Evidence for the first multi-species shark nursery area in Atlantic Africa (Boa Vista Island, Cabo Verde)
 Rui Rosa1,2,3*
Rui Rosa1,2,3*  Emanuel Nunes1,4
Emanuel Nunes1,4  Vasco Pissarra1,3
Vasco Pissarra1,3  Catarina Pereira Santos1,3,5
Catarina Pereira Santos1,3,5  Jaquelino Varela1,3
Jaquelino Varela1,3  Miguel Baptista1
Miguel Baptista1  Joana Castro1,6
Joana Castro1,6  José Ricardo Paula1,2 Tiago Repolho1,2
José Ricardo Paula1,2 Tiago Repolho1,2  Tiago A. Marques2,7,8
Tiago A. Marques2,7,8  Rui Freitas9
Rui Freitas9  Catarina Frazão Santos1,2,3,5
Catarina Frazão Santos1,2,3,5- 1MARE – Marine and Environmental Sciences Centre/ARNET – Aquatic Research Network, Laboratório Marítimo da Guia, Faculdade de Ciências, Universidade de Lisboa, Cascais, Portugal
- 2Departamento de Biologia Animal, Faculdade de Ciências, Universidade de Lisboa, Lisboa, Portugal
- 3Sphyrna Association, Boa Vista Island, Sal Rei, Cape Verde
- 4Inspeção Geral das Pescas, Ministério do Mar, Sal Rei, Boa Vista, Cape Verde
- 5Environmental Economics Knowledge Center, Nova School of Business and Economics, New University of Lisbon, Carcavelos, Portugal
- 6AIMM – Associação para a Investigação do Meio Marinho, Lisboa, Portugal
- 7Centro de Estatística e Aplicações, Universidade de Lisboa, Lisboa, Portugal
- 8Centre for Research into Ecological and Environmental Modelling, University of St Andrews, St Andrews, Scotland
- 9Instituto de Engenharia e Ciências do Mar, Universidade Técnica do Atlântico, Mindelo, São Vicente, Cape Verde
This study describes the first potential multi-species shark nursery area in Atlantic Africa (Sal Rei Bay – SRB, Boa Vista Island, Cabo Verde). From August 2016 to September 2019, 6162 neonates and juveniles of 5 different shark species were observed in SRB using beach gillnet-based bycatch surveys, namely milk (Rhizoprionodon acutus; n= 4908), scalloped hammerhead (Sphyrna lewini; n= 1035), blacktip (Carcharhinus limbatus; n=115), Atlantic weasel (Paragaleus pectoralis; n= 93) and nurse (Ginglymostoma cirratum; n= 12) sharks. Except for nurse sharks, significant seasonal variations in shark relative abundance were observed, with higher levels being recorded during summer and autumn. These findings, together with local knowledge (interviews to fishermen), denote the consistent use of SRB by juvenile sharks and its preference relative to other areas in the region. Ensuring the protection and conservation of SRB nursery area is especially relevant as, according to IUCN, all identified shark species are threatened with extinction over the near-future – in particular, scalloped hammerheads (critically endangered) and Atlantic weasel sharks (endangered). The effective protection of SRB will not only support the conservation of shark populations, but also of other charismatic fauna (e.g., loggerhead turtles) and broader benthic and pelagic ecosystems.
Introduction
Most sharks occupy high trophic levels in marine ecosystems, thus exerting a key influence on their structure and function (1990; Compagno, 1984). Yet, contrary to most fishes, sharks generally have a K-selected life history strategy, which means slow growth rates, late maturity age, low fecundity, long gestation period, few offspring, and long-life spans (Dulvy et al., 2014; Wheeler et al., 2020). This, allied to a general tendency for segregation by age and sex, makes them especially vulnerable to human impacts (Baum et al., 2003; García et al., 2008; Ferretti et al., 2010; Roff et al., 2016). In fact, shark populations have been plummeting over the past few decades, namely driven by intense fishing pressure, with key implications for their sustainability and conservation (Queiroz et al., 2019; Dulvy et al., 2021; Pacoureau et al., 2021).
Understanding the habitat-use patterns of sharks, namely through identification of key aggregation sites and nursery grounds, is essential to recognize the potential effects of human activities on these populations and design effective conservation and management strategies (Knipp et al., 2010; Speed et al., 2010; Diemer et al., 2011; Henderson et al., 2016; Queiroz et al., 2016; Heupel et al., 2019; Queiroz et al., 2019). Among several hypotheses concerning the role of nursery areas, it is generally accepted that such areas provide enhanced food availability and protection against predation (Springer, 1967; Branstetter, 1987; Heupel and Simpfendorfer, 2002; Heupel et al., 2007). According to Heupel et al. (2007), three criteria must be met so that a particular marine area can be considered as a shark nursery ground, namely: i) preference – sharks are found more often in the specific area than in neighboring ones, ii) residency – sharks tend to remain in the area (or return) for extended periods, and iii) consistency – the area or habitat is used repeatedly by sharks over the years.
While great efforts have been made to identify and describe sharks’ nursery areas around the world, there are still strong climate, habitat, and taxonomic bias in the literature (Heupel et al., 2019). Moreover, and because the identification of such important nursery areas is often dependent on long-term sample size datasets, most studies do not comply with all three criteria defined by Heupel et al. (2007), fulfilling only one or two criteria. For the Atlantic African region in particular, seven potential shark nursery areas were described over the past decade, all of them being for single species (see Supplementary Table 1). These pertained to areas used by angel sharks (Squatina squatina) in the Canary Islands, leafscale gulper sharks (Centrophorus squamosus) in Mauritania and Namibia, and great white (Carcharodon carcharias), smooth hound (Mustelus mustelus), and blue (Prionace glauca) sharks in South Africa (see details and respective references in Supplementary Table 1).
Communal nurseries are locations where juveniles of multiple shark species occur and the adults are mostly absent (Simpfendorfer and Milward, 1993). Yet, in such nurseries, the juveniles face a tradeoff between lower predation risk and increased competition – while the latter is potentially reduced via partitioning of food resources (Kinney et al., 2011). Within this context, here we describe, for the first time, a potential multi-species/communal shark nursery area in the Atlantic African region. More specifically, we describe the first potential nursery of milk (Rhizoprionodon acutus), scalloped hammerhead (Sphyrna lewini), blacktip (Carcharhinus limbatus), Atlantic weasel (Paragaleus pectoralis), and nurse (Ginglymostoma cirratum) sharks in Sal Rei Bay (SRB), Boa Vista Island, Cabo Verde. Here we assess: (i) the diversity of shark species occurring in the SRB; (ii) the size frequency distribution of juvenile sharks; (iii) inter and intra-year patterns in the relative abundance of juvenile sharks (catch per unit of effort data); and (iv) the spatial variation in species composition and abundance around Boa Vista Island based on interviews to local fishermen.
