455971), I. hexagonus (JX193818), Rhipicephalus evertsi (JX193846), R. appendiculatus (DQ159967), R. microplus (EU301808), R. sanguineus (JX193845), R. haemaphysaloides (KP677498), R. annulatus (JX193844), R. decoloratus (JX193843), R. zambeziensis (GFPF01005851), R. bursa (GFZJ01017781), R. pulchellus (GACK01006228), Dermacentor silvarum (JX856138), D. sinicus (KM115649), D. marginatus (KU973622), D. variabilis (AY652657), D. reticulatus (JX193847), Amblyomma variegatum (JX193824), A. hebraeum (EU262598), A. cajennense (JX193823), A. americanum (JX193819), A. maculatum (JX193825), A. aureolatum (GFAC01005925), A. triste (GBBM01002796), A. sculptum (GFAA01000261), Hyalomma anatolicum (KT981976), H. rufipes (JX193849, H. marginatum (DQ159971), H. excavatum (GEFH01000904), Haemaphysalis longicornis (EU289292), Hae. elliptica (JX193850), Hae. qinghaiensis (EU326281, Hae. flava (KJ829652), Hae. punctata (DQ159972), Ornithodoros moubata (JX193852), O. savignyi (JX193851), O. turicata (GDIE01114362), O. erraticus (HM622148) and O. rostratus (GCJJ01005500).)
The phylogenetic analysis of subolesin gene sequences was performed in 42 species belonging to 6 and 1 genera of hard (family Ixodidae) and soft (family Argasidae) ticks, respectively. The analysis corroborated previous results showing a reductive evolution in protein length (de la Fuente et al., 2006a; Figure 1B). The Subolesin amino acid (aa) sequence evolved from 173 to 184 aa in Ornithodoros, Ixodes and Haemaphysalis spp. to 161 aa in Rhipicephalus spp. (Figure 1B). It is generally accepted that evolution proceeds toward greater complexity at both the organismal and genomic levels. However, numerous examples of reductive evolution of parasites and symbionts have been described to challenge this notion (Wolf and Koonin, 2013). Wolf and Koonin (2013) proposed that quantitatively, the evolution of genomes appears to be dominated by reduction and simplification, punctuated by episodes of complexification. The reductive evolution process has been particularly documented in genomes that replicate within the domain of a host genome (Andersson and Kurland, 1998; Driscoll et al., 2017), but it has also been proposed to be involved in the origin of bacteria from eukaryotes (Staley, 2017). In arthropods, reductive evolution has been implicated in the evolutionary origin of other proteins such as type IV classical cadherins (Sasaki et al., 2017).
The protein length is subjected to systematic variation that relates to the cellular context in which it functions (Wang et al., 2011). For growth rate-optimized cells, the reduction in protein length constitutes an advantage by increasing their mass-normalized kinetic efficiencies (Ehrenberg and Kurland, 1984; Kurland et al., 2007; Wang et al., 2011). Consequently, shorter proteins that retain maximum functional rates are expected to support faster cell growth rates than longer proteins with similar kinetic characteristics. Wang et al. (2011) proposed the use of the terms “domain” and “linker” to refer to protein folded domains and nondomain regions, respectively. Proteins with nondomain sequences are proteins intrinsically unstructured or natively unfolded that lack a stable tertiary structure but have a dynamic range of conformations (Orengo and Thornton, 2005). These proteins appear to be more abundant in eukaryotes when compared to prokaryotes and are usually involved in binding and molecular recognition (Brown et al., 2011). Subolesin/Akirin were characterized as a linker with three predicted disordered nondomain regions that resulted in unstructured proteins (Prudencio et al., 2010; see also below). These results agreed with the findings of Wang et al. (2011) that the evolutionary reductive constraints on protein lengths are preferentially expressed in linker sequences.
It is difficult to establish a comprehensive record of ticks developmental rates because most of the experiments in previously published papers have been done at different regimes of temperature, relative humidity and photoperiod, all factors affecting the time in which ticks complete each developmental stage. However, data compiled by Hoogstraal (1956), Morel (2003) and Horak et al. (2018) under similar conditions established that in the range of 24–28°C, ticks of the genera Hyalomma and Rhipicephalus complete their life cycle in about 33% less time than ticks of the genera Ixodes or Amblyomma. The Hyalomma and Rhipicephalus spp. are considered the two most recent genera of ticks, while Ixodes spp. and Amblyomma are among the most ancient splits of tick lineages (Mans et al., 2016; Figure 1B). Therefore, it is possible that the reductive evolution of Subolesin is associated with faster developmental rates in Rhipicephalus and Hyalomma spp. when compared to more ancient tick species even if they are sympatric. The faster developmental rate in recently evolved tick species may be associated with increasing cell growth rates that have been associated with reductive evolution (Wang et al., 2011). However, the complete association between existing data about developmental rates and evolutionary features of ticks requires further research.
At the genome level, subolesin/akirin exon-intron architecture shows a clear evolutionary pattern (Figure 2). As shown for subolesin/akirin coding sequences (Figure 1A), the vertebrate-specific paralogs form a separate clade from invertebrate genes (Figure 2). The subolesin/akirin genes evolved from 4 exons in invertebrates to 5 exons in vertebrates. The exon sizes showed a pattern of larger to shorter for vertebrate akirin1 exons I to V and mosquito akirin exons I to IV (Figure 2). However, for tick subolesin and vertebrate akirin2 genes, the size of exons II and III, respectively, were larger or similar than that of preceding exon (Figure 2). Furthermore, while the length of 11 nucleotides (nt) in the last exon was conserved across evolution, the length of the penultimate exon evolved from 157 nt in mosquitoes to 72 nt in ticks and vertebrates (Figure 2). However, the length of the exon-intron sequence increased from 3,992 nt in mosquitoes to 22,829 nt in ticks and then decreased to 8,862 nt in fish to increase back again to 26,088 nt in humans (Figure 2). These results do not correlate with genome sizes of these organisms (Gregory, 2005), and may originated from still unknown evolutionary events.
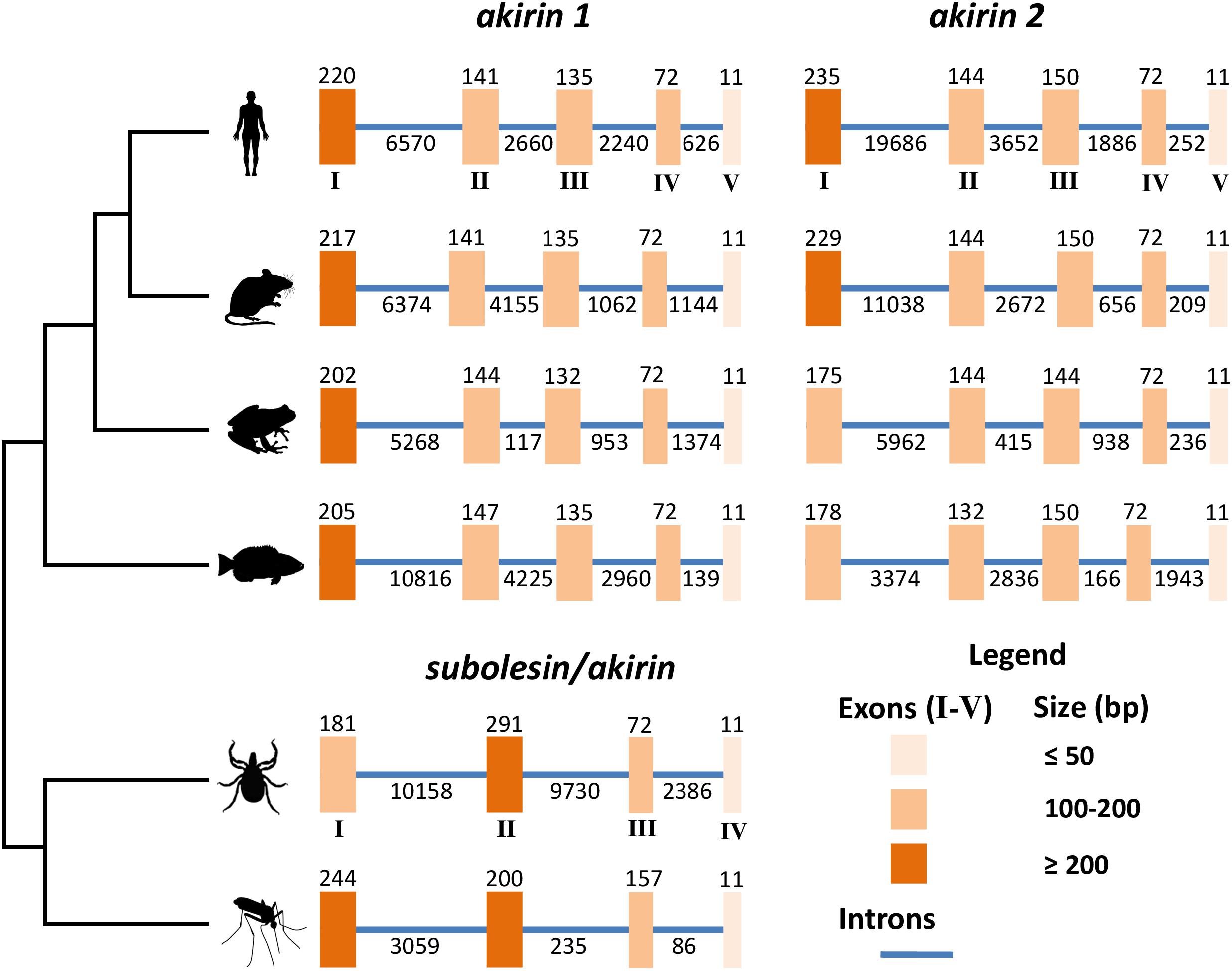
FIGURE 2. Genomic organization of subolesin/akirin orthologs across selected eukaryotic species. The genomic organization of the coding regions of tick (I. scapularis), human (Homo sapiens), mouse (M. musculus), frog (Xenopus laevis), fish (Danio rerio) and mosquito (Anopheles gambiae) subolesin/akirin is shown. The genomic organization of human, mouse, frog and fish akirins was previously reported (Liu et al., 2015). The genomic organization of tick and mosquito subolesin/akirin was collected from VectorBase (https://www.vectorbase.org; Giraldo-Calderón et al., 2015). Latin numerals correspond to the size of exons/introns in base pairs.
Subolesin/Akirin Structure and Its Interactions With Dna and Transcription Factors
Akirins are involved in cellular processes that are regulated by specific domains and binding sites. The rat Akirin2 (or 14-3-3β Interactant 1) described by Komiya et al. (2008) is 48% identical to the I. scapularis Subolesin and was previously used to identify the conserved Akirin nuclear localization signal (NLS) domains and its binding sites (Macqueen and Johnston, 2009). The pairwise sequence alignment shows that both NLS domains of the rat Akirin2 are conserved in Subolesin, but with variations in a few binding sites (Figure 3A). Subolesin binding sites 1 and 4 are similar to Akirin2 with Subolesin binding sites 2 and 3 each possessing a single substitution. The Subolesin binding site 5, however, has a double Leu insertion when compared to Akirin2 (Figure 3A). Currently there are no resolved Subolesin/Akirin structures and standard sequence-based bioinformatics methods lack parameters for locating structural homologs in the Protein Databank (PDB). Therefore, several logical steps were taken to correctly model the I. scapularis Subolesin. The Subolesin sequence (Figure 3A) was initially submitted to I-TASSER (Zhang, 2008), a protein multiple threading algorithm that is considered a top competitor in the Critical Assessment of Structure Prediction1. The I-TASSER algorithm resulted in five distinct Subolesin models that were then individually submitted to the DALI server (Holm and Laakso, 2016) for identifying PDB structural homologs of similar length with minimalα-carbon backbone deviations between the two global structures. Since Subolesin/Akirin are effectors of the IMD/TNF/TLR Relish/NF-kB signaling pathways (Goto et al., 2008; Naranjo et al., 2013), a match with homologous structures that potentially participate within these pathways was also a criterion in selecting an appropriate Subolesin model. This logical approach resulted in a Subolesin model homologous in structure, with only 7% residue conservation, to the genetically engineered catalytic domain of the transposase Sleeping Beauty (Ivics et al., 1997; Zanesi et al., 2013; Figure 3B).
