 Javier Fernández1,2,3
Javier Fernández1,2,3 F. J. Moreno4
F. J. Moreno4 Agustín Olano4
Agustín Olano4 Alfonso Clemente5
Alfonso Clemente5 Claudio J. Villar1,2,3
Claudio J. Villar1,2,3 Felipe Lombó1,2,3*
Felipe Lombó1,2,3*- 1Research Unit “Biotechnology in Nutraceuticals and Bioactive Compounds-BIONUC”, Departamento de Biología Funcional, Área de Microbiología, Universidad de Oviedo, Oviedo, Spain
- 2Instituto Universitario de Oncología del Principado de Asturias, Oviedo, Spain
- 3Instituto de Investigación Sanitaria del Principado de Asturias, Oviedo, Spain
- 4Instituto de Investigación en Ciencias de la Alimentación (CIAL-CSIC), Madrid, Spain
- 5Estación Experimental del Zaidín (EEZ-CSIC), Granada, Spain
Colorectal cancer (CRC) is one of the most common neoplasias worldwide, and its incidence is increasing. Consumption of prebiotics is a useful strategy in order to prevent this important disease. These nutraceutical compounds might exert protective biological functions as antitumors. In order to test the chemopreventive effect of GOS-Lu (galacto-oligosaccharides derived from lactulose) prebiotic preparation against this cancer, an animal model (Rattus norvegicus F344) was used. In this model, two doses of azoxymethane (10 mg/kg) and two treatments with dextran sodium sulfate (DSS) were administered to the animals. Animals were fed for 20 weeks, and either control drinking water or drinking water containing 10% (w/w) GOS-Lu prebiotic preparation was provided to them. Animals were sacrificed after those 20 weeks, and their digestive tract tissues were analyzed. The results revealed a statistically significant reduction in the number of colon tumors in the GOS-Lu cohort with respect to control animals. Metagenomics sequencing was used for studying colon microbiota populations, revealing significant reductions in populations of pro-inflammatory bacteria families and species, and significant increases in interesting beneficial populations, such as Bifidobacterium. Therefore, oral administration of the prebiotic GOS-Lu preparation may be an effective strategy for preventing CRC.
Introduction
Prebiotics have been recently redefined as a substrate that is selectively utilized by host microorganisms conferring a health benefit (Gibson et al., 2017). Prebiotics are typically metabolized by bifidobacteria and lactobacilli and their major beneficial effects seem to occur in the large intestine due to the slow transit of the substrates susceptible of fermentation and their effects on microbial diversity and metabolic fingerprinting, which play an important role in host health (Bindels et al., 2015). These effects include growth inhibition of potential pathogens, immune response stimulation, modulation of intestinal epithelial cells and production of short-chain fatty acids (SCFAs) as metabolic endpoints of carbohydrate fermentation. The most abundant SCFAs are acetate, butyrate and propionate (which constitute more than 95% of total SCFA content), playing a key role in maintaining intestinal homeostasis. These metabolites are linked to specific health aspects at gut level and elsewhere in the body, including pathogen exclusion, colonocyte function, epithelial cell proliferation and differentiation, energy intake and control of body weight, levels of secondary bile acids, mineral absorption (Ca, Fe, and Mg), cholesterol biosynthesis, glucose metabolism and insulin sensitivity (Canfora et al., 2015; Fernández et al., 2015; Koh et al., 2016).
Prebiotic compounds are non-digestible oligosaccharides with various origins and chemical characteristics. They differ in the chain length, monosaccharide composition, linkage type and degree of branching (Clemente, 2014). The most deeply studied oligosaccharides are human milk oligosaccharides (HMO), inulin, fructo-oligosaccharides (FOS), the disaccharide lactulose and galacto-oligosaccharides (GOS) derived from lactose. Recently, novel galacto-oligosaccharides derived from lactulose (GOS-Lu) have demonstrated their potential as prebiotic compounds. These can be enzymatically produced by transgalactosylation of lactulose (4-O-β-D-galactopyranosyl-D-fructose) using β-galactosidases from different microbial sources (Martínez-Villaluenga et al., 2008; Cardelle-Cobas et al., 2012; Díez-Municio et al., 2014). Scientific evidences supporting their potential application as emerging prebiotic ingredients exerting beneficial effects on the gastrointestinal tract have been gathered (Moreno et al., 2014). Thus, GOS-Lu have shown to be selective for bifidobacteria and lactic-acid bacteria following several in vitro fermentation studies (Cardelle-Cobas et al., 2008, 2009, 2011). This bifidogenic effect was further corroborated in growing rats fed 1% (w/w) of GOS-Lu (Hernández-Hernández et al., 2012), together with a significant and selective increase of Bifidobacterium animalis found in the caecum and colon sections (Marín-Manzano et al., 2013). In vitro (Ferreira-Lazarte et al., 2017) and in vivo (Hernández-Hernández et al., 2012) studies have revealed that GOS-Lu are significantly less digestible in rats than conventional GOS (enzymatically synthetized from lactose). The higher resistance to gastrointestinal digestion together with the presence of non-transgalactosylated lactulose, which is itself a prebiotic, instead of lactose suggests that GOS-Lu have a lower calorific content than conventional GOS products (Rastall, 2013). Another prebiotic-mediated beneficial effect ascribed to GOS-Lu is their capacity to improve iron absorption in an iron-deficient rat model (Laparra et al., 2014). Lastly, GOS-Lu has been reported to inhibit in vitro production of pro-inflammatory factors, such as TNF-α and IL-1β, by intestinal epithelial cells (Caco-2) stimulated by the pathogen Listeria monocytogenes CECT 935 (Laparra et al., 2013); in addition, GOS-Lu has been reported to exert preventive intestinal anti-inflammatory effects in the trinitrobenzenesulfonic acid model of rat colitis (Algieri et al., 2014).
Colorectal cancer (CRC) is one of the leading causes of cancer-related mortality worldwide in both men (after lung and prostate cancers) and women (after breast cancer), and it is expected to increase by 60% to more than 1.1 million deaths by 2030 (Merrill and Anderson, 2011; Bray et al., 2013; Ferlay et al., 2015; Arnold et al., 2017). CRC is a complex and heterogeneous disease that reflects a combination of hereditary (such as mutations in specific genes such as apc), environmental (such as tobacco, alcohol, etc.) and dietary factors (such as saturated fat, nitrosamines, benzopyrenes, low consumption of fruit and vegetables) (Jemal et al., 2011; Brenner et al., 2014). Accumulating data suggest that the gut microbiota, and particularly their metabolic end products, might exert a protective role against CRC development by influencing inflammation, DNA damage and apoptosis (Louis et al., 2014). Prebiotics have been showed to improve biomarkers associated to CRC, as they stimulate the growth and activity of gut beneficial bacterial populations (Gibson and Roberfroid, 1995; Roberfroid, 2007; Pompei et al., 2008; Bosscher et al., 2009), which generate diverse short-chain fatty acids (SCFAs) as acetate, propionate, butyrate, isobutyrate and valerate when feeding on these prebiotic fibers (Rumessen et al., 1990). Some of these SCFAs exert interesting antitumor properties, as they inhibit histone deacetylases, causing changes in the expression of diverse cell cycle key modulators, and inducing apoptosis in tumor colon cells (Roller et al., 2004a; Pool-Zobel, 2005; Verghese et al., 2005; Kim and Milner, 2007; Scharlau et al., 2009; Stein et al., 2012; Fernández et al., 2015).
Consequently, the aim of this work was to evaluate the potential chemopreventive effects of orally ingested GOS-Lu against CRC in an animal model (Rattus norvegicus F344). Tumors were chemically induced with azoxymethane (AOM) and dextran sodium sulfate (DSS). Diverse biochemical, physical and microbiological parameters were analyzed in these rats: body weight, number of hyperplastic Peyer’s patches, caecum weight, number of colon polyps and total tumor-affected area. The intestinal microbiota was also examined in the two animal cohorts (control rat feed, GOS-Lu), revealing significant differences.
Materials and Methods
Production and Characterization of GOS-Lu
Galacto-oligosaccharides derived from lactulose (Lu) were synthesized using a commercial lactulose preparation (670 g of lactulose per liter; Duphalac, Abbott Biologicals BV, Olst, Netherlands) and β-galactosidase from Aspergillus oryzae (16 U/mL; Sigma, St. Louis, MO, United States) (López-Sanz et al., 2015). The enzymatic reaction took place at pH 5.4, achieved after the addition of 3 mL of KOH 2M at 800 mL of Duphalac, and 50°C in an orbital shaker at 300 rpm for 24 h. Afterwards, the enzymatic reaction was stopped by heating at 110°C for 10 min. The resulting mixture contained 66% (w:w) of total carbohydrates.
The carbohydrate fraction was qualitatively and quantitatively determined by gas chromatography-flame ionization detector (GC-FID) as trimethyl silylated oxime (TMSO) derivatives following previous approaches (Cardelle-Cobas et al., 2009; Hernández-Hernández et al., 2012). The carbohydrate composition of GOS-Lu, whose main involved glycosidic linkage was β(1→6), was as follows: fructose (19.5%), galactose (12.4%), glucose (1.2%), lactulose (24.7%), GOS-Lu disaccharides (13.6%), GOS-Lu trisaccharides (22.6%), GOS-Lu tetrasaccharides (5.1%) and GOS-Lu pentasaccharides (1.0%).
Animals and Experimental Design
Twenty male Fischer 344 rats (5 weeks old) were kept at the authorized facility No. ES330440003591 (University of Oviedo), and experiments were started after approval by the Ethics Committee of the Principado de Asturias (PROAE 16/2015).
Rats were separated in two groups of 10 animals each one. Groups 1 and 2 were fed ad libitum with universal feed (2014 Teklad Global 14% Protein Rodent Maintenance Harlan diet feed, Harlan Laboratories, Barcelona, Spain). The composition of this diet is the following one1: protein 14.3%, fat 4%, carbohydrate 48%, crude fiber 4.1%, neutral detergent fiber 18%, ash 4.7%, energy 2.9 kcal/g.
Group 1 received drinking water. Group 2 received 10% (w/w) GOS-Lu dissolved in the drinking water (average daily intake of GOS-Lu per rat was 2 g).
Rats number 9 and 10 of each group were kept free of CRC induction, as absolute control animals.
Colorectal Cancer Induction and Monitoring
One week after the animals arrived, drinking water or 10% (w/w) GOS-Lu in drinking water was provided continuously. After 1 week of drinking the corresponding liquid, CRC was induced in eight rats from each cohort. The two other rats were kept free of CRC induction as absolute control animals. Azoxymethane (AOM, Sigma-Aldrich, Madrid, Spain) was used for CRC induction in 8 rats of each group. AOM was dissolved in sterile saline (0.9% w:v NaCl) at 2 mg/mL and injected intraperitoneally (10 mg/kg body weight). The AOM treatment was repeated 1 week after first injection (weeks 2 and 3). The 2 control rats per group received sterile saline injections.
Rats received 3% and 2% (w:v) dextran sodium sulfate (DSS, 40.000 g/mol, VWR) in drinking water during 7 days, on weeks 4 and 15, respectively. This ulcerative colitis challenge was repeated twice in order to reinforce the pro-carcinogenic exerted by AOM. All rats were sacrificed at weeks 20 (pneumothorax). During those 20 weeks, rats were monitored for stool consistency, rectal bleeding and body weight.
Body Weight
Weight was measured along the 20 experimental weeks: arrival of animals (week 1), both AOM administrations (weeks 2 and 3), both DSS challenges (weeks 4 and 15), at week 6 and at sacrifice.
Blood and Tissue Samples
At week 20, rats were anesthetized with isoflurane and sacrificed (pneumothorax). All caecums were weighed and frozen at -20°C.
