 Céline Blaye
Céline Blaye Thomas Boyer1
Thomas Boyer1 Florent Peyraud
Florent Peyraud Charlotte Domblides
Charlotte Domblides Nicolas Larmonier
Nicolas Larmonier- 1Centre National de la Recherche Scientific (CNRS) Unité Mixte de Recherche (UMR) 5164, ImmunoConcEpT, Bordeaux, France
- 2Department of Medical Oncology, Institut Bergonié, Bordeaux, France
- 3Service d’Oncologie Médicale, Centre Hospitalo-Universitaire (CHU) Bordeaux, Bordeaux, France
- 4Department of Biological and Medical Sciences, University of Bordeaux, Bordeaux, France
Breast cancers are commonly associated with an immunosuppressive microenvironment responsible for tumor escape from anti-cancer immunity. Cells of the myeloid lineage account for a major part of this tumor-promoting landscape. These myeloid cells are composed of heterogeneous subsets at different stages of differentiation and have traditionally been described by their cardinal ability to suppress innate and adaptive anticancer immunity. However, evidence has accumulated that, beyond their immunosuppressive properties, breast cancer-induced myeloid cells are also equipped with a broad array of “non-immunological” tumor-promoting functions. They therefore represent major impediments for anticancer therapies, particularly for immune-based interventions. We herein analyze and discuss current literature related to the versatile properties of the different myeloid cell subsets engaged in breast cancer development. We critically assess persisting difficulties and challenges in unequivocally discriminate dedicated subsets, which has so far prevented both the selective targeting of these immunosuppressive cells and their use as potential biomarkers. In this context, we propose the concept of IMCGL, “pro-tumoral immunosuppressive myeloid cells of the granulocytic lineage”, to more accurately reflect the contentious nature and origin of granulocytic cells in the breast tumor microenvironment. Future research prospects related to the role of this myeloid landscape in breast cancer are further considered.
Highlights
Beyond their cardinal immunosuppressive properties, many subsets of myeloid cells are equipped with multiple tumor-promoting functions impacting most steps of cancer development.
Introduction
Centered for years on the intrinsic characteristics of tumor cells, the field of cancer research has evolved toward the notion that cancers emerge and develop in a dedicated tumor-promoting environment. The cross-talks between malignant cells and components of this tumor-specific landscape dictate the fate of cancer (persistence or elimination) and further shape the nature of this microenvironment (1, 2). In this context, the influence of the immune system on cancer development has been widely evidenced, and many strategies have been developed to induce, restore and enhance anti-cancer immunity. Successes of these immune-based approaches in inducing efficient anti-tumor responses and improving cancer patient survival have brought some of them to the forefront of cancer therapeutics in recent years (3). However, it has also become clear that cancers can escape from immune detection and destruction by many mechanisms resulting in the establishment of an immunosuppressive tumor environment, which represents a major obstacle for efficient immunotherapies. Compelling evidences have indicated that inhibition of these immunosuppressive networks is an important prerequisite to uncover the full potential of immune-based interventions. It is noteworthy that, although several immunotherapies provide clinical benefits in melanoma, lung, bladder and colon cancers, breast cancer patients have yet to fully experience these breakthroughs. Indeed, except for triple-negative cancers which are more immunogenic and have obtained FDA approval of immunotherapies in the neo-adjuvant (4) and metastatic (5) settings, most immune-based therapeutic attempts in breast cancers have ended in failure.
For many years, the environment of breast tumors has been described as “immunologically cold”, as defined by the sparsity or absence of tumor-infiltrating lymphocytes (TILs) (6). This description is somewhat inaccurate insofar as it usually does not take into account cells of myeloid origin, despite their many diverse roles in the environment of mammary cancers. The lack of anti-tumoral immune response in breast cancers has indeed been associated with a hostile immuno-inhibitory microenvironment, the major components of which being cells of myeloid origin (7). Tumor-associated macrophages (TAMs), tumor-associated neutrophils (TANs), tolerogenic dendritic cells (tDC) and immature subsets of myeloid cells endowed with immunosuppressive properties termed “myeloid-derived suppressor cells” (MDSCs) have been identified as such myeloid subpopulations, present not only within the tumor environment, but also at the sites of priming of antitumoral immune responses (secondary lymphoid organs), in the bloodstream and in the pre-metastatic and metastatic sites.
Besides their ability to impair anti-tumor immunity at different steps of immune responses (initiation, priming, effector stages), these myeloid cells are also endowed with a large array of “non-immunologic” tumor-promoting functions. They can indeed contribute to the epithelial-to-mesenchymal transition (EMT), participate to local tissue invasion at the primary tumor site, foster blood or lymphatic vessel intravasation and extravasation of migrating cancer cells, associate with circulating tumor cells protecting them in the bloodstream, and prepare the pre-metastatic niches thus enhancing metastatic processes. Furthermore, these myeloid cells can also directly promote primary tumor cell survival and proliferation and foster tumor neoangiogenesis and cancer cell stemness (Figure 1). The role of breast cancer-induced myeloid cells in resistance to chemotherapy as well as endocrine therapy has also been described, making them potential targets for the development of new immunotherapies.
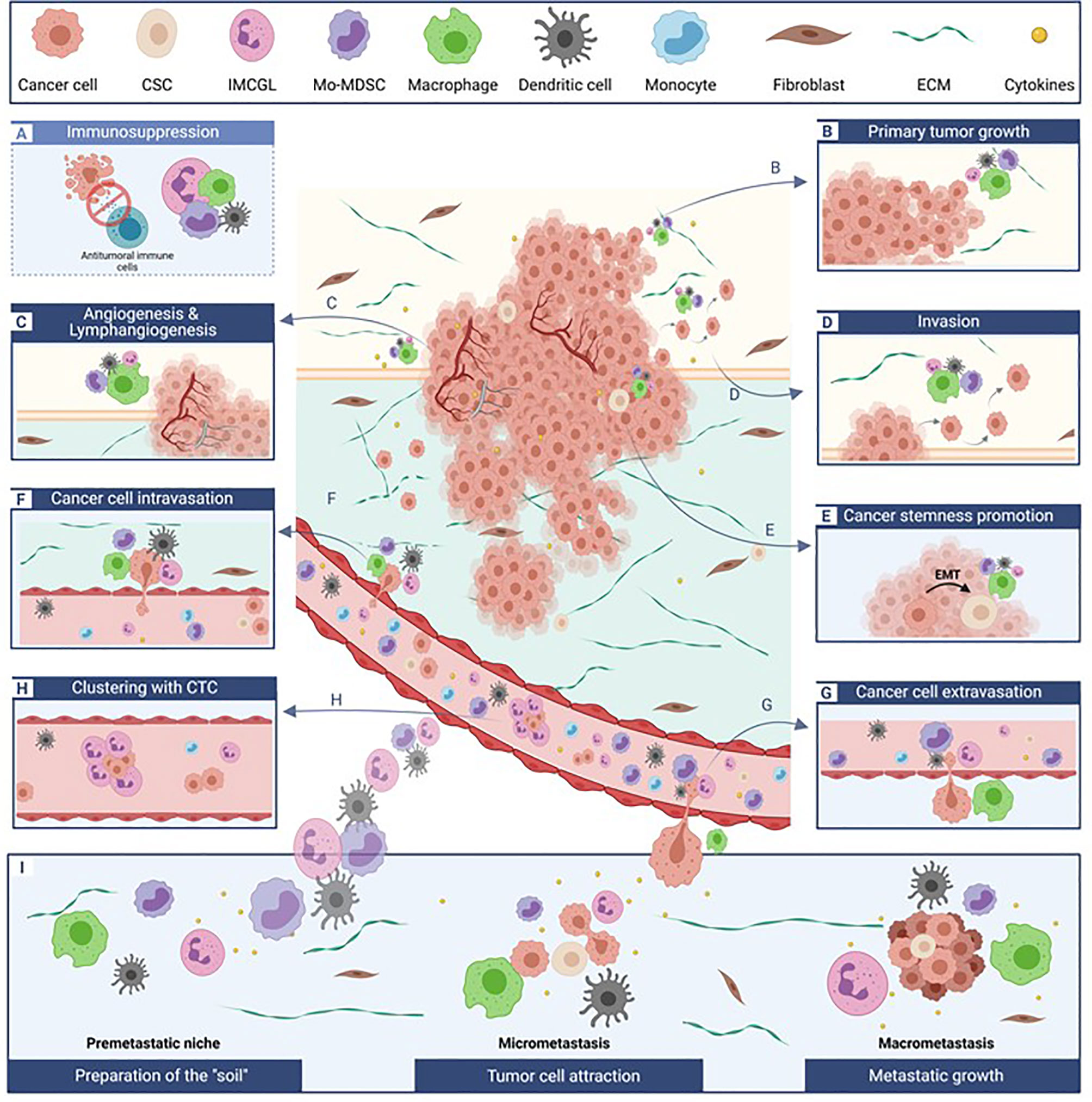
Figure 1 Tumor-promoting myeloid cells critically affect multiple and distinct steps of cancer development. Besides impairing anti-tumor immunity. (A), dedicated subpopulations of myeloid cells differentially impact primary tumor survival and growth (B), tumor vascularization (C), local tissue invasion (D) cancer stemness (E), tumor cell intra- (F) and extravasation (G) in and from blood vessels, associate with circulation tumor as beneficial clusters (H), and participate to metastatic site preparation and development (I). The relative contribution of each myeloid cell subset to a specific process (each illustrated in a separate box) is depicted by the differential size of the cells. CSC, cancer stem cells; CTC, circulating tumor cells; ECM, extra-cellular matrix; EMT, epithelial-to-mesenchymal transition; IMCGL, immunosuppressive myeloid cells from granulocytic lineage; Mo-MDSCs, monocyctic myeloid-derived suppressor cells.
Recently, major advances in the characterization of the phenotypes, functions and origins of myeloid cell subpopulations in breast cancers have been made, particularly by single cell RNA sequencing approaches. In this review, we discuss the equivocal identify of some subsets, particularly “polymorphonuclear-MDSCs” (“PMN-MDSCs”) and “immunosuppressive neutrophils”, and examine and discuss the polyvalent tumor-promoting functions of these myeloid cells in the breast cancer environment in light of recent literature, with a specific emphasis on the “non-immunologic” pro-tumoral properties of these multitasking cells.
Macrophages in Breast Cancers: A Multifunctional Impact on Tumor Promotion
Macrophage Phenotype and Function in the Context of Breast Cancer
Macrophages have been one of the most widely studied population of myeloid cells in cancer, specifically in the context of breast malignancies. In breast cancer patients, tumor-associated macrophages (TAM) have been associated with aggressive features (size ≥ 2 cm, higher tumor grade, higher Ki67) and estrogen receptor (ER) negative breast cancers (8). However, the prognostic value of these cells remains controversial and depends on the cancer subtype, the macrophage subset (M1 vs M2, see below) and their localization (9). Indeed, some authors have described an improved survival of ER- or triple-negative breast cancer (TNBC) patients with a CD163+CD68+ macrophage infiltrate (10), while others correlated the presence of tumor-infiltrating CD163+ macrophages with a worse prognosis of TNBC patients (11). In a gene-expression based study using a CIBERSORT deconvolution method, macrophages have been associated with a significant poorer outcome in both ER+ and ER- BC patients, and were predictive of a worse response to chemotherapy in ER- patients (12). Independently of BC subtype, immune population clustering identified 2 clusters enriched in pro-tumorigenic macrophages, which have been associated with significantly worse outcome in BC patients (12).
The polyvalent functions and the high degree of plasticity of macrophages are partly responsible for these conflicting results. Macrophages have originally been broadly discriminated in two different types with opposite roles. “M1” macrophages have been described as classically activated, pro-inflammatory, anti-tumoral effectors, whereas “M2” macrophages correspond to alternatively activated cells endowed with “wound-healing” and tumor-promoting functions. M1 and M2 represent in fact two extreme polarization states of a highly plastic differentiation program controlled by environmental cues (13). M1 macrophages can be induced by TLRs ligands such as LPS, and/or IFNγ. Their differentiation is driven by STAT1, IRF5, NF-kB (14). M2 macrophages are primarily induced by IL-4 and/or IL-13 (+/- IL-10) via the IL-4Rα receptor and their differentiation depend mainly on STAT3, STAT6, IRF4 activation. Different markers have been used to distinguish between M1 and M2 macrophages, but most of these molecules are expressed by both types, although at different levels (14). In immunohistochemical studies, CD68 is often used as a “pan-macrophages” marker. M1 are described as iNOS-expressing cells, with high expression of MHC class II and detectable co-stimulatory molecules CD80 and CD86. M2 macrophages conventional markers include CD163, the scavenger receptor CD204 and mannose receptors CD206, as well as a high expression of Arg1 (9). However, this M1/M2 dichotomy has been challenged and it may not be fully relevant in the context of chronic, non resolutive inflammation such as cancer (14). In fact, recent transcriptomic data, RNA sequencing and mass-cytometry analyses argue for a more complex and heterogeneous phenotypic identity of breast cancer-associated macrophages. A single-cell RNA-seq analysis of 8 tumors (matched to healthy tissues from the same patients) uncovered numerous clusters of immune cells. Among them, three different clusters of TAMs were described (15). Whether TAM may originate from bone-marrow-derived monocytes or from tissue-resident macrophages, which derive from embryonic macrophages that colonize developing organs during the process of embryogenesis and that persist in mature developed adult organs, has been debated. The abovementioned study indicated that the three identified distinct TAM clusters originate either from monocytes or from resident macrophages. Interestingly, among these TAMs, the M1 gene signature correlated with that of M2, advocating for a simultaneous activation of these different genes (15). Along these lines, a mass-cytometry analysis of 144 breast tumors (compared with 46 matched juxta-tumoral tissue and four mammoplasties from cancer-free individuals) defined 19 clusters of myeloid cells and highlighted a frequent co-expression (although at different levels) of phenotypic markers of both M1 and M2 by TAMs such as CD169, CD86, CD204, CD206 and CD163 (16). Consistent with these studies, a RNA-seq analysis indicated that TAMs from breast and endometrial cancers did not exhibit typical M2 gene signatures (17). In this study, TAMs from these two different types of cancers revealed very small similarities, emphasizing the crucial role of the TME in differentially shaping macrophage phenotype and function (17). These data thus indicate that TAMs in breast cancers exhibit complex overlapping phenotypic and functional characteristics and cannot be simplistically categorized as conventional M1 vs M2. With regard to the origin of TAMs in breast cancers, the aforementioned single-cell RNA-seq analysis indicated that these cells can originate either from resident macrophages or from monocyte differentiation (15).