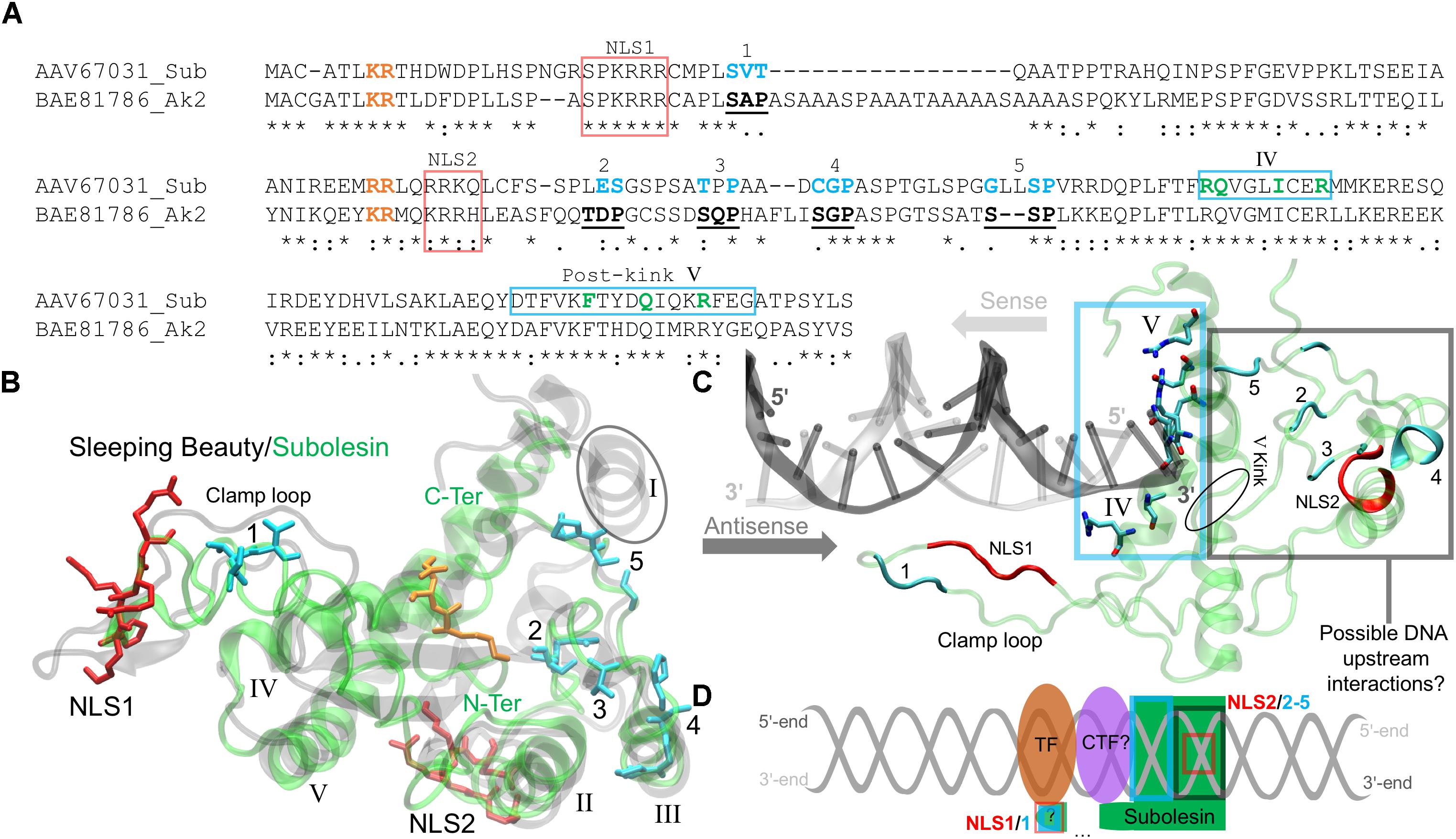
FIGURE 3. The I. scapularis Subolesin structure and its interactions with DNA and transcription factors. (A) The pairwise sequence alignment of the I. scapularis Subolesin (Sub) and the rat Arkinin2 (Ak2), accession numbers indicated, was generated using the MAFFT alignment program at default settings (Katoh et al., 2017). The NLS 1 and 2 domains (red box), binding sites 1-5 (bold-underlined in Ak2 and cyan for Sub), and the novel DNA binding sites (green and enclosed in a cyan box) are shown. The residues color-coded orange are extensions of the NLS domains. (B) The superposed tertiary structures of Sleeping Beauty (transparent black) and Subolesin (transparent green) are represented with the clamp loop labeled and the five α-helices of Sleeping Beauty (PDB: 5CR4) annotated in roman numerals. The tertiary residue positions of the labeled Subolesin NLS domains and binding sites are, respectively, color-coded as in the pairwise alignment. The Subolesin termini positions are color-labeled (green). (C) The Subolesin-DNA complex, modeled from the Mos1-DNA (PDB: 3HOS) show the residues of the novel DNA-binding site on α-helices IV-V, enclosed by a cyan box that were predicted by I-TASSER (Zhang, 2008). The DNA prime ends are color-labeled for the respective directions (indicated by arrows) of the sense (gray) and antisense (dark gray) strands. The residue positions of the Subolesin clamp loop, NLS domains and binding sites are color-coded as in previous panels A and B. (D) The schematic representation of the upstream DNA (gray helix) interactions with Subolesin NLS2, binding sites 2-5, and the potential clamp loop interaction (via NLS1 and binding site 1) with an unknown co-transcription factor (CTF?) and unknown (?) transcription factor (TF).
As part of the Tc1/mariner transposon superfamily, the Sleeping Beauty transposase has been engineered for genetic screening studies, leading to the discovery of several genes activated by Sleeping Beauty transposon insertions that participate in the NF-kB signaling pathway (Zanesi et al., 2013). The Sleeping Beauty transposase sequence is composed of an N-terminus paired-like domain with a leucine zipper (∼90 residues long) and the C-terminus folds as the catalytic domain (Ivics et al., 1997). The catalytic domain of Sleeping Beauty was resolved with a DNA transposon end and modeled with a target DNA revealing the mechanism of hyperactive Sleeping Beauty mutation screening studies while discovering novel variants for future screenings (Voigt et al., 2016). The Sleeping Beauty crystal structure details that its catalytic domain has a global homology to Ribonuclease H (RNase H) (Voigt et al., 2016). The RNase H-like protein fold forms a catalytic triad (Asp-Asp-Glu) that coordinates metal ions involved in excision and insertions of DNA (Voigt et al., 2016). By resolving the Sleeping Beauty catalytic domain, Voigt et al. (2016) also discovered that the Gly-rich box (located on the clamp loop) is involved in protein-protein interactions, specifically with partnering monomers in the DNA complex. The conserved positions of the catalytic triad and the Gly-rich box, however, are not present in Subolesin/Akirin sequences.
Prior to acting in the Tc1/mariner transposon system, Sleeping Beauty must enter the nucleus. Passage to the nucleus is controlled by NLS domains that have a strong affinity to karyopherin/importin receptors, proteins responsible for transporting NLS-tagged “cargo” in and out of the nucleus via nuclear pores (Leung et al., 2003). The N-terminus of the Sleeping Beauty catalytic domain contains a NLS domain that is quite long (17 residues) and is actually a bipartite NLS (Ivics et al., 1997). A monopartite NLS domain has the formulation Lys-Lys/Arg-[X]-Lys-Lys/Arg, where [X] is any other (∼2) amino acids. A bipartite NLS domain has a linker sequence, where [X] is ∼10 residues long (Makkerh et al., 1996). Mutations of these upstream NLS residues prior to the linker sequence [X] has shown to reduce protein entry into the nucleus (Dingwall et al., 1988), and inhibit NLS binding to karyopherin/importin receptors (Leung et al., 2003). These upstream, positively charged residue pairs are in the aligned sequences of Figure 3A (orange-labeled residues), indicating that Subolesin/Akirin2 NLS1 is bipartite. The two positively charged residues highlighted near NLS2 (Figure 3A) indicate that the NLS2 of Subolesin/Akirin is actually a longer monopartite domain. The modeled Subolesin structure has its NLS1 positioned on the clamp loop, which differs from Sleeping Beauty that is coordinated at the N-terminus α-helix (I) (Figure 3B). The Subolesin NLS2 domain, not present in the catalytic domain of Sleeping Beauty, is located on an α-helix (II) outside the central core of the protein. The absence of the catalytic triad of Sleeping Beauty (Voigt et al., 2016) and metal binding sites in Subolesin/Akirin support that these proteins do not act as a transposase. Additionally, the Subolesin/Akirin binding sites have long been recognized by mutation studies (Komiya et al., 2008), and Subolesin/Akirin RNA interference (RNAi) experiments have shown to disrupt the Relish signaling pathway (Goto et al., 2008; Naranjo et al., 2013). However, as discussed bellow, the Subolesin/Akirin interactome has not been fully characterized, and whether Subolesin/Akirin binding partners are only proteins or also include nucleic acids.
The superposed structures of Subolesin and Sleeping Beauty depict a global homology with a low α-carbon backbone deviation of 0.3 nm (Figure 3B). There are, however, missing and disordered secondary structures. The Sleeping Beauty catalytic domain has five β-sheets surrounded by five α-helices. As previously mentioned, the N-terminus α-helix I of Sleeping Beauty that contains its NLS domain is missing in the Subolesin model (encircled in Figure 3B), thereby shifting the Subolesin NLS1 domain to the clamp loop. Moreover, the β-sheets of Subolesin are highly disordered. Future experiments should resolve the stacking and conformations of the disordered Subolesin β-sheets by X-ray crystallography. Nevertheless, the remaining four α-helices (II-V) are structurally conserved, and the Subolesin/Akirin binding sites 2-5 are positioned on or approximating α-helix III (Figure 3B). As in the primary sequence (Figure 3A), the Subolesin/Akirin binding site 1 is structurally distant from the other sites, located on the N-terminus clamp loop near the position of NLS1 (Figure 3B). As a transposase, the clamp loop of the resolved Sleeping Beauty catalytic domain is not in its DNA-bound conformation. Therefore, Voigt et al. (2016) modeled the clamp loop after the DNA-bound transposase, Mos1, from Drosophila mauritiana (Richardson et al., 2009). The Mos1 also has poor sequence identity to Subolesin (<5%), but are structurally homologous with α-carbon backbone deviation of 0.34 nm. This led to a subsequent I-TASSER simulation using the template Mos1 as conducted by Voigt et al. (2016). The resulting model has Subolesin bound to a DNA duplex with an adequate clamp loop conformation that extends downstream the duplex (Figure 3C).
The Subolesin-DNA complex show several residues on α-helices IV and V that approximate the DNA 5′-end of the sense strand and the 3′-end of the antisense strand (Figure 3C). Four of the seven residues positioned on α-helix IV mainly interact with the phosphate backbone of the antisense strand. The remaining three residues that interact with both strands are after the pivotal kink of α-helix V (encircled in Figure 3C). The alignment in Figure 3A highlights these novel DNA-binding residues and indicates the α-helix on which they are positioned. The binding sites 2–5 and NLS2 are distal to the DNA interacting site, while the clamp loop containing NLS1 and binding site one is downstream the DNA duplex. Given the structural coordination of Subolesin bound to DNA (Figure 3C), binding sites 2–5 and NLS2 may interact with nucleotides upstream the DNA or with additional co-transcription factors (CTF) (Figure 3D). The transcription factors (TF) Relish/NF-kB form DNA-protein complexes with CTFs, and Subolesin is hypothesized to act as a CTF of Relish via an intermediate CTF (Figure 3D and described in the next section). Furthermore, the extended DNA downstream position of the Subolesin clamp loop with binding site one will hypothetically coordinate the CTF and possibly the TF (Figure 3D). If Subolesin is a CTF, how will it conform while the mRNA is being transcribed? Positively charged residues, specifically Lys, recognize RNA strands via electrostatic interactions (Law et al., 2006). Given conformational flexibility of the Subolesin clamp loop and the fact that it does not contain the Sleeping Beauty Gly-rich box, the proximity of positively charged Subolesin NLS1 domain residues downstream the DNA duplex may guide transcribing mRNAs for post-transcriptional processing.
Function of Subolesin/Akirin
The innate immune response acts as the first line of defense against pathogen infection in all metazoans, and constitutes the only immune response in invertebrates (Medzhitov et al., 1997; Shaw et al., 2017). It has been shown that invertebrate Subolesin/Akirin and vertebrate Akirin2 act in concert with Relish/NF-kB to induce the expression of a subset of downstream pathway elements in the IMD and TNF/TLR signaling pathways involved in the immune response to pathogen infection. This function of Subolesin/Akirin has been documented in ticks (de la Fuente et al., 2008, 2017a; Naranjo et al., 2013; Gulia-Nuss et al., 2016; Shaw et al., 2017), fruit fly (Beutler and Moresco, 2008; Goto et al., 2008; Wan and Lenardo, 2010; Bonnay et al., 2014), human (Goto et al., 2008), salmon (Macqueen et al., 2010b), shrimp (Hou et al., 2013), Japanese flounder (Yang et al., 2013), amphioxus (Yan et al., 2013), rock bream (Kasthuri et al., 2013), Chinese loach (Xue et al., 2014), Hong Kong oyster (Qu et al., 2014), turbot (Yang et al., 2011), sea louse (Valenzuela-Muñoz and Gallardo-Escárate, 2014), mouse (Beutler and Moresco, 2008; Goto et al., 2008; Wan and Lenardo, 2010; Tartey et al., 2015), croaker (Liu et al., 2015); shrimp (Liu Y. et al., 2016; Liu et al., 2016), and seahorse (Pavithiran et al., 2018; Figures 4, 5).