Material and methods
Temporal changes in juvenile shark relative abundance in SRB, Boa Vista Island (Cabo Verde)
Cabo Verde is a small archipelagic country, located in the Atlantic Ocean (Supplementary Figure 1), which has been long recognized as a global hotspot of marine biodiversity (Roberts et al., 2002; Freitas et al., 2019). Boa Vista Island is the easternmost (windward) island of the archipelago (Supplementary Figure 1), with a coastline mostly composed of sandy and rocky beaches and high-energy exposed shores (Gomes, 2019). While Boa Vista is a well-known nesting area for loggerhead turtles (Caretta caretta) in the eastern Atlantic (Marco et al., 2012; Martins et al., 2022), and breeding area for the endangered North Atlantic humpback whale (Megaptera novaeangliae) (Wenzel et al., 2020), biological knowledge on other marine taxa, namely on sharks, is largely absent. A particular bay in the island, the SRB (Supplementary Figure 2), comprises a marine area of c. 22 km2, mostly of sandy substrate, and is locally known (e.g., local communities, artisanal fishermen) to bear a variety of shark juveniles. Therefore, from August 2016 to September 2019, in SRB, and with the help of local fishermen, beach gillnet-based bycatch was surveyed (4 cm square-mesh monofilament gill net, with 30 meters in length and 3 m deep) on a monthly basis. This type of artisanal fishing gear is used by the local fishermen to catch small pelagic fish (sparids, bigeye scad, tuna, among others). The gill net was always set perpendicular to the shore, with soak time ranging from 2 to 4 h, depending on tidal and weather (season) conditions. Juvenile sharks were identified to the species-level. To prevent post-release casualties, the net was regularly surveyed, and when necessary, the animals were manually moved through the water during release to promote recuperation. Catch per unit effort (CPUE) was calculated as the number of sharks caught per hour per net meter square (sharks h-1 m-2).
Spatial differences in juvenile shark abundance
To understand if sharks are found more often in SBR than in other areas of the island, a short questionnaire was conducted close to local artisanal and semi-industrial fishermen. From a list of 92 licensed fishermen in Boa Vista Island, a total of 55 interviews were done (~60%). They were conducted in Cabo Verdean creole, by telephone, in August 2022. Prior to the interviews, fishermen were informed about the purpose of the survey, anonymity, and confidential treatment of the obtained data, and asked for verbal consent to participating. The interview entailed the following 4 questions:
i) “Are you aware of any specific areas in Boa Vista Island where one can find higher abundance of shark juveniles?”
ii) “Among those areas, which one shows the highest values of abundance of shark juveniles”?
iii) “Which species do you find in that particular area”?
and
iv) “Where do you usually fish”?
Total length distributions
Total length (TL) of juvenile sharks was measured to the nearest 0.5 cm. TL frequency distributions of milk (n= 2165), scalloped hammerheads (n= 404), blacktip (n=115), Atlantic weasel (n=94) and nurse sharks (n=12) were evaluated and compared with key biological information obtained from previous studies, namely species’ length at first maturity and size at birth in the Atlantic Ocean (see respective data and references in Supplementary Table 2).
Statistical analyses
To evaluate changes over time generalized additive models (GAMs) per species were used, with a smooth of time (year) and a cyclic smooth of month. We considered the relative abundance (catch per unit of effort) response to be Gaussian, with a log link. To ensure that the cyclic nature of the variable month was respected we considered a cyclic basis for the month spline. Models were implemented in the R library mgcv, following Wood (2017). The residuals of the fitted models were checked for temporal autocorrelation, and since no serious reasons for concern were found, we did not include an autocorrelation term in the models.
Results
Temporal changes in juvenile shark relative abundance in SRB
From August 2016 to September 2019, juveniles of five different shark species were observed during bycatch surveys in SRB, namely milk (n=4908), scalloped hammerhead (n=1035), blacktip (n=115), Atlantic weasel (n=93) and nurse (n=12) sharks (Figures 1, 2). The higher CPUE values were observed for the milk shark (reaching a maximum of 3.55 individuals h-1 m-2 in August 2017), followed by the hammerhead (a maximum of 0.52 individuals h-1 m-2 in September 2019), blacktip (a maximum of 0.06 individuals h-1 m-2 in November 2017), Atlantic weasel (a maximum of 0.03 individuals h-1 m-2 in June 2017), and nurse sharks (a maximum of 0.01 individuals h-1 m-2 in January 2019). Except for the nurse sharks, all other species revealed significant seasonal variations in CPUE values (Table 1), with highest values being observed during summer or autumn periods (Figures 1, 2).