Colon was opened along main axis, washed with PBS (phosphate buffer saline) and kept in 4% v:v formaldehyde (4°C). The tumors number (from 1 to 9.5 mm diameter) were counted in the colon mucosa. Tumor morphologies were annotated as pedunculated, circular, spherical, and plane irregular, in order to get the total tumor-affected area.
GC-MS Quantification of SCFAs in Feces Using Deuterated Standards
Four hundred milligrams of frozen caecum feces were thawed and resuspended in 1,716 μL milli-Q H2O in 5 mL glass vials, homogenized in vortexed. Then, deuterated SCFAs standards were added as internal controls: deuterated acetate, butyrate, propionate and valerate (Cambridge Isotope Laboratories, United States), to a final concentration of 0.4 mM each one. Finally, 400 μL of 50% H2SO4 and 800 mg NaCl were added as well. This mixture was resuspended and 1 mL of ethyl acetate was added as extraction solvent. Samples were stirred for 1 h at 300 rpm and 25°C, and centrifuged for 5 min at 3500 rpm. 500 μL supernatants were taken to a new vial. This extraction was repeated twice.
The gas chromatography-mass spectrometry (GC-MS) equipment was an Agilent 7890A (Agilent Technologies), equipped with an inert crosslink mass selective detector (XL MSD) with triple-Axis detector. Acquisition was done using Chemstation software. The capillary chromatographic column was DB-FFAP (30 m, 0.25 mm ID, 0.25 μm film thickness). Helium was used as the carrier gas at 1 mL/min. Injection was made in splitless mode with an injection volume of 1 μL and an injector temperature of 200°C. A glass liner with a glass wool plug at the lower end of the liner was used to avoid the contamination of the GC column with non-volatile fecal material. A blank sample was inserted between experimental samples, to check for memory effects.
The column temperature was initially 50°C (1 min), then was increased to 150°C at 5°C/min, and finally to 230°C at 15°C/min (total time 20 min). The temperature of the ion source, quadrupole, and interface were 230, 150, and 220°C, respectively. Scanning ions were 45 and 76 m/z for deuterated propionic acid, 45 and 74 m/z for propionic acid, 43 and 73 m/z for isobutiric acid, 63 and 77 m/z for deuterated butyric acid, 60 and 73 m/z for butyric acid, 60 and 87 m/z for isovaleric acid, 63 and 77 m/z for deuterated isovaleric acid, 60 and 73 m/z for valeric acid and 60, 73, and 87 m/z for hexanoic acid. Identification of the different SCFAs was based on the retention time of standards and with the assistance of the Wiley 7 library.
Genomic DNA Extraction and 16S Ribosomal RNA Sequencing for Metagenomics
E.Z.N.A.® DNA Stool Kit (Ref. D4015-02, VWR, Madrid, Spain) was used for genomic DNA (gDNA) extraction (200 mg of frozen caecum feces). A BioPhotometer® (Eppendorf, Madrid, Spain) was used for gDNA quantification, a prior step before preparing working solutions diluted to 6 ng/μL, which were needed for PCR amplification using the Ion 16TM Metagenomics kit (Thermo Fischer Scientific, Madrid, Spain).
PCR amplicons were used to generate a library (Ion Plus Fragment Library kit for AB Library BuilderTM System, Cat. No. 4477597, Thermo Fischer Scientific, Madrid, Spain). The indexing of each sample was carried out with the Ion XpressTM Barcode Adapters 1-96 kit (Cat. No. 4474517, Thermo Fischer Scientific, Madrid, Spain). The ION OneTouchTM 2 System and the ION PGMTM Hi-QTM OT2 kit (Cat. No. A27739, Thermo Fischer Scientific, Madrid, Spain) was used for preparing the templates. The IONTM PGM Hi-QTM Sequencing kit (Cat. No. A25592, Thermo Fischer Scientific, Madrid, Spain) on the ION PGMTM System was used for metagenomics sequencing. The ION 318TM v2 Chip (Cat. No. 4484355, Thermo Fischer Scientific, Madrid, Spain) was used.
Phylogenetic Analysis
For each rat metagenomics, the consensus spreadsheet (ION Reporter software 5.6, Life Technologies Holdings Pte. Ltd., Singapore) included the percentages for each phylum, class, order, family or genus/species. These data were used in order to compare frequencies between experimental groups. Taxonomic adscription up to species level was performed using the QIIME 2 (v.2017.6.0) open-source bioinformatics pipeline. Analysis of the microbiome community was carried out using R software (v3.2.4): non-supervised multivariate analysis (PCA). For LDA analysis, tab-delimited files were generated in R and computed at family level using Galaxy. Graphical representation of Galaxy output included only discriminative features with logarithmic LDA score higher than 3. The reference library used was the Curated MicroSEQ(R) 16S Reference Library v2013.1; Curated Greengenes v13.5. The number of mapped reads (after the ignored ones due to less than 10 copies) per sample was always over 60.000. Total number of reads was always over 110.000. Counts were normalized by sum scaling. All raw metagenomics data have been deposited at NCBI SRA database (submission ID SRP155959).
Statistical Methods
Shapiro–Wilk’s test was used for calculating the Gaussian distribution of the different variables. Data were then expressed as the mean value ± SEM (standard error of mean). t-Test and other parametric methods were used for showing these data. Levene’s test was used for checking the similarity of variances. In the case of normal distribution, unpaired t-Test (when variances were similar) or Welch t-Test (when variances were not similar) were used for determining the statistical differences. In the case of no normal distribution, the non-parametric Mann–Whitney test was used for determining the statistical differences among cohorts.
GraphPad Prism software version 7 (GraphPad Software, San Diego, CA, United States) was used for the graphical representations: a p-value < 0.05 was considered statistically significant.
Results
Effect of GOS-Lu on Body Weight
Body weight gain were similar for all rat groups along the 20 experimental weeks (the first AOM challenge for CRC induction took place at week 2) (see Supplementary Table S2). When the animals were sacrificed, the mean value for the control cohort was 391.1 ± 40.5 g whereas for the GOS-Lu cohort was 367.1 ± 17.3 g (Figure 1).
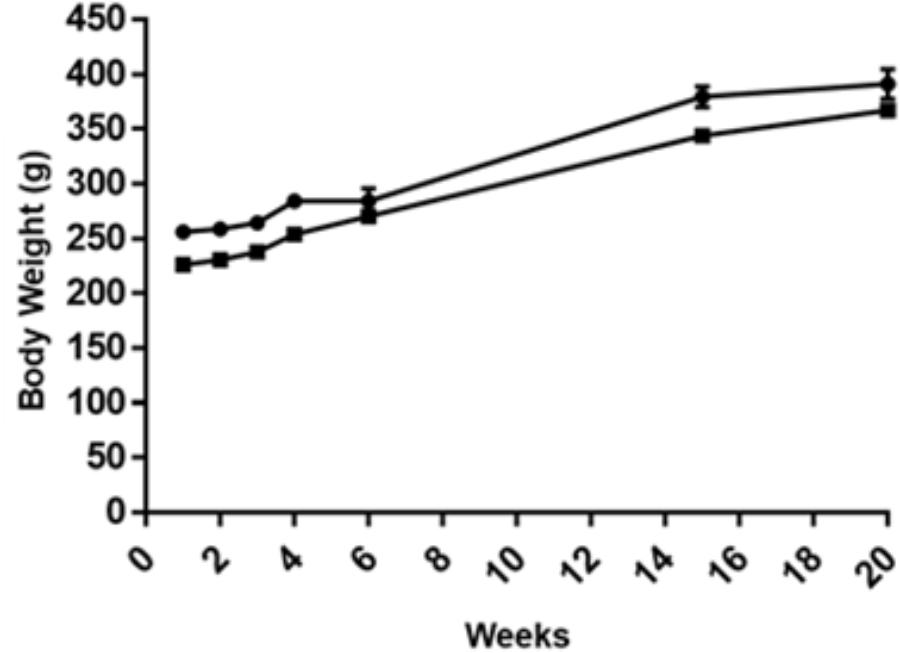
FIGURE 1. Body weight along the experimental time for the eight animals with CRC induction in the two groups: control (circles), GOS-Lu (squares). Body weight was measured at weeks 1, 2, 3, 4, 7, 15, and 20.
The second DSS challenge caused the death of rat number 3 (control cohort) due to intense rectal bleeding. This transitional ulcerative colitis process was a pro-inflammatory step necessary to increase the final tumor numbers and sizes.
Effect of GOS-Lu on Caecum Weight
Statistically significant differences in the caecum weight values were observed between the control cohort and the GOS-Lu cohort. These mean values were increased in the GOS-Lu cohort (7.63 ± 0.4 g) with respect to the control cohort (5.64 ± 0.4 g) and these differences were statistically significant (p-values 0.001) (Figure 2) (see Supplementary Table S2). Caecums from GOS-Lu cohort showed a 35.28% increase, due to the stimulation of bacterial populations caused by the presence of prebiotic compounds. In rodents, fermentation of prebiotic compounds starts in this organ.
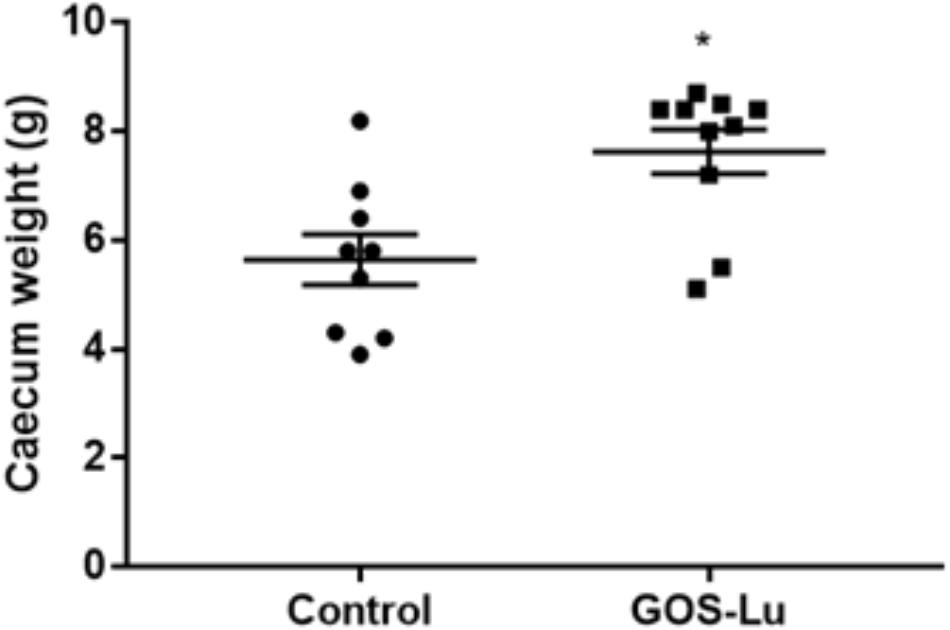
FIGURE 2. Mean of caecum weight in grams for each cohort.
Effect of GOS-Lu on Number of Polyps and Tumor-Affected Area
The colonic mucosa from each animal was analyzed for the number of polyps. Polyp diameter ranged from 1 to 9.5 mm (see Supplementary Table S2). A statistically significant difference was observed between rats in the control cohort and those in the GOS-Lu cohort. Polyps number decreased in the case of the GOS-Lu group (24.8 ± 1.5) with respect to the control group (58.5 ± 9.5) and this difference was statistically significant (p-value 0.0022). The GOS-Lu cohort showed a drastic 57.5% reduction in the number of polyps (Figure 3A).