TAMs at the Primary Tumor Site
At the primary tumor site, crosstalks between macrophages and cancer cells contribute to the recruitment and activation of TAMs, which in turn foster tumor progression through many mechanisms. Particularly, the immunosuppressive activity of these cells has been extensively described. Indeed, macrophages can suppress anti-tumoral T lymphocytes responses via their catabolism of L-arginine and/or tryptophan (expression of iNOS, IDO, arginase), production of immunosuppressive cytokines such as IL-10, IL-4, IL-17, CXCL1, or the expression of ligands for immune checkpoint inhibitory receptors such as PD-L1. They also produce chemoattractant chemokines that further recruit immunosuppressive cells such as neutrophils, immature DCs and/or Tregs [reviewed in (18)]. Along these lines, IL-1ß production by TAM has been shown to participate to the recruitment of immunosuppressive cells and thus to overall suppression of adaptive immune responses (19, 20). In mice, specific targeting of these immunosuppressive macrophages or inhibition of their immunoinhibitory functions can restore anti-tumor immune responses (21).
However, tumor-associated macrophages can also display pro-angiogenic functions and can promote cancer cell stemness. For instance, the transcription factor POU class 1 homeobox 1 (POU1F1, also known as Pit-1), a protein expressed by breast cancer cells, has been reported to increase macrophage recruitment and to promote their polarization towards VEGFA-expressing tumor-promoting macrophages. In turn, these macrophages foster tumor growth, angiogenesis and extravasation of breast cancer cells in a CXCL12-dependent manner in vitro (22). Likewise, the expression by breast cancer cells of the ID4 protein (a member of inhibitors of differentiation family of proteins), which is associated with a basal, stem-like phenotype and poor prognosis in TNBC, induces the activation of a pro-angiogenic program in macrophages with upregulation of angiogenesis-related transcripts (23). It is noteworthy that pro-tumorigenic TAM infiltration is more prominent in inflammatory breast cancer (IBC), a disease with a very poor prognosis, compared to other breast cancer subtypes. These macrophages are recruited and polarized into a pro-tumoral phenotype (upregulation of CD206, CD163 and CD209) by CSF-1, CXCL2, VEGFA and CCL18 produced by cancer cells (24). In hypoxic zones, breast cancer cells produce Oncostatin M (OSM) that induces macrophage polarization toward a tumor-promoting phenotype (higher expression of CD163, CD206, Arg1 and Cox-2) (25). Hypoxia enhances TAM expression of galectin-3, a ß-galactoside binding protein modulating TAM apoptosis, migratory and adhesive properties. These macrophages have been shown to promote the proliferation, invasion and migration of MDA-MB-231 breast cancer cells and angiogenesis in vitro. Furthermore, in vivo experiments in Balb/c mice exposed to hypoxia indicate that targeting Galectin-3 decreases lung metastasis burden and reduces endothelial cell in the primary tumor (26). The presence of sexual steroids in the TME, particularly the presence of estrogens, is a specificity of breast malignancies as breasts are made of adipose tissues producing sexual steroids in the environment. ER+ breast cancers arising in this environment are uniquely capable of responding to these signals and grow. Interestingly, it has been reported that in BC patients, estrogens induce the production of CCL2 and CCL5 within the tumor beds, leading to the recruitment and polarization of macrophages towards a pro-tumorigenic phenotype (27). The reversal of estrogen effects using Tamoxifen led to a reduced infiltration of these pro-tumoral macrophages in the primary tumor, a finding further confirmed in murine models (28).
Macrophages recruited and accumulating at the tumor site also contribute to tumor development through the promotion of cancer cell stemness via secretion of IL-8 and CXCL1, 2 and 3 (24). Cancer stem cells (CSCs) are described as “tumor-initiating cells” with the capability of self-renewal and asymmetric proliferation, and are characterized by a reduced sensitivity to drugs and irradiation compared to non-CSCs. These CSCs are critical for cancer dissemination and metastasis (29). The acquisition of stemness properties by cancer cells has been associated with the induction of the EMT (epithelial-to-mesenchymal transition) program. EMT is controlled by transcription factors such as TWIST, ZEB1, SNAIL, or SLUG, and is characterized by specific phenotypic changes whereby epithelial cancer cells acquire a mesenchymal-like phenotype, which increases their invasive and migratory potency (29). CCL2 and CXCL12 produced by breast cancer-associated fibroblasts and tumor cells promote the recruitment and differentiation of monocytes into immunosuppressive TAMs. In turn, these TAMs upregulate the expression of Vimentin, decrease the expression of E-cadherin, and induce Twists, Snail and Slug expression by breast cancer cells, thereby promoting the acquisition of mesenchymal and stemness properties by the latter (30). In a xenograft mouse model, CD68+ TAMs have been demonstrated to promote breast cancer cell stemness through expression of the transmembrane protein LSECtin, which engaged BTN3A3 (B7 family member) on breast tumor cells (31). In the same study, the authors have found a co-localization between LSECtin-expressing macrophages and breast cancer cells expressing CD90 – a stemness marker in breast cancer (31). The role of CD90 in the anchorage of monocytes/macrophages to cancer cells had previously been highlighted in a previous study (32). This CD90-dependent bound leads to the production of cytokines such as IL-6, IL-8, GM-CSF by cancer stem cells which further support cancer stemness (32). Finally, in the inflammatory context of obesity, mammary adipose tissue macrophages can be reprogrammed into a pro-inflammatory metabolically activated phenotype (MMe), which can promote tumor initiation and triple negative breast cancer stem-like properties through an IL-6/GP130-dependent mechanism (33).
TAMs in the Metastatic Process
TAMs also play an essential role at most steps of breast cancer metastasis. As outlined above, these myeloid cells contribute to breast cancer EMT and stemness, two essential initial steps required for tumor systemic dissemination (24, 30–32). TAMs located in the tumor beds or at their vicinity have also been reported to promote intravasation of migrating cancer cells from the primary tumor in blood vessels, while TAMs at the metastatic sites may contribute to the preparation of the pre-metastatic niches before colonization by cancer cells, and enhance breast cancer cell extravasation from blood capillaries in distal metastatic tissues (9).
Many reports have described the influence of TAMs in breast cancer cells intravasation, but the underlying mechanisms in vivo have not been extensively studied. A real-time imaging analysis in the MMTV-PyMT mouse model has indicated that VEGF-A produced by Tie2-expressing macrophages induced the loss of vascular junctions and transient vascular permeability, allowing for breast tumor cell intravasation (34). More recent studies have identified proteins involved in pro-tumoral macrophage promotion of cancer cell invasion in vitro assays. Chitinase 3-like protein 1 (CHI3L1, a glycoprotein highly expressed in solid tumors) secreted by macrophages has been shown to enhance breast cancer cell invasion, migration and adhesion. CHI3L1 has been detected in the sera of patients with breast carcinomas but not in healthy individuals. Analysis of GEO databases has indicated that CHI3L1 is associated with a worse prognosis in breast cancer patients (35). Use of 2D, 3D and Transwell migration assays have also underlined the role of pro-tumorigenic macrophage-secreted CCL-18 in promoting breast cancer cell migration (36). TAMs can also indirectly foster tumor dissemination by promoting the expansion of pro-metastatic neutrophils by an IL-1ß-dependent mechanism (see neutrophil section below) (37). Even in early and non-invasive breast cancers (in situ carcinomas) in mice, CCL2-recruited tumor-infiltrating macrophages with pro-tumorigenic features (CD206+/Tie2+), downregulate expression of E-cadherin by malignant cells, thus destabilizing cell-cell junctions, which leads to cancer dissemination and metastasis. These data advocate further for a decisive role of these cells in the establishment of metastatic disease (38).
Macrophages can also contribute to the preparation of the pre-metastatic niches and the promotion of breast cancer cell extravasation from blood vessels in distant sites. Indeed, monocytes, recruited to pre-metastatic niches by the CCL2, have been reported to quickly differentiate into pro-metastatic macrophages, which contribute to metastatic disease (9, 18). More recently, the presence of CYP4A-expressing TAMs in uninvolved tumor draining lymph nodes has significantly been correlated with the expression of markers associated with pre-metastatic niche formation (VEGFR1, S100A8 and fibronectin), and with a reduced overall and relapse-free survival of patients. In the same study, the specific targeting of CYP4A using pharmacological approaches in 4T1 breast tumor-bearing mice reprogrammed tumor-infiltrated TAMs with a F4/80+CD206+ phenotype into TAM with a F4/80+iNOS+ “anti-tumor” phenotype, and reduced lung metastatic burden by impairing the preparation of the pre-metastatic niches (39). At the metastatic sites, macrophages further promote metastatic disease development by fostering vessel formation and directly enhancing cancer cell growth and survival, through the expression of VEGFA and downstream upregulation of MMP-9 (40).
TAMs have been recently described to promote lymphangiogenesis by two different mechanisms. First, expression of podoplanin, a transmembrane glycoprotein implicated in cell motility and adhesion, has been detected on TAMs at the vicinity of lymphatic vessels in the breast TME. The binding of podoplanin to the galectin 8 protein, a secreted glycan-binding protein expressed by lymphatic endothelial cells, promotes the secretion of the pro-migratory integrin ß1 by macrophages, which in turn fosters their migration and binding to lymphatics vessels where they induce matrix remodeling and promote vessel growth and lymphoinvasion. In the same study, podoplanin-expressing TAMs were associated with lymph node invasion and organ metastasis in a small cohort of breast cancer patients (41). Second, signaling through the sphingolipid sphingosine-1-phosphate receptor 1 (S1PR1) expressed on TAMs induced macrophage NLRP3 inflammasome expression, leading to the production of IL-1ß, which in turn directly acted on lymphatic endothelial cells to promote lymphangiogenesis. In mice deficient in S1PR1 in macrophages, lymphangiogenesis and metastatic growth are impaired. In human, NLRP3 expression in macrophages correlated with lymph nodes invasion and distant metastasis (42). Consistent with the aforementioned observations, in vitro experiments confirmed the role of macrophage-derived IL-1ß in the promotion of breast cancer cell adhesion to human lymphatic endothelial cells (43).
Breast Cancer Promotion by “Myeloid-Derived Suppressor Cells (MDSCs)”: beyond the Suppression of Anti-Tumor Immunity
“MDSCs”: A Functional Definition Rather Than a True Biological Subtype
The term “myeloid-derived suppressor cells (MDSCs)” was initially proposed by Gabrilovich et al. in 2007 in an effort to globally describe a heterogeneous population of myeloid cells exhibiting an immature phenotype and endowed with immunosuppressive functions (ability to suppress T lymphocytes), which accumulate in large numbers in the context of cancer (44). These cells have drawn intense scrutiny over the last 20 years and a considerable amount of data has been provided related to their participation to the complex immunoregulatory networks responsible for tumor immune escape. It has also become clear that they contribute to tumor development and dissemination through many different “immune-unrelated” mechanisms. “MDSCs”, in the context of cancer, derive from bone marrow hematopoietic precursors through aberrant myelopoiesis induced by tumor-derived factors (45). Many chemokines have been involved in MDSCs generation and recruitment to primary tumor sites or pre-metastatic niches, such as CXCL1, CXCL2, CXCL5, CXCL12, GM-CSF, G-CSF, M-CSF, VEGF, IL-6, IL-1ß or ß-FGF (46). More recently, breast cancer cells-derived exosomes have been shown to induce “MDSCs” from bone marrow myeloid progenitors (47), or to lead to their recruitment (48). In the specific estrogen-rich environment of breast cancer, these hormones have been shown to induce “MDSCs” recruitment via the activation of cancer associated fibroblasts, which in turn secrete CXCL12 (49). “MDSCs” have been defined as myeloid cells blocked at different stages in their differentiation toward mature terminally differentiated subsets such as macrophages and are thus associated with different degree of immaturity. This hallmark is however not always explored in many studies on “MDSCs”. The immunosuppressive capabilities of these cells, enabling them to block innate and adaptive anti-tumoral immune responses, represent their primary characteristic which must be systematically investigated for their identification as such (50).