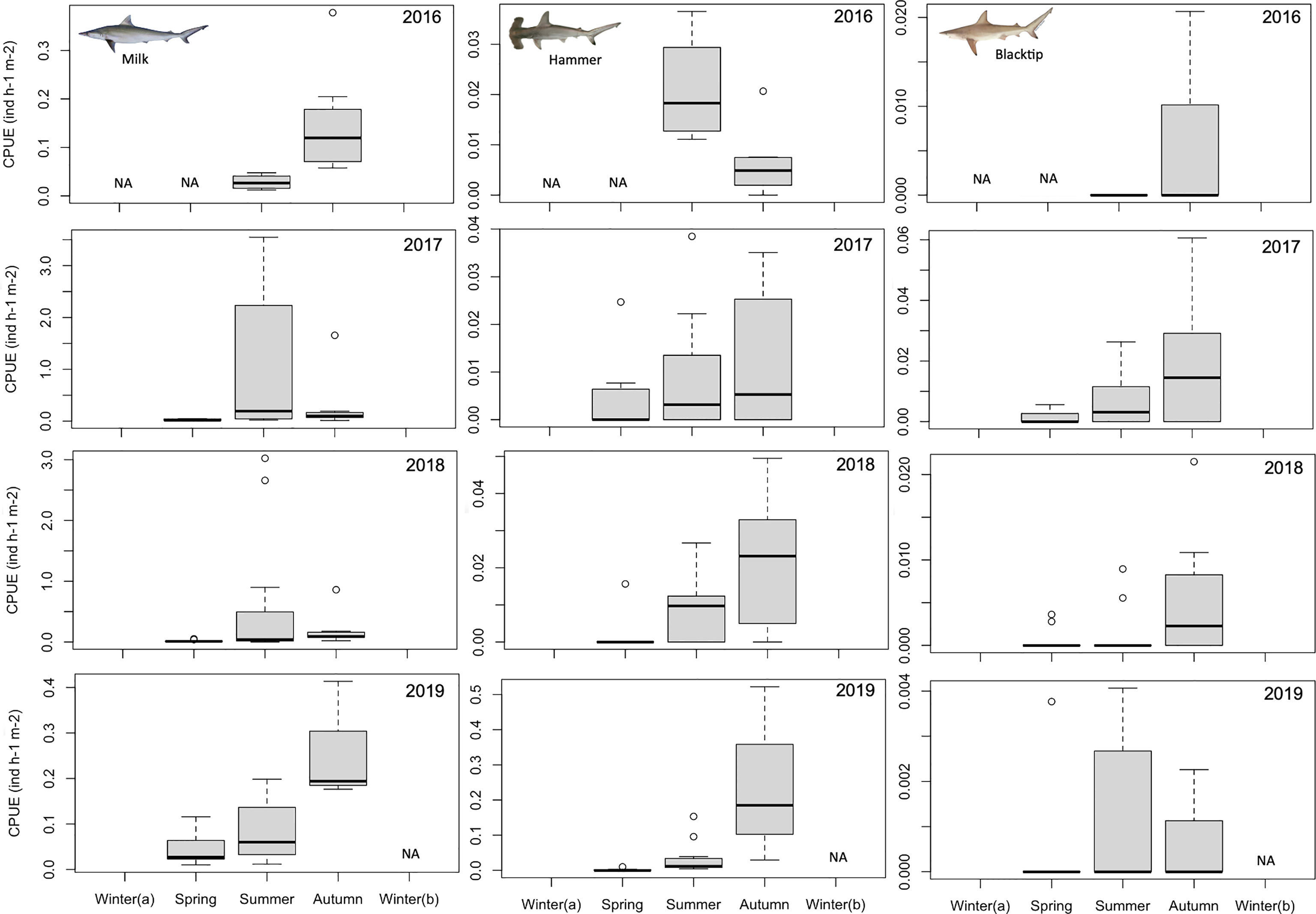
Figure 1 Temporal changes in the relative abundance (catch per unit of effort; number of individuals h-1 m-2) of juvenile milk (Rhizoprionodon acutus), scalloped hammerhead (Sphyrna lewini), and blacktip (Carcharhinus limbatus) sharks in Sal Rei Bay, Boa Vista Island, Cabo Verde. NA, not applicable.
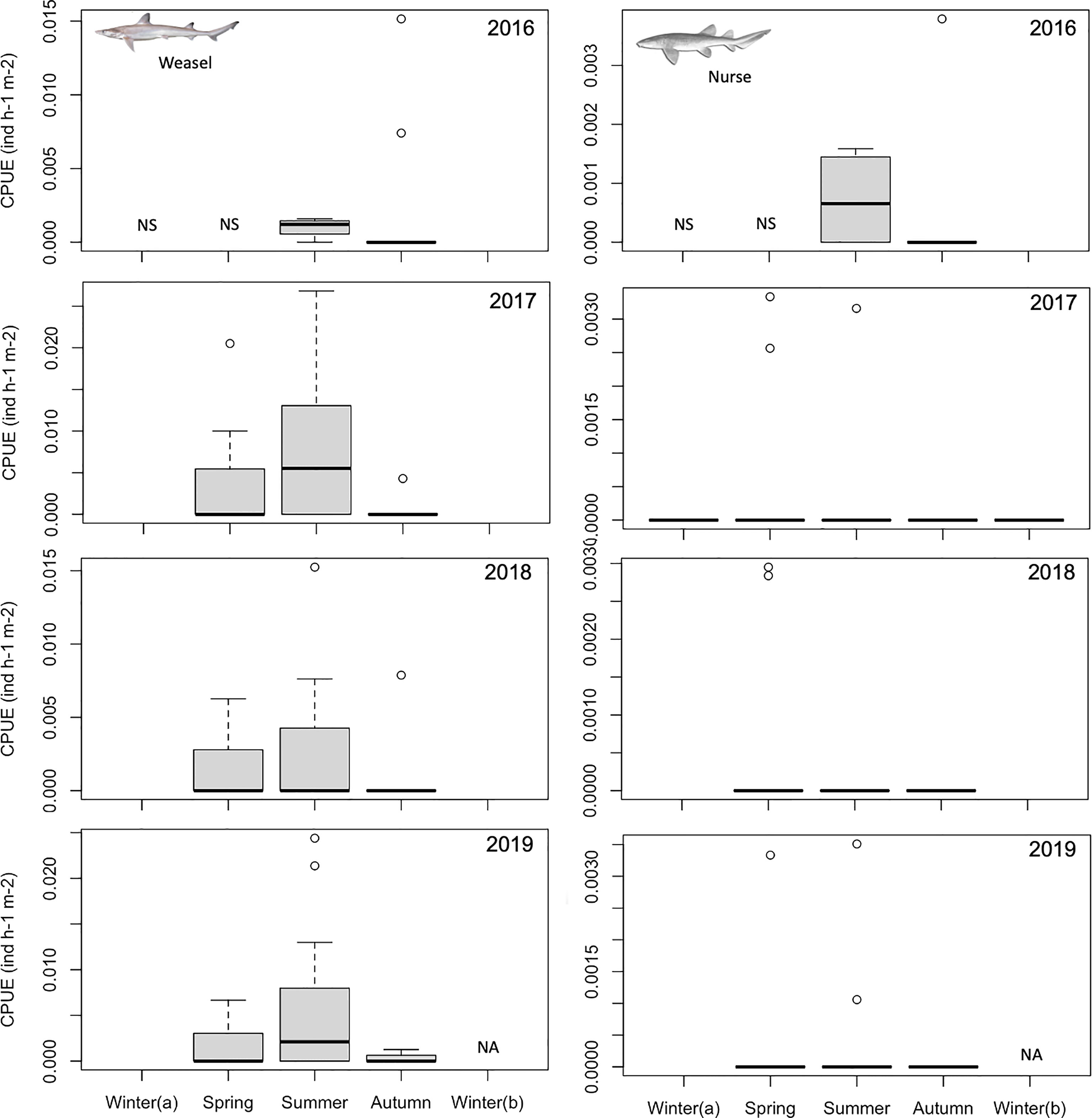
Figure 2 Temporal changes in the relative abundance (catch per unit of effort; number of individuals h-1 m-2) of juvenile Atlantic weasel (Paragaleus pectoralis) and nurse (Ginglymostoma cirratum) juvenile sharks in Sal Rei Bay, Boa Vista Island, Cabo Verde. NS, not sampled.