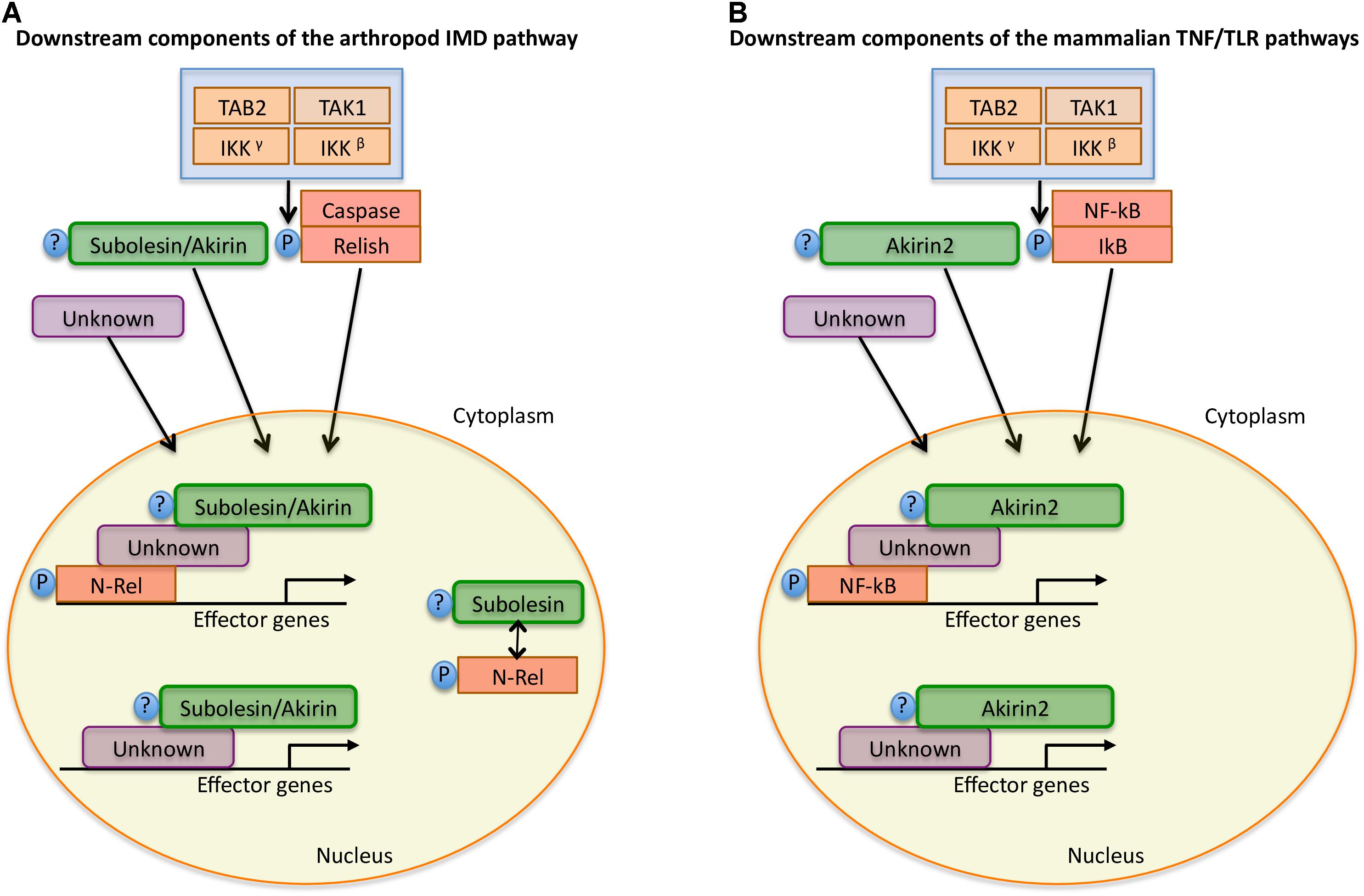
FIGURE 4. Model for Subolesin/Akirin function in immune response pathways. A simplified annotation of the downstream components of the arthropod IMD and mammalian TNF/TLR pathways (Goto et al., 2008; Beutler and Moresco, 2008; de la Fuente et al., 2008; Naranjo et al., 2013; Shaw et al., 2017). (A) After activation of the arthropod IMD pathway, the TGF-β (TAK1), Tak1-binding protein 2 (TAB2) and the I-KB kinase (IKK) complex are recruited, which leads to phosphorylation of the NF-κB transcription factor, Relish. After phosphorylation, the N-terminal domain of Relish (N-Rel) is cleaved by Caspase-8 homolog Dredd or a similar Caspase and is translocated to the nucleus. Subolesin/Akirin may be post-translationally modified and translocated to the nucleus. In the nucleus, N-Rel interacts with Subolesin/Akirin through unknown proteins to drive the production of anti-microbial peptides and other effector genes. In ticks, N-Rel and Subolesin may be reciprocally regulated. (B) In mammals, the activation of the TNF/TLR signaling pathways also results in the recruitment of the TAB2-TAK1 and IKK complexes, which results in the phosphorylation of the inhibitory regulator of NF-kB, IkB, resulting in the NF-kB translocation to the nucleus. As in arthropods, Akirin2 may be post-translationally modified and translocated to the nucleus. Once in the nucleus, NF-kB interacts with Akirin2 through unknown proteins for the activation of gene expression. In both arthropods and mammals, Subolesin/Akirin are involved in the regulation of genes that are Relish/NF-kB independent.
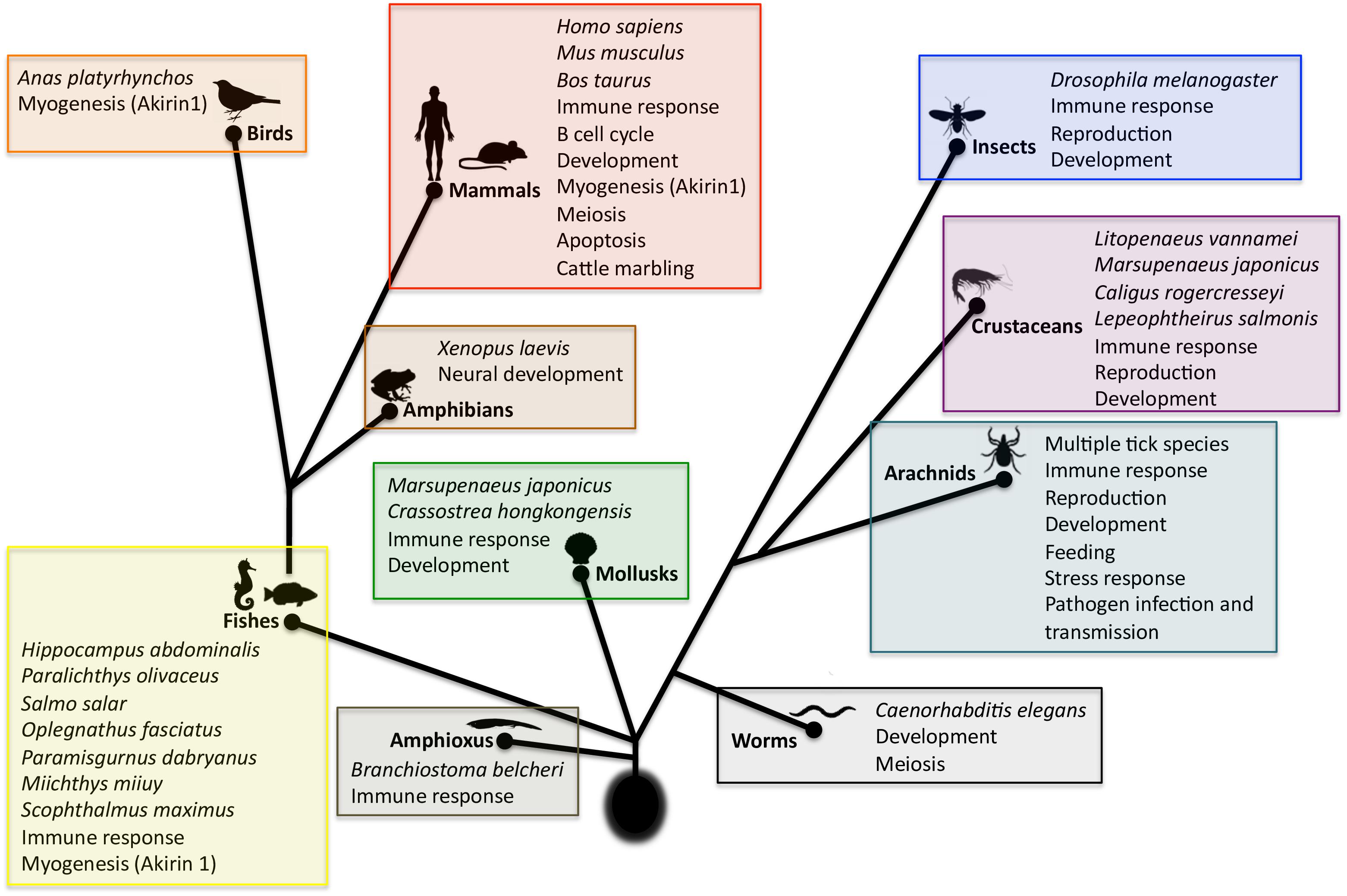
FIGURE 5. Scheme of the evolution and function of Subolesin/Akirin | Functional annotations were done based on published results for Subolesin/Akirin2. References are in the text of the paper. Myogenesis, attributed exclusively to Akirin1, was included and labeled as such. For each taxa, species in which studies were conducted are shown.
Subolesin/Akirin are also involved in the Relish/NF-kB independent gene regulation (Figure 4), thus playing a role in various biological processes in addition to the immune response (Figure 5). These processes include animal reproduction and development, causing lethal embryonic or reduced growth phenotypes in knockout mice, fruit fly, ticks, and nematodes (Maeda et al., 2001; de la Fuente et al., 2006a; Goto et al., 2008; Carpio et al., 2013; Qu et al., 2014), metazoan myogenesis (Marshall et al., 2008; Salerno et al., 2009; Macqueen et al., 2010a; Mobley et al., 2014; Sun et al., 2016), Xenopus neural development (Liu et al., 2017), meiosis/carcinogenesis (Komiya et al., 2008; Macqueen et al., 2010b; Clemons et al., 2013), tick stress response, feeding, growth and reproduction (Almazán et al., 2003, 2005; de la Fuente et al., 2006a, 2008; Smith et al., 2009; Busby et al., 2012; Rahman et al., 2018), pathogen infection and transmission in ticks (de la Fuente et al., 2006b, 2016, 2017a; Zivkovic et al., 2010a,b; Busby et al., 2012; Hajdušek et al., 2013) and turbot (Yang et al., 2011), human glioblastoma cell apoptosis (Krossa et al., 2015), cattle marbling (Sasaki et al., 2009; Watanabe et al., 2011; Kim et al., 2013), and mouse mitogen-induced B cell cycle progression and humoral immune responses (Tartey et al., 2015). For example, as previously reported in I. scapularis and other tick species (de la Fuente and Kocan, 2006; de la Fuente et al., 2006a,b, 2011, 2013; Merino et al., 2013a; de la Fuente and Contreras, 2015), Subolesin appears to function in multiple biological processes such as tick response to infection, feeding, reproduction, development and stress response (Figure 6).
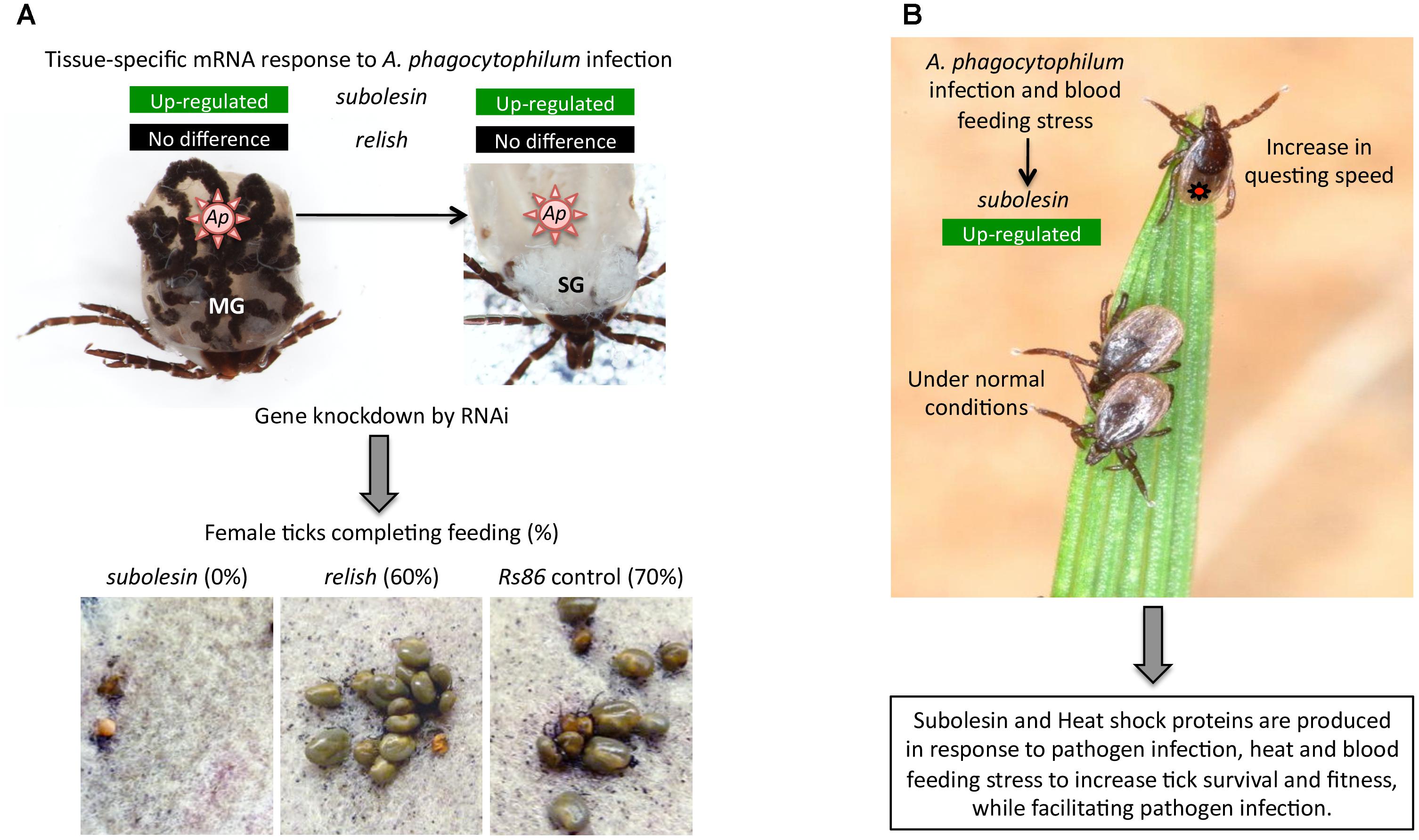
FIGURE 6. Examples of the role of tick Subolesin in different biological processes | (A) Role of tick Subolesin in A. phagocytophilum infection and blood feeding. The transcriptomics analysis in different I. scapularis tick tissues showed that subolesin (ISCW023283) but not relish (ISCW018935) mRNA levels significantly increased in response to A. phagocytophilum (Ap) infection in both midgut (MG) and salivary glands (SG). In addition, the subolesin gene knockdown phenotype in ticks injected with dsRNA resulted in a significant reduction in the number of female ticks completing feeding, oviposition and fertility. Results were reported by Ayllón et al. (2015). Photo of dissected I. scapularis partially fed adult female ticks courtesy of K. M. Kocan (Oklahoma State University, United States). (B) Role of tick Subolesin in A. phagocytophilum infection, blood feeding and questing speed. The response to A. phagocytophilum and stress increases subolesin levels, which together with heat shock proteins improve tick questing speed and survival. Results were reported by Busby et al. (2012). Photo of questing Ixodes ricinus courtesy of L. Grubhoffer & J. Erhard (Biology Center of the AS CR, Institute of Parasitology, Czechia).