If the term “MDSCs” was designed to globally encompass immature myeloid cells with many common features, it has nonetheless become confusing, particularly because it has contributed to consider these cells as a unique population of myeloid cells. However, “MDSCs” are made of highly heterogeneous populations, including cells from the monocytic and granulocytic lineage. In human, monocytic (M)-MDSCs have been defined as Lin-CD33+CD11b+HLA-DRlow/-CD14+CD15-, granulocytic (G) or polymorphonuclear (PMN)-MDSCs as CD33+CD11b+HLA-DRlow/-CD14-CD15+CD66b+, and “early stage” (more immature MDSC) (eMDSCs) as CD33+HLA-DR-Lin- (Lin: CD3, CD19, CD20, CD56, CD14, CD15) (50). In mice, MDSCs are CD11b+/Gr1+ cells, with Gr1 composed of two molecules, Ly6C (expressed on monocytic cells/M-MDSCs), and Ly6G (expressed on granulocytic cells/PMN-MDSCs) (51). As a main pitfall in the field, in many preclinical studies the phenotypical characterization of “MDSCs” has been limited to CD11b+Gr1+, which does not allow to discriminate between monocytic and granulocytic myeloid cells, each subset being endowed with distinct functions (52).
Granulocytic MDSC or “PMN-MDSC” constitute the majority of the MDSC pool in many cancers. However, phenotypically and functionally, these “PMN-MDSC” can hardly be distinguished from pro-tumoral immunosuppressive neutrophils and share the same phenotype as differentiated granulocytes. For these reasons, and since this overlap between PMN-MDSC and tumor-associated neutrophils remains a significant challenge in the field, we will discuss their phenotype and function together with that of cells of the granulocytic lineage in a dedicated section hereafter.
M-MDSCs and conventional monocytes share a similar phenotype, with however as main differences, lower expression of MHC Class II molecules and immunosuppressive capability for M-MDSC (53). However, “classical” CD14hiCD16lo monocytes may also exhibit low expression of HLA-DR, which is further reduced in the context of inflammation, sepsis, or cancer. In fact, cells with such a monocyte/M-MDSCs phenotype in the context of cancer have been shown to be immunosuppressive, blocking antitumoral T cell responses (54). In addition, both cell types have been reported to differentiate into pro-tumorigenic TAMs. This suggests a change of function of monocytes induced in the context of cancer (or other pathological conditions), rather than the occurrence of two different cell subtypes. Therefore, immunosuppressive monocytes and M-MDSC substantially overlap phenotypically and functionally. It is also noteworthy that some cells of myeloid origin exhibiting phenotypic characteristics that do not meet the classical definition of MDSC because they lack expression of specific MDSC markers (but which are nonetheless immunosuppressive and pro-tumoral) are excluded by this current MDSC terminology and may thus be overlooked. This is for instance the case of non-classical monocytes described in different cancers (55–58).
Overall, the “MDSCs” terminology is a conceptual approach, which was indispensable fifteen years ago to provide a comprehensive picture of a particular phenomenon observed in many cancers: the expansion of a myeloid cell population, more or less mature, with immunosuppressive properties. With the evolution of detection technologies such as scRNAseq, new knowledge of these cells has been brought, and it appears today as an essential requirement to regroup these myeloid cells according to their refined phenotype, in order to better identify and ultimately target them. However, since many studies do not allow such a discrimination (because of the use of an incomplete phenotype to identify these cells), in the next section, we will discuss the “all” MDSCs population (CD11b+/Gr1+ in mice, CD33+ in humans), the “early MDSCs” and the monocytic fraction (monocytes and M-MDSCs).
“MDSCs” in Breast Cancer Patients
In breast cancer patients, while some studies have reported that immunosuppressive monocytic cell number is increased compared to control patients (54), others did not observe this expansion (59). These cells have been associated with more advanced disease (60), and with a worse survival (61). Increased eMDSC numbers have also been correlated with a worse response to neo-adjuvant chemotherapy in TNBC patients (62). In breast cancer tissues, many reports have described, with various degree of accuracy, myeloid cells exhibiting an immature phenotype (CD33+CD13+CD14-CD15-) and immunosuppressive properties, which have been associated with adverse prognostic features (higher tumor grade, positive lymph nodes) (63). The composition of this myeloid infiltrate was different among studies, composed either with a majority of CD14+ monocytic immunosuppressive cells (64), or with granulocytic myeloid cells and early MDSCs (59). Together, these studies consistently advocate for the presence and role of suppressive myeloid cells in the tumor microenvironment, but their contradictory findings related to the exact phenotype of these cells highlights the extreme heterogeneity of this myeloid landscape, as further outlined more recently in scRNAseq and cytometry by time of flight (CyTOF) studies discussed in the next sections (15, 16).
Direct Effects of “MDSCs” on Tumor Cells
In primary tumor sites, suppressive myeloid cells recruited by cancer cells play an important role in inhibiting anti-tumor immune responses using many mechanisms extensively reviewed elsewhere (expression of ARG1, production of NO, ROS, and prostaglandin E2) (50, 51). However, many studies have also lent support to the notion that, besides their role as potent suppressor of cancer immunity, “MDSC” may also play an important role in breast cancer cell invasion, activate other stromal cells such as fibroblasts, and promote angiogenesis. Recruited at the hypoxic tumor sites, CD11b+Gr1+ MDSCs produce S100A8, an alarmin not only involved in the recruitment of additional MDSCs, but also implicated in the activation of endothelial cells. This activation led to the modification of tight junctions, leading to vascular leakage (65).
MDSCs also participate to breast cancer resistance to chemotherapy. It has indeed been shown that immunosuppressive CD33+ cells isolated from breast cancers patients are able to induce a stemness phenotype (associated with cancer cell chemoresistance) in the breast cancer cell line MCF-7 (64). Furthermore, the chemotherapeutic agent doxorubicin has been found to increase the levels of monocyte chemoattractant proteins (MCPs) 1 to 3 and particularly MCP1/CCL2 (48, 66). This chemotherapy-induced expression of CCL2 has also been reported in the metastatic sites such as the lungs. It results from the release of extracellular vesicles enriched in annexin-6 by chemoresistant cancer cells, which induced the recruitment of Ly6C+CCR2+ monocytes that participate to the pre-metastatic niche formation (48).
Preparation of the Metastatic Niche by Recruited “MDSCs”
CD11b+Gr1+ granulocytic and monocytic myeloid cells critically contribute to the metastatic dissemination of breast cancer cells (52). It has been observed that in the mouse 4T1.2 mammary cancer model, pro-inflammatory monocytes with MDSCs features (IL4R, CD49b, CD62L, CD11b) can be recruited by the chemokine CCL2 in the pre-metastatic lungs where they foster metastasis. CCL2 promotes the release of the alarmin S100A8/9 which further increases MDSC recruitment (67). CD11b+Gr1+ cells have been reported to progressively accumulate in the lungs of 4T1 tumor-bearing mice before the arrival of cancer cells, and to prepare the lung environment for seeding by metastatic cells via vascular remodeling and production of MMP9 (68). Interestingly, this recruitment of CCR2+ cells induced by inflammatory signals can be mediated by other sources of inflammation than tumors. Indeed, myocardial infarction represent a major cause of systemic stress and is accompanied by systemic monocytosis. It has been shown to be associated with a higher risk of relapse and cancer-specific mortality in early breast cancer patients (69). In tumor-bearing mice, myocardial infarction results in an important recruitment of Ly6C+ monocytes with immunosuppressive functions, which can differentiate into pro-tumorigenic macrophages at the tumor sites and accelerate primary tumor growth and metastasis (69). It has also been reported in the 4T1 triple negative mouse breast cancer model that Gr1+ cells primarily promote the metastatic cascade by facilitating extravasation of malignant cells at the distant metastatic lungs through IL1β and matrix metalloproteinase secretion (70). Along these lines, CXCR4-dependent mechanisms were involved in Gr1+ cell-mediated metastasis promotion in a mouse breast cancer model (71)
At the future metastatic site, MDSCs are implicated in the angiogenic switch. In two different studies in 4T1-bearing mice, CD11b+Gr1+ cells recruited in the lungs have been demonstrated to upregulate many pro-angiogenic factors such as Il1ß, Mmp9, Tnf, Tie2 (72), or to secrete platelet-derived growth factor-BB (PDGF-BB), which mediates angiogenesis (73).
Differentiation of “MDSCs” Into Other Cell Types in the Context of Breast Cancer
“MDSC” are endowed with a particularly high degree of plasticity. Indeed, many reports indicate that monocytes/M-MDSCs often differentiate into pro-tumorigenic macrophages at the tumor site or in the metastatic organs. This phenomenon has been tracked in vivo with the use of GFP+ expressing myeloid cells, transferred into E0771-bearing C57BL/6 mice: few hours after the transfer, classical monocytes were recruited in the metastatic lungs, where they differentiated into macrophages precursors, before becoming metastasis-associated macrophages (74). Exosomes derived from mesenchymal stem cells can promote the differentiation of M-MDSCs into highly immunosuppressive pro-tumorigenic macrophages (75).
Bone is one of the most important metastatic sites in breast cancer patients, with up to 70% of metastatic patients facing bone metastasis. MDSCs and monocytic cells play a major role in the formation of this metastatic site. Indeed, it has been demonstrated that MDSCs (defined as CD11b+Gr1+) cells can differentiate into osteoclasts in vitro and in vivo. These osteoclasts are capable of bone resorption (76, 77).
Immunosuppressive Myeloid Cells of the Granulocytic Lineage: Versatile Tumor-Promoting Functions in Breast Cancers
Granulocytic Cells in the Breast TME: Phenotypes, Functions and Controversies
Neutrophils constitute the more prominent leucocytes, primarily participating to the first lines of defense against infectious agents. They are produced in the bone marrow from granulocyte-monocyte myeloid progenitors (GMPs), which originate from lymphoid-primed multipotent progenitors, themselves derived from hematopoietic stem cells. Their maturation and differentiation depend on G-CSF and STAT3 activation.
In the context of cancer, and particularly in the context of breast cancers, it has been demonstrated that malignant cells could disrupt neutrophil homeostasis, hijacking their production and functions to their advantage through the production of TDFs such as G-CSF (78). The phenotypic characterization of tumor-associated neutrophils and the identification of specific subsets have remained the matter of intensive debates for the past few years. As mentioned in the previous section, the discrimination and possible relationship between tumor-associated neutrophils and polymorphonuclear (PMN)-MDSCs (MDSCs with a granulocyte phenotype) has remained an outstanding question in the field. Whether PMN-MDSC and tumor-associated neutrophils represent the same cell populations or are different subsets remains highly questionable. Indeed, in human, both are commonly identified as SSChigh, CD33+/medium, CD11bhigh, CD16+, CD15+, CD66b+, HLA-DRneg. Furthermore, many preclinical studies, primarily in mouse cancer models, on which most of our understandings of tumor-associated neutrophils have been based, do not clearly distinguish between neutrophils and PMN-MDSC. In these studies, cells with a CD11b+Gr1+Ly6G+LY6Cmed/low phenotype have been shown to expand over the course of cancer progression, and functional assays to assess their immunosuppressive properties have not always been performed (79, 80).. Although some markers have been proposed to discriminate between PMN-MDSC and “classical neutrophils” such as LOX-1 (81), CD84 and JAML (82), or sometimes the alarmins S100A8 or S100A9 (50), PMN-MDSC have mainly been defined by their immunosuppressive properties. Along these lines, because neutrophil density is higher than that of MDSC, it has also been proposed that density gradients may be used to separate physically tumor-associated neutrophils and tumor-induced PMN-MDSC. However, mature neutrophils have also been shown to exhibit immunosuppressive and pro-tumorigenic features in the TIME, and activated neutrophils fall in the “low-density” section of density gradients. Some authors also tried to establish a “N1/N2” dichotomy similar to that proposed for macrophages, N2 being pro-tumorigenic, immunosuppressive neutrophils (83). However, to date no reliable marker allow for a clear distinction of neutrophil different differentiation stages, and neutrophils in cancer are likely present as a heterogenous population, with cells at various activation states (84).
Recent reports have attempted to address this equivocal identity of PMN-MDSC and tumor-associated neutrophils. A single-cell transcriptomic analysis of the myeloid compartment in the splenocytes from two tumor-bearing mice (PyMT tumor model) and 2 tumor-free control animals has suggested that PMN-MDSCs differ from their normal myeloid counterparts and may originate from neutrophil progenitor cells undergoing an aberrant differentiation path (82). Data from another recent study have suggested that peripheral PMN-MDSCs from patients with metastatic breast cancer are more closely related to healthy donors’ neutrophils than to MDSCs induced in another pathological condition (Gram-positive sepsis) (85). Further mass-cytometry analysis revealed that unique subpopulations of these granulocytic cells were specifically present in cancer patients, with a majority of low density mature activated neutrophils and a minority of immature neutrophils lacking maturation markers (CD10, CD13, CD45) at different maturation stages. These cells were collectively referred to by the authors as “G-MDSCs” and proposed to constitute neutrophils at various differentiation stages (85). These data thus advocate for a differential differentiation and activation profile of neutrophils in the context of cancer.