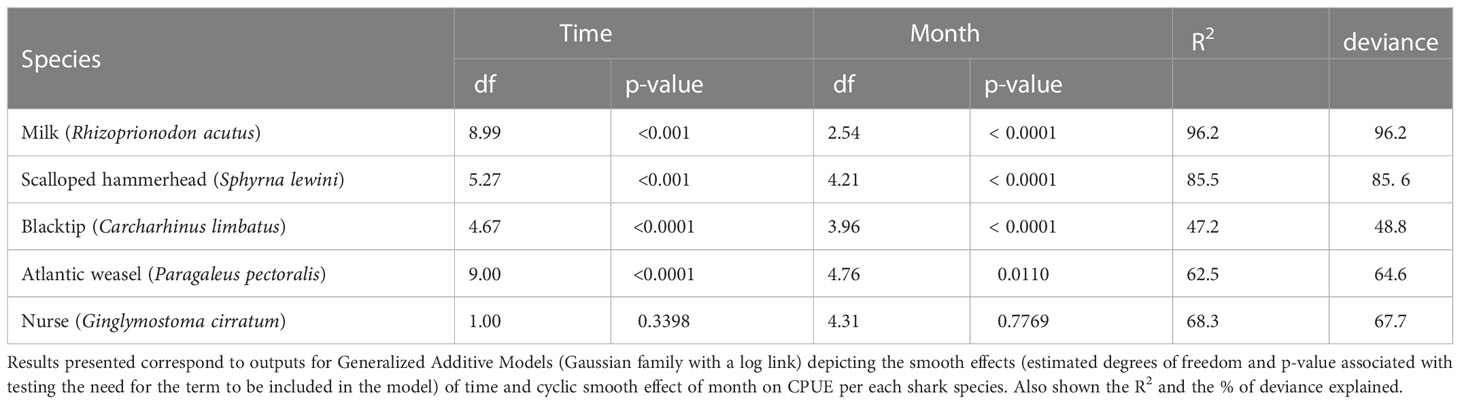
Table 1 Effects of time (year) and cyclic month effects on the catch per unit of effort (CPUE) for each of the five shark species.
Total length distributions
The size range of milk sharks (n=2165) was 30 to 70 cm total length (mode 40-50 cm interval), while for the scalloped hammerheads (n=404) was 30 to 65 cm (mode 50-60 cm interval). The blacktip sharks (n=115) presented a size range of 58 to 110 cm (mode 70-80 cm interval), the Atlantic weasel sharks (n=94) of 43 to 98 cm (mode 50-60 cm interval), and the nurse sharks (n=12) of 43 to 140 cm (mode 50-60 cm interval). All sampled individuals revealed sizes below species’ length at first maturity, except to two weasel shark individuals (with 96 and 98 cm total length; Figure 3).
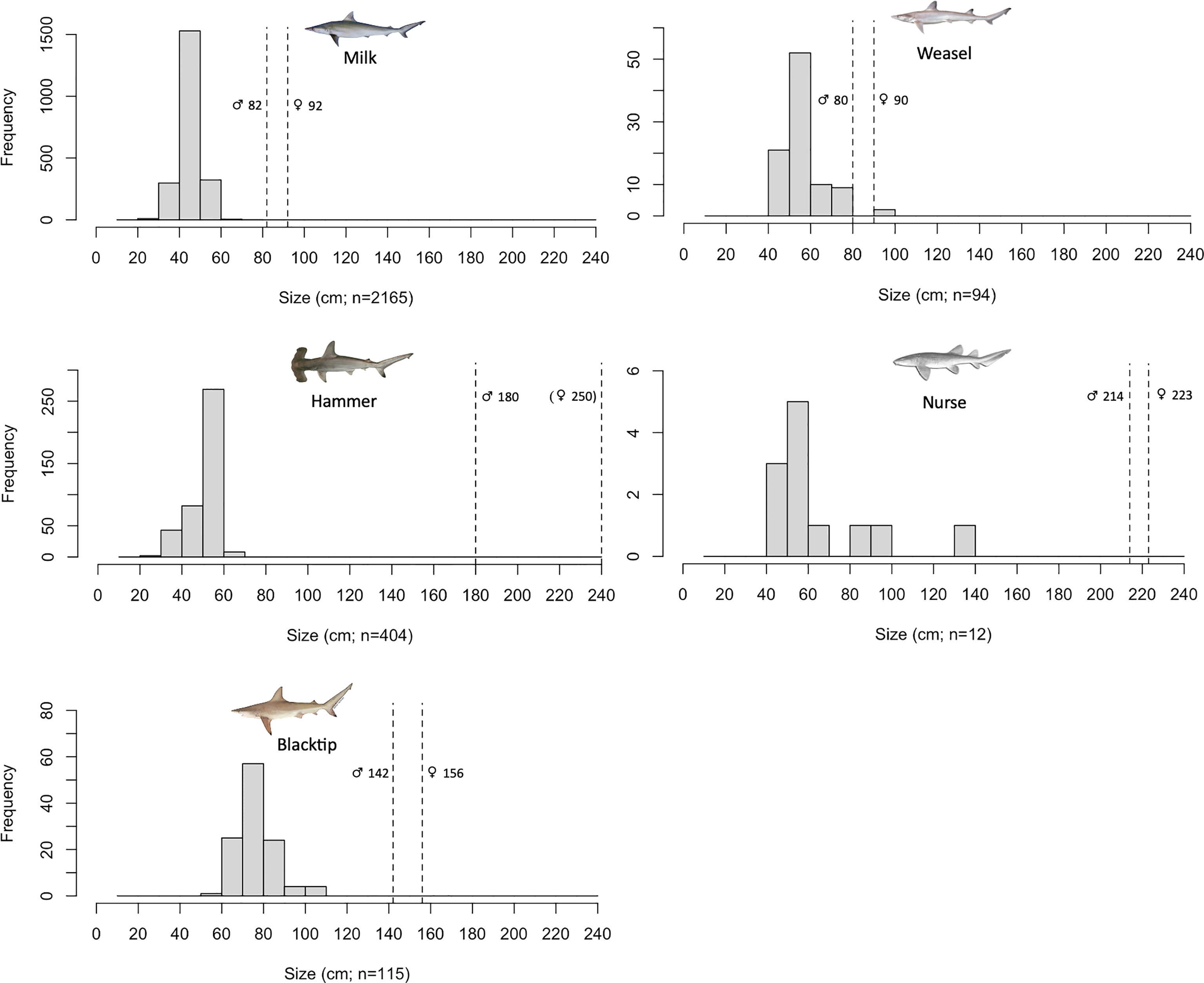
Figure 3 Length distributions of juvenile milk (Rhizoprionodon acutus), scalloped hammerhead (Sphyrna lewini), blacktip (Carcharhinus limbatus), Atlantic weasel (Paragaleus pectoralis) and nurse (Ginglymostoma cirratum) sharks in Sal Rei Bay, Boa Vista Island, Cabo Verde. Vertical dashed lines represent species’ length at first maturity for both males and females (see respective references in Supplementary Table 2).