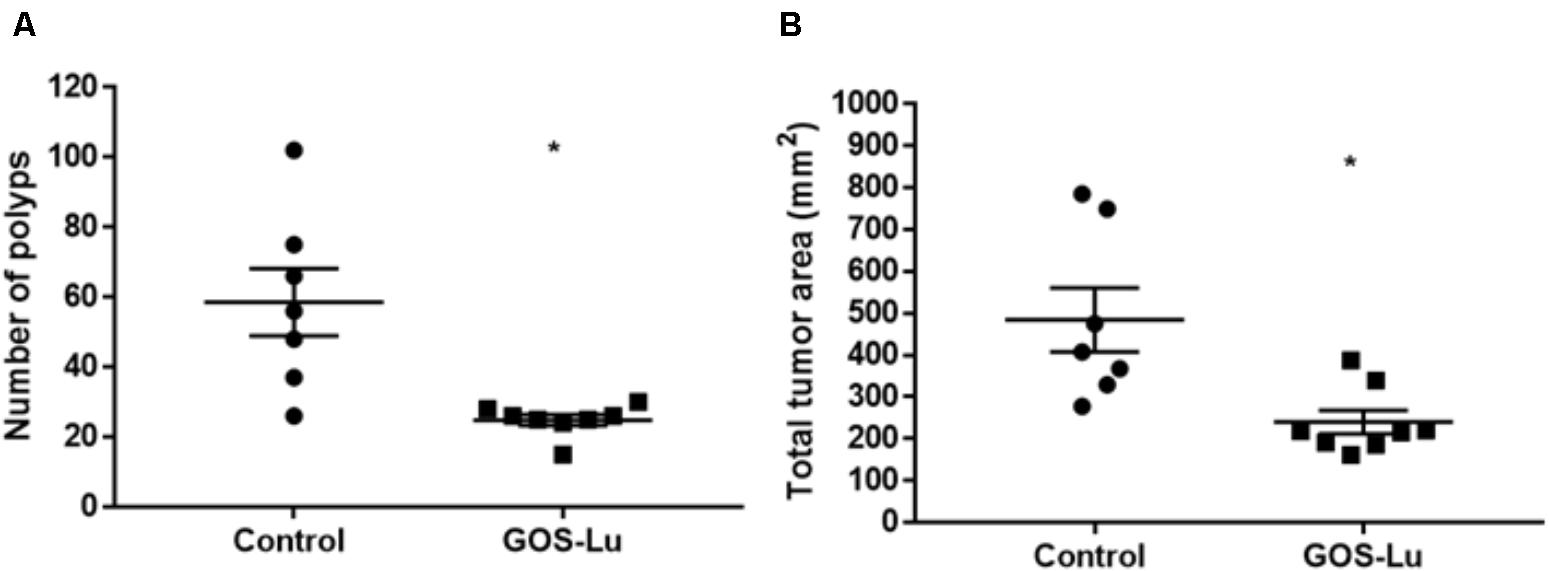
FIGURE 3. Measurements of colon polyps. (A) Average number of colon polyps. (B) Average sum of polyp areas.
Each polyp area was also calculated depending on its shape, computing the total tumor area for each rat. A statistically significant reduction was observed in the mean affected area of the GOS-Lu cohort (240 ± 28.1 mm2) with respect to the control cohort (484.8 ± 76.6 mm2). Therefore, the GOS-Lu cohort showed a 50.4% reduction in this parameter with respect to the control cohort (Figure 3B).
Effect of GOS-Lu on SCFAs Production
Cecal production of SCFAs is important, as these compounds show interesting antitumor properties regarding CRC prevention. Six SCFAs were identified and quantified in cecal content: propionate, butyrate, isobutyrate, valerate, isovalerate, and hexanoate. Propionate concentrations in control (0.86 ± 0.1) and GOS-Lu (1.35 ± 0.03) cohorts showed a statistically significant difference, with a 56.9% higher production in the GOS-Lu cohort (p-value 0.003) (Figure 4A). Concentrations of butyrate in control (1.29 ± 0.2) and GOS-Lu (0.51 ± 0.1) cohorts, of hexanoate in control (0.73 ± 0.2) and GOS-Lu (0.13 ± 0.06) cohorts, of valerate in control (0.07 ± 0.004) and GOS-Lu (0.06 ± 0.008) cohorts, of isobutyrate in control (0.04 ± 0.01) and GOS-Lu (0.03 ± 0.009) cohorts, and of isovalerate in control (0.004 ± 0.0007) and GOS-Lu (0.003 ± 0.0006) cohorts showed no statistically significant differences (Figure 4) (see Supplementary Table S2).
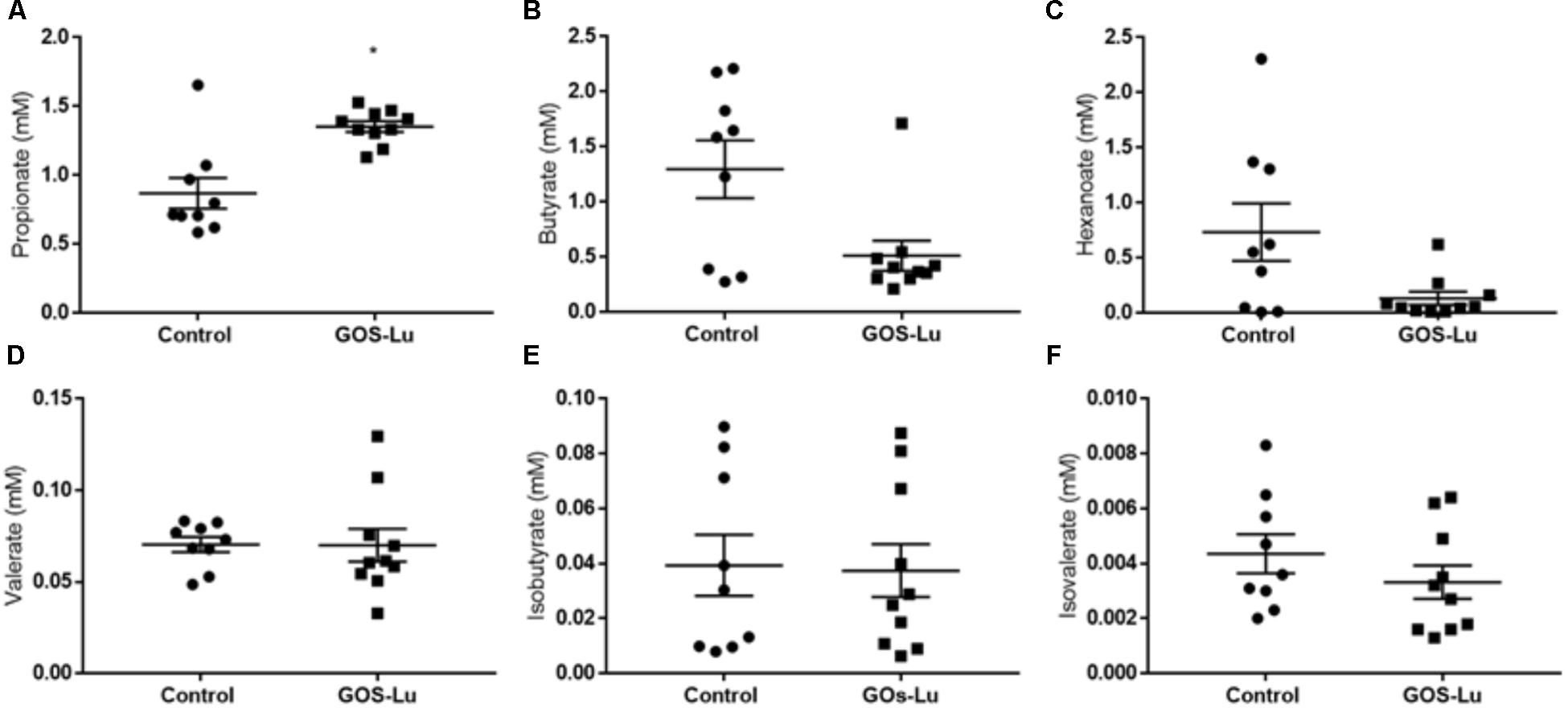
FIGURE 4. Cecal short chain fatty acids concentrations. (A) Propionate. (B) Butyrate. (C) Hexanoate. (D) Valerate. (E) Isobutyrate. (F) Isovalerate.
Effect of GOS-Lu on Intestinal Microbiota
Average phyla compositions of the two cohorts showed important differences between both animal groups (Table 1). The main differences between these cohorts is a 39.18% higher proportion in Bacteroidetes in the GOS-Lu cohort, a 19.35% reduction in Firmicutes populations in GOS-Lu cohort, and the near absence of Proteobacteria in the rats of the control cohort (0.34%), whereas Proteobacteria is the third most common phylum in the GOS-Lu cohort (9.49%) (Table 1). The predominant phylum in both cohorts was Firmicutes (69.14% in the control cohort, 55.76% in the GOS-Lu cohort) (Table 1). GOS-Lu increased the abundance of Actinobacteria and decreased Tenericutes (Table 1) (see Supplementary Table S1).
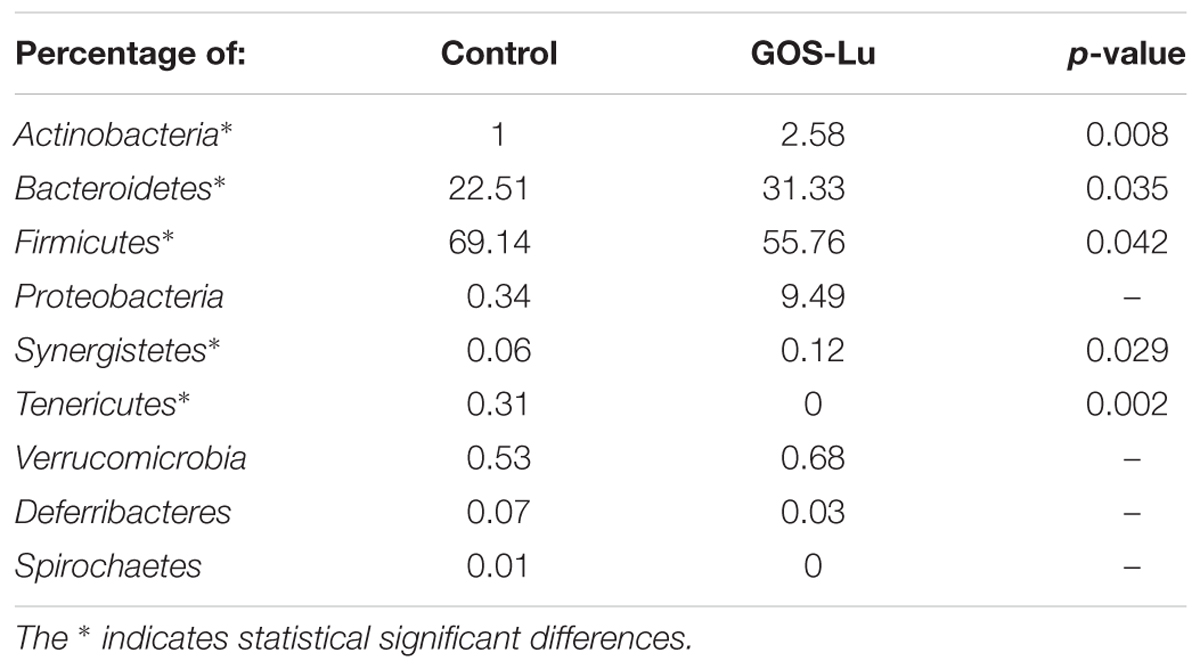
TABLE 1. Average percentage composition of intestinal microbiota at phylum level for the two cohorts studied.
The Firmicutes/Bacteroidetes ratio, described as an obesity microbiome marker, showed that it was significantly higher in the control cohort (3.86 ± 0.75) than in GOS-Lu cohort (1.95 ± 0.25) (p-value 0.03) (Figure 5).