Considering this significant phenotypic and functional overlap between so-called “PMN-MDSC” and “immunosuppressive neutrophils” we propose to refer to these cells more accurately using the term “immunosuppressive myeloid cells of the granulocytic lineage” or “IMCGL” until definitive phenotypic markers or functional assays are available to unequivocally distinguish them. (Figure 2). We believe that, compared to the terms “immunosuppressive neutrophils” or “PMN-MDSC”, the denomination ““IMCGL” allows to group these highly overlapping cell types to better study them and partly address the current controversies and challenges in distinguishing between immunosuppressive neutrophils and PMN-MDSC (Table 1). The physiological relevance and clinical usefulness of discriminating these cells also remains to be addressed.

Figure 2 Overlaps between subsets of tumor-promoting myeloid cells (monocytic vs gralulocytic origine). Cells formerly referred to as “Myeloid-Derived Suppressor Cells, MDSCs” encompass undifferentiated CD33+CD11b+ (“early-MDSCs”), immunosuppressive cells of the granulocytic lineage that we propose to call “immunosuppressive Myeloid Cell of the Granulocytic lineage, IMCGL” (“formal PMN-MDSCs”), and monocytes endowed with pro-tumoral properties. These tumor-promoting monocytes (“formal m-MDSCs”) can differentiate into pro-tumoral macrophages, Mo-dendritic cells or osteoclasts, each endowed with multiple dedicated tumor-promoting activities. IMCG, immunosuppressive myeloid cells from granulocytic lineage; e-MDSCs, early-myeloid-derived suppressor cells; Mo-Dendritic cell, monocyte-derived dendritic cells.
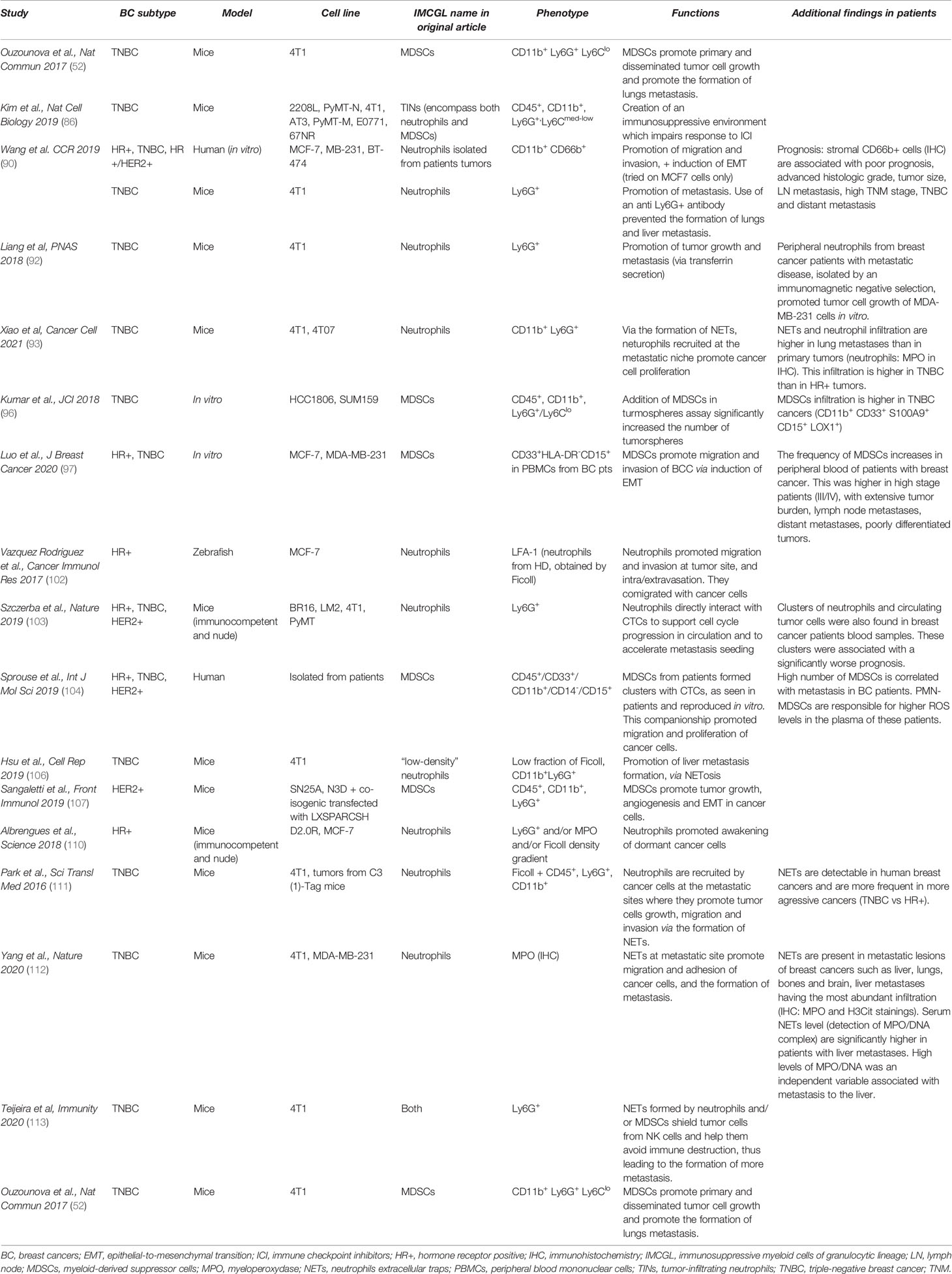
Table 1 IMCGL terminology, phenotype and non-immunological functions.
IMCGL in Breast Cancer Patients
In breast cancers, IMCGL have historically been considered as a major obstacle to anti-cancer immunity because of their immunosuppressive activities, a concept supported by recent findings in triple-negative breast cancer mouse models, where IMCGL-infiltrated tumors do not respond to immunotherapy (86). In breast cancer patients, tumor-induced expansion of circulating IMCGL has been corelated with a worst prognosis (87, 88). Interestingly, IMCGL are more frequently found in the tumor beds than in “healthy” adjacent tissues (59), and in most studies IMCGL have been associated with a higher tumor stage (89), or with a worse prognosis and an impaired response to chemotherapy (90). Tumor-induced IMCGL have been reported to impair T cell activation, particularly in advanced tumors, through increased production of ROS, NO or ARG1 or expression of immunoinhibitory ligands such as PDL1 (91).
IMCGL at the Primary Tumor Sites
IMCGL expansion and recruitment is directly promoted by breast cancer cells by various mechanisms. GM-CSF secretion by malignant cells induces the production of transferrin in Ly6G+ cells from 4T1-bearing mice that, in turn significantly enhances primary tumor growth in vivo and in vitro. In humans, the transferrin, TFR1 gene has been found to be up-regulated in breast cancers, and higher levels of this protein have been associated with higher tumor grades/stages, but also with a significantly worse survival (92). It has been shown that G-CSF production by breast cancer cells induces the recruitment of IMCGL that accumulate in the periphery of tumor-bearing PyMT mice (78). Cathepsin C (CTSC) produced by cancer cells has also been reported to induce recruitment and activation of IMCGL in 4T1 tumor bearing-mice. CTSC expression in human breast cancer is associated with metastasis and IMCGL occurrence (93). IMCGL recruitment can also be indirectly promoted by IL-1ß-secreting TAMs, which are recruited by Wnt ligands in p53-deficient cancer cells (94), or by cancer cell secretion of CCL2 (37).
Besides being equipped with immunosuppressive properties, IMCGL are also endowed with versatile tumor-promoting functions. Indeed, IMCGL can also induce and promote angiogenesis, participate to the remodeling of the extracellular matrix, contribute to tumor cell invasion, and participate to metastatic dissemination (84). IMCGL have also been shown to form Neutrophil Extracellular Traps (NETs) involved in tumor cell capture and growth as detailed hereafter (95).
IMCGL have been reported to display direct effects on breast cancer cells. Recent studies have indeed demonstrated that these cells participate to the acquisition of a stem-cell phenotype by malignant cells. Breast cancer cells expressing the ΔNp63 protein secrete CCL22 and CXCL2 that recruit IMCGL, which, in tumorsphere assays, promote the stemness phenotype of breast tumor cells via the secretion of CHI3L1 and MMP9 (96). Along these lines, it has also been reported that tumor-infiltrating IMCGL from breast cancer patients induce EMT in the MCF-7 cancer cell line, and promoted migration and invasion in in vitro assays (90). Consistent with these data, CCL3-recruited IMCGL have been observed to foster the EMT in breast cancer cells and enhance their proliferation, migratory and invasive properties through the PI3K/Akt/mTOR pathway (97).
IMCGL in the Metastatic Process
Most studies in the field have however shown that, primarily, IMCGL foster the process of breast cancer invasion and metastasis, with limited effects on primary tumor growth. Indeed, the elimination of these cells in pre-clinical models resulted mainly in dampening metastatic dissemination, with limited influence on primary tumor development (90, 94, 98–101). In zebrafish injected with MCF-7 cells, IMCGL from healthy human donors are capable of promoting cancer cell migration and intravasation at the tumor injection site in an Estradiol-dependent manner. In this model, IMCGL have been shown to migrate together with circulating cancer cells and to extravasate together in distant sites, thereby supporting disseminated tumor cell establishment in new metastatic niches (102). Two recent reports have brought substantial new insights into this joint migration of IMCGL with cancer cells. Indeed, evidence has emerged that some subpopulations of IMCGL can form companionship clusters with circulating tumor cells (CTC), and chaperone these CTC, protecting them in the circulating blood. Furthermore, IMCGL may foster CTC seeding at distant sites. CTC primarily circulate alone in the peripheral blood where the vast majority die in this environment. Only a limited number of these CTC (2-4%) have been detected as circulating homotypic or heterotypic clusters (103). These clusters have been associated with a significantly worse prognosis in breast cancer patients (103, 104). Heterotypic clusters are composed of CTC with white blood cells, most of them being myeloid cells of the granulocytic lineage (103, 104). Sprouse et al. have recently explored the cross-talks in these clusters between CTC and CD33+CD11b+CD15+ IMCGL cells (defined by the authors as PMC-MDSC) in the PBMC fraction of the peripheral blood (104). IMCGL induce upregulation of Notch1 receptor expression in CTCs through the ROS-NRF2-ARE axis, while CTCs induce pro-tumorigenic differentiation of IMCGL through paracrine Nodal signaling. Importantly, in mice, co-injection of breast cancer cells with IMCGL leads to an early dissemination of malignant cells to the lungs and brain (104). Szczerba et al. have also studied the interactions between CTC and IMCGL in heterotypic clusters from breast-cancer patients and from mammary tumor-bearing mice (103). Within these clusters, CTC exhibit a marked enrichment in positive regulators of cell cycle and DNA replication programs. Furthermore, RNA sequencing analysis has identified cytokines implicated in these cellular cross-talks and determined that IMCGL secrete TNFα, OSM, IL-1ß and IL-6, while CTC express CSF1, CSF3 (G-CSF), TGF-ß and IL-15 (103). However, the exact identity of each cellular partners within these clusters, the nature and importance of their interactions as it relates to the metastatic process, and the mechanisms underlying the promotion of CTC seeding and development at the metastatic niches remain to be fully uncovered. Along these lines, not only do IMCGL represent the main immune cells present at the metastatic site (105), but they also critically participate to the preparation of pre-metastatic niches. In tumor-bearing mice, myeloid cells accumulating in distant tissues are essentially composed of Ly6G+ immunosuppressive IMCGL (78). In 4T1 tumor-bearing mice, accumulation of IMCGL in the lungs (52) or in the liver (106) promotes metastatic cancer growth, and disseminated malignant cell proliferation. Importantly, depletion of IMCGL with an anti-Ly6G antibody suppresses metastasis in both studies (52, 106). It is noteworthy that although Ly6G has been used to deplete granulocytic cells, whether this approach results in the elimination of all IMCGL subsets remains to be determined.