Spatial differences in juvenile shark abundance
Local fishermen identified 11 areas of occurrence of juvenile sharks around the island. Yet, SRB collected the highest level of agreement by far, with 78% of respondents identifying it as an area of juvenile sharks’ occurrence, and 60% as the area with the highest number of juvenile sharks in the entire island (Figures 4A, B). Some areas within the SRB were further emphasized by fishermen, such as “Djeu”, “Praia do Estoril”, “Morro de Areia”, “Praia de Chaves”, or “Caramboa” (n=14). Ervatão and Santa Mónica were also identified as areas of juvenile sharks’ occurrence (24% and 22%, respectively), and Ervatão and Esgata as having the highest number of juvenile sharks, only to a lower extent (11% and 7%, respectively; Figures 4A, B). When asked about what species were present in the area with the highest number of juvenile sharks, 16% of respondents identified blacktip sharks (Figure 4C), 20% identified hammerhead sharks (Figure 4D), and 60% identified “cação” – the latter is the common name used locally to refer several species, including milk and Atlantic weasel sharks (Figure 4E). In all cases, SRB was the area that collected most responses for each species (from 64% to 92%; Figures 4C–E). This preference was irrespective from respondents fishing grounds, as 80% of the fishermen that selected SRB do not use it as a fishing ground (Supplementary Figure 3). Only a very small percentage of respondents did not provide any information on shark juveniles (2 fishermen were not knowledgeable on areas with juvenile sharks, and 1 did not want to share information). Most fishermen identified only one type of shark (n=45, 82%; Supplementary Figure 4). Only a small number of fishermen identified two types of sharks simultaneously (n=4) or was not able to identify any particular species (n=3; Supplementary Figure 4).
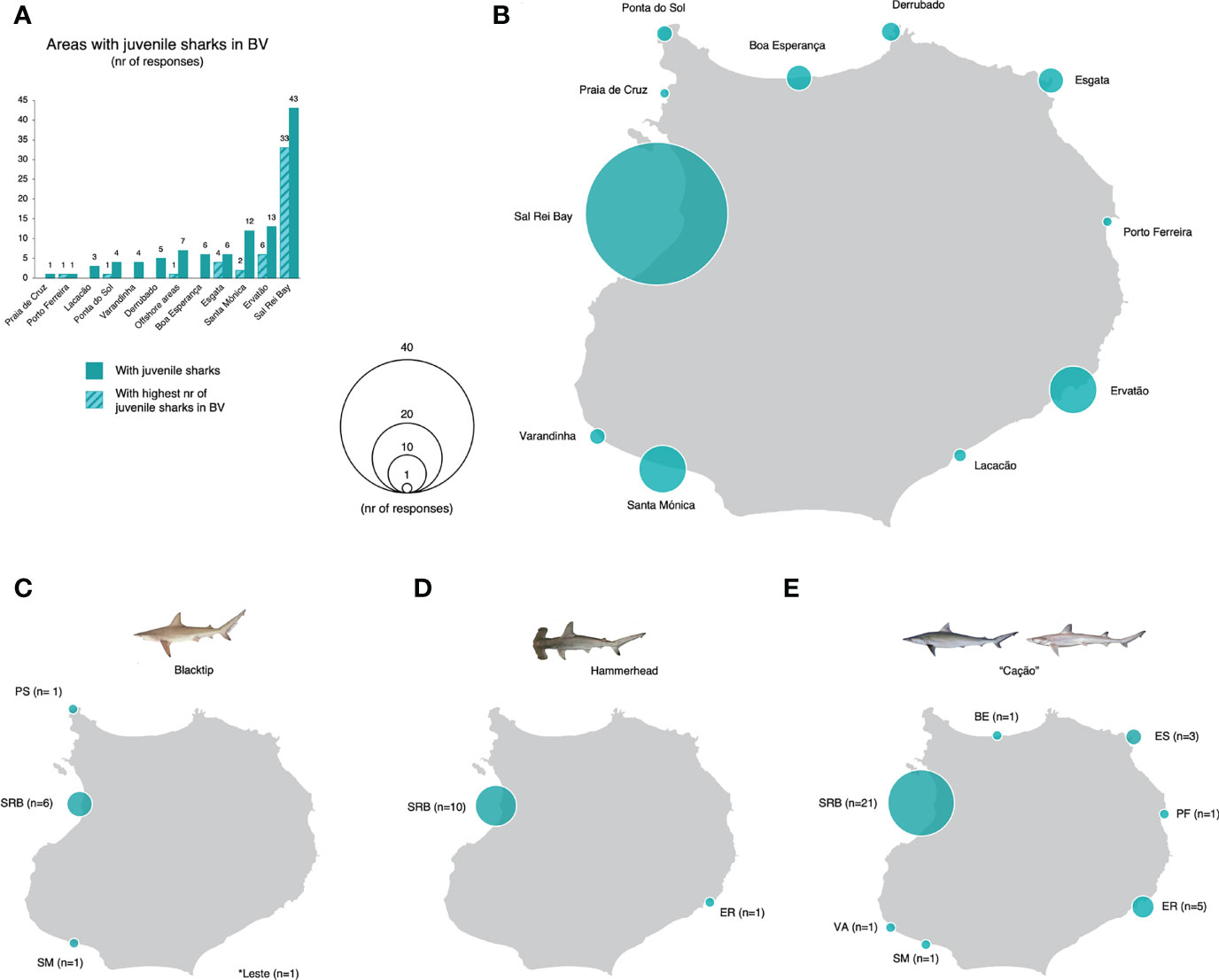
Figure 4 (A) Areas of occurrence of juvenile sharks in Boa Vista Island, Cabo Verde, according to local fishermen. Eleven areas were identified based on dedicated interviews (n=55) to local registered commercial and artisanal fishermen; six of these areas were considered as the areas with the highest number of juvenile sharks in Boa Vista Island. (B) Spatial distribution of the identified areas around the island of Boa Vista. Sal Rei Bay (SRB) collected the highest level of agreement among fishermen, both as an area with juvenile sharks (n=43; 78%) and as the area with the highest number of juvenile sharks (n=33; 60%). Respondents identified (C) areas of occurrence of juvenile blacktip sharks (n=9; 16%), (D) juvenile hammerhead sharks (n=11; 20%), and (E) juvenile “cação” (n=33; 60%). “Cação” is the common name used locally to refer to both milk and Atlantic weasel sharks – only two fishermen referred to Atlantic weasel sharks specifically, and one to “boca cumprido” referring to milk sharks. Identified offshore fishing grounds include “Leste”, “Rio de Janeiro”, “Costa de Mar”, “Txom Branco”, “Verde”, and “West” (these are large areas off the coast, locally known as “pesqueiros”). BV, Boa Vista.