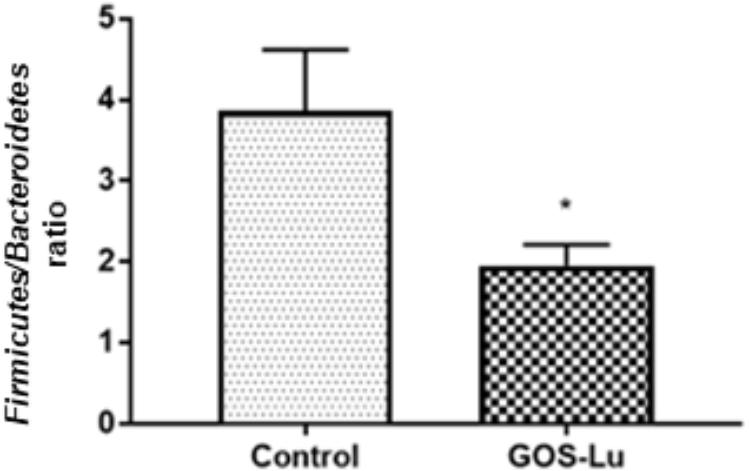
FIGURE 5. Graphical representation of the Firmicutes/Bacteroidetes ratio in both rat cohorts. ∗ Means a statistical significant difference between both cohorts.
As a general description of the gut microbiota composition, the Shannon diversity index was increased in the GOS-Lu cohort. This index is 3.47 (±0.17) for control cohort and 4.32 (±0.09) for GOS-Lu cohort (statistical significant differences, with a p-value of 0.0004) (Figure 6A). The Chao and the Simpson diversity indexes indicate an increase in the gut microbiota diversity in GOS-Lu cohort as well. Chao index is 33.56 (±3.66) for control cohort and 46.2 (±1.13) for GOS-Lu cohort (statistical significant differences, with a p-value of 0.0086) (Figure 6B). Simpson index is 0.8494 (±0.02) for control cohort and 0.9177 (±0.01) for GOS-Lu cohort (statistical significant differences, with a p-value of 0.0086) (Figure 6C). A homogeneous distribution between all rats in each cohort was found at phyla level.
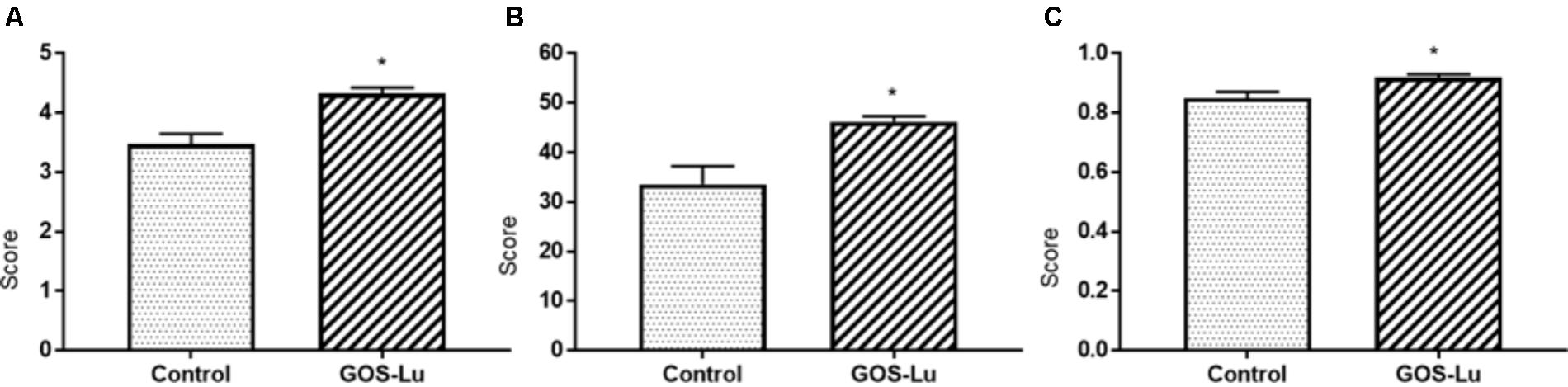
FIGURE 6. Graphical representation of the diversity indexes associated to the gut microbiota. (A) Shannon. (B) Chao. (C) Simpson.
At the family level, in the control cohort, the most abundant families were Lachnospiraceae (33.47%), Porphyromonadaceae (14.17%), Clostridiaceae (10.56%), Lactobacillaceae (9.96%), and Bacteroidaceae (4.22%) (Figure 7). In the case of GOS-Lu cohort, the most abundant ones were Lachnospiraceae (14.18%), Porphyromonadaceae (13.78%), Lactobacillaceae (13.06%) Clostridiaceae (9.49%), and Bacteroidaceae (9.40%) (Figure 7). These compositions at family level were homogeneous between animals in each cohort (Figure 8) (see Supplementary Table S1).
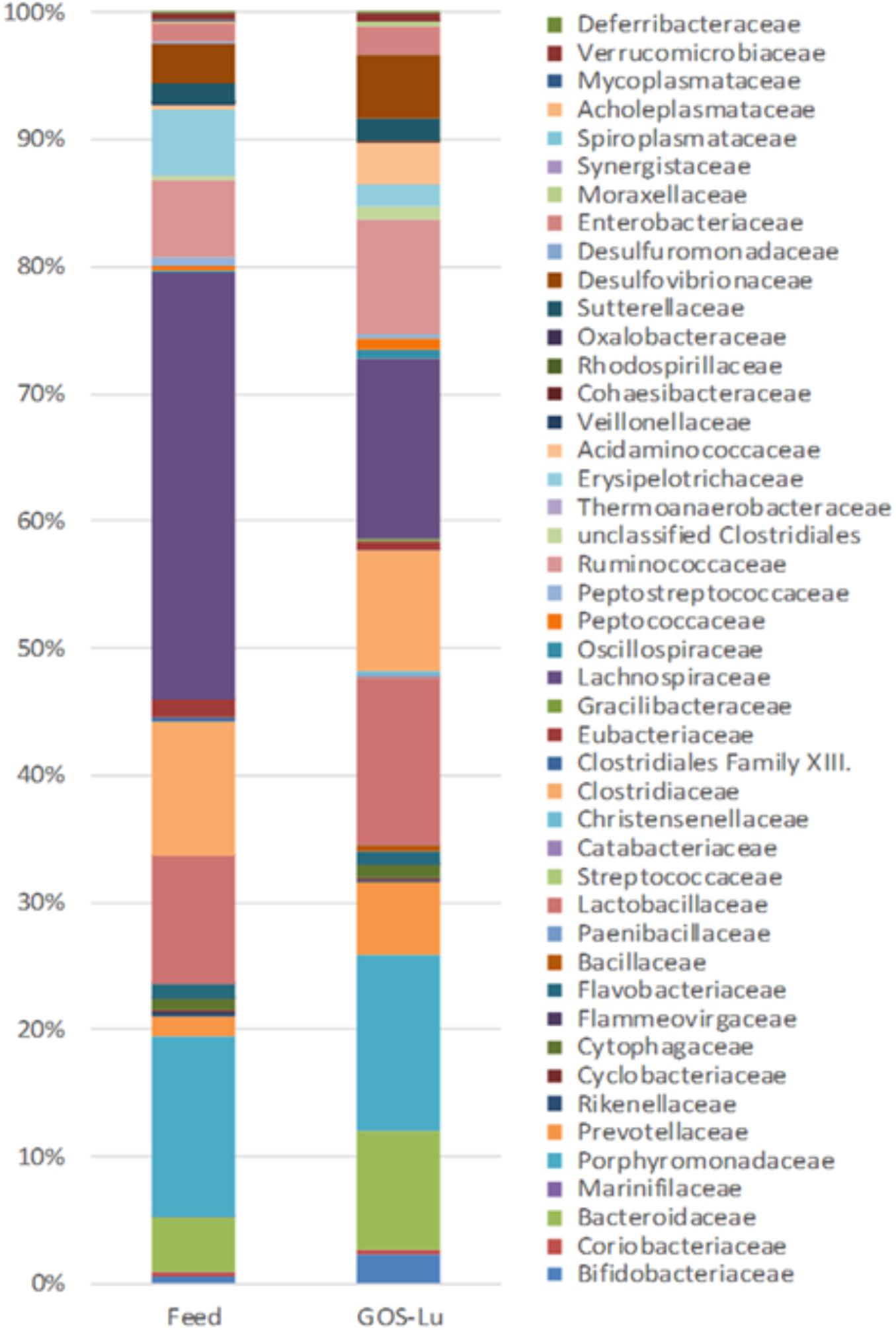
FIGURE 7. Differences in average intestinal microbiota composition at the family level for both cohorts.
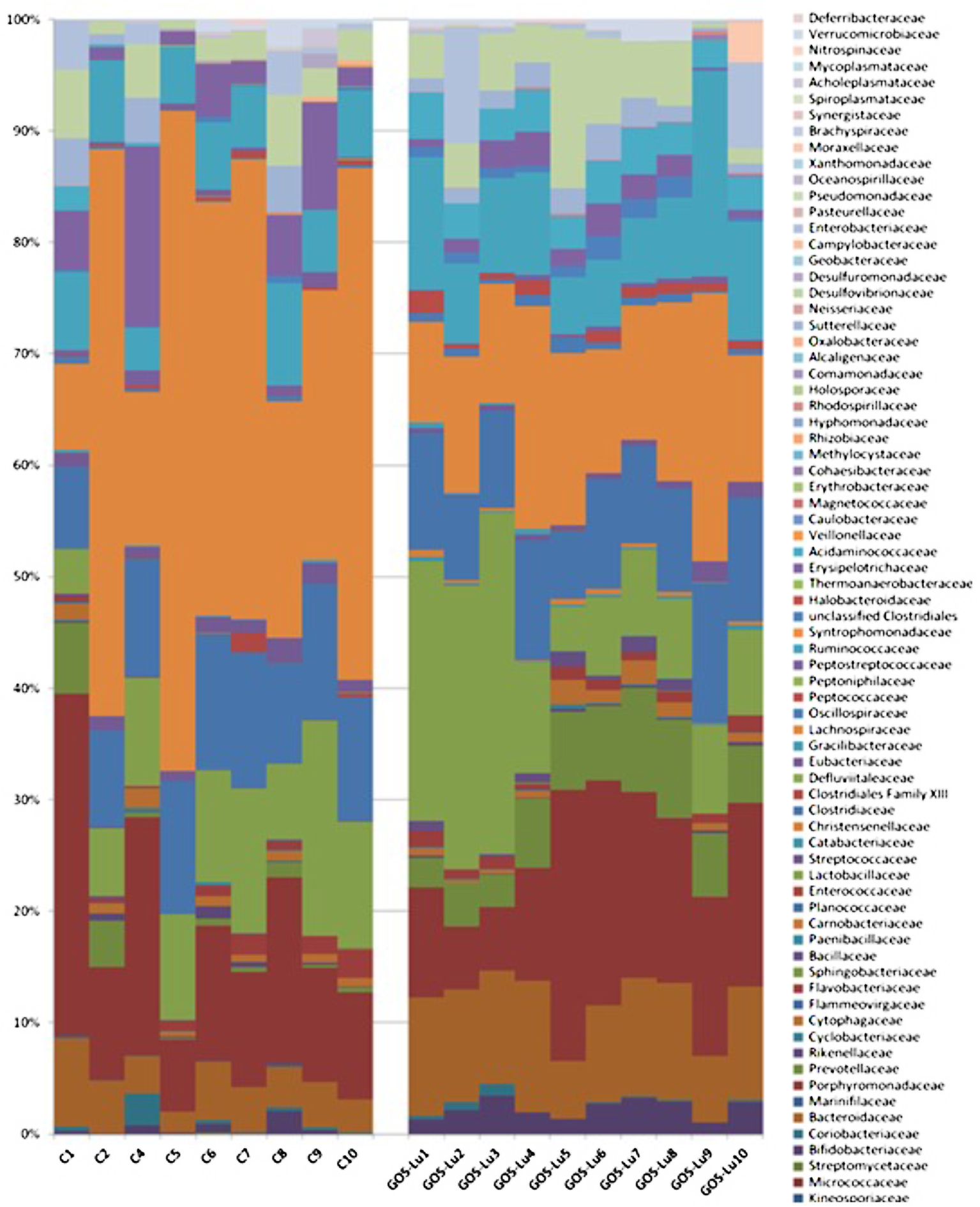
FIGURE 8. Intestinal microbiota composition at the family level in the rats belonging to the control cohort (C1–C10), and in the rats belonging to the GOS-Lu cohort (GOS-Lu1 to GOS-Lu10).