Importance of NETs in Breast Cancer
Neutrophils have been described for their capacity of releasing neutrophil extra-cellular traps or “NETs”, a function that has also been described for PMN-MDSCs (107). NETosis is the process by which neutrophils release large web-like structures composed of cytosolic and granule proteins assembled on de-condensed chromatin. NETosis has been proposed to be a specific defense mechanism harbored by neutrophils against some pathogens like funga. The phenomenon of NETosis has also been observed in cancer where it can be triggered in part by G-CSF produced by many malignant cells (95). The impact of this process on cancer progression and on disease-associated complications such as thrombosis is being increasingly acknowledged (95, 108). A recent breakthrough in breast cancer was the findings that NETs may contribute to the awakening of dormant cancer cells. Reactivation of dormant cancer cells is of utmost importance in breast cancers, since half of patient relapses occur more than 5 years after the initial diagnosis, and in some cases even up to 20 years (109). Using breast cancer models that usually do not metastasize in mice, Albrengues et al. have demonstrated that neutrophil-derived NETs, induced by inflammatory conditions such as prolonged tobacco exposure or LPS instillations, lead to dormant cancer cell awakening and development into aggressive lungs metastases. In this setting, inflammation triggers NETs extrusion, which forms a scaffold allowing the sequential cleavage of laminin by neutrophil elastase (NE) and MMP9, as well as thrombospondin 1 (TSP-1). This laminin cleavage activates an α3β1-associated signaling in dormant cancer cells, leading to their reactivation (110). TSP-1 is a key matricellular protein that has been reported to inhibit metastasis. As outlined hereabove, CTSC secreted by cancer cells promotes the recruitment and activation of neutrophils in the metastatic niches, which upon activation form NETs that degrade the extracellular matrix, in part by cleavage of TSP-1, thereby allowing cancer cell proliferation and establishment (93). Other reports have indicated that in the context of breast cancers, NETs actively contribute to the formation of the pre-metastatic niches (111, 112). Indeed, these structures have been observed in the lungs of mice early after injection of 4T1 cells, thus before arrival and seeding of breast cancer cells. Furthermore, evidence has been provided that, in vitro, NETs stimulate invasion and migration of cancer cells. Consistently, NETs digestion with deoxyribonuclease I (DNase I) has been reported to significatively reduce the occurrence of lung metastasis (111). In humans, NETs have been detected in large amount in the metastatic lungs, and circulating NETs levels are higher in metastatic breast cancer patients compared to early-stage cancer patients (93). Suggesting a role of NETs in metastatic tumor cell organotropism regulation, Yang et al. have demonstrated that NETs contribute to the formation of metastases in the liver but not in the lungs. Furthermore, the authors have identified the protein CCDC25 expressed at the surface of cancer cells as a specific sensor of NETs DNA, and responsible for malignant cells migration, adhesion and proliferation induced by NETs. In breast cancer patients, CCDC25 has been detected in cancer cells with a clear membrane staining at the border of the tumor, and higher levels of CCDC25 in the primary tumors have correlated with a reduced survival (112).
Finally, NETs act as “shields” for cancer cells, wrapping them to avoid destruction by cytotoxic CD8 T cells of NK cells, adding to their multiple tumor-promoting functions (113).
Roles of “Tolerogenic/Regulatory” Dendritic Cells in Breast Cancer Development
Dendritic Cell Alterations in Breast Cancers
Dendritic cells (DC) play a central role in cancer immunosurveillance. They capture antigenic material from neoplastic cells, process tumor-specific antigens and present the derived peptides onto MHC class I or class II. Upon migration to the secondary lymphoid tissues, they activate effector tumor-specific CD8+ CTL and CD4+ Th lymphocytes. DC can also promote the anti-tumoral functions of NK, NKT and γδ T cells (114). However, in most cancers, DC are phenotypically and functionally impaired leading to dampened anti-cancer immunity (115). Although the nature of the microenvironment of breast cancers greatly varies depending on the tumor subtype and stage of the disease, in most cases it negatively influences DC capability to induce and sustain anti-tumor immunity. These DC alterations in breast malignancies have been attributed: a) to DC elimination; b) to the blockade of the generation of these cells from DC precursors; c) to the triggering of functional deficiency in DC (reduced antigen capture, processing, presentation and ability to activate T lymphocytes); and d) to the generation of immunosuppressive and tolerogenic DC capable of blocking anti-cancer T cells, inducing T lymphocyte anergy or inducing tumor-promoting regulatory T cells (Treg) (116–118). These defects are induced by different tumor-derived factors [extensively reviewed in (116, 117)], among which are VEGF (119), TGFβ (120), IL10 (121), PGE2 (122) or tumor-produced polyamines (123), and are responsible for a deficient induction of anti-cancer T lymphocyte proliferation and activation, thus contributing to breast cancer evasion from immunosurveillance.
In addition, different studies have demonstrated that breast malignancies are associated with the induction of different subpopulations of DC (myeloid, mDC or plasmacytoid, pDC) at different stages of maturation in the tumor, lymph nodes or blood, which actively promote T cell anergy and suppression and/or which trigger tumor-promoting Treg induction by a variety of mechanisms such as L-Arginine depletion (124), PD-L1 (125), TGFβ, IDO (126) or ICOS-ligand (127). Breast cancer-derived thymic stromal lymphopoietin (TSLP) has been identified as an inducer of OX40L on DC infiltrating primary breast cancer (128). These OX40L+ DC participate to the induction of IL-13- and TNF- producing Th2 cells thus contributing to the promotion of an environment permissible for breast tumor growth (128).
Role of DC in Breast Cancer Angiogenesis and Metastasis
Although many studies have extensively reported on the immune-modulatory role of tolerogenic/regulatory DC in breast cancers, much sparser reports are available as it relates to the tumor-promoting pro-angiogenic, pro-invasive and pro-metastatic properties of these cells. In this context, a study has correlated the presence of immature DC in highly angiogenic tumors (129), but the mechanistic bases underlying neoangiogenesis promotion remains to be determined. Similarly, the role of DC in breast cancer metastasis remains incompletely elucidated. A recent report indicates that CD303+ pDC accumulating in human breast cancer beds of patients with positive lymph nodes promote CXCR4 expression by cancer cells, suggesting that these tumor-associated pDC may participate to malignant cell metastasis to lymph nodes expressing SDF-1 through a CXCR4/SDF-1-dependent mechanism (130). Since DC conditioned by the tumor microenvironments can produce TGFβ, these cells may also contribute to the epithelial-mesenchymal transition (EMT) precluding tumor cell migration from primary tissues to metastatic sites, but a formal demonstration of this effect in breast cancer remains to be provided. Likewise, the possibility that DC may contribute to the preparation of pre-metastatic niches, before seeding of metastasizing cancer cells has yet to be formally demonstrated. In this context, a recent study has suggested that, in the mouse breast cancer model E0771, glucose-regulated protein 78 (GRP78) produced by tumor reduces DC MHC class II expression in the liver in the early stage of metastasis. However, the actual role of DC in the preparation of the pre-metastatic liver has not been demonstrated (131). Along these lines, recent data have also indicated that CD11c+ DC exposed to conditioned medium of RANKL+ T cells from the bone marrow of 4T1 mammary tumor-bearing mice can differentiate into osteoclast-like cells, suggesting that DC may participate to the osteolytic process occurring in metastatic breast cancer patients (132).
Conclusion, Perspectives and Challenges
The critical contribution of immune cells of the myeloid lineage to the mechanisms of cancer escape from immune detection and elimination is now widely recognized, and many studies have deciphered the various modes of action underlying the immunosuppressive properties of these cells. The notion that, beside this cardinal role in antitumor immunity, different myeloid cell populations are also endowed with a variety of “non-immunologic” tumor-promoting functions has drawn less scrutiny, until recently.
The heterogeneous nature of tumor-promoting myeloid cells, with some likely phenotypical and functional overlaps between subsets (illustrated in Figure 2) remains a major challenge preventing the unequivocal identification of distinct subpopulations. This current problem is probably best illustrated by the difficulty to draw a clear line between PMN-MDSC and immunosuppressive neutrophils in breast cancer patients or in mammary tumor models. Recent extensive studies, which have attempted to establish dedicated genomic, proteomic and biochemical profiles to better characterize these cells, have actually highlighted further the complexity and the high degree of plasticity of this myeloid landscape, advocating for instance that PMN-MDSC may actually correspond to neutrophils at different maturation stages in breast malignancies. These considerations have prompted us to refer to these tumor-promoting granulocytic cells occurring in cancer as “immunosuppressive myeloid cells of the granulocytic lineage, IMCGL”, which, we believe, better depicts their origins and functions. It would be clinically relevant to clearly decipher whether IMCGL are constituted of different subsets with dedicated properties and predictive or prognostic values. Better identification of these cells is a prerequisite to further determining whether they may serve as useful biomarkers and therapeutic targets, which warrants the urgent need to discover novel marker(s) and/or strategies allowing for a clear discrimination of the multiple subsets of these myeloid cells.
An additional outstanding question that still needs to be fully addressed relates to the “division of labor” among these tumor-promoting myeloid cell populations. As outlined in the previous section, it appears that all the main myeloid populations, TAM, MDSCs, IMCGL, and to some extend DC, are endowed with the capacity to exert many pro-tumoral activities. Recent single cell transcriptomic analysis suggest that, within each of these populations, a dedicated subset or even a single cell, may be equipped with concomitant multitasking activities (through the co-expression of factors involved for instance in immunosuppression, extracellular matrix remodeling, metastasis promotion…). The possibility that dedicated subsets or individual cells may sequentially acquire and lose one of these properties at a given time and depending on the nature of their environment and therefore on their location and on tumor stage is also conceivable, but remains to be clearly demonstrated. This functional plasticity of tumor-associated myeloid cells over time and space may be essential to fulfill the specific needs of growing tumors at each of the sequential stages of their development in the primary tumor sites (promotion of tumor growth, EMT, invasion, angiogenesis, intravasation, immunosuppression, production of chemokines involved in the recruitment of tumor-promoting cells), as CTC in the bloodstream (shielding in heterophilic clusters), and in the pre-metastatic and metastatic niches (soil preparation, ECM remodeling, extravasation, chemoattraction, immunosuppression).
Lastly, questions remain concerning the differences between breast cancer subtypes. Few studies have studied precisely the myeloid landscape and compared the different subtypes. Recent RNAseq findings show that myeloid infiltration is present in all main subtypes (“luminal” or HR positive BC, HER2+, TNBC) at different levels (133). A majority of studies discussed in this review focus on the TNBC subtype, some showing a higher infiltration of IMCGL in the tumors of these patients (90, 93, 96, 111). Macrophages are very represented across the different subtypes (133), though their role and exact phenotype in each subtype is unclear.
As these myeloid cells are essential contributors to many tumor-promoting networks, their therapeutic targeting (elimination, inactivation, reprogramming) has logically led to promising anti-tumor responses. However, the high phenotypic and functional heterogeneity and plasticity of these cells over time and depending on their tissue location has, to date, been a major hurdle for both their use as definitive biomarkers and the development of therapeutic strategies that would specifically interfere with their generation, development and multifaced tumor-promoting functions, which underlines the need to further characterize this myeloid landscape.
Author Contributions
CB and NL conceived and wrote the article. TB drew figures and wrote the article. FP and CD wrote the article. All authors contributed to the article and approved the submitted version.