Discussion
The present study shows that SRB is used by juveniles of, at least, 5 threatened shark species. According to the IUCN Red List of Threatened Species (IUCN, 2022), S. lewini is designated by as “Critically Endangered”, P. pectoralis as “Endangered”, and R. acutus, C. limbatus and G. cirratum as “Vulnerable”. Furthermore, although not caught during the present surveys, information acquired through citizen science and preliminary data based on baited remote underwater videos (BRUVs), suggest the presence of juveniles of other shark species, namely spinner (Carcharhinus brevipinna) and lemon (Negaprion brevirostris) sharks (see Supplementary Figure 5).
The high consistency of the results obtained based on local fishermen’s knowledge clearly showcases that shark juveniles are found more often in SRB than in other coastal areas of the island (Figure 4). This preference is potentially related to the fact that SRB is a shallow, relatively wave-protected area (mostly with less than 10 m of depth). Because the predominant direction of waves in Boa Vista Island is from the Northeastern quadrant (Gomes, 2019) under the influence of the Canary Current (Peña-Izquierdo et al., 2012), SRB has relatively low wave action, especially during summer months. Moreover, there is a small islet at the north side of the bay that provides further protection (Supplementary Figure 2). By contrast, most other areas in Boa Vista Island are high-energy exposed shores without noticeable barriers (e.g., reefs or small islets) (Gomes, 2019).
SRB is repeatedly used by juvenile sharks over the years (Figures 1, 2), with higher CPUE values during summer and autumn periods. With this information, we are confident that SRB fully meets both criteria I (i.e., sharks are found more often in the specific area than in neighboring ones) and III (i.e., the area is used repeatedly by sharks over the years), as defined by Heupel et al. (2007). While no definite conclusions can be drawn regarding criterion II (i.e., sharks tend to remain in the area for extended periods), the observation of neonates with umbilical scars at different healing stages (preliminary findings) together with the general range of sizes observed (Figure 4) suggest that these animals are likely to make use of the bay for at least a few weeks after birth, and, thus, likely to meet criteria II. In this context, and although direct efforts to definitively confirm criterion II are still required (e.g., mark-recapture studies, remote tracking, stable isotope analysis), we argue that the importance of the SRB as a potential nursery area for multiple threatened shark species must not be understated.
Ensuring the effective protection of the SRB potential nursery area is of the utmost relevance as all identified shark species – in particular scalloped hammerheads and Atlantic weasel sharks – are threatened by anthropogenic pressures, and vulnerable to extinction over the near future (IUCN, 2022). There are many types of potential marine protected areas (MPAs), from full to minimal protection, from the ones that exist in practice (implemented) to the ones only on paper (Grorud-Colvert et al., 2021). The SRB is partially encompassed by two of the Boa Vista Island protected areas, however these do not focus on the marine realm, and are not MPAs but “natural reserves” – over 80% of their extension corresponds to terrestrial area (Cabo Verde Parliament, 2014a; 2014b). Indeed, the Boa Esperança Natural Reserve and the Morro de Areia Natural Reserve do include “peripheral areas for marine protection” that extend up to 300 m offshore (Cabo Verde Parliament, 2014a; 2014b). Yet, the latter represent only a small fraction of the SRB (10%), with an area of c. 2.7 km2 (see Supplementary Figure 2). At the same time, these natural reserves are still pending implementation, which means that they do not yet have any management instruments in place (e.g., management plans) (Boa Vista Municipality, 2022). The latter is especially significant as benefits of MPAs are highly dependent on their effective implementation and management (Grorud-Colvert et al., 2021). Also, the lack of enforcement further undermines the effectiveness of MPAs, as observed in other islands of Cabo Verde (Vasconcelos et al., 2015).
New opportunities and risk also arise from the recently approved coastal and marine spatial plan of Boa Vista Island – Plano de Ordenamento da Orla Costeira e Mar adjacente da ilha da Boa Vista (POOCM) (Cabo Verde Parliament, 2020). The POOCM establishes a planning unit for SRB (the Sal Rei Bay Integrated Management Area) whose general goals are to: (1) minimize the risk of environmental impacts; and (2) regulate fishing, nautical sports, and other recreational activities to make them compatible with the protection and valorization of marine ecosystems. Still, human activities such as artisanal fisheries, aquaculture, renewable energy, and maritime transportation are generally allowed in the planning unit (Cabo Verde Parliament, 2020). At the same time, while the POOCM has several references to the protection of sea turtles and marine mammals, no references are found for sharks, reflecting the lack of attention to this particular taxonomic group. Also, the POOCM is to be further implemented by dedicated management plans and regulations that are still to be developed, and which sometimes take long to be put in place because of social-political factors (Frazão Santos et al., 2021). Finally, the close proximity to two “tourism development zones” (the Integral Tourism Development Zone of Chave and Integral Tourism Development Zone of Morro de Areia) makes the SRB further vulnerable to human pressures and impacts (Cabo Verde Parliament, 2008; 2009).
Further action is therefore needed to ensure the effective conservation of shark populations, and broader benthic and pelagic ecosystems, in the SRB. Such protection will largely depend on the specific conservation measures and monitoring plans that are put in place, but also on capacity building and awareness raising actions targeting national and international ocean users and stakeholders. Only then will it be possible to ensure an equitable and sustainable use and conservation of the SRB.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
Ethical review and approval was not required for the animal study because It entailed surveys on local artisanal bycatch fishery.