Akirin1 and Akirin2 have also different functions in vertebrates, which is illustrated by the role of Akirin1 in myogenesis while Akirin2 promotes meiosis/carcinogenesis (Macqueen and Johnston, 2009; Macqueen et al., 2010a,b; Figure 4). These different functions may be related to the Akirin subcellular localization. While Akirin1 is found in the nucleus, Subolesin/Akirin2 is located in both cytoplasm and nucleus (de la Fuente et al., 2011; Antunes et al., 2014; Krossa et al., 2015; Pavithiran et al., 2018; Figure 7). The subcellular localization of Subolesin/Akirin2 is probably related to its structure, which as discussed above contains NLS domains that are involved in protein transport in and out of the nucleus via nuclear pores (Leung et al., 2003).
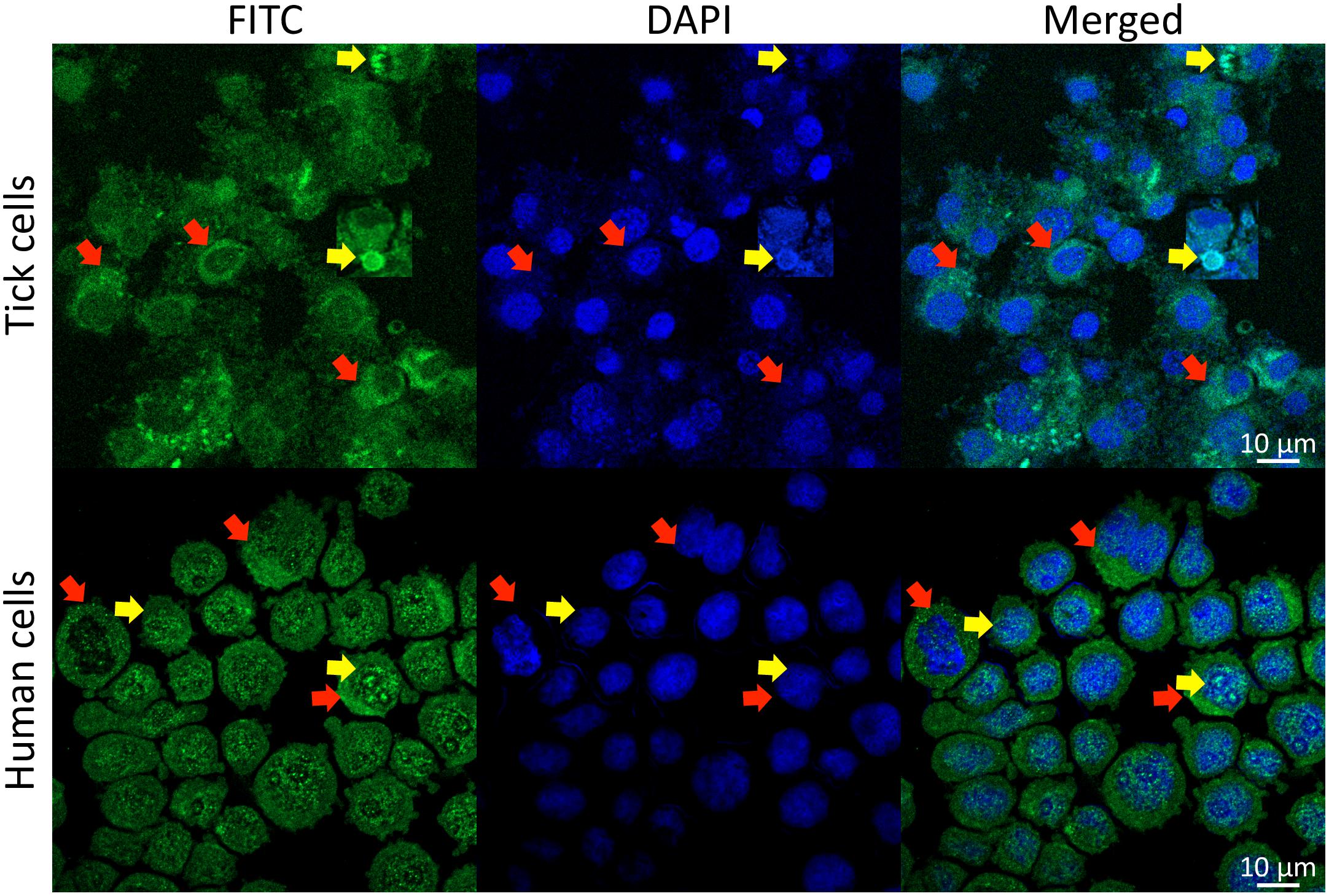
FIGURE 7. Subcellular localization of Subolesin/Akirin2. Representative images of immunofluorescence analysis of I. scapularis ISE6 and human HL60 cells incubated with anti-Subolesin and anti-Akirin2 antibodies, respectively. The cells were fixed with 4% paraformaldehyde in PBS for 15 min, permeabilized with 0.5% Triton X-100 in PBS for 5 min and blocked with blocking buffer (3% BSA in PBS) for 1 h. Then, the cells were incubated overnight at 4°C with anti-Subolesin (Antunes et al., 2014) or anti-Akirin2 (Abcam, Cambridge, United Kingdom) antibodies (1/50 dilution in 3% BSA in PBS). After 3 washes with PBS, the slides were incubated with fluorescein isothiocyanate (FITC) conjugated anti-rabbit secondary antibodies (Sigma-Aldrich, St. Louis, MO, United States; green) at 1/160 dilution in 3% BSA in PBS for 1 h at room temperature. Cells were counterstained with ProLong Antifade containing 4′,6-diamidino-2-phenylindole (DAPI) (Molecular Probes, Eugene, OR, United States; blue), and imaged with a Zeiss LSM800 confocal microscope using a 63× oil immersion lens (Carl Zeiss, Oberkochen, Germany). Yellow arrows show examples of protein localization in the nucleus while red arrows illustrate protein localization in the cytoplasm.
In summary and based on current information, Subolesin/Akirin evolved with similar functions in both invertebrates and vertebrates (Figure 5). The annotation of some biological processes described in certain taxa only may be due to the presence of species-specific functions or more likely a consequence of the still incomplete characterization of Subolesin/Akirin function in the different species.
Subolesin/Akirin Role in Cell Interactome and Regulome
Subolesin/Akirin are proteins without catalytic or DNA-binding capacity. How Subolesin/Akirin regulate gene expression is unknown but likely involve interactions with proteins with DNA-binding or chromatin-remodeling capacity (de la Fuente et al., 2008; Beutler and Moresco, 2008; Nowak and Baylies, 2012; Nowak et al., 2012; Naranjo et al., 2013). In this way, Subolesin/Akirin link the activities of transcription factors with those of chromatin remodeling complexes to influence gene expression in a context-dependent manner (Nowak and Baylies, 2012; Nowak et al., 2012). For example, Subolesin/Akirin and Akirin2 as key components of the innate immune response can directly or indirectly interact with other regulatory proteins such as “14-3-3” proteins, DNA methyltransferase-associated protein 1 (DMAP1) and the basic helix–loop–helix transcription factor (Twist) to up- or down-regulate transcription (Gonzalez and Baylies, 2005; Goto et al., 2008, 2014; Komiya et al., 2008; de la Fuente et al., 2008; Beutler and Moresco, 2008; Nowak et al., 2012; Naranjo et al., 2013; Tartey et al., 2014; Gulia-Nuss et al., 2016; Shaw et al., 2017).
The Subolesin/Akirin role in the cell interactome and regulome in response to different stimuli has not been characterized. Recently, we proposed a method based on the graph theory for the analysis of human and tick cell proteome in response to A. phagocytophilum infection (Estrada-Peña et al., 2018). This approach resulted in a network of interacting proteins and cell processes clustered in biological pathways, and ranked with indexes representing the topology of the proteome influenced by features of the interactome and regulome. The results evidenced differences in the response to A. phagocytophilum infection between human and tick cells, and supported that human neutrophils but not tick cells limit pathogen infection through differential representation of ras-related proteins (Estrada-Peña et al., 2018). Herein, this method was applied to predict the position of Subolesin in the regulome of tick cells and in response to A. phagocytophilum infection (Figure 8). The results showed that Subolesin is deeply involved in the core of transcription processes but also in other secondary processes such as transcription from RNA polymerase II promoter, DNA repair, and chromatin remodeling (Figure 8). Furthermore, other processes that change in infected cells when compared to uninfected cells (i.e., signal transduction, regulation of transcription, and response to heat) are deeply linked to the central transcription process. The putative Subolesin role in these processes varied between infected and uninfected cells (the width of the lines is proportional to the importance of the link between proteins and processes; Figure 8). For example, it appears that particularly in infected cells other proteins but Subolesin have a more prominent role in the strong protein link with transcription and transcription from RNA polymerase II promoter processes (Figure 8). These results predict the role that Subolesin plays in the regulation of different biological processes, and its differential role in response to A. phagocytophilum infection in tick cells. However, the lack of a more prominent role for Subolesin may respond to the fact that this protein does not bind directly to DNA but interacts with other proteins to exert its regulatory function.
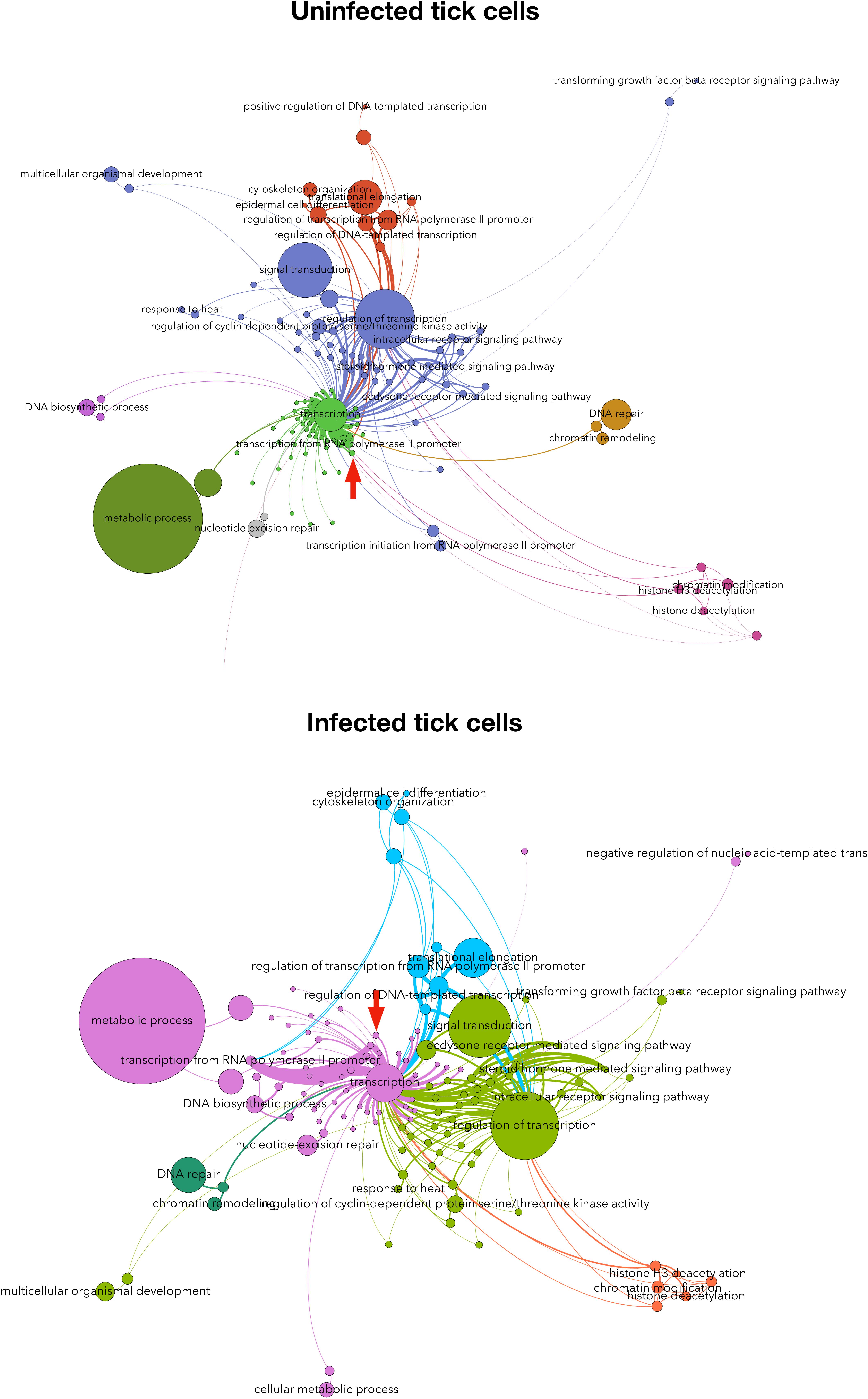
FIGURE 8. Subolesin regulome in tick cells. The network of proteins and processes associated to transcription in tick cells uninfected and infected with Anaplasma phagocytophilum. The nodes (circles) are either proteins or processes (labeled). The size of each circle is proportional to its centrality index. The networks show (clusters of interacting proteins and processes in colors. The width of each link is proportional to the strength of the interaction. The networks show the topology of the tick interactome and regulome. The networks were built with the annotated proteins represented in either uninfected or infected cells, and a directed network was built for each protein linked to the processes in which it is involved. The weight of each link is proportional to the number of reads of the protein. This weighted degree of each link was used to calculate the centrality indexes, mainly the Betweenness Centrality, which is represented in the panels. Only the proteins annotated as involved in processes associated with transcription (i.e., linked by one or more protein(s) simultaneously annotated as transcripiton or other cellular process). The topology of the networks was obtained with the Lovaine algorithm. In both networks, the topological position of Subolesin is marked with a red arrow. Methods were described in Estrada-Peña et al. (2018).)