Also at family level, statistically significant differences were found mainly in the case of Lachnospiraceae (33.47% ± 5.87 in control cohort and 14.18% ± 1.49 in GOS-Lu cohort), Bacteroidaceae (4.22% ± 0.57 in control cohort and 9.40% ± 0.68 in GOS-Lu cohort), Prevotellaceae (1.58% ± 0.72 and 5.84 ± 0.002, respectively), Acidaminococcaceae (0.32% ± 0.24 and 3.27% ± 0.18, respectively), Bifidobacteriaceae (0.50% ± 0.22 and 2.30% ± 0.27, respectively), Eubacteriaceae (1.33% ±0.14 and 0.71% ± 0.16, respectively), Peptococcaceae (0.27% ± 0.05 and 0.86% ± 0.16, respectively), Oscillospiraceae (0.25% ± 0.06 and 0.71% ± 0.10, respectively) and Acholeplasmataceae (0.23% ± 0.16 and 0%, respectively) (Figure 9).
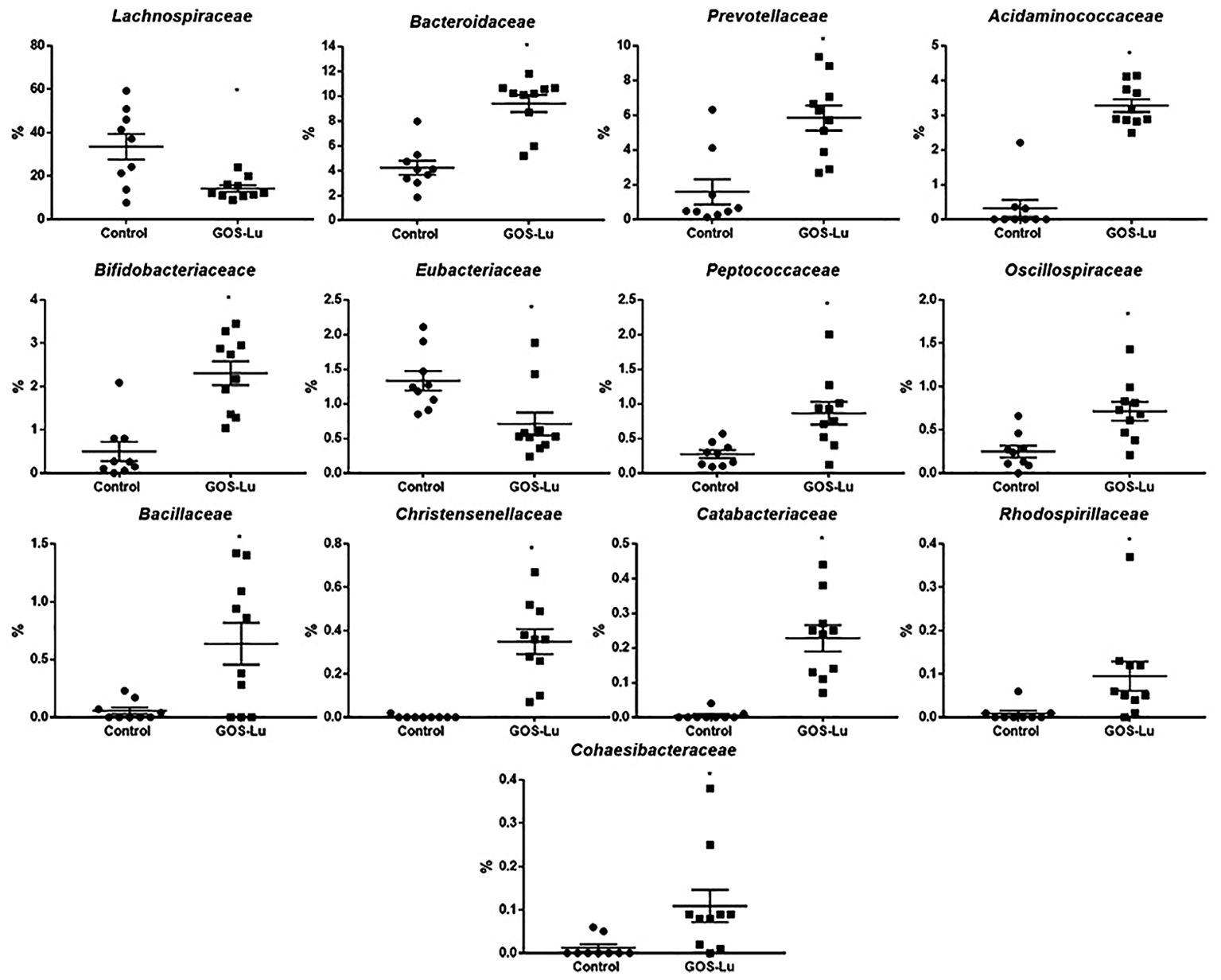
FIGURE 9. Families showing statistical significant differences between the rats belonging to the control and GOS-Lu cohorts.
The observed statistical significant increase in Actinobacteria phylum (Table 1) was mainly due to an increment in the Bifidobacteriaceae family (0.5% in control cohort, 2.31% in GOS-Lu cohort) (Figures 7, 9). In the case of Tenericutes phylum, its observed decrease was mainly due to a reduction in Acholeplasmataceae family (0.23% in control cohort, 0% in GOS-Lu cohort) (Figure 7).
The Bacteroidaceae (4.22% in control cohort, 9.4% in GOS-Lu cohort) and Prevotellaceae (1.59% in control cohort, 5.84% in GOS-Lu cohort) families were the remarkable groups from the Bacteroidetes phylum that showed an increase in the GOS-Lu cohort, compared to the control group (Figures 7, 9).
The decrease in Firmicutes phylum which took place in the GOS-Lu cohort was mainly due to a reduction of the Lachnospiraceae family (33.47% in control cohort, 14.18% in GOS-Lu cohort) and to a lesser extent, to a reduction in Eubacteriaceae family (1.33% in control cohort, 0.71% in GOS-Lu cohort) as well (Figures 7, 9).
Despite the decline of Firmicutes phylum in the GOS-Lu cohort, several families showed a small but significant increase in their numbers, such as Bacillaceae, Oscillospiraceae, Peptococcaceae, Catabacteriaceae, Christensenellaceae, unclassified Clostridiales and Acidaminococcaceae (Figures 7, 9). PCA of gut microbiota composition divided the animals in two clusters, indicating differences in the gut microbiota composition associated to both dietary interventions, feed and GOS-Lu supplementation (Figure 10A). Bacterial genera and species with significant differences in their relative abundances between the GOS-Lu and the control groups are indicated in Table 2 and the LDA analysis is represented in Figure 10B: in total, 27 families explain in a significant way the two types of diet. Finally, the relative abundance at genus and species levels was studied. Most significant differences were associated to higher proportion of some genera in the GOS-Lu cohort, such us Bifidobacterium, Bacteroides, Parabacteroides and others (Table 2). Only a few significant genera and species showed a reduction in GOS-Lu cohort, such as Lactobacillus intestinalis, Turicibacter spp., and Desulfovibrio spp. (Table 2).
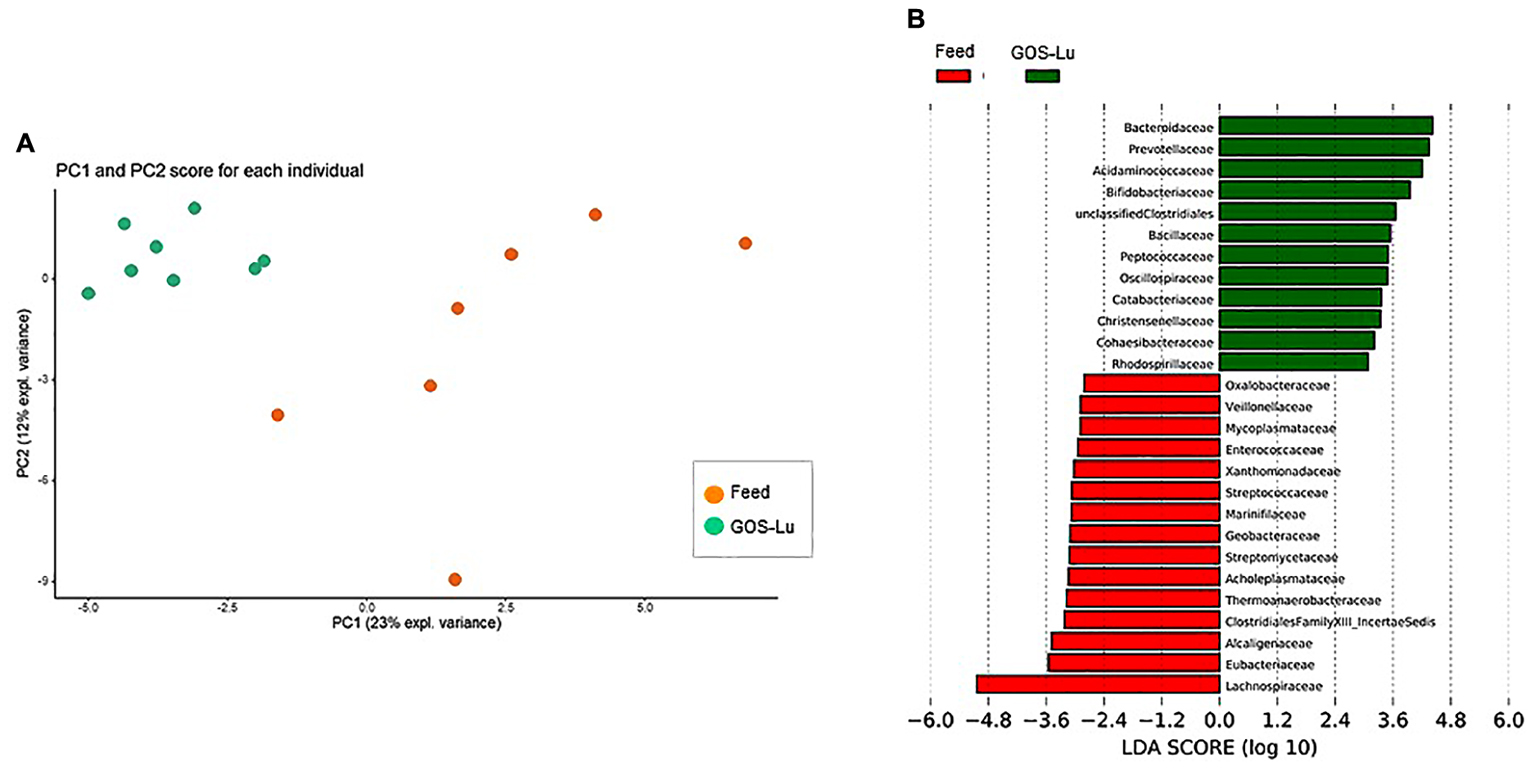
FIGURE 10. (A) Gut microbiota PCA cluster analysis. (B) LDA analysis showing the families that better discriminate between both cohort conditions.