Author contributions
RR conceptualized the study. RR, EN, VP, CS, JV, JP, MB, TR, JC and CF collected field data. RR and TM performed data analyses. RR and CF wrote the original draft of the manuscript. All authors reviewed and edited the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This research is part of project NGANDU (The Importance of Shark Populations and Sustainable Ocean Use for Human Well-being in Cabo Verde and São Tomé and Príncipe, West Africa) funded by the Portuguese Foundation for Science and Technology (FCT) and the Aga Khan Development Network (AKDN) under grant agreement FCT AGA-KHAN/541746579/2019. All authors acknowledge funding from FCT under the strategic project UIDB/04292/2020 granted to MARE and project LA/P/0069/2020 granted to the Associate Laboratory ARNET. TM acknowledges funding from the strategic project UIDB/00006/2020 granted to CEAUL. CF acknowledges funding from FCT research contract 2020.03704.CEECIND and FCT grant agreement PTDC/CTA-AMB/30226/2017. VP acknowledges funding from FCT PhD grant 2020.05435.BD. CS acknowledges funding from FCT PhD grant SFRH/BD/117890/2016, FCT research grants under PTDC/CTA-AMB/30226/2017 through FCiencias.ID and AGA-KHAN/541746579/2019 through Nova School of Business and Economics. EN acknowledges funding from FCT PhD grant SFRH/BD/135438/2017. JRP acknowledges funding from FCT research contract 2021.01030.CEECIND. JV acknowledges funding from the Intergovernmental Panel on Climate Change (PhD grant, IPCC Scholarship Programme–Prince Albert II of Monaco Foundation) and the Camões–Instituto da Cooperação e da Língua, I.P. (Scholarship Programme).
Acknowledgments
The authors thanks the help provided by Eduardo Lima and João Pina during field work.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2023.1077748/full#supplementary-material
References
Baum J. K., Myers R. A., Kehler D. G., Worm B., Harley S. J., Doherty P. A. (2003). Collapse and conservation of shark populations in the Northwest Atlantic. Science 299, 389–392. doi: 10.1126/science.1079777
Boa Vista Municipality (2022) Boa Vista protected areas. Available at: www.municipiodaboavista.com/municipio/areas-protegidas/ (Accessed 01/09/2022).
Branstetter S. (1987). Age, growth and reproductive biology of the silky shark, Carcharhinus falciformes, and the scalloped hammerhead, Sphyrna lewini, from the northwestern gulf of Mexico. Environ. Biol. Fish. 19, 161–173. doi: 10.1007/BF00005346
Cabo Verde Parliament (2008). Ruling no. 20/2008 – Portaria n.° 20/2008, de 7 de julho. Cabo Verde: Boletim Oficial da República de Cabo Verde, I Série, 25, 502–530.
Cabo Verde Parliament (2009). Ruling no. 1/2009 – Portaria Conjunta n.° 1/2009, de 2 de fevereiro. Cabo Verde: Boletim Oficial da República de Cabo Verde, I Série, 5, 85–109.
Cabo Verde Parliament (2014a). Implementing degree no. 16/2014. Decreto-Regulamentar n.° 16/2014, de 10 de fevereiro. Cabo Verde: Boletim Oficial da República de Cabo Verde, I Série, 8, 280–282.
Cabo Verde Parliament (2014b). Implementing degree no. 17/2014. Decreto-Regulamentar n.° 17/2014, de 10 de fevereiro. Cabo Verde: Boletim Oficial da República de Cabo Verde, I Série, 8. 282–283.
Cabo Verde Parliament (2020). Ruling no. 112/2020 – Republicação n.° 112/2020, de 10 de setembro. Cabo Verde: Boletim Oficial da República de Cabo Verde, I Série, 107. 2518–2560.
Compagno L. J. V. (1984). FAO species catalogue, vol. 4. sharks of the world; an annotated and illustrated catalogue of shark species known to date. part 2. carcharhiniformes. FAO Fish. Synopsis 125, 250–655.
Compagno L. J. V. (1990). Alternative life-history styles of cartilaginous fishes in time and space. Environ. Biol. Fish. 28, 33–75. doi: 10.1007/BF00751027
Diemer K. M., Mann B. Q., Hussey N. E. (2011). Distribution and movement of scalloped hammerhead Sphryna lewini and smooth hammerhead Sphyrna zygaena sharks along the east coast of southern Africa. Afr. J. Mar. Sci. 33, 229–238. doi: 10.2989/1814232X.2011.600291
Dulvy N. K., Fowler S. L., Musick J. A., Cavanagh R. D., Kyne P. M., Harrison L. R., et al. (2014). Extinction risk and conservation of the world's sharks and rays. Elife 3, e00590. doi: 10.7554/eLife.00590
Dulvy N. K., Pacoureau N., Rigby C. L., Pollom R. A., Jabado R. W., Ebert D. A., et al. (2021). Overfishing drives over one-third of all sharks and rays toward a global extinction crisis. Curr. Biol. 31, 5118–5119. doi: 10.1016/j.cub.2021.11.008
Ferretti F., Worm B., Britten G. L., Heithaus M. R., Lotze H. K. (2010). Patterns and ecosystem consequences of shark declines in the ocean. Ecol. Lett. 13, 1055–1071. doi: 10.1111/j.1461-0248.2010.01489.x
Frazão Santos C., Agardy T., Andrade F., Crowder L. B., Ehler C. N., Orbach M. K. (2021). Major challenges in developing marine spatial planning. Mar. Policy 132, 103248. doi: 10.1016/j.marpol.2018.08.032
Freitas R., Romeiras M., Silva L., Cordeiro R., Madeira P., Gonzalez J. A., et al. (2019). Restructuring of the 'Macaronesia' biogeographic unit: A marine multitaxon biogeographical approach. Sci. Rep. 9, 15792. doi: 10.1038/s41598-019-51786-6
García V. B., Lucifora L. O., Myers R. A. (2008). The importance of habitat and life history to extinction risk in sharks, skates, rays and chimaeras. Proceeding R. Soc. B 275, 83–89. doi: 10.1098/rspb.2007.1295
Gomes C. C. (2019). Climate change impacts on island coastal evolution: The case of boa vista (Cabo Verde) (Lisboa, Portugal: Faculdade de Ciências, Universidade de Lisboa). PhD Thesis.