In an attempt to provide information on the Subolesin/Akirin interactome, the information on Subolesin/Akirin-protein physical and functional interactions was compiled from the String protein-protein interactions database2 (Figure 9 and Supplementary Dataset S1). Based on the analysis of protein-protein interactions, the results did not allow establishing an evolutionary signature of the Subolesin/Akirin2 interactome (Figure 9), probably due to the limited information available. Nevertheless, similar Subolesin/Akirin2 interacting proteins were described in fly and mouse (B7PRT9, Brahma/SWI2-related protein BRG-1) and in fish and rat (B7P8Y4, Arginyl-tRNA synthetase) suggesting possible evolutionarily conserved protein-protein interactions (Figure 9). To further gain insight into the evolution of the Subolesin/Akirin2 interactome, instead of looking only at protein-protein interactions, the interacting proteins were annotated according to the biological processes in which they are involved (Figure 9). The results of this analysis showed that the biological processes affected by the Subolesin/Akirin interactome are evolutionarily conserved, with metabolic process (MP), cellular process (CP) and biological regulation (BR) being among the most represented processes in all organisms (Figure 9).
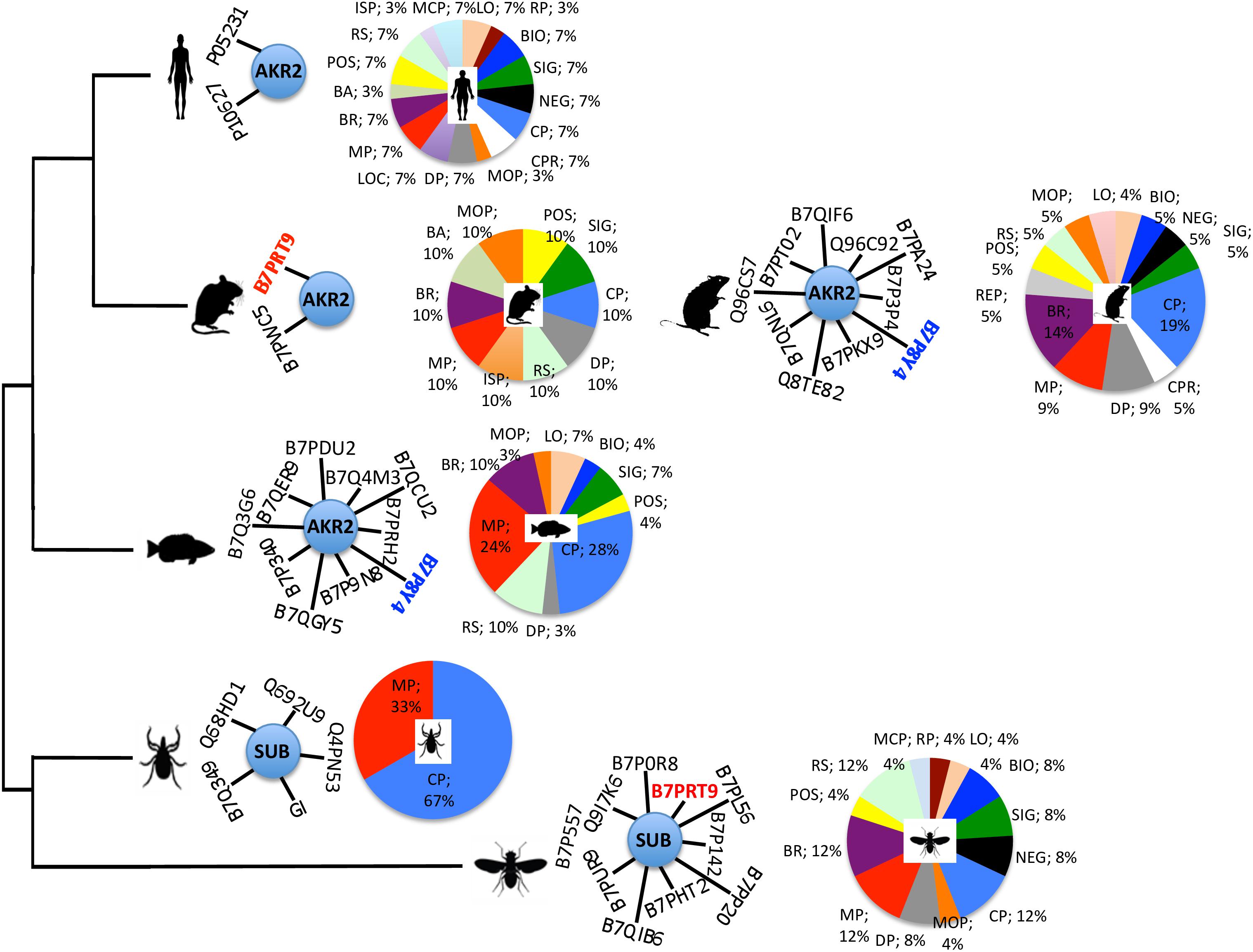
FIGURE 9. Characterization of the Subolesin/Akirin2 interactome. The information on Subolesin/Akirin-protein physical and functional interactions was compiled from the String protein-protein interactions database v.10.5 (https://string-db.org). The central node of the networks represent Subolesin/Akirin2 while the edges correspond to the predicted functional associations. Only predictions with medium (or better) confidence ( > 0.4) limited to the top 10 interactions with protein-protein interaction (PPI) enrichment p-value ≤ 0.5 were considered. To compare the different species, protein annotations were standardize by identity to I. scapularis/I. ricinus-D. melanogaster-H. sapiens order of priority (see Supplementary Dataset S1 for complete annotations). For illustration purposes, the species included in the analysis correspond to D. melanogaster, I. scapularis, Danio rerio, Mus musculus, Rattus norvegicus, and H. sapiens. Identical proteins in two different species are highlighted in red and blue letters. The functional annotation of the Subolesin/Akirin2 interacting proteins according to the biological processes (level 2) in which they are involved was done using Blast2GO (www.blast2go.com), and represented in pies with different colors for each process and the percentage of proteins on each process. Abbreviations: LO, localization (sepia); RP, rhythmic process (sangria); BIO, biogenesis (blue); SIG, signaling (green); NEG, negative regulation of biological process (black); CP, cellular process (azure); CPR, cell proliferation (white); MCP, multi-organism process (sky); DP, developmental process (gray); LOC, locomotion (violet); MP, metabolic process (red); BR, biological regulation (byzantine); BA, biological adhesion (moss); POS, positive regulation of biological process (yellow); RS, response to stimulus (tea); ISP, immune system process (gold); MOP, multicellular organismal process (orange); REP, reproductive process (smoke). Color code was established according to color thesaurus (https://graf1x.com/list-of-colors-with-color-names/).
Protective Capacity of Subolesin/Akirin for the Control of Ectoparasite Vector Infestations and Pathogen Infection
Subolesin was discovered and characterized as a tick protective antigen for the control of I. scapularis infestations (Almazán et al., 2003, 2005; Sonenshine et al., 2006). Since then, Subolesin/Akirin showed a protective capacity in vaccines for the control of infestations by different arthropod species and pathogen infection and transmission (reviewed by de la Fuente et al., 2006a, 2011, 2013; de la Fuente and Kocan, 2006, 2014; Merino et al., 2013a,b; de la Fuente and Contreras, 2015). The putative mechanism for Subolesin vaccine protection was described by de la Fuente et al. (2011). They showed that by still unknown mechanisms anti-Subolesin antibodies could enter into tick cells where they can interact with cytosolic Subolesin to prevent its translocation to the nucleus and therefore the possibility to exert it regulatory functions.
The development of vaccines for the control of multiple arthropod ectoparasites constitutes a priority for targeting the various species infesting the same host (de la Fuente et al., 2011; Contreras and de la Fuente, 2016b). In this context, Subolesin/Akirin appears as a promising vaccine protective antigen due to its conservation in sequence and function through evolution (Figures 1, 5, 9). In fact, recent results support the potential of Subolesin/Akirin as a vaccine protective antigen for the control of multiple ectoparasite vector species and transmitted pathogens (Figure 10). The arthropod ectoparasite species in which Subolesin/Akirin have shown protective capacity by affecting different phases of their life cycles include the genera Aedes, Anopheles, Phlebotomus, Caligus, Dermanyssus, Ornithodoros, Ixodes, Haemaphysalis, Amblyomma, Dermacentor, Hyalomma and Rhipicephalus (Almazán et al., 2003, 2005, 2010, 2012; de la Fuente et al., 2006a, 2010, 2011, 2013; Canales et al., 2009; Harrington et al., 2009; Zivkovic et al., 2010a; Moreno-Cid et al., 2011, 2013; Carpio et al., 2011; Bensaci et al., 2012; Merino et al., 2011a,b, 2013b; Carreón et al., 2012; Manzano-Román et al., 2012, 2015; de la Fuente and Kocan, 2014; da Costa et al., 2014; Shakya et al., 2014; Torina et al., 2014; Contreras et al., 2015; de la Fuente and Contreras, 2015; Lu et al., 2016; Olds et al., 2016; Contreras and de la Fuente, 2016a; Kumar et al., 2017; Villar et al., 2017; Figure 10). The reduction in pathogen infection has been shown for tick-borne pathogens, Anaplasma marginale, A. phagocytophilum, Borrelia burgdorferi s.s. and Babesia bovis, and the mosquito-borne pathogen, Plasmodium berghei (de la Fuente et al., 2011; Merino et al., 2011b, 2013a; Bensaci et al., 2012; de la Fuente and Kocan, 2014; da Costa et al., 2014; Figure 10). Furthermore, protective epitopes were mapped in Subolesin/Akirin (Prudencio et al., 2010) and chimeric antigens were designed showing protective capacity in vaccinated hosts against tick infestations (Merino et al., 2013b; Contreras and de la Fuente, 2016a).
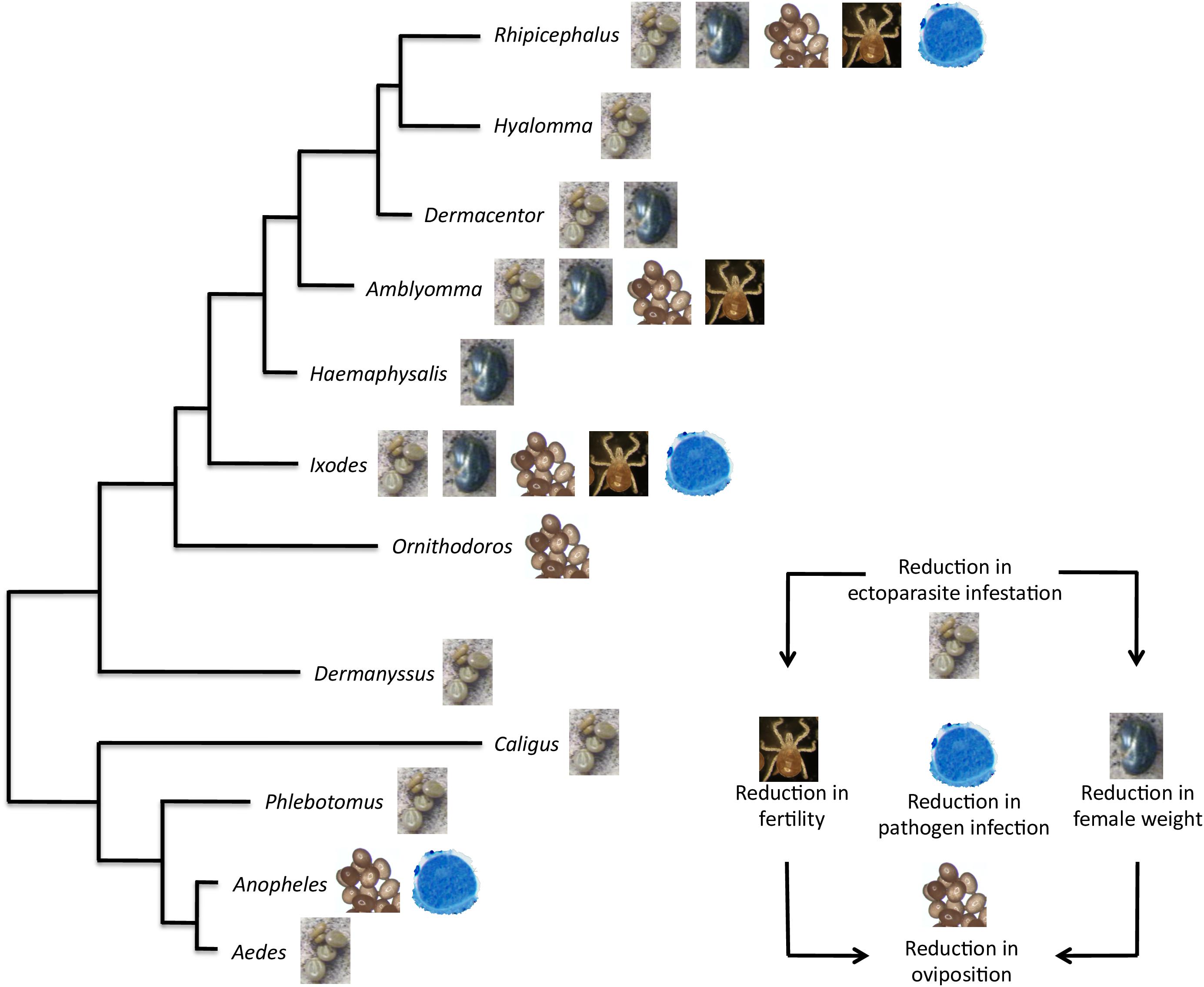
FIGURE 10. Protective capacity of Subolesin/Akirin in vaccines for the control of arthropod ectoparasite infestations and pathogen infection. The effect of Subolesin/Akirin vaccination is shown on evolutionarily diverse arthropod genera. The effect of the vaccine was recorded on different phases of ectoparasite life cycle in the form of reduction in ectoparasite infestation (number of ectoparasites completing feeding), weight (weight of engorged female ectoparasites), oviposition (number of eggs per female), and fertility (number of larvae per female) in ectoparasites fed on vaccinated hosts when compared to controls. The reduction in pathogen infection was recorded as differences in pathogen levels between ectoparasites fed on vaccinated and control hosts.