Funding
Supported by the French national league against cancer and the SIRIC-BRIO (NL), and the Fondation ARC (CB).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Rondeau E, Larmonier N, Pradeu T, Bikfalvi A. Characterizing Causality in Cancer. Elife (2019) 8:1–6. doi: 10.7554/eLife.53755
2. Laplane L, Duluc D, Bikfalvi A, Larmonier N, Pradeu T. Beyond the Tumour Microenvironment. Int J Cancer (2019) 145(10):2611–8. doi: 10.1002/ijc.32343
3. Sharma P, Siddiqui BA, Anandhan S, Yadav SS, Subudhi SK, Gao J, et al. The Next Decade of Immune Checkpoint Therapy. Cancer Discov (2021) 11(4):838–57. doi: 10.1158/2159-8290.CD-20-1680
4. Schmid P, Cortes J, Pusztai L, McArthur H, Kummel S, Bergh J, et al. Pembrolizumab for Early Triple-Negative Breast Cancer. N Engl J Med (2020) 382(9):810–21. doi: 10.1056/NEJMoa1910549
5. Cortes J, Cescon DW, Rugo HS, Nowecki Z, Im SA, Yusof MM, et al. Pembrolizumab Plus Chemotherapy Versus Placebo Plus Chemotherapy for Previously Untreated Locally Recurrent Inoperable or Metastatic Triple-Negative Breast Cancer (KEYNOTE-355): A Randomised, Placebo-Controlled, Double-Blind, Phase 3 Clinical Trial. Lancet (2020) 396(10265):1817–28. doi: 10.1016/S0140-6736(20)32531-9
6. Vonderheide RH, Domchek SM, Clark AS. Immunotherapy for Breast Cancer: What Are We Missing? Clin Cancer Res (2017) 23(11):2640–6. doi: 10.1158/1078-0432.CCR-16-2569
7. Tekpli X, Lien T, Rossevold AH, Nebdal D, Borgen E, Ohnstad HO, et al. An Independent Poor-Prognosis Subtype of Breast Cancer Defined by a Distinct Tumor Immune Microenvironment. Nat Commun (2019) 10(1):5499. doi: 10.1038/s41467-019-13329-5
8. Klingen TA, Chen Y, Aas H, Wik E, Akslen LA. Tumor-Associated Macrophages are Strongly Related to Vascular Invasion, non-Luminal Subtypes, and Interval Breast Cancer. Hum Pathol (2017) 69:72–80. doi: 10.1016/j.humpath.2017.09.001
9. Qiu SQ, Waaijer SJH, Zwager MC, de Vries EGE, van der Vegt B, Schroder CP. Tumor-Associated Macrophages in Breast Cancer: Innocent Bystander or Important Player? Cancer Treat Rev (2018) 70:178–89. doi: 10.1016/j.ctrv.2018.08.010
10. Pelekanou V, Villarroel-Espindola F, Schalper KA, Pusztai L, Rimm DL. CD68, CD163, and Matrix Metalloproteinase 9 (MMP-9) Co-Localization in Breast Tumor Microenvironment Predicts Survival Differently in ER-Positive and -Negative Cancers. Breast Cancer Res (2018) 20(1):154. doi: 10.1186/s13058-018-1076-x
11. Zhang WJ, Wang XH, Gao ST, Chen C, Xu XY, Sun Q, et al. Tumor-Associated Macrophages Correlate With Phenomenon of Epithelial-Mesenchymal Transition and Contribute to Poor Prognosis in Triple-Negative Breast Cancer Patients. J Surg Res (2018) 222:93–101. doi: 10.1016/j.jss.2017.09.035
12. Ali HR, Chlon L, Pharoah PD, Markowetz F, Caldas C. Patterns of Immune Infiltration in Breast Cancer and Their Clinical Implications: A Gene-Expression-Based Retrospective Study. PloS Med (2016) 13(12):e1002194. doi: 10.1371/journal.pmed.1002194
13. Mosser DM, Edwards JP. Exploring the Full Spectrum of Macrophage Activation. Nat Rev Immunol (2008) 8(12):958–69. doi: 10.1038/nri2448
14. Murray PJ. Macrophage Polarization. Annu Rev Physiol (2017) 79:541–66. doi: 10.1146/annurev-physiol-022516-034339
15. Azizi E, Carr AJ, Plitas G, Cornish AE, Konopacki C, Prabhakaran S, et al. Single-Cell Map of Diverse Immune Phenotypes in the Breast Tumor Microenvironment. Cell (2018) 174(5):1293–308.e36. doi: 10.1016/j.cell.2018.05.060
16. Wagner J, Rapsomaniki MA, Chevrier S, Anzeneder T, Langwieder C, Dykgers A, et al. A Single-Cell Atlas of the Tumor and Immune Ecosystem of Human Breast Cancer. Cell (2019) 177(5):1330–45.e18. doi: 10.1016/j.cell.2019.03.005
17. Cassetta L, Fragkogianni S, Sims AH, Swierczak A, Forrester LM, Zhang H, et al. Human Tumor-Associated Macrophage and Monocyte Transcriptional Landscapes Reveal Cancer-Specific Reprogramming, Biomarkers, and Therapeutic Targets. Cancer Cell (2019) 35(4):588–602.e10. doi: 10.1016/j.ccell.2019.02.009
18. Williams CB, Yeh ES, Soloff AC. Tumor-Associated Macrophages: Unwitting Accomplices in Breast Cancer Malignancy. NPJ Breast Cancer (2016) 2:1–12. doi: 10.1038/npjbcancer.2015.25
19. Bent R, Moll L, Grabbe S, Bros M. Interleukin-1 Beta-A Friend or Foe in Malignancies? Int J Mol Sci (2018) 19(8):1–34. doi: 10.3390/ijms19082155
20. Mantovani A, Barajon I, Garlanda C. IL-1 and IL-1 Regulatory Pathways in Cancer Progression and Therapy. Immunol Rev (2018) 281(1):57–61. doi: 10.1111/imr.12614
21. Racioppi L, Nelson ER, Huang W, Mukherjee D, Lawrence SA, Lento W, et al. CaMKK2 in Myeloid Cells is a Key Regulator of the Immune-Suppressive Microenvironment in Breast Cancer. Nat Commun (2019) 10(1):2450. doi: 10.1038/s41467-019-10424-5
22. Seoane S, Martinez-Ordonez A, Eiro N, Cabezas-Sainz P, Garcia-Caballero L, Gonzalez LO, et al. POU1F1 Transcription Factor Promotes Breast Cancer Metastasis via Recruitment and Polarization of Macrophages. J Pathol (2019) 249(3):381–94. doi: 10.1002/path.5324
23. Donzelli S, Milano E, Pruszko M, Sacconi A, Masciarelli S, Iosue I, et al. Expression of ID4 Protein in Breast Cancer Cells Induces Reprogramming of Tumour-Associated Macrophages. Breast Cancer Res (2018) 20(1):59. doi: 10.1186/s13058-018-0990-2
24. Valeta-Magara A, Gadi A, Volta V, Walters B, Arju R, Giashuddin S, et al. Inflammatory Breast Cancer Promotes Development of M2 Tumor-Associated Macrophages and Cancer Mesenchymal Cells Through a Complex Chemokine Network. Cancer Res (2019) 79(13):3360–71. doi: 10.1158/0008-5472.CAN-17-2158
25. Shrivastava R, Asif M, Singh V, Dubey P, Ahmad Malik S, Lone MU, et al. M2 Polarization of Macrophages by Oncostatin M in Hypoxic Tumor Microenvironment is Mediated by Mtorc2 and Promotes Tumor Growth and Metastasis. Cytokine (2019) 118:130–43. doi: 10.1016/j.cyto.2018.03.032
26. Wang L, Li YS, Yu LG, Zhang XK, Zhao L, Gong FL, et al. Galectin-3 Expression and Secretion by Tumor-Associated Macrophages in Hypoxia Promotes Breast Cancer Progression. Biochem Pharmacol (2020) 178:114113. doi: 10.1016/j.bcp.2020.114113
27. Trenti A, Tedesco S, Boscaro C, Trevisi L, Bolego C, Cignarella A. Estrogen, Angiogenesis, Immunity and Cell Metabolism: Solving the Puzzle. Int J Mol Sci (2018) 19(3):2–16. doi: 10.3390/ijms19030859
28. Svensson S, Abrahamsson A, Rodriguez GV, Olsson AK, Jensen L, Cao Y, et al. CCL2 and CCL5 Are Novel Therapeutic Targets for Estrogen-Dependent Breast Cancer. Clin Cancer Res (2015) 21(16):3794–805. doi: 10.1158/1078-0432.CCR-15-0204
29. Dittmer J. Breast Cancer Stem Cells: Features, Key Drivers and Treatment Options. Semin Cancer Biol (2018) 53:59–74. doi: 10.1016/j.semcancer.2018.07.007
30. Gok Yavuz B, Gunaydin G, Gedik ME, Kosemehmetoglu K, Karakoc D, Ozgur F, et al. Cancer Associated Fibroblasts Sculpt Tumour Microenvironment by Recruiting Monocytes and Inducing Immunosuppressive PD-1(+) TAMs. Sci Rep (2019) 9(1):3172. doi: 10.1038/s41598-019-39553-z
31. Liu D, Lu Q, Wang X, Wang J, Lu N, Jiang Z, et al. LSECtin on Tumor-Associated Macrophages Enhances Breast Cancer Stemness via Interaction With its Receptor BTN3A3. Cell Res (2019) 29(5):365–78. doi: 10.1038/s41422-019-0155-6
32. Lu H, Clauser KR, Tam WL, Frose J, Ye X, Eaton EN, et al. A Breast Cancer Stem Cell Niche Supported by Juxtacrine Signalling From Monocytes and Macrophages. Nat Cell Biol (2014) 16(11):1105–17. doi: 10.1038/ncb3041
33. Tiwari P, Blank A, Cui C, Schoenfelt KQ, Zhou G, Xu Y, et al. Metabolically Activated Adipose Tissue Macrophages Link Obesity to Triple-Negative Breast Cancer. J Exp Med (2019) 216(6):1345–58. doi: 10.1084/jem.20181616
34. Harney AS, Arwert EN, Entenberg D, Wang Y, Guo P, Qian BZ, et al. Real-Time Imaging Reveals Local, Transient Vascular Permeability, and Tumor Cell Intravasation Stimulated by TIE2hi Macrophage-Derived VEGFA. Cancer Discovery (2015) 5(9):932–43. doi: 10.1158/2159-8290.CD-15-0012
35. Chen Y, Zhang S, Wang Q, Zhang X. Tumor-Recruited M2 Macrophages Promote Gastric and Breast Cancer Metastasis via M2 Macrophage-Secreted CHI3L1 Protein. J Hematol Oncol (2017) 10(1):36. doi: 10.1186/s13045-017-0408-0
36. Little AC, Pathanjeli P, Wu Z, Bao L, Goo LE, Yates JA, et al. IL-4/IL-13 Stimulated Macrophages Enhance Breast Cancer Invasion Via Rho-GTPase Regulation of Synergistic VEGF/CCL-18 Signaling. Front Oncol (2019) 9:456. doi: 10.3389/fonc.2019.00456
37. Kersten K, Coffelt SB, Hoogstraat M, Verstegen NJM, Vrijland K, Ciampricotti M, et al. Mammary Tumor-Derived CCL2 Enhances Pro-Metastatic Systemic Inflammation Through Upregulation of IL1beta in Tumor-Associated Macrophages. Oncoimmunology (2017) 6(8):e1334744. doi: 10.1080/2162402X.2017.1334744
38. Linde N, Casanova-Acebes M, Sosa MS, Mortha A, Rahman A, Farias E, et al. Macrophages Orchestrate Breast Cancer Early Dissemination and Metastasis. Nat Commun (2018) 9(1):21. doi: 10.1038/s41467-017-02481-5
39. Chen XW, Yu TJ, Zhang J, Li Y, Chen HL, Yang GF, et al. CYP4A in Tumor-Associated Macrophages Promotes Pre-Metastatic Niche Formation and Metastasis. Oncogene (2017) 36(35):5045–57. doi: 10.1038/onc.2017.118
40. Celus W, Di Conza G, Oliveira AI, Ehling M, Costa BM, Wenes M, et al. Loss of Caveolin-1 in Metastasis-Associated Macrophages Drives Lung Metastatic Growth Through Increased Angiogenesis. Cell Rep (2017) 21(10):2842–54. doi: 10.1016/j.celrep.2017.11.034
41. Bieniasz-Krzywiec P, Martin-Perez R, Ehling M, Garcia-Caballero M, Pinioti S, Pretto S, et al. Podoplanin-Expressing Macrophages Promote Lymphangiogenesis and Lymphoinvasion in Breast Cancer. Cell Metab (2019) 30(5):917–936 e10. doi: 10.1016/j.cmet.2019.07.015
42. Weichand B, Popp R, Dziumbla S, Mora J, Strack E, Elwakeel E, et al. S1PR1 on Tumor-Associated Macrophages Promotes Lymphangiogenesis and Metastasis via NLRP3/IL-1beta. J Exp Med (2017) 214(9):2695–713. doi: 10.1084/jem.20160392
43. Storr SJ, Safuan S, Ahmad N, El-Refaee M, Jackson AM, Martin SG. Macrophage-Derived Interleukin-1beta Promotes Human Breast Cancer Cell Migration and Lymphatic Adhesion In Vitro. Cancer Immunol Immunother (2017) 66(10):1287–94. doi: 10.1007/s00262-017-2020-0
44. Gabrilovich DI, Bronte V, Chen SH, Colombo MP, Ochoa A, Ostrand-Rosenberg S, et al. The Terminology Issue for Myeloid-Derived Suppressor Cells. Cancer Res (2007) 67(1):425.doi: 10.1158/0008-5472.CAN-06-3037
45. Gabrilovich DI, Ostrand-Rosenberg S, Bronte V. Coordinated Regulation of Myeloid Cells by Tumours. Nat Rev Immunol (2012) 12(4):253–68. doi: 10.1038/nri3175
46. Condamine T, Ramachandran I, Youn JI, Gabrilovich DI. Regulation of Tumor Metastasis by Myeloid-Derived Suppressor Cells. Annu Rev Med (2015) 66:97–110. doi: 10.1146/annurev-med-051013-052304
47. Liu QW, Chen Y, Li JY, Xiao L, Zhang WJ, Zhao JL, et al. Bone Marrow Cells are Differentiated Into MDSCs by BCC-Ex Through Down-Regulating the Expression of CXCR4 and Activating STAT3 Signalling Pathway. J Cell Mol Med (2021) 25(12):5497–510. doi: 10.1111/jcmm.16559