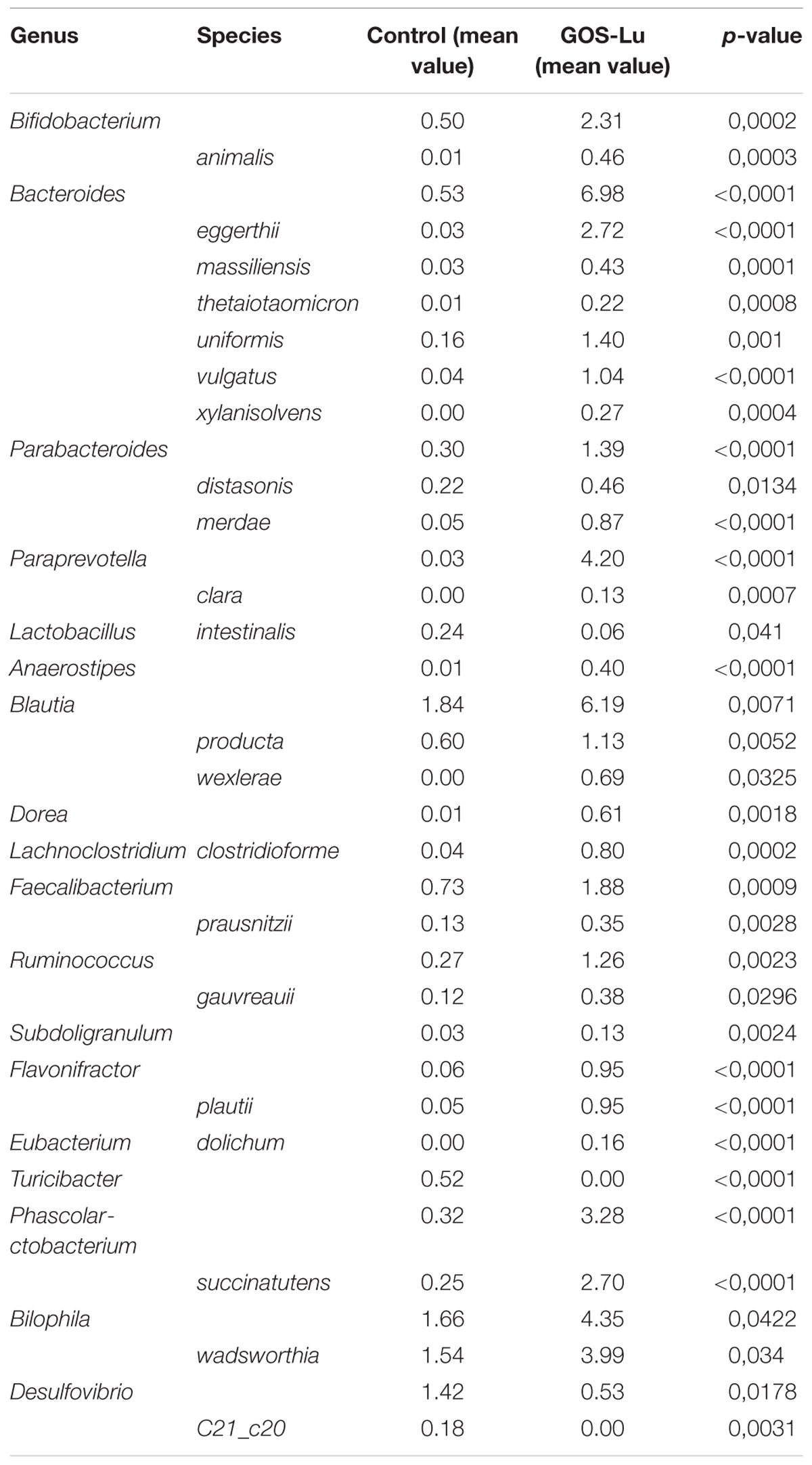
TABLE 2. Statistical significant differences in intestinal microbiota composition at the genus and species level for the rats belonging to the control and GOS-Lu cohorts.
Discussion
In this work, the antitumor effect of GOS-Lu as a prebiotic compound was studied in an animal model where CRC was generated using AOM/DSS. This GOS-Lu prebiotic preparation contains some amounts of monosaccharides (33.1%) and lactulose (24.7%) from the enzymatic synthesis reaction. It contains also prebiotic disaccharides (13.6%), trisaccharides (22.6%), tetrasaccharides (5.1%), and pentasaccharides (1%). In total, these prebiotics (galactosyl-fructoses) account for 42.3%, a nearly two-fold proportion to lactulose. GOS-Lu was used here without any prior purification step (including monosaccharides and the precursor lactulose substrate) in a similar way to the conventional GOS used by food industry. This is a common practice for prebiotics acting as food ingredients (for instance, following their addition to infant formulae) to make their use more cost-effective and feasible, since the removal of mono- and disaccharides would make the manufacturing process substantially more expensive. Therefore, GOS-Lu was tested without any previous purification to provide a more straightforward potential industrial application.
Rat number 3 in the control group died during the second DSS challenge, as this transient ulcerative colitis stage causes rectal bleeding. No significant differences on the body weight gain was observed between both animal cohorts during the 20 experimental weeks.
At week 20th, all 19 surviving animals were sacrificed and diverse histological measures were analyzed: caecum weight, number of colon tumors, and total tumor area in the colon mucosa (Figures 2, 3).
Caecum weight was significantly increased in animals from GOS-Lu cohort, an indication of the prebiotic properties of this preparation, as caecum is the main section of the digestive tract where prebiotic fermentative process occurs, being carried out by gut microbiota. Monosaccharides present in GOS-Lu preparation are absorbable carbohydrates at the small intestinal level, and they do not reach significant concentrations at colon level. Therefore, these free sugars (glucose and fructose) should not have any major impact in the outcome of these CRC animal model results. Regarding lactulose in the GOS-Lu preparation, it is well established that it undergoes fermentation rapidly in the proximal colon (Venema et al., 2003; Undseth et al., 2015). In consequence, the additional interest of GOS-Lu relies on the fact that it possesses lower fermentation rates as compared with lactulose because longer carbohydrate chains are normally fermented slower (Perrin et al., 2002), so their action takes place in the distal parts of the colon more efficiently (Díez-Municio et al., 2014). In this context, it is worth to mention a previous work demonstrating that GOS-Lu showed a better anti-inflammatory profile than lactulose in the trinitrobenzenesulfonic acid rat animal model of colitis, derived from the improvement of the luminal microbiota balance and a greater SCFA production (Algieri et al., 2014).
In the presence of prebiotics, some bacterial populations flourish in the caecum, contributing to a larger organ (Figure 2) (Roller et al., 2004b; Wang et al., 2010; Chen et al., 2011). GOS-Lu has been already described as prebiotic, and in this work, its daily intake of 2 g per rat has been able to clearly increase caecum weight (Moreno et al., 2014).
A factor which is directly correlated with the presence of prebiotics in the caecum is the bacterial fermentation of these compounds in this organ, giving rise to diverse SCFAs. In this study, a 56.9% increase in the caecum production of propionate was clearly observed, in a statistically significant way (Figure 4A). For the other detected SCFAs, these differences were not observed (Figure 4). These results will be commented below, after describing the changes in intestinal microbiota composition between both cohorts.
The number of colon tumors and the total tumor area in the colon mucosa clearly demonstrated a protective effect of GOS-Lu supplementation with respect to CRC. There was a statistically significant tumors number and area reduction (57.5 and 50.4, respectively) in the GOS-Lu cohort (Figure 3).
After analyzing these histological parameters, the intestinal microbiota composition was determined through metagenomics 16S ribosomal RNA (rRNA) sequencing of cecal content. The taxonomic diversity of a given metagenomics sample can be calculated using diverse alpha-diversity indexes. In this work, Shannon, Chao and Simpson indexes show that caecum bacterial richness in the animals belonging to GOS-Lu cohort is higher than in control cohort animals (Figure 6).
Analysis of gut microbiota at phylum level showed important differences between both cohorts (Table 1). More specifically, a high Firmicutes/Bacteroidetes ratio has been proposed as a microbiome marker for obesity and type II diabetes (two inflammatory and metabolic conditions) in human and animal studies (Ley et al., 2005, 2006). In this study, animals from GOS-Lu cohort showed increased Bacteroidetes and reduced Firmicutes populations, lowering significantly this ratio, a protective effect according to literature.
At the family level, the significant increase observed in the GOS-Lu cohort in Bacteroidetes phylum was mainly due to an increase in the abundance of Bacteroidaceae family (more specifically Bacteroides genus) and Prevotellaceae family (Paraprevotella genus), together with an increase in the Parabacteroides genus (Table 2 and Figures 7, 9). These three genera have been described as good propionate producers (Polansky et al., 2016). Phascolarctobacterium genus (which belongs to the Acidaminococcaceae family, a Firmicutes) is also a good propionate producer, and it is increased in GOS-Lu cohort as well. These three Bacteroidetes genera and one Firmicutes genus are the main responsible ones of the high increase in propionate production in the GOS-Lu cohort (Table 2 and Figures 9, 10A), which is a plausible explanation for the strong reduction in tumor numbers in this prebiotic cohort (Figure 3), as it has been described before for this SCFA (Ooi et al., 2010).
The significant reduction in Firmicutes phylum is due to lower population levels in members of the Lachnospiraceae family in general (Figures 7, 9). However, some genera of this family are slightly more abundant in the GOS-Lu cohort (Lachnoclostridium, Anaerostipes, Dorea, Blautia). This Lachnospiraceae family has been involved in high butyrate production (Polansky et al., 2016), and its reduction might be the main responsible for the reduced butyrate levels observed in the GOS-Lu cohort. One of the species showing an important reduction within Lachnospiraceae family is the mucolytic bacteria Ruminococcus gnavus, which has been reported to be more abundant in Crohn’s Disease (CD) and Intestinal Bowel Disease (IBD) patients (Png et al., 2010; Willing et al., 2010; Joossens et al., 2011) and may play an important role in inducing chronic intestinal inflammation (Eun et al., 2014). Eubacteriaceae family (from Firmicutes phylum) is also reduced in GOS-Lu cohort. This can have also a protective role against pro-inflammatory conditions, as an increase in this family has been described in dogs suffering IBD (Omori et al., 2017).
Three Firmicutes families are more abundant in the GOS-Lu cohort. These are Oscillospiraceae, Christensenellaceae, and Ruminococcaceae (Faecalibacterium, and Subdoligranulum genera) (Table 2). This is remarkable, as Faecalibacterium has been described as an anti-inflammatory genus, as it secretes metabolites able to block NF-kB activation, an important pro-inflammatory cellular mediator which is also involved in colon carcinogenesis (Sokol et al., 2006; Han et al., 2016; Huang et al., 2017). In a similar way, Christensenellaceae family has been associated with beneficial effects in the host, as lower obesity risk (Goodrich et al., 2014).
At the level of the Actinobacteria phylum, GOS-Lu cohort shows an increase in this bacteria, mainly due to more abundant populations of Bifidobacteriaceae family, and specially of Bifidobacterium genus, being associated to health benefits including protection against pro-inflammatory gut conditions but also against CRC (Table 1 and Figure 9). This protection against CRC has been associated to NF-kB repression by these probiotic bacteria (Roller et al., 2004a; Kim et al., 2010; Celiberto et al., 2017; Paveljšek et al., 2018).
Finally, regarding Proteobacteria phylum, Desulfovibrio genus (Desulfovibrionaceae family) populations are reduced in GOS-Lu cohort (Table 2). This bacteria have been associated to apoptosis induction in the colon mucosa in in vitro models and to a pro-inflammatory changes in UC patients, such as decreased sulphation of mucin, as a result of the metabolic activity of these bacteria (Lennon et al., 2014; Coutinho et al., 2017). Also, this bacterial genus has been associated to increasing levels of damage (also toward CRC) at the mucosal level, caused by reduction of the mucin barrier, the natural protection against luminal microbiota pro-inflammatory challenges (Song et al., 2018). However, another member of this family, Bilophila wadsworthia, is increased in GOS-Lu cohort. Although this species has been associated to a pro-inflammatory effect, some reports associate this mainly to the presence of low levels of the anti-inflammatory IL-10 (such as those found in IL-10-/- mice mutants colon mucosa) (Rennick et al., 1997; Devkota et al., 2012; Devkota and Chang, 2015). Therefore, in a wild type rat, as in this work, the pro-inflammatory effect of B. wadsworthia could be attenuated at different level in the gut mucosa.