Considering the protective capacity shown by Subolesin/Akirin on different vector and pathogen species, future research directions will include the characterization of its protective capacity in other arthropod ectoparasite species, and the combination with other vector-derived and pathogen-derived antigens to increase vaccine efficacy for the control of both vector infestations and pathogen infection (Schetters et al., 2016; de la Fuente et al., 2017b; de la Fuente, 2018).
Conclusion and Future Directions
Significant advances have been made recently toward understanding the evolution and function of Subolesin/Akirin. Our results suggest that Subolesin/Akirin evolved conserving not only its sequence and structure, but also its function and role in cell interactome and regulome in response to pathogen infection and other biological processes. However, major challenges remain in fully characterizing the interactome and function of these proteins, their role in the cell regulome in response to different stimuli, and how their evolution can meet species-specific demands. Furthermore, the structure of Subolesin/Akirin and interacting molecules should be resolved by X-ray crystallography to better understand their function. Finally, the conserved functional evolution of Subolesin/Akirin correlates with the protective capacity shown by these proteins in vaccine formulations for the control of different arthropod and pathogen species, and encourage further research to develop new vaccine formulations by combining Subolesin/Akirin with interacting proteins for the control of multiple ectoparasite infestations and pathogen infection.
Author Contributions
JdlF conceived the paper. SA-J, MV, AC-C, JV, and AE-P performed the data analyses. PA and SA-J performed the microscopy studies. JF, SA-J, JV, and AC-C wrote the manuscript. All authors approved and contributed to the final version of the manuscript.
Funding
This work was financially supported by the Ministerio de Economía, Industria y Competitividad (Spain) grant BFU2016-79892-P. MV was funded by the Universidad de Castilla La Mancha, Spain. JV was supported by Project FIT (Pharmacology, Immunotherapy, nanoToxicology), funded by the European Regional Development Fund. The publication fee was partially supported by the CSIC Open Access Publication Support Initiative through its Unit of Information Resources for Research (URICI).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We acknowledge a grant for the development of a research organization RVO: RO0516 from the Veterinary Research Institute, Brno, Czechia.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fphys.2018.01612/full#supplementary-material
FIGURE S1 | Phylogenetic tree of akirin and subolesin nucleotide sequences. The figure displays a Neighbor Joining (NJ) phylogenetic tree of 361 nucleotide sequences belonging to 152 families, 73 orders and 15 classes. GenBank accession numbers and species names are shown. Tree reconstruction method was as described in Figure 1.
DATASET S1 | Annotation of the Subolesin/Akirin interactome. The information on Subolesin/Akirin-protein interactions was compiled from the literature and String protein-protein interactions database v.10.5 (https://string-db.org). Only predictions with medium (or better) confidence (0.4) limited to the top 10 interactions with protein-protein interaction (PPI) enrichment p-value ≤ 0.5 were considered. To compare the different species, protein annotations were standardize by identity to I. scapularis/I. ricinus-D. melanogaster-H. sapiens order of priority.
Footnotes
References
Adipietro, K. A., Mainland, J. D., and Matsunami, H. (2012). Functional evolution of mammalian odorant receptors. PLoS Genet. 8:e1002821. doi: 10.1371/journal.pgen.1002821
Almazán, C., Blas-Machado, U., Kocan, K. M., Yoshioka, J. H., Blouin, E. F., Mangold, A. J., et al. (2005). Characterization of three Ixodes scapularis cDNAs protective against tick infestations. Vaccine 23, 4403–4416. doi: 10.1016/j.vaccine.2005.04.012
Almazán, C., Kocan, K. M., Bergman, D. K., Garcia-Garcia, J. C., Blouin, E. F., and de la Fuente, J. (2003). Identification of protective antigens for the control of Ixodes scapularis infestations using cDNA expression library immunization. Vaccine 21, 1492–1501. doi: 10.1016/S0264-410X(02)00683-7
Almazán, C., Lagunes, R., Villar, M., Canales, M., Rosario-Cruz, R., Jongejan, F., et al. (2010). Identification and characterization of Rhipicephalus (Boophilus) microplus candidate protective antigens for the control of cattle tick infestations. Parasitol. Res. 106, 471–479. doi: 10.1007/s00436-009-1689-1
Almazán, C., Moreno-Cantú, O., Moreno-Cid, J. A., Galindo, R. C., Canales, M., Villar, M., et al. (2012). Control of tick infestations in cattle vaccinated with bacterial membranes containing surface-exposed tick protective antigens. Vaccine 30, 265–272. doi: 10.1016/j.vaccine.2011.10.102
Andersson, S. G., and Kurland, C. G. (1998). Reductive evolution of resident genomes. Trends Microbiol. 6, 263–268. doi: 10.1016/S0966-842X(98)01312-2
Antunes, S., Merino, O., Mosqueda, J., Moreno-Cid, J. A., Bell-Sakyi, L., Fragkoudis, R., et al. (2014). Tick capillary feeding for the study of proteins involved in tick-pathogen interactions as potential antigens for the control of tick infestation and pathogen infection. Parasit. Vectors 7:42. doi: 10.1186/1756-3305-7-42
Ayllón, N., Villar, M., Galindo, R. C., Kocan, K. M., Šíma, R., López, J. A., et al. (2015). Systems biology of tissue-specific response to Anaplasma phagocytophilum reveals differentiated apoptosis in the tick vector Ixodes scapularis. PLoS Genet. 11:e1005120. doi: 10.1371/journal.pgen.1005120
Bensaci, M., Bhattacharya, D., Clark, R., and Hu, L. T. (2012). Oral vaccination with Vaccinia virus expressing the tick antigen subolesin inhibits tick feeding and transmission of Borrelia burgdorferi. Vaccine 30, 6040–6046. doi: 10.1016/j.vaccine.2012.07.053
Beutler, B., and Moresco, E. M. (2008). Akirins versus infection. Nat. Immunol. 9, 7–9. doi: 10.1038/ni0108-7
Bonnay, F., Nguyen, X. H., Cohen-Berros, E., Troxler, L., Batsche, E., Camonis, J., et al. (2014). Akirin specifies NF-κB selectivity of Drosophila innate immune response via chromatin remodeling. EMBO J. 33, 2349–2362. doi: 10.15252/embj.201488456
Brown, C. J., Johnson, A. K., Dunker, A. K., and Daughdrill, G. W. (2011). Evolution and disorder. Curr. Opin. Struct. Biol. 21, 441–446. doi: 10.1016/j.sbi.2011.02.005
Busby, A. T., Ayllón, N., Kocan, K. M., Blouin, E. F., de la Fuente, G., Galindo, R. C., et al. (2012). Expression of heat-shock proteins and subolesin affects stress responses, Anaplasma phagocytophilum infection and questing behavior in the tick, Ixodes scapularis. Med. Vet. Entomol. 26, 92–102. doi: 10.1111/j.1365-2915.2011.00973.x
Canales, M., Naranjo, V., Almazán, C., Molina, R., Tsuruta, S. A., Szabó, M. P. J., et al. (2009). Conservation and immunogenicity of the mosquito ortholog of the tick protective antigen, subolesin. Parasitol. Res. 105, 97–111. doi: 10.1007/s00436-009-1368-2
Carpio, Y., Basabe, L., Acosta, J., Rodríguez, A., Mendoza, A., Lisperger, A., et al. (2011). Novel gene isolated from Caligus rogercresseyi: a promising target for vaccine development against sea lice. Vaccine 29, 2810–2820. doi: 10.1016/j.vaccine.2011.01.109
Carpio, Y., García, C., Pons, T., Haussmann, D., Rodríguez-Ramos, T., Basabe, L., et al. (2013). Akirins in sea lice: first steps toward a deeper understanding. Exp. Parasitol. 135, 188–199. doi: 10.1016/j.exppara.2013.06.018
Carreón, D., Pérez de la Lastra, J. M., Almazán, C., Canales, M., Ruiz-Fons, F., Boadella, M., et al. (2012). Vaccination with BM86, subolesin and akirin protective antigens for the control of tick infestations in white tailed deer and red deer. Vaccine 30, 273–279. doi: 10.1016/j.vaccine.2011.10.099
Clemons, A. M., Brockway, H. M., Yin, Y., Kasinathan, B., Butterfield, Y. S., Jones, S. J., et al. (2013). Akirin is required for diakinesis bivalent structure and synaptonemal complex disassembly at meiotic prophase I. Mol. Biol. Cell. 24, 1053–1067. doi: 10.1091/mbc.E12-11-0841
Contreras, M., and de la Fuente, J. (2016a). Control of Ixodes ricinus and Dermacentor reticulatus tick infestations in rabbits vaccinated with the Q38 Subolesin/Akirin chimera. Vaccine 34, 3010–3013. doi: 10.1016/j.vaccine.2016.04.092
Contreras, M., and de la Fuente, J. (2016b). Vaccinomics approach to the development of vaccines for the control of multiple ectoparasite infestations. Nova Acta Leopold. 411, 185–200. doi: 10.1186/1471-2164-12-105
Contreras, M., Moreno-Cid, J. A., Domingos, A., Canales, M., Díez-Delgado, I., Pérez de la Lastra, J. M., et al. (2015). Bacterial membranes enhance the immunogenicity and protective capacity of the surface exposed tick Subolesin-Anaplasma marginale MSP1a chimeric antigen. Ticks Tick Borne Dis. 6, 820–828. doi: 10.1016/j.ttbdis.2015.07.010
da Costa, M., Pinheiro-Silva, R., Antunes, S., Moreno-Cid, J. A., Villar, M., de la Fuente, J., et al. (2014). Mosquito Akirin as a potential antigen for malaria control. Malar. J. 13:470. doi: 10.1186/1475-2875-13-470
DasGupta, R., Kaykas, A., Moon, R. T., and Perrimon, N. (2005). Functional genomic analysis of the Wnt-wingless signaling pathway. Science 308, 826–833. doi: 10.1126/science.1109374
de la Fuente, J. (2018). Controlling ticks and tick-borne diseases…looking forward. Ticks Tick Borne Dis. 9, 1354–1357. doi: 10.1016/j.ttbdis.2018.04.001
de la Fuente, J., Almazaìn, C., Blas-Machado, U., Naranjo, V., Mangold, A. J., Blouin, E. F., et al. (2006a). The tick protective antigen, 4D8, is a conserved protein involved in modulation of tick blood ingestion and reproduction. Vaccine 24, 4082–4095.
de la Fuente, J., Almazán, C., Blouin, E. F., Naranjo, V., and Kocan, K. M. (2006b). Reduction of tick infections with Anaplasma marginale and A. phagocytophilum by targeting the tick protective antigen subolesin. Parasitol. Res. 100, 85–91.
de la Fuente, J., Antunes, S., Bonnet, S., Cabezas-Cruz, A., Domingos, A., Estrada-Peña, A., et al. (2017a). Tick-pathogen interactions and vector competence: identification of molecular drivers for tick-borne diseases. Front. Cell. Infect. Microbiol. 7:114. doi: 10.3389/fcimb.2017.00114
de la Fuente, J., Contreras, M., Estrada-Peña, A., and Cabezas-Cruz, A. (2017b). Targeting a global health problem: vaccine design and challenges for the control of tick-borne diseases. Vaccine 35, 5089–5094. doi: 10.1016/j.vaccine.2017.07.097
de la Fuente, J., and Contreras, M. (2015). Tick vaccines: current status and future directions. Expert Rev. Vaccines 14, 1367–1376. doi: 10.1586/14760584.2015.1076339
de la Fuente, J., and Kocan, K. M. (2006). Strategies for development of vaccines for control of ixodid tick species. Parasite Immunol. 28, 275–283. doi: 10.1111/j.1365-3024.2006.00828.x
de la Fuente, J., and Kocan, K. M. (2014). “Development of vaccines for control of tick infestations and interruption of pathogen transmission,” in Biology of Ticks, 2nd Edn, eds D. E. Sonenshine and R. M. Roe (New York, NY: Oxford University Press), 333–352.
de la Fuente, J., Manzano-Roman, R., Naranjo, V., Kocan, K. M., Zivkovic, Z., Blouin, E. F., et al. (2010). Identification of protective antigens by RNA interference for control of the lone star tick, Amblyomma americanum. Vaccine 28, 1786–1795. doi: 10.1016/j.vaccine.2009.12.007
de la Fuente, J., Maritz-Olivier, C., Naranjo, V., Ayoubi, P., Nijhof, A. M., Almazán, C., et al. (2008). Evidence of the role of tick subolesin in gene expression. BMC Genomics 9:372. doi: 10.1186/1471-2164-9-372
de la Fuente, J., Moreno-Cid, J. A., Canales, M., Villar, M., Pérez de la Lastra, J. M., Kocan, K. M., et al. (2011). Targeting arthropod subolesin/akirin for the development of a universal vaccine for control of vector infestations and pathogen transmission. Vet. Parasitol. 181, 17–22. doi: 10.1016/j.vetpar.2011.04.018
de la Fuente, J., Moreno-Cid, J. A., Galindo, R. C., Almazán, C., Kocan, K. M., Merino, O., et al. (2013). Subolesin/Akirin vaccines for the control of arthropod vectors and vector-borne pathogens. Transbound. Emerg. Dis. 60(Suppl. 2), 172–178. doi: 10.1111/tbed.12146
de la Fuente, J., Villar, M., Cabezas-Cruz, A., Estrada-Peña, A., Ayllón, N., and Alberdi, P. (2016). Tick-host-pathogen interactions: conflict and cooperation. PLoS Pathog. 12:e1005488. doi: 10.1371/journal.ppat.1005488
Dingwall, C., Robbins, J., Dilworth, S. M., Roberts, B., and Richardson, W. D. (1988). The nucleoplasmin nuclear location sequence is larger and more complex than that of SV-40 large T antigen. J. Cell Biol. 107, 841–849. doi: 10.1083/jcb.107.3.841
Driscoll, T. P., Verhoeve, V. I., Guillotte, M. L., Lehman, S. S., Rennoll, S. A., Beier-Sexton, M., et al. (2017). Wholly Rickettsia! Reconstructed metabolic profile of the quintessential bacterial parasite of eukaryotic cells. mBio 8:e859-17. doi: 10.1128/mBio.00859-17
Ehrenberg, M., and Kurland, C. G. (1984). Costs of accuracy determined by a maximal growth rate constraint. Q. Rev. Biophys. 17, 45–82. doi: 10.1017/S0033583500005254
Estrada-Peña, A., Villar, M., Artigas-Jerónimo, S., López, V., Alberdi, P., Cabezas-Cruz, A., et al. (2018). Use of graph theory to characterize human and arthropod vector cell protein response to infection. Front. Cell. Infect. Microbiol. 8:265. doi: 10.3389/fcimb.2018.00265
Felsenstein, J. (1985). Confidence limits on phylogenies: an approach using the bootstrap. Evolution 39, 783–791. doi: 10.1111/j.1558-5646.1985.tb00420.x
Galindo, R. C., Doncel-Pérez, E., Zivkovic, Z., Naranjo, V., Gortazar, C., Mangold, A. J., et al. (2009). Tick subolesin is an ortholog of the akirins described in insects and vertebrates. Dev. Comp. Immunol. 33, 612–617. doi: 10.1016/j.dci.2008.11.002
Giraldo-Calderón, G. I., Emrich, S. J., MacCallum, R. M., Maslen, G., Dialynas, E., Topalis, P., et al. (2015). VectorBase: an updated bioinformatics resource for invertebrate vectors and other organisms related with human diseases. Nucleic Acids Res. 43, D707–D713. doi: 10.1093/nar/gku1117
Gonzalez, K., and Baylies, M. (2005). “bhringi: A novel Twist co-regulator”, in Proceedings of the Program and Abstracts 46th Annual Drosophila Research Conference, San Diego, CA, 46:320B.