48. Keklikoglou I, Cianciaruso C, Guc E, Squadrito ML, Spring LM, Tazzyman S, et al. Chemotherapy Elicits Pro-Metastatic Extracellular Vesicles in Breast Cancer Models. Nat Cell Biol (2019) 21(2):190–202. doi: 10.1038/s41556-018-0256-3
49. Ouyang L, Chang W, Fang B, Qin J, Qu X, Cheng F. Estrogen-Induced SDF-1alpha Production Promotes the Progression of ER-Negative Breast Cancer via the Accumulation of MDSCs in the Tumor Microenvironment. Sci Rep (2016) 6:39541. doi: 10.1038/srep39541
50. Bronte V, Brandau S, Chen SH, Colombo MP, Frey AB, Greten TF, et al. Recommendations for Myeloid-Derived Suppressor Cell Nomenclature and Characterization Standards. Nat Commun (2016) 7:12150. doi: 10.1038/ncomms12150
51. Veglia F, Perego M, Gabrilovich D. Myeloid-Derived Suppressor Cells Coming of Age. Nat Immunol (2018) 19(2):108–19. doi: 10.1038/s41590-017-0022-x
52. Ouzounova M, Lee E, Piranlioglu R, El Andaloussi A, Kolhe R, Demirci MF, et al. Monocytic and Granulocytic Myeloid Derived Suppressor Cells Differentially Regulate Spatiotemporal Tumour Plasticity During Metastatic Cascade. Nat Commun (2017) 8:14979. doi: 10.1038/ncomms14979
53. Thomas GD, Hamers AAJ, Nakao C, Marcovecchio P, Taylor AM, McSkimming C, et al. Human Blood Monocyte Subsets: A New Gating Strategy Defined Using Cell Surface Markers Identified by Mass Cytometry. Arterioscler Thromb Vasc Biol (2017) 37(8):1548–58. doi: 10.1161/ATVBAHA.117.309145
54. Bergenfelz C, Larsson AM, von Stedingk K, Gruvberger-Saal S, Aaltonen K, Jansson S, et al. Systemic Monocytic-MDSCs Are Generated From Monocytes and Correlate With Disease Progression in Breast Cancer Patients. PloS One (2015) 10(5):e0127028. doi: 10.1371/journal.pone.0127028
55. Jung K, Heishi T, Incio J, Huang Y, Beech EY, Pinter M, et al. Targeting CXCR4-Dependent Immunosuppressive Ly6C(low) Monocytes Improves Antiangiogenic Therapy in Colorectal Cancer. Proc Natl Acad Sci U.S.A. (2017) 114(39):10455–60. doi: 10.1073/pnas.1710754114
56. Jung K, Heishi T, Khan OF, Kowalski PS, Incio J, Rahbari NN, et al. Ly6Clo Monocytes Drive Immunosuppression and Confer Resistance to Anti-VEGFR2 Cancer Therapy. J Clin Invest (2017) 127(8):3039–51. doi: 10.1172/JCI93182
57. Jeong J, Suh Y, Jung K. Context Drives Diversification of Monocytes and Neutrophils in Orchestrating the Tumor Microenvironment. Front Immunol (2019) 10:1817. doi: 10.3389/fimmu.2019.01817
58. Jeong J, Kim DK, Park JH, Park DJ, Lee HJ, Yang HK, et al. Tumor-Infiltrating Neutrophils and Non-Classical Monocytes May Be Potential Therapeutic Targets for HER2(negative) Gastric Cancer. Immune Netw (2021) 21(4):e31. doi: 10.4110/in.2021.21.e31
59. Toor SM, Syed Khaja AS, El Salhat H, Faour I, Kanbar J, Quadri AA, et al. Myeloid Cells in Circulation and Tumor Microenvironment of Breast Cancer Patients. Cancer Immunol Immunother (2017) 66(6):753–64. doi: 10.1007/s00262-017-1977-z
60. Safarzadeh E, Mohammadi A, Mansoori B, Duijf PHG, Hashemzadeh S, Khaze V, et al. STAT3 Silencing and TLR7/8 Pathway Activation Repolarize and Suppress Myeloid-Derived Suppressor Cells From Breast Cancer Patients. Front Immunol (2020) 11:613215. doi: 10.3389/fimmu.2020.613215
61. Wang PF, Song SY, Wang TJ, Ji WJ, Li SW, Liu N, et al. Prognostic Role of Pretreatment Circulating MDSCs in Patients With Solid Malignancies: A Meta-Analysis of 40 Studies. Oncoimmunology (2018) 7(10):e1494113. doi: 10.1080/2162402X.2018.1494113
62. Salvador-Coloma C, Santaballa A, Sanmartin E, Calvo D, Garcia A, Hervas D, et al. Immunosuppressive Profiles in Liquid Biopsy at Diagnosis Predict Response to Neoadjuvant Chemotherapy in Triple-Negative Breast Cancer. Eur J Cancer (2020) 139:119–34. doi: 10.1016/j.ejca.2020.08.020
63. Yu J, Du W, Yan F, Wang Y, Li H, Cao S, et al. Myeloid-Derived Suppressor Cells Suppress Antitumor Immune Responses Through IDO Expression and Correlate With Lymph Node Metastasis in Patients With Breast Cancer. J Immunol (2013) 190(7):3783–97. doi: 10.4049/jimmunol.1201449
64. Peng D, Tanikawa T, Li W, Zhao L, Vatan L, Szeliga W, et al. Myeloid-Derived Suppressor Cells Endow Stem-Like Qualities to Breast Cancer Cells Through IL6/STAT3 and NO/NOTCH Cross-Talk Signaling. Cancer Res (2016) 76(11):3156–65. doi: 10.1158/0008-5472.CAN-15-2528
65. Bauer R, Udonta F, Wroblewski M, Ben-Batalla I, Santos IM, Taverna F, et al. Blockade of Myeloid-Derived Suppressor Cell Expansion With All-Trans Retinoic Acid Increases the Efficacy of Antiangiogenic Therapy. Cancer Res (2018) 78(12):3220–32. doi: 10.1158/0008-5472.CAN-17-3415
66. Liu L, Yang L, Yan W, Zhai J, Pizzo DP, Chu P, et al. Chemotherapy Induces Breast Cancer Stemness in Association With Dysregulated Monocytosis. Clin Cancer Res (2018) 24(10):2370–82. doi: 10.1158/1078-0432.CCR-17-2545
67. Eisenblaetter M, Flores-Borja F, Lee JJ, Wefers C, Smith H, Hueting R, et al. Visualization of Tumor-Immune Interaction - Target-Specific Imaging of S100A8/A9 Reveals Pre-Metastatic Niche Establishment. Theranostics (2017) 7(9):2392–401. doi: 10.7150/thno.17138
68. Yan HH, Pickup M, Pang Y, Gorska AE, Li Z, Chytil A, et al. Gr-1+CD11b+ Myeloid Cells Tip the Balance of Immune Protection to Tumor Promotion in the Premetastatic Lung. Cancer Res (2010) 70(15):6139–49. doi: 10.1158/0008-5472.CAN-10-0706
69. Koelwyn GJ, Newman AAC, Afonso MS, van Solingen C, Corr EM, Brown EJ, et al. Myocardial Infarction Accelerates Breast Cancer via Innate Immune Reprogramming. Nat Med (2020) 26(9):1452–8. doi: 10.1038/s41591-020-0964-7
70. Spiegel A, Brooks MW, Houshyar S, Reinhardt F, Ardolino M, Fessler E, et al. Neutrophils Suppress Intraluminal NK Cell-Mediated Tumor Cell Clearance and Enhance Extravasation of Disseminated Carcinoma Cells. Cancer Discovery (2016) 6(6):630–49. doi: 10.1158/2159-8290.CD-15-1157
71. Hiratsuka S, Duda DG, Huang Y, Goel S, Sugiyama T, Nagasawa T, et al. C-X-C Receptor Type 4 Promotes Metastasis by Activating P38 Mitogen-Activated Protein Kinase in Myeloid Differentiation Antigen (Gr-1)-Positive Cells. Proc Natl Acad Sci U.S.A. (2011) 108(1):302–7. doi: 10.1073/pnas.1016917108
72. Ghouse SM, Vadrevu SK, Manne S, Reese B, Patel J, Patel B, et al. Therapeutic Targeting of Vasculature in the Premetastatic and Metastatic Niches Reduces Lung Metastasis. J Immunol (2020) 204(4):990–1000. doi: 10.4049/jimmunol.1901208
73. Hsu YL, Yen MC, Chang WA, Tsai PH, Pan YC, Liao SH, et al. CXCL17-Derived CD11b(+)Gr-1(+) Myeloid-Derived Suppressor Cells Contribute to Lung Metastasis of Breast Cancer Through Platelet-Derived Growth Factor-BB. Breast Cancer Res (2019) 21(1):23. doi: 10.1186/s13058-019-1114-3
74. Kitamura T, Doughty-Shenton D, Cassetta L, Fragkogianni S, Brownlie D, Kato Y, et al. Monocytes Differentiate to Immune Suppressive Precursors of Metastasis-Associated Macrophages in Mouse Models of Metastatic Breast Cancer. Front Immunol (2017) 8:2004. doi: 10.3389/fimmu.2017.02004
75. Biswas S, Mandal G, Roy Chowdhury S, Purohit S, Payne KK, Anadon C, et al. Exosomes Produced by Mesenchymal Stem Cells Drive Differentiation of Myeloid Cells Into Immunosuppressive M2-Polarized Macrophages in Breast Cancer. J Immunol (2019) 203(12):3447–60. doi: 10.4049/jimmunol.1900692
76. Sawant A, Deshane J, Jules J, Lee CM, Harris BA, Feng X, et al. Myeloid-Derived Suppressor Cells Function as Novel Osteoclast Progenitors Enhancing Bone Loss in Breast Cancer. Cancer Res (2013) 73(2):672–82. doi: 10.1158/0008-5472.CAN-12-2202
77. Danilin S, Merkel AR, Johnson JR, Johnson RW, Edwards JR, Sterling JA. Myeloid-Derived Suppressor Cells Expand During Breast Cancer Progression and Promote Tumor-Induced Bone Destruction. Oncoimmunology (2012) 1(9):1484–94. doi: 10.4161/onci.21990
78. Casbon AJ, Reynaud D, Park C, Khuc E, Gan DD, Schepers K, et al. Invasive Breast Cancer Reprograms Early Myeloid Differentiation in the Bone Marrow to Generate Immunosuppressive Neutrophils. Proc Natl Acad Sci U.S.A. (2015) 112(6):E566–75. doi: 10.1073/pnas.1424927112
79. Gustafson MP, Lin Y, Maas ML, Van Keulen VP, Johnston PB, Peikert T, et al. A Method for Identification and Analysis of non-Overlapping Myeloid Immunophenotypes in Humans. PloS One (2015) 10(3):e0121546. doi: 10.1371/journal.pone.0121546
80. Lakschevitz FS, Hassanpour S, Rubin A, Fine N, Sun C, Glogauer M. Identification of Neutrophil Surface Marker Changes in Health and Inflammation Using High-Throughput Screening Flow Cytometry. Exp Cell Res (2016) 342(2):200–9. doi: 10.1016/j.yexcr.2016.03.007
81. Condamine T, Dominguez GA, Youn JI, Kossenkov AV, Mony S, Alicea-Torres K, et al. Lectin-Type Oxidized LDL Receptor-1 Distinguishes Population of Human Polymorphonuclear Myeloid-Derived Suppressor Cells in Cancer Patients. Sci Immunol (2016) 1(2):1–32. doi: 10.1126/sciimmunol.aaf8943
82. Alshetaiwi H, Pervolarakis N, McIntyre LL, Ma D, Nguyen Q, Rath JA, et al. Defining the Emergence of Myeloid-Derived Suppressor Cells in Breast Cancer Using Single-Cell Transcriptomics. Sci Immunol (2020) 5(44):1–28. doi: 10.1126/sciimmunol.aay6017
83. Fridlender ZG, Sun J, Kim S, Kapoor V, Cheng G, Ling L, et al. Polarization of Tumor-Associated Neutrophil Phenotype by TGF-Beta: “N1” Versus “N2” TAN. Cancer Cell (2009) 16(3):183–94. doi: 10.1016/j.ccr.2009.06.017
84. Coffelt SB, Wellenstein MD, de Visser KE. Neutrophils in Cancer: Neutral No More. Nat Rev Cancer (2016) 16(7):431–46. doi: 10.1038/nrc.2016.52
85. Mehmeti-Ajradini M, Bergenfelz C, Larsson AM, Carlsson R, Riesbeck K, Ahl J, et al. Human G-MDSCs are Neutrophils at Distinct Maturation Stages Promoting Tumor Growth in Breast Cancer. Life Sci Alliance (2020) 3(11):1–19. doi: 10.26508/lsa.202000893
86. Kim IS, Gao Y, Welte T, Wang H, Liu J, Janghorban M, et al. Immuno-Subtyping of Breast Cancer Reveals Distinct Myeloid Cell Profiles and Immunotherapy Resistance Mechanisms. Nat Cell Biol (2019) 21(9):1113–26. doi: 10.1038/s41556-019-0373-7
87. Huszno J, Kolosza. Prognostic Value of the Neutrophil-Lymphocyte, Platelet-Lymphocyte and Monocyte-Lymphocyte Ratio in Breast Cancer Patients. Oncol Lett (2019) 18(6):6275–83. doi: 10.3892/ol.2019.10966
88. Ethier JL, Desautels D, Templeton A, Shah PS, Amir E. Prognostic Role of Neutrophil-To-Lymphocyte Ratio in Breast Cancer: A Systematic Review and Meta-Analysis. Breast Cancer Res: BCR (2017) 19(1):1–13. doi: 10.1186/s13058-016-0794-1
89. Wu L, Saxena S, Goel P, Prajapati DR, Wang C, Singh RK. Breast Cancer Cell-Neutrophil Interactions Enhance Neutrophil Survival and Pro-Tumorigenic Activities. Cancers (Basel) (2020) 12(10):1–18. doi: 10.3390/cancers12102884
90. Wang Y, Chen J, Yang L, Li J, Wu W, Huang M, et al. Tumor-Contacted Neutrophils Promote Metastasis by a CD90-TIMP-1 Juxtacrine-Paracrine Loop. Clin Cancer Res (2019) 25(6):1957–69. doi: 10.1158/1078-0432.CCR-18-2544