In summary, this GOS-Lu supplementation experiment created a distinct distribution of main taxonomic families in gut microbiota, in contrast to feed control diet (Figure 10).
Based on these results, it can be noted that the addition of the prebiotic GOS-Lu to the animal diet was able to diminish colon tumors to a greater extent (57.5%, Figure 3), an effect that can be mainly attributed to an increase in propionate gut concentrations. This SCFA shows antitumor properties (induction of apoptosis in colon cancer cells) (Ooi et al., 2010), and its higher concentrations in the GOS-Lu cohort can be associated to increased populations of bacterial lineages described as good propionate producers, such as Bacteroides, Paraprevotella (both Bacteroidetes genera) and Phascolarctobacterium (a Firmicutes genus).
Also, the protective role of GOS-Lu supplementation can be associated with a reduction in gut pro-inflammatory bacterial populations, such as Desulfovibrio (a Proteobacteria genus), but above all, it can be associated with the greatest abundance of bacterial genera with strong anti-inflammatory and antitumor properties, as Bifidobacterium (an Actinobacteria genus) and Faecalibacterium (a Firmicutes genus).
Conclusion
Based on these results, GOS-Lu can be described as a prebiotic mixture with protective effects against CRC onset and progression in this animal model (57.5% reduction in colorectal tumors), which opens the way to its wide use in human populations as a food supplement. Also, GOS-Lu supplementation causes changes in intestinal microbiota, favoring anti-inflammatory taxons and reducing pro-inflammatory genera.
Author Contributions
AC, FL, and FM conceived and designed the experiments. JF and AO performed the experiments. JF, CV, and FL analyzed the data. FM and AO provided the GOS-Lu prebiotic. AC, JF, FL, FM, AO, and CV wrote the paper.
Funding
This work was funded by the research project grants RTC-2014-1525 from MINECO (Spanish Ministry of Economy, Industry and Competitiveness), AGL2017-84614-C2-1-R from MINECO, and AGR2011-7626 from Junta de Andalucía (Spain).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Servicios Científico-Técnicos from the University of Oviedo (Sequencing Unit) and from ISPA (Biostatistics and Epidemiology Platform).
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2018.02004/full#supplementary-material
TABLE S1 | Phyla, families and genera data for each of the animals used in this study.
TABLE S2 | Raw data on animals weight, caecums weight, number of Peyer’s patches, number of polyps, total tumoral area and short fatty acids concentrations for each of the animals used in this study.
Footnotes
References
Algieri, F., Rodríguez-Nogales, A., Garrido-Mesa, N., Vezza, T., Garrido-Mesa, J., Utrilla, M. P., et al. (2014). Intestinal anti-inflammatory Effects of Oligosaccharides derived from Lactulose in the Trinitrobenzenesulfonic acid model of rat colitis. J. Agric. Food Chem. 62, 4285–4297. doi: 10.1021/jf500678p
Arnold, M., Sierra, M. S., Laversanne, M., Soerjomataram, I., Jemal, A., and Bray, F. (2017). Global patterns and trends in colorectal cancer incidence and mortality. Gut 66, 683–691. doi: 10.1136/gutjnl-2015-310912
Bindels, L. B., Delzenne, N. M., Cani, P. D., and Walter, J. (2015). Towards a more comprehensive concept for prebiotics. Nat. Rev. Gastroenterol. Hepatol. 12, 303–310. doi: 10.1038/nrgastro.2015.47
Bosscher, D., Breynaert, A., Pieters, L., and Hermans, N. (2009). Food-based strategies to modulate the composition of the intestinal microbiota and their associated health effects. J. Physiol. Pharmacol. 60(Suppl. 6), 5–11.
Bray, F., Ren, J.-S., Masuyer, E., and Ferlay, J. (2013). Global estimates of cancer prevalence for 27 sites in the adult population in 2008. Int. J. cancer 132, 1133–1145. doi: 10.1002/ijc.27711
Brenner, H., Kloor, M., and Pox, C. P. (2014). Colorectal cancer. Lancet 383, 1490–1502. doi: 10.1016/S0140-6736(13)61649-9
Canfora, E. E., Jocken, J. W., and Blaak, E. E. (2015). Short-chain fatty acids in control of body weight and insulin sensitivity. Nat. Rev. Endocrinol. 11, 577–591. doi: 10.1038/nrendo.2015.128
Cardelle-Cobas, A., Corzo, N., Olano, A., Peláez, C., Requena, T., and Ávila, M. (2011). Galactooligosaccharides derived from lactose and lactulose: influence of structure on Lactobacillus, Streptococcus and Bifidobacterium growth. Int. J. Food Microbiol. 149, 81–87. doi: 10.1016/j.ijfoodmicro.2011.05.026
Cardelle-Cobas, A., Fernández, M., Salazar, N., Martínez-Villaluenga, C., Villamiel, M., Ruas-Madiedo, P., et al. (2009). Bifidogenic effect and stimulation of short chain fatty acid production in human faecal slurry cultures by oligosaccharides derived from lactose and lactulose. J. Dairy Res. 76, 317–325. doi: 10.1017/S0022029909004063
Cardelle-Cobas, A., Martínez-Villaluenga, C., Villamiel, M., Olano, A., and Corzo, N. (2008). Synthesis of oligosaccharides derived from lactulose and pectinex ultra SP-L. J. Agric. Food Chem. 56, 3328–3333. doi: 10.1021/jf073355b
Cardelle-Cobas, A., Olano, A., Corzo, N., Villamiel, M., Collins, M., Kolida, S., et al. (2012). in vitro fermentation of lactulose-derived oligosaccharides by mixed fecal microbiota. J. Agric. Food Chem. 60, 2024–2032. doi: 10.1021/jf203622d
Celiberto, L. S., Bedani, R., Dejani, N. N., Ivo de Medeiros, A., Sampaio Zuanon, J. A., Spolidorio, L. C., et al. (2017). Effect of a probiotic beverage consumption (Enterococcus faecium CRL 183 and Bifidobacterium longum ATCC 15707) in rats with chemically induced colitis. PLoS One 12:e0175935. doi: 10.1371/journal.pone.0175935
Chen, Q., Swist, E., Beckstead, J., Green, J., Matias, F., Roberts, J., et al. (2011). Dietary fructooligosaccharides and wheat bran elicit specific and dose-dependent gene expression profiles in the proximal colon epithelia of healthy Fischer 344 rats. J. Nutr. 141, 790–797. doi: 10.3945/jn.110.133421
Clemente, A. (2014). “In vivo assessment of the bioactivity of food oligosaccharides,” in Food Oligosaccharides: Production, Analysis and Bioactivity, eds F. J. Moreno and M. L. Sanz (Hoboken, NJ: John Wiley & Sons), 238–254. doi: 10.1002/9781118817360.ch14
Coutinho, C. M. L. M., Coutinho-Silva, R., Zinkevich, V., Pearce, C. B., Ojcius, D. M., and Beech, I. (2017). Sulphate-reducing bacteria from ulcerative colitis patients induce apoptosis of gastrointestinal epithelial cells. Microb. Pathog. 112, 126–134. doi: 10.1016/j.micpath.2017.09.054
Devkota, S., and Chang, E. B. (2015). Interactions between diet, bile acid metabolism, gut microbiota, and inflammatory bowel diseases. Dig. Dis. 33, 351–356. doi: 10.1159/000371687
Devkota, S., Wang, Y., Musch, M. W., Leone, V., Fehlner-Peach, H., Nadimpalli, A., et al. (2012). Dietary-fat-induced taurocholic acid promotes pathobiont expansion and colitis in Il10-/- mice. Nature 487, 104–108. doi: 10.1038/nature11225
Díez-Municio, M., Herrero, M., Olano, A., and Moreno, F. J. (2014). Synthesis of novel bioactive lactose-derived oligosaccharides by microbial glycoside hydrolases. Microb. Biotechnol. 7, 315–331. doi: 10.1111/1751-7915.12124
Eun, C. S., Mishima, Y., Wohlgemuth, S., Liu, B., Bower, M., Carroll, I. M., et al. (2014). Induction of bacterial antigen-specific colitis by a simplified human microbiota consortium in gnotobiotic interleukin-10-/- mice. Infect. Immun. 82, 2239–2246. doi: 10.1128/IAI.01513-13
Ferlay, J., Soerjomataram, I., Dikshit, R., Eser, S., Mathers, C., Rebelo, M., et al. (2015). Cancer incidence and mortality worldwide: sources, methods and major patterns in GLOBOCAN 2012. Int. J. cancer 136, E359–E386. doi: 10.1002/ijc.29210
Fernández, J., Redondo-Blanco, S., Miguélez, E. M., Villar, C. J., Clemente, A., and Lombó, F. (2015). Healthy effects of prebiotics and their metabolites against intestinal diseases and colorectal cancer. AIMS Microbiol. 1, 48–71. doi: 10.3934/microbiol.2015.1.48
Ferreira-Lazarte, A., Olano, A., Villamiel, M., and Moreno, F. J. (2017). Assessment of in vitro digestibility of dietary carbohydrates using rat small intestinal extract. J. Agric. Food Chem. 65, 8046–8053. doi: 10.1021/acs.jafc.7b01809
Gibson, G. R., Hutkins, R., Sanders, M. E., Prescott, S. L., Reimer, R. A., Salminen, S. J., et al. (2017). Expert consensus document: the International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 14, 491–502. doi: 10.1038/nrgastro.2017.75
Gibson, G. R., and Roberfroid, M. B. (1995). Dietary modulation of the human colonic microbiota: introducing the concept of prebiotics. J. Nutr. 125, 1401–1412.
Goodrich, J. K., Waters, J. L., Poole, A. C., Sutter, J. L., Koren, O., Blekhman, R., et al. (2014). Human genetics shape the gut microbiome. Cell 159, 789–799. doi: 10.1016/j.cell.2014.09.053
Han, Y.-M., Jeong, M., Park, J.-M., Kim, M.-Y., Go, E.-J., Cha, J. Y., et al. (2016). The ω-3 polyunsaturated fatty acids prevented colitis-associated carcinogenesis through blocking dissociation of β-catenin complex, inhibiting COX-2 through repressing NF-κB, and inducing 15-prostaglandin dehydrogenase. Oncotarget 7, 63583–63595. doi: 10.18632/oncotarget.11544
Hernández-Hernández, O., Marín-Manzano, M. C., Rubio, L. A., Moreno, F. J., Sanz, M. L., and Clemente, A. (2012). Monomer and linkage type of galacto-oligosaccharides affect their resistance to ileal digestion and prebiotic properties in rats. J. Nutr. 142, 1232–1239. doi: 10.3945/jn.111.155762
Huang, T.-H., Wu, S.-Y., Huang, Y.-J., Wei, P.-L., Wu, A. T., and Chao, T.-Y. (2017). The identification and validation of Trichosstatin A as a potential inhibitor of colon tumorigenesis and colon cancer stem-like cells. Am. J. Cancer Res. 7, 1227–1237.