Goto, A., Fukuyama, H., Imler, J. L., and Hoffmann, J. A. (2014). The chromatin regulator DMAP1 modulates activity of the nuclear factor B (NF-B) transcription factor Relish in the Drosophila innate immune response. J. Biol. Chem. 289, 20470–20476. doi: 10.1074/jbc.C114.553719
Goto, A., Matsushita, K., Gesellchen, V., El Chamy, L., Kuttenkeuler, D., Takeuchi, O., et al. (2008). Akirins are highly conserved nuclear proteins required for NF-kappaB-dependent gene expression in Drosophila and mice. Nat. Immunol. 9, 97–104. doi: 10.1038/ni1543
Gregory, T. R. (2005). Animal Genome Size Database. Avilable at: http://www.genomesize.com
Gulia-Nuss, M., Nuss, A. B., Meyer, J. M., Sonenshine, D. E., Roe, R. M., Waterhouse, R. M., et al. (2016). Genomic insights into the Ixodes scapularis tick vector of Lyme disease. Nat. Commun. 7:10507. doi: 10.1038/ncomms10507
Hajdušek, O., Šíma, R., Ayllón, N., Jalovecká, M., Perner, J., de la Fuente, J., et al. (2013). Interaction of the tick immune system with transmitted pathogens. Front. Cell. Infect. Microbiol. 3:26. doi: 10.3389/fcimb.2013.00026
Harrington, D., Canales, M., de la Fuente, J., de Luna, C., Robinson, K., Guy, J., et al. (2009). Immunisation with recombinant proteins subolesin and Bm86 for the control of Dermanyssus gallinae in poultry. Vaccine 27, 4056–4063. doi: 10.1016/j.vaccine.2009.04.014
Holm, L., and Laakso, L. M. (2016). Dali server update. Nucleic Acids Res. 44, W351–W355. doi: 10.1093/nar/gkw357
Hoogstraal, H. (1956). African Ixodoidea. I. Ticks of the Sudan (with Special Reference to Equatoria Province and with Preliminary Reviews of the Genera Boophilus, Margaropus and Hyalomma). Washington, DC: Department of the Navy, 1101.
Horak, I. G., Heyne, H., Williams, R., Gallivan, G. J., Spickett, A. M., Bezuidenhout, J. D., et al. (2018). The Ixodid Ticks (Acari: Ixodidae) of Southern Africa. Hamburg: Springer, 675. doi: 10.1007/978-3-319-70642-9
Hou, F., Wang, X., Qian, Z., Liu, Q., Liu, Y., He, S., et al. (2013). Identification and functional studies of Akirin, a potential positive nuclear factor of NF-κB signaling pathways in the Pacific white shrimp, Litopenaeus vannamei. Dev. Comp. Immunol. 41, 703–714. doi: 10.1016/j.dci.2013.08.005
Ivics, Z., Hackett, P. B., Plasterk, R. H., and Izsvak, Z. (1997). Molecular reconstruction of Sleeping Beauty, a Tc1-like transposon from fish, and its transposition in human cells. Cell 91, 501–510. doi: 10.1016/S0092-8674(00)80436-5
Kasthuri, S. R., Umasuthan, N., Whang, I., Wan, Q., Lim, B. S., Jung, H. B., et al. (2013). Akirin2 homologues from rock bream, Oplegnathus fasciatus: genomic and molecular characterization and transcriptional expression analysis. Fish Shellfish Immunol. 35, 740–747. doi: 10.1016/j.fsi.2013.06.006
Katoh, K., Rozewicki, J., and Yamada, K. D. (2017). MAFFT online service: multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. doi: 10.1093/bib/bbx108 [Epub ahead of print].
Katoh, K., and Standley, D. M. (2013). MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol. Biol. Evol. 30, 772–780. doi: 10.1093/molbev/mst010
Kim, H., Lee, S. K., Hong, M. W., Park, S. R., Lee, Y. S., Kim, J. W., et al. (2013). Association of a single nucleotide polymorphism in the akirin 2 gene with economically important traits in Korean native cattle. Anim. Genet. 44, 750–753. doi: 10.1111/age.12055
Komiya, Y., Kurabe, N., Katagiri, K., Ogawa, M., Sugiyama, A., Kawasaki, Y., et al. (2008). A novel binding factor of 14-3-3beta functions as a transcriptional repressor and promotes anchorage-independent growth, tumorigenicity, and metastasis. J. Biol. Chem. 283, 18753–18764. doi: 10.1074/jbc.M802530200
Koonin, E. V. (2005). Orthologs, paralogs, and evolutionary genomics. Annu. Rev. Genet. 39, 309–338. doi: 10.1371/journal.pgen.1002821
Krossa, S., Schmitt, A. D., Hattermann, K., Fritsch, J., Scheidig, A. J., Mehdorn, H. M., et al. (2015). Down regulation of Akirin-2 increases chemosensitivity in human glioblastomas more efficiently than Twist-1. Oncotarget 6, 21029–21045. doi: 10.18632/oncotarget.3763
Kumar, B., Manjunathachar, H. V., Nagar, G., Ravikumar, G., de la Fuente, J., Saravanan, B. C., et al. (2017). Functional characterization of candidate antigens of Hyalomma anatolicum and evaluation of its cross-protective efficacy against Rhipicephalus microplus. Vaccine 35, 5682–5692. doi: 10.1016/j.vaccine.2017.08.049
Kumar, S., Stecher, G., and Tamura, K. (2016). MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 33, 1870–1874. doi: 10.1093/molbev/msw054
Kurland, C. G., Canbäck, B., and Berg, O. G. (2007). The origins of modern proteomes. Biochimie 89, 1454–1463. doi: 10.1016/j.biochi.2007.09.004
Law, M. J., Linde, M. E., Chambers, E. J., Oubridge, C., Katsamba, P. S., Nilsson, L., et al. (2006). The role of positively charged amino acids and electrostatic interactions in the complex of U1A protein and U1 hairpin II RNA. Nucleic Acids Res. 34, 275–285. doi: 10.1093/nar/gkj436
Leung, S. W., Harreman, M. T., Hodel, M. R., Hodel, A. E., and Corbett, A. H. (2003). Dissection of the karyopherin alpha nuclear localization signal (NLS)-binding groove: functional requirements for NLS binding. J. Biol. Chem. 278, 41947–41953. doi: 10.1074/jbc.M307162200
Liu, N., Wang, X. W., Sun, J. J., Wang, L., Zhang, H. W., Zhao, X. F., et al. (2016). Akirin interacts with Bap60 and 14-3-3 proteins to regulate the expression of antimicrobial peptides in the kuruma shrimp (Marsupenaeus japonicus). Dev. Comp. Immunol. 55, 80–89. doi: 10.1016/j.dci.2015.10.015
Liu, T., Gao, Y., and Xu, T. (2015). Evolution of akirin family in gene and genome levels and coexpressed patterns among family members and rel gene in croaker. Dev. Comp. Immunol. 52, 17–25. doi: 10.1016/j.dci.2015.04.010
Liu, X., Xia, Y., Tang, J., Ma, L., Li, C., Ma, P., et al. (2017). Dual roles of Akirin2 protein during Xenopus neural development. J. Biol. Chem. 292, 5676–5684. doi: 10.1074/jbc.M117.777110
Liu, Y., Song, L., Sun, Y., Liu, T., Hou, F., and Liu, X. (2016). Comparison of immune response in Pacific white shrimp, Litopenaeus vannamei, after knock down of Toll and IMD gene in vivo. Dev. Comp. Immunol. 60, 41–52. doi: 10.1016/j.dci.2016.02.004
Lu, P., Zhou, Y., Yu, Y., Cao, J., Zhang, H., Gong, H., et al. (2016). RNA interference and the vaccine effect of a subolesin homolog from the tick Rhipicephalus haemaphysaloides. Exp. Appl. Acarol. 68, 113–126. doi: 10.1007/s10493-015-9987-z
Macqueen, D. (2009). Commentary on Galindo et al. [Dev. Comp. Immunol. 33(4) (2009) 612-617]. Dev. Comp. Immunol. 33, 877; authorrely 878–879. doi: 10.1016/j.dci.2009.02.011
Macqueen, D. J., and Johnston, I. A. (2009). Evolution of the multifaceted eukaryotic akirin gene family. BMC Evol. Biol. 9:34. doi: 10.1186/1471-2148-9-34
Macqueen, D. J., Bower, N. I., and Johnston, I. A. (2010a). Positioning the expanded akirin gene family of Atlantic salmon within the transcriptional networks of myogenesis. Biochem. Biophys. Res. Commun. 200, 599–605. doi: 10.1016/j.bbrc.2010.08.110
Macqueen, D. J., Kristjánsson, B. K., and Johnston, I. A. (2010b). Salmonid genomes have a remarkably expanded akirin family, coexpressed with genes from conserved pathways governing skeletal muscle growth and catabolism. Physiol. Genomics 42, 134–148. doi: 10.1152/physiolgenomics.00045.2010
Maeda, I., Kohara, Y., Yamamoto, M., and Sugimoto, A. (2001). Large-scale analysis of gene function in Caenorhabditis elegans by high-throughput RNAi. Curr. Biol. 11, 171–176. doi: 10.1016/S0960-9822(01)00052-5
Makkerh, J. P., Dingwall, C., and Laskey, R. A. (1996). Comparative mutagenesis of nuclear localization signals reveals the importance of neutral and acidic amino acids. Curr. Biol. 6, 1025–1027. doi: 10.1016/S0960-9822(02)00648-6
Mans, B. J., De Castro, M. H., Pienaar, R., De Klerk, D., Gaven, P., Genu, S., et al. (2016). Ancestral reconstruction of tick lineages. Ticks Tick Borne Dis. 7, 509–535. doi: 10.1016/j.ttbdis.2016.02.002
Manzano-Román, R., Díaz-Martín, V., Oleaga, A., and Pérez-Sánchez, R. (2015). Identification of protective linear B-cell epitopes on the subolesin/akirin orthologues of Ornithodoros spp. soft ticks. Vaccine 33, 1046–1055. doi: 10.1016/j.vaccine.2015.01.015
Manzano-Román, R., Díaz-Martín, V., Oleaga, A., Siles-Lucas, M., and Pérez-Sánchez, R. (2012). Subolesin/akirin orthologs from Ornithodoros spp. soft ticks: cloning, RNAi gene silencing and protective effect of the recombinant proteins. Vet. Parasitol. 185, 248–259. doi: 10.1016/j.vetpar.2011.10.032
Marshall, A., Salerno, M. S., Thomas, M., Davies, T., Berry, C., Dyer, K., et al. (2008). Mighty is a novel promyogenic factor in skeletal myogenesis. Exp. Cell Res. 314, 1013–1029. doi: 10.1016/j.yexcr.2008.01.004
Medzhitov, R., and Janeway, C. A. Jr. (1997). Innate immunity: impact on the adaptive immune response. Curr. Opin. Immunol. 9, 4–9. doi: 10.1016/S0952-7915(97)80152-5
Merino, O., Alberdi, P., Pérez de la Lastra, J. M., and de la Fuente, J. (2013a). Tick vaccines and the control of tick-borne pathogens. Front. Cell. Infect. Microbiol. 3:30. doi: 10.3389/fcimb.2013.00030
Merino, O., Antunes, S., Mosqueda, J., Moreno-Cid, J. A., Pérez de la Lastra, J. M., Rosario-Cruz, R., et al. (2013b). Vaccination with proteins involved in tick-pathogen interactions reduces vector infestations and pathogen infection. Vaccine 31, 5889–5896. doi: 10.1016/j.vaccine.2013.09.037
Merino, O., Almazán, C., Canales, M., Villar, M., Moreno-Cid, J. A., Estrada-Peña, A., et al. (2011a). Control of Rhipicephalus (Boophilus) microplus infestations by the combination of subolesin vaccination and tick autocidal control after subolesin gene knockdown in ticks fed on cattle. Vaccine 29, 2248–2254. doi: 10.1016/j.vaccine.2011.01.050
Merino, O., Almazán, C., Canales, M., Villar, M., Moreno-Cid, J. A., Galindo, R. C., et al. (2011b). Targeting the tick protective antigen subolesin reduces vector infestations and pathogen infection by Anaplasma marginale and Babesia bigemina. Vaccine 29, 8575–8579. doi: 10.1016/j.vaccine.2011.09.023
Mobley, C. B., Fox, C. D., Ferguson, B. S., Amin, R. H., Dalbo, V. J., Baier, S., et al. (2014). L-leucine, beta-hydroxy-beta-methylbutyric acid (HMB) and creatine monohydrate prevent myostatin-induced Akirin-1/Mighty mRNA down-regulation and myotube atrophy. J. Int. Soc. Sports Nutr. 11:38. doi: 10.1186/1550-2783-11-38
Morel, P. C. (2003). Les Tiques d’Afrique et du Bassin Meìditerraneìen (1965–1995). Can Tho: CIRAD-EMVT, 1342.