Grorud-Colvert K., Sullivan-Stack J., Roberts C., Constant V., Costa B. H. E., Pike E. P., et al. (2021). The MPA guide: A framework to achieve global goals for the ocean. Science 373, abf0861. doi: 10.1126/science.abf0861
Henderson A. C., Jourdan A., Bell K. (2016). Assessing the incidental value of a marine reserve to a lemon shark Negaprion brevirostris nursery. Aquat. Conserv.: Mar. Freshw. Ecosyst. 26, 482–491. doi: 10.1002/aqc.2627
Heupel M. R., Carlson J. K., Simpfendorfer C. A. (2007). Shark nursery areas: concepts, definition, characterization and assumptions. Mar. Ecol. Prog. Ser. 337, 287–297. doi: 10.3354/meps337287
Heupel M. R., Kanno S., Martins A. P. B., Simpfendorfer C. A. (2019). Advances in understanding the roles and benefits of nursery areas for elasmobranch populations. Mar. Freshw. Res. 70, 897–907. doi: 10.1071/MF18081
Heupel M. R., Simpfendorfer C. A. (2002). Estimation of mortality of juvenile blacktip sharks, Carcharhinus limbatus, within a nursery area using telemetry data. Can. J. Fish. Aquat. Sci. 59, 624–632. doi: 10.1139/f02-036
IUCN (2022) The IUCN red list of threatened species. version 2022-1. Available at: www.iucnredlist.org (Accessed 8/8/2022).
Kinney M. J., Hussey N. E., Fisk A. T., Tobin A. J., Simpfendorfer C. A. (2011). Communal or competitive? Stable isotope analysis provides evidence of resource partitioning within a communal shark nursery. Mar. Ecol. Prog. Ser. 439, 263–276. doi: 10.3354/meps09327
Knipp D. M., Heupel M. R., Simpfendorfer C. A. (2010). Sharks in nearshore environments: Models, importance and consequences. Mar. Ecol. Prog. Ser. 402, 1–11. doi: 10.3354/meps08498
Marco A., Abella E., Liria-Loza A., Martins S., Lopez O., Jimenez-Bordon S., et al. (2012). Abundance and exploitation of loggerhead turtles nesting in Boa Vista island, Cape Verde: the only substantial rookery in the eastern Atlantic. Anim. Conserv. 15, 351–360. doi: 10.1111/j.1469-1795.2012.00547.x
Martins S., Patricio R., Clarke L. J., Loureiro N. D., Marco A. (2022). High variability in nest site selection in a loggerhead turtle rookery, in Boa Vista island, Cabo Verde. J. Exp. Mar. Biol. Ecol. 556, 151798. doi: 10.1016/j.jembe.2022.151798
Pacoureau N., Rigby C. L., Kyne P. M., Sherley R. B., Winker H., Carlson J. K., et al. (2021). Half a century of global decline in oceanic sharks and rays. Nature 589, 567–56+. doi: 10.1038/s41586-020-03173-9
Peña-Izquierdo J., Pelegrí J. L., Pastor M., Castellanos P., Emelianov M., Gasser M., et al. (2012). El Sistema de corrientes de talud continental entre Cabo Verde y las Islas Canarias. Sci. Marina 76, 65–78. doi: 10.3989/scimar.03607.18C
Queiroz N., Humphries N. E., Couto A., Vedor M., Costa I., Sequeira A. M. M., et al. (2019). Global spatial risk assessment of sharks under the footprint of fisheries. Nature 572, 461–46+. doi: 10.1038/s41586-019-1444-4
Queiroz N., Humphries N. E., Mucientes G., Hammerschlag N., Lima F. P., Scales K. L., et al. (2016). Ocean-wide tracking of pelagic sharks reveals extent of overlap with longline fishing hotspots. Proc. Natl. Acad. Sci. USA 113, 1582–1587. doi: 10.1073/pnas.1510090113
Roberts C. M., Mcclean C. J., Veron J. E. N., Hawkins J. P., Allen G. R., Mcallister D. E., et al. (2002). Marine biodiversity hotspots and conservation priorities for tropical reefs. Science 295, 1280–1284. doi: 10.1126/science.1067728
Roff G., Doropoulos C., Rogers A., Bozec Y. M., Krueck N. C., Aurellado E., et al. (2016). The ecological role of sharks on coral reefs. Trends Ecol. Evol. 31, 395–407. doi: 10.1016/j.tree.2016.02.014
Simpfendorfer C. A., Milward N. E. (1993). Utilisation of a tropical bay as a nursery area by sharks of the families Carcharhinidae and Sphyrnidae. Environ. Biol. Fish. 37, 337–345. doi: 10.1007/BF00005200
Speed C. W., Field I. C., Meekan M. G., Bradshaw C. J. A. (2010). Complexities of coastal shark movements and their implications for management. Mar. Ecol. Prog. Ser. 408, 275–293. doi: 10.3354/meps08581
Springer S. (1967). “Social organization of shark populations,” in Sharks, skates and rays. Eds. Gilbert P. W., Matheson R. F., Rall. D. P. (Baltimore: John Hopkins Press), 149–174.
Vasconcelos R., Freitas R., Hazevoet C. J. (2015). História natural das ilhas desertas – Santa luzia, branco e raso (Porto, Portugal: Sociedade Caboverdiana de Zoologia).
Wenzel F. W., Broms F., Lopez-Suarez P., Lopes K., Veiga N., Yeoman K., et al. (2020). Humpback whales (Megaptera novaeangliae) in the Cape Verde islands: Migratory patterns, resightings, and abundance. Aquat. Mammals 46, 21–31. doi: 10.1578/AM.46.1.2020.21
Wheeler C. R., Gervais C. R., Johnson M. S., Vance S., Rosa R., Mandelman J. W., et al. (2020). Anthropogenic stressors influence reproduction and development in elasmobranch fishes. Rev. Fish Biol. Fish. 30, 373–386. doi: 10.1007/s11160-020-09604-0
Keywords: elasmobranchs, juveniles, parturition area, marine conservation, Western Africa, sharks
Citation: Rosa R, Nunes E, Pissarra V, Santos CP, Varela J, Baptista M, Castro J, Paula JR, Repolho T, Marques TA, Freitas R and Frazão Santos C (2023) Evidence for the first multi-species shark nursery area in Atlantic Africa (Boa Vista Island, Cabo Verde). Front. Mar. Sci. 10:1077748. doi: 10.3389/fmars.2023.1077748
Received: 23 October 2022; Accepted: 13 January 2023;
Published: 01 February 2023.
Edited by:
Elizabeth Grace Tunka Bengil, University of Kyrenia, CyprusReviewed by:
Colombo Estupiñán-Montaño, Fundación Alium Pacific, ColombiaDominic Swift, Texas A&M University Corpus Christi, United States
Copyright © 2023 Rosa, Nunes, Pissarra, Santos, Varela, Baptista, Castro, Paula, Repolho, Marques, Freitas and Frazão Santos. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Rui Rosa, rrosa@fc.ul.pt