Jemal, A., Bray, F., Center, M. M., Ferlay, J., Ward, E., and Forman, D. (2011). Global cancer statistics. CA Cancer J. Clin. 61, 69–90. doi: 10.3322/caac.20107
Joossens, M., Huys, G., Cnockaert, M., Preter, V., De Verbeke, K., Rutgeerts, P., et al. (2011). Dysbiosis of the faecal microbiota in patients with Crohn’s disease and their unaffected relatives. Gut 60, 631–637. doi: 10.1136/gut.2010.223263
Kim, S. W., Kim, H. M., Yang, K. M., Kim, S.-A., Kim, S.-K., An, M. J., et al. (2010). Bifidobacterium lactis inhibits NF-kappaB in intestinal epithelial cells and prevents acute colitis and colitis-associated colon cancer in mice. Inflamm. Bowel Dis. 16, 1514–1525. doi: 10.1002/ibd.21262
Kim, Y. S., and Milner, J. A. (2007). Dietary modulation of colon cancer risk. J. Nutr. 137, 2576S–2579S. doi: 10.1093/jn/137.11.2576S
Koh, A., De Vadder, F., Kovatcheva-Datchary, P., and Bäckhed, F. (2016). From dietary fiber to host physiology: short-chain fatty acids as key bacterial metabolites. Cell 165, 1332–1345. doi: 10.1016/j.cell.2016.05.041
Laparra, J. M., Díez-Municio, M., Herrero, M., and Moreno, F. J. (2014). Structural differences of prebiotic oligosaccharides influence their capability to enhance iron absorption in deficient rats. Food Funct. 5, 2430–2437. doi: 10.1039/c4fo00504j
Laparra, J. M., Hernandez-Hernandez, O., Moreno, F. J., and Sanz, Y. (2013). Neoglycoconjugates of caseinomacropeptide and galactooligosaccharides modify adhesion of intestinal pathogens and inflammatory response(s) of intestinal (Caco-2) cells. Food Res. Int. 54, 1096–1102. doi: 10.1016/j.foodres.2012.10.034
Lennon, G., Balfe,Á, Bambury, N., Lavelle, A., Maguire, A., Docherty, N. G., et al. (2014). Correlations between colonic crypt mucin chemotype, inflammatory grade and Desulfovibrio species in ulcerative colitis. Colorectal Dis. 16,O161–O169. doi: 10.1111/codi.12503
Ley, R. E., Bäckhed, F., Turnbaugh, P., Lozupone, C. A., Knight, R. D., and Gordon, J. I. (2005). Obesity alters gut microbial ecology. Proc. Natl. Acad. Sci. U.S.A. 102, 11070–11075. doi: 10.1073/pnas.0504978102
Ley, R. E., Turnbaugh, P. J., Klein, S., and Gordon, J. I. (2006). Microbial ecology: human gut microbes associated with obesity. Nature 444, 1022–1023. doi: 10.1038/4441022a
López-Sanz, S., Montilla, A., Moreno, F. J., and Villamiel, M. (2015). Stability of oligosaccharides derived from lactulose during the processing of milk and apple juice. Food Chem. 183, 64–71. doi: 10.1016/j.foodchem.2015.03.020
Louis, P., Hold, G. L., and Flint, H. J. (2014). The gut microbiota, bacterial metabolites and colorectal cancer. Nat. Rev. Microbiol. 12, 661–672. doi: 10.1038/nrmicro3344
Marín-Manzano, M. C., Abecia, L., Hernández-Hernández, O., Sanz, M. L., Montilla, A., Olano, A., et al. (2013). Galacto-oligosaccharides derived from lactulose exert a selective stimulation on the growth of Bifidobacterium animalis in the large intestine of growing rats. J. Agric. Food Chem. 61, 7560–7567. doi: 10.1021/jf402218z
Martínez-Villaluenga, C., Cardelle-Cobas, A., Olano, A., Corzo, N., Villamiel, M., and Jimeno, M. L. (2008). Enzymatic synthesis and identification of two trisaccharides produced from lactulose by transgalactosylation. J. Agric. Food Chem. 56, 557–563. doi: 10.1021/jf0721343
Merrill, R. M., and Anderson, A. E. (2011). Risk-adjusted colon and rectal cancer incidence rates in the United States. Dis. Colon Rectum 54, 1301–1306. doi: 10.1097/DCR.0b013e3182242bd3
Moreno, F. J., Montilla, A., Villamiel, M., Corzo, N., and Olano, A. (2014). Analysis, structural characterization, and bioactivity of oligosaccharides derived from lactose. Electrophoresis 35, 1519–1534. doi: 10.1002/elps.201300567
Omori, M., Maeda, S., Igarashi, H., Ohno, K., Sakai, K., Yonezawa, T., et al. (2017). Fecal microbiome in dogs with inflammatory bowel disease and intestinal lymphoma. J. Vet. Med. Sci. 79, 1840–1847. doi: 10.1292/jvms.17-0045
Ooi, C. C., Good, N. M., Williams, D. B., Lewanowitsch, T., Cosgrove, L. J., Lockett, T. J., et al. (2010). Efficacy of butyrate analogues in HT-29 cancer cells. Clin. Exp. Pharmacol. Physiol. 37, 482–489. doi: 10.1111/j.1440-1681.2009.05335.x
Paveljšek, D., Juvan, P., Košir, R., Rozman, D., Hacin, B., Ivičak-Kocjan, K., et al. (2018). Lactobacillus fermentum L930BB and Bifidobacterium animalis subsp. animalis IM386 initiate signalling pathways involved in intestinal epithelial barrier protection. Benef. Microbes 9, 515–525. doi: 10.3920/BM2017.0107
Perrin, S., Fougnies, C., Grill, J. P., Jacobs, H., and Schneider, F. (2002). Fermentation of chicory fructo-oligosaccharides in mixtures of different degrees of polymerization by three strains of bifidobacteria. Can. J. Microbiol. 48, 759–763. doi: 10.1139/w02-065
Png, C. W., Lindén, S. K., Gilshenan, K. S., Zoetendal, E. G., McSweeney, C. S., Sly, L. I., et al. (2010). Mucolytic bacteria with increased prevalence in IBD mucosa augment in vitro utilization of mucin by other bacteria. Am. J. Gastroenterol. 105, 2420–2428. doi: 10.1038/ajg.2010.281
Polansky, O., Sekelova, Z., Faldynova, M., Sebkova, A., Sisak, F., and Rychlik, I. (2016). Important metabolic pathways and biological processes expressed by chicken cecal microbiota. Appl. Environ. Microbiol. 82, 1569–1576. doi: 10.1128/AEM.03473-15
Pompei, A., Cordisco, L., Raimondi, S., Amaretti, A., Pagnoni, U. M., Matteuzzi, D., et al. (2008). in vitro comparison of the prebiotic effects of two inulin-type fructans. Anaerobe 14, 280–286. doi: 10.1016/j.anaerobe.2008.07.002
Pool-Zobel, B. L. (2005). Inulin-type fructans and reduction in colon cancer risk: review of experimental and human data. Br. J. Nutr. 93(Suppl. 1), S73–S90. doi: 10.1079/BJN20041349
Rastall, R. A. (2013). Gluco and galacto-oligosaccharides in food: update on health effects and relevance in healthy nutrition. Curr. Opin. Clin. Nutr. Metab. Care 16, 675–678. doi: 10.1097/MCO.0b013e328365b5fd
Rennick, D. M., Fort, M. M., and Davidson, N. J. (1997). Studies with IL-10-/- mice: an overview. J. Leukoc. Biol. 61, 389–396. doi: 10.1002/jlb.61.4.389
Roberfroid, M. B. (2007). Inulin-type fructans: functional food ingredients. J. Nutr. 137, 2493S–2502S. doi: 10.1093/jn/137.11.2493S
Roller, M., Pietro Femia, A., Caderni, G., Rechkemmer, G., and Watzl, B. (2004a). Intestinal immunity of rats with colon cancer is modulated by oligofructose-enriched inulin combined with Lactobacillus rhamnosus and Bifidobacterium lactis. Br. J. Nutr. 92, 931–938.
Roller, M., Rechkemmer, G., and Watzl, B. (2004b). Prebiotic inulin enriched with oligofructose in combination with the probiotics Lactobacillus rhamnosus and Bifidobacterium lactis modulates intestinal immune functions in rats. J. Nutr. 134, 153–156.
Rumessen, J. J., Bodé, S., Hamberg, O., and Gudmand-Høyer, E. (1990). Fructans of Jerusalem artichokes: intestinal transport, absorption, fermentation, and influence on blood glucose, insulin, and C-peptide responses in healthy subjects. Am. J. Clin. Nutr. 52, 675–681. doi: 10.1093/ajcn/52.4.675
Scharlau, D., Borowicki, A., Habermann, N., Hofmann, T., Klenow, S., Miene, C., et al. (2009). Mechanisms of primary cancer prevention by butyrate and other products formed during gut flora-mediated fermentation of dietary fibre. Mutat. Res. 682, 39–53. doi: 10.1016/j.mrrev.2009.04.001
Sokol, H., Seksik, P., Rigottier-Gois, L., Lay, C., Lepage, P., Podglajen, I., et al. (2006). Specificities of the fecal microbiota in inflammatory bowel disease. Inflamm. Bowel Dis. 12, 106–111. doi: 10.1097/01.MIB.0000200323.38139.c6
Song, H., Wang, W., Shen, B., Jia, H., Hou, Z., Chen, P., et al. (2018). Pretreatment with probiotic Bifico ameliorates colitis-associated cancer in mice: transcriptome and gut flora profiling. Cancer Sci. 109, 666–677. doi: 10.1111/cas.13497
Stein, K., Borowicki, A., Scharlau, D., Schettler, A., Scheu, K., Obst, U., et al. (2012). Effects of synbiotic fermentation products on primary chemoprevention in human colon cells. J. Nutr. Biochem. 23, 777–784. doi: 10.1016/j.jnutbio.2011.03.022
Undseth, R., Jakobsdottir, G., Nyman, M., Berstad, A., and Valeur, J. (2015). Low serum levels of short-chain fatty acids after lactulose ingestion may indicate impaired colonic fermentation in patients with irritable bowel syndrome. Clin. Exp. Gastroenterol. 8, 303–308. doi: 10.2147/CEG.S94084
Venema, K., van Nuenen, M. H. M. C., van den Heuvel, E. G., Pool, W., and van der Vossen, J. M. B. M. (2003). The Effect of Lactulose on the composition of the intestinal microbiota and short-chain fatty acid production in human volunteers and a computer-controlled model of the proximal large intestine. Microb. Ecol. Health Dis. 15, 94–105. doi: 10.1080/08910600310019895
Verghese, M., Walker, L. T., Shackelford, L., and Chawan, C. B. (2005). Inhibitory effects of nondigestible carbohydrates of different chain lengths on azoxymethane-induced aberrant crypt foci in Fisher 344 rats. Nutr. Res. 25, 859–868. doi: 10.1016/j.nutres.2005.09.007
Wang, Y., Zeng, T., Wang, S., Wang, W., Wang, Q., and Yu, H.-X. (2010). Fructo-oligosaccharides enhance the mineral absorption and counteract the adverse effects of phytic acid in mice. Nutrition 26, 305–311. doi: 10.1016/j.nut.2009.04.014
Willing, B. P., Dicksved, J., Halfvarson, J., Andersson, A. F., Lucio, M., Zheng, Z., et al. (2010). A pyrosequencing study in twins shows that gastrointestinal microbial profiles vary with inflammatory bowel disease phenotypes. Gastroenterology 139, 1844.e1–1854.e1. doi: 10.1053/j.gastro.2010.08.049
Keywords: prebiotic, colorectal cancer, prevention, galacto-oligosaccharides, gut microbiota
Citation: Fernández J, Moreno FJ, Olano A, Clemente A, Villar CJ and Lombó F (2018) A Galacto-Oligosaccharides Preparation Derived From Lactulose Protects Against Colorectal Cancer Development in an Animal Model. Front. Microbiol. 9:2004. doi: 10.3389/fmicb.2018.02004
Received: 03 May 2018; Accepted: 08 August 2018;
Published: 31 August 2018.
Edited by:
Andrea Gomez-Zavaglia, Centro de Investigación y Desarrollo en Criotecnología de Alimentos (CIDCA), ArgentinaReviewed by:
Carlos Vera Vera, Universidad de Santiago de Chile, ChileJose M. Bruno-Barcena, North Carolina State University, United States
Copyright © 2018 Fernández, Moreno, Olano, Clemente, Villar and Lombó. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Felipe Lombó, lombofelipe@uniovi.es