91. Jaillon S, Ponzetta A, Di Mitri D, Santoni A, Bonecchi R, Mantovani A. Neutrophil Diversity and Plasticity in Tumour Progression and Therapy. Nat Rev Cancer (2020) 20(9):485–503. doi: 10.1038/s41568-020-0281-y
92. Liang W, Li Q, Ferrara N. Metastatic Growth Instructed by Neutrophil-Derived Transferrin. Proc Natl Acad Sci USA (2018) 115(43):11060–5. doi: 10.1073/pnas.1811717115
93. Xiao Y, Cong M, Li J, He D, Wu Q, Tian P, et al. Cathepsin C Promotes Breast Cancer Lung Metastasis by Modulating Neutrophil Infiltration and Neutrophil Extracellular Trap Formation. Cancer Cell (2020) 39(3):423–37.e7. doi: 10.1016/j.ccell.2020.12.012
94. Wellenstein MD, Coffelt SB, Duits DEM, van Miltenburg MH, Slagter M, de Rink I, et al. Loss of P53 Triggers WNT-Dependent Systemic Inflammation to Drive Breast Cancer Metastasis. Nature (2019) 572(7770):538–42. doi: 10.1038/s41586-019-1450-6
95. Papayannopoulos V. Neutrophil Extracellular Traps in Immunity and Disease. Nat Rev Immunol (2018) 18(2):134–47. doi: 10.1038/nri.2017.105
96. Kumar S, Wilkes DW, Samuel N, Blanco MA, Nayak A, Alicea-Torres K, et al. DeltaNp63-Driven Recruitment of Myeloid-Derived Suppressor Cells Promotes Metastasis in Triple-Negative Breast Cancer. J Clin Invest (2018) 128(11):5095–109. doi: 10.1172/JCI99673
97. Luo A, Meng M, Wang G, Han R, Zhang Y, Jing X, et al. Myeloid-Derived Suppressor Cells Recruited by Chemokine (C-C Motif) Ligand 3 Promote the Progression of Breast Cancer via Phosphoinositide 3-Kinase-Protein Kinase B-Mammalian Target of Rapamycin Signaling. J Breast Cancer (2020) 23(2):141–61. doi: 10.4048/jbc.2020.23.e26
98. Janiszewska M, Tabassum DP, Castano Z, Cristea S, Yamamoto KN, Kingston NL, et al. Subclonal Cooperation Drives Metastasis by Modulating Local and Systemic Immune Microenvironments. Nat Cell Biol (2019) 21(7):879–88. doi: 10.1038/s41556-019-0346-x
99. Robichaud N, Hsu BE, Istomine R, Alvarez F, Blagih J, Ma EH, et al. Translational Control in the Tumor Microenvironment Promotes Lung Metastasis: Phosphorylation of Eif4e in Neutrophils. Proc Natl Acad Sci U.S.A. (2018) 115(10):E2202–9. doi: 10.1073/pnas.1717439115
100. Coffelt SB, Kersten K, Doornebal CW, Weiden J, Vrijland K, Hau CS, et al. IL-17-Producing γδ T Cells and Neutrophils Conspire to Promote Breast Cancer Metastasis. Nature (2015) 522(7556):345–8. doi: 10.1038/nature14282
101. Li P, Lu M, Shi J, Hua L, Gong Z, Li Q, et al. Dual Roles of Neutrophils in Metastatic Colonization are Governed by the Host NK Cell Status. Nat Commun (2020) 11(1):4387. doi: 10.1038/s41467-020-18125-0
102. Vazquez Rodriguez G, Abrahamsson A, Jensen LD, Dabrosin C. Estradiol Promotes Breast Cancer Cell Migration via Recruitment and Activation of Neutrophils. Cancer Immunol Res (2017) 5(3):234–47. doi: 10.1158/2326-6066.CIR-16-0150
103. Szczerba BM, Castro-Giner F, Vetter M, Krol I, Gkountela S, Landin J, et al. Neutrophils Escort Circulating Tumour Cells to Enable Cell Cycle Progression. Nature (2019) 566(7745):553–7. doi: 10.1038/s41586-019-0915-y
104. Sprouse ML, Welte T, Boral D, Liu HN, Yin W, Vishnoi M, et al. PMN-MDSCs Enhance CTC Metastatic Properties Through Reciprocal Interactions via ROS/Notch/Nodal Signaling. Int J Mol Sci (2019) 20(8):1–20. doi: 10.3390/ijms20081916
105. Ombrato L, Nolan E, Kurelac I, Mavousian A, Bridgeman VL, Heinze I, et al. Metastatic-Niche Labelling Reveals Parenchymal Cells With Stem Features. Nature (2019) 572(7771):603–8. doi: 10.1038/s41586-019-1487-6
106. Hsu BE, Tabaries S, Johnson RM, Andrzejewski S, Senecal J, Lehuede C, et al. Immature Low-Density Neutrophils Exhibit Metabolic Flexibility That Facilitates Breast Cancer Liver Metastasis. Cell Rep (2019) 27(13):3902–3915 e6. doi: 10.1016/j.celrep.2019.05.091
107. Sangaletti S, Talarico G, Chiodoni C, Cappetti B, Botti L, Portararo P, et al. SPARC Is a New Myeloid-Derived Suppressor Cell Marker Licensing Suppressive Activities. Front Immunol (2019) 10:1369. doi: 10.3389/fimmu.2019.01369
108. Snoderly HT, Boone BA, Bennewitz MF. Neutrophil Extracellular Traps in Breast Cancer and Beyond: Current Perspectives on NET Stimuli, Thrombosis and Metastasis, and Clinical Utility for Diagnosis and Treatment. Breast Cancer Res (2019) 21(1):145. doi: 10.1186/s13058-019-1237-6
109. Pan H, Gray R, Braybrooke J, Davies C, Taylor C, McGale P, et al. 20-Year Risks of Breast-Cancer Recurrence After Stopping Endocrine Therapy at 5 Years. N Engl J Med (2017) 377(19):1836–46. doi: 10.1056/NEJMoa1701830
110. Albrengues J, Shields MA, Ng D, Park CG, Ambrico A, Poindexter ME, et al. Neutrophil Extracellular Traps Produced During Inflammation Awaken Dormant Cancer Cells in Mice. Science (2018) 361(6409):1–30. doi: 10.1126/science.aao4227
111. Park J, Wysocki RW, Amoozgar Z, Maiorino L, Fein MR, Jorns J, et al. Cancer Cells Induce Metastasis-Supporting Neutrophil Extracellular DNA Traps. Sci Transl Med (2016) 8(361):361ra138. doi: 10.1126/scitranslmed.aag1711
112. Yang L, Liu Q, Zhang X, Liu X, Zhou B, Chen J, et al. DNA of Neutrophil Extracellular Traps Promotes Cancer Metastasis via CCDC25. Nature (2020) 583(7814):133–8. doi: 10.1038/s41586-020-2394-6
113. Teijeira A, Garasa S, Gato M, Alfaro C, Migueliz I, Cirella A, et al. CXCR1 and CXCR2 Chemokine Receptor Agonists Produced by Tumors Induce Neutrophil Extracellular Traps That Interfere With Immune Cytotoxicity. Immunity (2020) 52(5):856–871 e8. doi: 10.1016/j.immuni.2020.03.001
114. Hanke N, Alizadeh D, Katsanis E, Larmonier N. Dendritic Cell Tumor Killing Activity and its Potential Applications in Cancer Immunotherapy. Crit Rev Immunol (2013) 33(1):1–21. doi: 10.1615/CritRevImmunol.2013006679
115. Wculek SK, Cueto FJ, Mujal AM, Melero I, Krummel MF, Sancho D. Dendritic Cells in Cancer Immunology and Immunotherapy. Nat Rev Immunol (2020) 20(1):7–24. doi: 10.1038/s41577-019-0210-z
116. Zong J, Keskinov AA, Shurin GV, Shurin MR. Tumor-Derived Factors Modulating Dendritic Cell Function. Cancer Immunol Immunother (2016) 65(7):821–33. doi: 10.1007/s00262-016-1820-y
117. Gabrilovich D. Mechanisms and Functional Significance of Tumour-Induced Dendritic-Cell Defects. Nat Rev Immunol (2004) 4(12):941–52. doi: 10.1038/nri1498
118. Janikashvili N, Bonnotte B, Katsanis E, Larmonier N. The Dendritic Cell-Regulatory T Lymphocyte Crosstalk Contributes to Tumor-Induced Tolerance. Clin Dev Immunol (2011) 2011:430394. doi: 10.1155/2011/430394
119. Long J, Hu Z, Xue H, Wang Y, Chen J, Tang F, et al. Vascular Endothelial Growth Factor (VEGF) Impairs the Motility and Immune Function of Human Mature Dendritic Cells Through the VEGF Receptor 2-RhoA-Cofilin1 Pathway. Cancer Sci (2019) 110(8):2357–67. doi: 10.1111/cas.14091
120. Sisirak V, Vey N, Goutagny N, Renaudineau S, Malfroy M, Thys S, et al. Breast Cancer-Derived Transforming Growth Factor-Beta and Tumor Necrosis Factor-Alpha Compromise Interferon-Alpha Production by Tumor-Associated Plasmacytoid Dendritic Cells. Int J Cancer (2013) 133(3):771–8. doi: 10.1002/ijc.28072
121. Ruffell B, Chang-Strachan D, Chan V, Rosenbusch A, Ho CM, Pryer N, et al. Macrophage IL-10 Blocks CD8+ T Cell-Dependent Responses to Chemotherapy by Suppressing IL-12 Expression in Intratumoral Dendritic Cells. Cancer Cell (2014) 26(5):623–37. doi: 10.1016/j.ccell.2014.09.006
122. Pockaj BA, Basu GD, Pathangey LB, Gray RJ, Hernandez JL, Gendler SJ, et al. Reduced T-Cell and Dendritic Cell Function is Related to Cyclooxygenase-2 Overexpression and Prostaglandin E2 Secretion in Patients With Breast Cancer. Ann Surg Oncol (2004) 11(3):328–39. doi: 10.1245/ASO.2004.05.027
123. Gervais A, Leveque J, Bouet-Toussaint F, Burtin F, Lesimple T, Sulpice L, et al. Dendritic Cells are Defective in Breast Cancer Patients: A Potential Role for Polyamine in This Immunodeficiency. Breast Cancer Res (2005) 7(3):R326–35. doi: 10.1186/bcr1001
124. Norian LA, Rodriguez PC, O'Mara LA, Zabaleta J, Ochoa AC, Cella M, et al. Tumor-Infiltrating Regulatory Dendritic Cells Inhibit CD8+ T Cell Function via L-Arginine Metabolism. Cancer Res (2009) 69(7):3086–94. doi: 10.1158/0008-5472.CAN-08-2826
125. Ning Y, Shen K, Wu Q, Sun X, Bai Y, Xie Y, et al. Tumor Exosomes Block Dendritic Cells Maturation to Decrease the T Cell Immune Response. Immunol Lett (2018) 199:36–43. doi: 10.1016/j.imlet.2018.05.002
126. Hanks BA, Holtzhausen A, Evans KS, Jamieson R, Gimpel P, Campbell OM, et al. Type III TGF-Beta Receptor Downregulation Generates an Immunotolerant Tumor Microenvironment. J Clin Invest (2013) 123(9):3925–40. doi: 10.1172/JCI65745
127. Faget J, Bendriss-Vermare N, Gobert M, Durand I, Olive D, Biota C, et al. ICOS-Ligand Expression on Plasmacytoid Dendritic Cells Supports Breast Cancer Progression by Promoting the Accumulation of Immunosuppressive CD4+ T Cells. Cancer Res (2012) 72(23):6130–41. doi: 10.1158/0008-5472.CAN-12-2409
128. Pedroza-Gonzalez A, Xu K, Wu TC, Aspord C, Tindle S, Marches F, et al. Thymic Stromal Lymphopoietin Fosters Human Breast Tumor Growth by Promoting Type 2 Inflammation. J Exp Med (2011) 208(3):479–90. doi: 10.1084/jem.20102131
129. Fainaru O, Almog N, Yung CW, Nakai K, Montoya-Zavala M, Abdollahi A, et al. Tumor Growth and Angiogenesis are Dependent on the Presence of Immature Dendritic Cells. FASEB J (2010) 24(5):1411–8. doi: 10.1096/fj.09-147025
130. Gadalla R, Hassan H, Ibrahim SA, Abdullah MS, Gaballah A, Greve B, et al. Tumor Microenvironmental Plasmacytoid Dendritic Cells Contribute to Breast Cancer Lymph Node Metastasis via CXCR4/SDF-1 Axis. Breast Cancer Res Treat (2019) 174(3):679–91. doi: 10.1007/s10549-019-05129-8
131. Chen L, Zheng H, Yu X, Liu L, Li H, Zhu H, et al. Tumor-Secreted GRP78 Promotes the Establishment of a Pre-Metastatic Niche in the Liver Microenvironment. Front Immunol (2020) 11:584458. doi: 10.3389/fimmu.2020.584458
132. Monteiro AC, Bonomo A. Dendritic Cells Development Into Osteoclast-Type APCs by 4T1 Breast Tumor T Cells Milieu Boost Bone Consumption. Bone (2020) 143:115755. doi: 10.1016/j.bone.2020.115755
Keywords: breast cancer, tumor-promoting myeloid cells, immunosuppression, immunotherapies, tumor microenvironment
Citation: Blaye C, Boyer T, Peyraud F, Domblides C and Larmonier N (2022) Beyond Immunosuppression: The Multifaceted Functions of Tumor-Promoting Myeloid Cells in Breast Cancers. Front. Immunol. 13:838040. doi: 10.3389/fimmu.2022.838040
Received: 17 December 2021; Accepted: 02 February 2022;
Published: 03 March 2022.
Edited by:
Giovanna Schiavoni, National Institute of Health (ISS), ItalyReviewed by:
Keehoon Jung, Seoul National University, South KoreaLinda Schuler, University of Wisconsin-Madison, United States
Copyright © 2022 Blaye, Boyer, Peyraud, Domblides and Larmonier. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Nicolas Larmonier, nicolas.larmonier@u-bordeaux.fr