Moreno-Cid, J. A., Jiménez, M., Cornelie, S., Molina, R., Alarcón, P., Lacroix, M.-N., et al. (2011). Characterization of Aedes albopictus akirin for the control of mosquito and sand fly infestations. Vaccine 29, 77–82. doi: 10.1016/j.vaccine.2010.10.011
Moreno-Cid, J. A., Pérez de la Lastra, J. M., Villar, M., Jiménez, M., Pinal, R., Estrada-Peña, A., et al. (2013). Control of multiple arthropod vector infestations with subolesin/akirin vaccines. Vaccine 31, 1187–1196. doi: 10.1016/j.vaccine.2012.12.073
Naranjo, N., Ayllón, N., Pérez de la Lastra, J. M., Galindo, R. C., Kocan, K. M., Blouin, E. F., et al. (2013). Reciprocal regulation of NF-kB (Relish) and Subolesin in the tick vector, Ixodes scapularis. PLoS One 8:e65915. doi: 10.1371/journal.pone.0065915
Nowak, S. J., Aihara, H., Gonzalez, K., Nibu, Y., and Baylies, M. K. (2012). Akirin links twist-regulated transcription with the Brahma chromatin remodeling complex during embryogenesis. PLoS Genet. 8:e1002547. doi: 10.1371/journal.pgen.1002547
Nowak, S. J., and Baylies, M. K. (2012). Akirin: a context-dependent link between transcription and chromatin remodeling. Bioarchitecture 2, 209–213. doi: 10.4161/bioa.22907
Olds, C. L., Mwaura, S., Odongo, D. O., Scoles, G. A., Bishop, R., and Daubenberger, C. (2016). Induction of humoral immune response to multiple recombinant Rhipicephalus appendiculatus antigens and their effect on tick feeding success and pathogen transmission. Parasit. Vectors 9:484. doi: 10.1186/s13071-016-1774-0
Orengo, C. A., and Thornton, J. M. (2005). Protein families and their evolution-a structural perspective. Annu. Rev. Biochem. 74, 867–900. doi: 10.1146/annurev.biochem.74.082803.133029
Pavithiran, A., Bathige, S. D. N. K., Kugapreethan, R., Priyathilaka, T. T., Yang, H., Kim, M. J., et al. (2018). A comparative study of three akirin genes from big belly seahorse Hippocampus abdominalis: molecular, transcriptional and functional characterization. Fish Shellfish Immunol. 74, 584–592. doi: 10.1016/j.fsi.2018.01.025
Peña-Rangel, M. T., Rodriguez, I., and Riesgo-Escovar, J. R. (2002). A misexpression study examining dorsal thorax formation in Drosophila melanogaster. Genetics 160, 1035–1050.
Prudencio, C. R., Pérez de la Lastra, J. M., Canales, M., Villar, M., and de la Fuente, J. (2010). Mapping protective epitopes in the tick and mosquito subolesin ortholog proteins. Vaccine 28, 5398–5406. doi: 10.1016/j.vaccine.2010.06.021
Qu, F., Xiang, Z., Zhang, Y., Li, J., Zhang, Y., and Yu, Z. (2014). The identification of the first molluscan Akirin2 with immune defense function in the Hong Kong oyster Crassostrea hongkongensis. Fish Shellfish Immunol. 41, 455–465. doi: 10.1016/j.fsi.2014.09.029
Rahman, M. K., Saiful Islam, M., and You, M. (2018). Impact of subolesin and cystatin knockdown by RNA interference in adult female Haemaphysalis longicornis (Acari: Ixodidae) on blood engorgement and reproduction. Insects 9:E39. doi: 10.3390/insects9020039
Richardson, J. M., Colloms, S. D., Finnegan, D. J., and Walkinshaw, M. D. (2009). Molecular architecture of the Mos1 paired-end complex: the structural basis of DNA transposition in a eukaryote. Cell 138, 1096–1108. doi: 10.1016/j.cell.2009.07.012
Rioualen, C., Da Costa, Q., Chetrit, B., Charafe-Jauffret, E., Ginestier, C., and Bidaut, G. (2017). HTS-Net: an integrated regulome-interactome approach for establishing network regulation models in high-throughput screenings. PLoS One 12:e0185400. doi: 10.1371/journal.pone.0185400
Salerno, M. S., Dyer, K., Bracegirdle, J., Platt, L., Thomas, M., Siriett, V., et al. (2009). Akirin1 (Mighty), a novel promyogenic factor regulates muscle regeneration and cell chemotaxis. Exp. Cell Res. 315, 2012–2021. doi: 10.1016/j.yexcr.2009.04.014
Sasaki, M., Akiyama-Oda, Y., and Oda, H. (2017). Evolutionary origin of type IV classical cadherins in arthropods. BMC Evol. Biol. 17:142. doi: 10.1186/s12862-017-0991-2
Sasaki, S., Yamada, T., Sukegawa, S., Miyake, T., Fujita, T., Morita, M., et al. (2009). Association of a single nucleotide polymorphism in akirin 2 gene with marbling in Japanese Black beef cattle. BMC Res. Notes 2:131. doi: 10.1186/1756-0500-2-131
Schetters, T., Bishop, R., Crampton, M., Kopáčk, P., Lew-Tabor, A., Maritz-Olivier, C., et al. (2016). Cattle tick vaccine researchers join forces in CATVAC. Parasit. Vectors 9:105. doi: 10.1186/s13071-016-1386-8
Shakya, M., Kumar, B., Nagar, G., de la Fuente, J., and Ghosh, S. (2014). Subolesin: a candidate vaccine antigen for the control of cattle tick infestations in Indian situation. Vaccine 32, 3488–3494. doi: 10.1016/j.vaccine.2014.04.053
Shaw, D. K., Wang, X., Brown, L. J., Oliva Chávez, A. S., Reif, K. E., Smith, A. A., et al. (2017). Infection-derived lipids elicit an immune deficiency circuit in arthropods. Nat. Commun. 8:14401. doi: 10.1038/ncomms14401
Smith, A., Guo, X., de la Fuente, J., Naranjo, N., Kocan, K. M., and Kaufman, W. R. (2009). The impact of RNA interference of the subolesin and voraxin genes in male Amblyomma hebraeum (Acari: Ixodidae) on female engorgement and oviposition. Exp. Appl. Acarol. 47, 71–86. doi: 10.1007/s10493-008-9195-1
Sonenshine, D. E., Kocan, K. M., and de la Fuente, J. (2006). Tick control: further thoughts on a research agenda. Trends Parasitol. 22, 550–551. doi: 10.1016/j.pt.2006.09.003
Staley, J. T. (2017). Domain Cell Theory supports the independent evolution of the Eukarya, Bacteria and Archaea and the Nuclear Compartment Commonality hypothesis. Open Biol. 7:170041. doi: 10.1098/rsob.170041
Sun, W., Huang, H., Ma, S., Gan, X., Zhu, M., Liu, H., et al. (2016). Akirin2 could promote the proliferation but not the differentiation of duck myoblasts via the activation of the mTOR/p70S6K signaling pathway. Int. J. Biochem. Cell. Biol. 79, 298–307. doi: 10.1016/j.biocel.2016.08.032
Tartey, S., Matsushita, K., Imamura, T., Wakabayashi, A., Ori, D., Mino, T., et al. (2015). Essential function for the nuclear protein Akirin2 in B cell activation and humoral immune responses. J. Immunol. 195, 519–527. doi: 10.4049/jimmunol.1500373
Tartey, S., Matsushita, K., Vandenbon, A., Ori, D., Imamura, T., Mino, T., et al. (2014). Akirin2 is critical for inducing inflammatory genes by bridging IκB-ζ and the SWI/SNF complex. EMBO J. 33, 2332–2348. doi: 10.15252/embj.201488447
Torina, A., Moreno-Cid, J. A., Blanda, V., Fernández de Mera, I. G., Pérez de la Lastra, J. M., Scimeca, S., et al. (2014). Control of tick infestations and pathogen prevalence in cattle and sheep farms vaccinated with the recombinant Subolesin-Major Surface Protein 1a chimeric antigen. Parasit. Vectors 7:10. doi: 10.1186/1756-3305-7-10
Valenzuela-Muñoz, V., and Gallardo-Escárate, C. (2014). TLR and IMD signaling pathways from Caligus rogercresseyi (Crustacea: Copepoda): in silico gene expression and SNPs discovery. Fish Shellfish Immunol. 36, 428–434. doi: 10.1016/j.fsi.2013.12.019
Villar, M., Marina, A., and de la Fuente, J. (2017). Applying proteomics to tick vaccine development: where are we? Expert Rev. Proteomics 14, 211–221. doi: 10.1080/14789450.2017.1284590
Voigt, F., Wiedemann, L., Zuliani, C., Querques, I., Sebe, A., Mates, L., et al. (2016). Sleeping Beauty transposase structure allows rational design of hyperactive variants for genetic engineering. Nat. Commun. 7:11126. doi: 10.1038/ncomms11126
Wan, F., and Lenardo, M. J. (2010). The nuclear signaling of NF-kappaB: current knowledge, new insights, and future perspectives. Cell Res. 20, 24–33. doi: 10.1038/cr.2009.137
Wang, M., Kurland, C. G., and Caetano-Anollés, G. (2011). Reductive evolution of proteomes and protein structures. Proc. Natl. Acad. Sci U.S.A. 108, 11954–11958. doi: 10.1073/pnas.1017361108
Watanabe, N., Satoh, Y., Fujita, T., Ohta, T., Kose, H., Muramatsu, Y., et al. (2011). Distribution of allele frequencies at TTN g.231054C > T, RPL27A g.3109537C > T and AKIRIN2 c.∗188G > A between Japanese Black and four other cattle breeds with differing historical selection for marbling. BMC Res. Notes 4:10. doi: 10.1186/1756-0500-4-10
Wolf, Y. I., and Koonin, E. V. (2013). Genome reduction as the dominant mode of evolution. Bioessays 35, 829–837. doi: 10.1002/bies.201300037
Xue, X., Wang, L., Chen, Y., Zhang, X., Luo, H., Li, Z., et al. (2014). Identification and molecular characterization of an Akirin2 homolog in Chinese loach (Paramisgurnus dabryanus). Fish Shellfish Immunol. 36, 435–443. doi: 10.1016/j.fsi.2013.12.021
Yan, J., Dong, X., Kong, Y., Zhang, Y., Jing, R., and Feng, L. (2013). Identification and primary immune characteristics of an amphioxus akirin homolog. Fish Shellfish Immunol. 35, 564–571. doi: 10.1016/j.fsi.2013.05.020
Yang, C. G., Wang, X. L., Wang, L., Zhang, B., and Chen, S. L. (2011). A new Akirin1 gene in turbot (Scophthalmus maximus): molecular cloning, characterization and expression analysis in response to bacterial and viral immunological challenge. Fish Shellfish Immunol 30, 1031–1041. doi: 10.1016/j.fsi.2011.01.028
Yang, C. G., Wang, X. L., Zhang, B., Sun, B., Liu, S. S., and Chen, S. L. (2013). Screening and analysis of PoAkirin1 and two related genes in response to immunological stimulants in the Japanese flounder (Paralichthys olivaceus). BMC Mol. Biol. 14:10. doi: 10.1186/1471-2199-14-10
Zanesi, N., Balatti, V., Riordan, J., Burch, A., Rizzotto, L., Palamarchuk, A., et al. (2013). A Sleeping Beauty screen reveals NF-kB activation in CLL mouse model. Blood 121, 4355–4358. doi: 10.1182/blood-2013-02-486035
Zhang, Y. (2008). I-TASSER server for protein 3D structure prediction. BMC Bioinformatics 9:40. doi: 10.1186/1471-2105-9-40
Zivkovic, Z., Esteves, E., Almazán, C., Daffre, S., Nijhof, A. M., Kocan, K. M., et al. (2010a). Differential expression of genes in salivary glands of male Rhipicephalus (Boophilus) microplus in response to infection with Anaplasma marginale. BMC Genomics 11:186. doi: 10.1186/1471-2164-11-186
Keywords: immune response, vaccine, interactome, regulome, phylogeny, tick, Anaplasma phagocytophilum
Citation: Artigas-Jerónimo S, Villar M, Cabezas-Cruz A, Valdés JJ, Estrada-Peña A, Alberdi P and de la Fuente J (2018) Functional Evolution of Subolesin/Akirin. Front. Physiol. 9:1612. doi: 10.3389/fphys.2018.01612
Received: 03 July 2018; Accepted: 25 October 2018;
Published: 13 November 2018.
Edited by:
Itabajara Da Silva Vaz Jr., Universidade Federal do Rio Grande do Sul (UFRGS), BrazilReviewed by:
Sukanya Narasimhan, Yale University, United StatesCarlos Alexandre Sanchez Ferreira, Pontifícia Universidade Católica do Rio Grande do Sul, Brazil
Copyright © 2018 Artigas-Jerónimo, Villar, Cabezas-Cruz, Valdés, Estrada-Peña, Alberdi and de la Fuente. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: José de la Fuente, jose_delafuente@yahoo.com
†These authors have contributed equally to this work