 Vanaja Konduri1Damilola Oyewole-Said1Jonathan Vazquez-Perez1Scott A. Weldon2Matthew M. Halpert1
Vanaja Konduri1Damilola Oyewole-Said1Jonathan Vazquez-Perez1Scott A. Weldon2Matthew M. Halpert1 Jonathan M. Levitt1,3,4
Jonathan M. Levitt1,3,4 William K. Decker1,3,5*
William K. Decker1,3,5*- 1Department of Pathology & Immunology, Baylor College of Medicine, Houston, TX, United States
- 2Michael E. DeBakey Department of Surgery, Baylor College of Medicine, Houston, TX, United States
- 3Dan L. Duncan Cancer Center, Baylor College of Medicine, Houston, TX, United States
- 4Scott Department of Urology, Baylor College of Medicine, Houston, TX, United States
- 5Center for Cell and Gene Therapy, Baylor College of Medicine, Houston, TX, United States
NK1.1 and its human homolog CD161 are expressed on NK cells, subsets of CD4+ and CD8+ T cells, and NKT cells. While the expression of NK1.1 is thought to be inhibitory to NK cell function, it is reported to play both costimulatory and coinhibitory roles in T-cells. CD161 has been extensively studied and characterized on subsets of T-cells that are MR1-restricted, IL-17 producing CD4+ (TH17 MAIT cells) and CD8+ T cells (Tc17 cells). Non-MAIT, MR1-independent CD161-expressing T-cells also exist and are characterized as generally effector memory cells with a stem cell like phenotype. Gene expression analysis of this enigmatic subset indicates a significant enhancement in the expression of cytotoxic granzyme molecules and innate like stress receptors in CD8+NK1.1+/CD8+CD161+ cells in comparison to CD8+ cells that do not express NK1.1 or CD161. First identified and studied in the context of viral infection, the role of CD8+CD161+ T-cells, especially in the context of tumor immunology, is still poorly understood. In this review, the functional characteristics of the CD161-expressing CD8+ T cell subset with respect to gene expression profile, cytotoxicity, and tissue homing properties are discussed, and application of this subset to immune responses against infectious disease and cancer is considered.
Introduction
Establishment of long-lived memory T cell populations with enhanced cytotoxicity kinetics and that provide durable immunity against reinfection are critical to the cell mediated immune response (1). An intriguing subset of such memory T cells is identified by the expression of natural cytotoxicity receptor NK1.1 in mice and CD161 in humans and has been under investigation by several groups (2–9). In mice, the NK1.1 receptor is expressed on almost all NK cells and subsets of T cells including CD4+, CD8+, and NKT cells (5). While majority of the NK1.1+ T cells are CD4+, a significant proportion of viral antigen specific CD8+ T cells are reported to express NK cell markers in mice subjected to experimental viral infection (6, 10, 11). Unlike classical NKT cells, the CD8+NK1.1+ T cell subset is not only generated in NKT cell-deficient mice but also expresses a diverse polyclonal TCR repertoire suggesting these are not classical NKT cells (6). While some NK1.1+ CD8+ T cells are CD1d restricted, CD1d independent NK1.1+CD8+ cells with a memory phenotype have been found to be significantly increased in the liver and to persist following allo-HCT. Establishment of this TCRαβ+ T cell population derived from donor splenocytes and not bone marrow precursors could be abrogated by depletion of CD8+ cells but not NK1.1+ cells from the donor HCT inoculum, providing significant evidence that CD8+ T cells can acquire the expression of NK markers (12).
CD161, the human homolog of the mouse NK1.1 is expressed by NK cells and 24% of T cells including both γδ and αβ TCR-expressing subsets, NKT cells, monocytes, and dendritic cells (13, 14). CD161 expressing T cells exhibit varying levels of NK-cell like innate activity and are considered in some contexts as markers of “innateness” among T cells (15). Peripheral blood analysis of T cells from healthy donors indicates that CD161 is also preferentially expressed on memory T cell subsets (13). While CD4+ T cells express intermediate levels of CD161, CD8+ T cells may express CD161 at either intermediate or high levels (7, 9). CD161high CD8+ T cells displaying a TH17 phenotype with upregulated expression of RORγt, CCR6, and IL18R are well characterized and described as Tc17 cells (Figure 1) (7). CD161neg subsets exhibit naïve and central memory phenotypes while the CD161high subset displays an effector memory phenotype (16). With no evidence of IL-17 secretion, CD161int cells are reported to be a unique population of memory CD8+ cells with enhanced effector functions (7, 17, 18). Based on the expression of CCR7 and CD45RA, CD161high, CD161int, and CD161neg subsets have been characterized as naïve, effector memory, and TEMRA (T effector memory with RA) phenotypes (18). While a majority of the CD161high cells display a TEM phenotype, CD161neg cells display a TCM phenotype, and CD161int cells display both TEM and TEMRA phenotypes.

Figure 1 CD161-expressing CD8+ T cells are classified as tissue homing MAIT (Mucosal Associated Invariant T) cells, cytotoxic Tc17 cells, or Tmem (stem cell like memory) cells that display tissue homing and enhanced cytotoxic characteristics in addition to memory. Tmem cells respond to viral infections and can be activated by antigen presenting cells loaded with viral peptides or intracellular bacteria. In response to infection or antigenic stimulation, Tmem cells upregulate chemokine and stress receptors and secrete granzyme, INF-γ, and in some instances, IL-17. CD161+ Tmem cells home to vital organs like lungs, liver, gut, pancreas, and brain where they respond to viral infections and also display anti-tumor properties.
Previous studies have extensively characterized the shared pre-programmed phenotype of the MAIT and non-MAIT CD8+CD161high T cells and their ability to respond to cytokine stimulation (9). MR1-restricted MAIT cells are the best characterized CD161+ CD8+ T cells and have been shown to detect a variety of microbes through the recognition of vitamin B metabolites presented by MR1 (19). Non-MAIT CD161+CD8+ T cells on the other hand exhibit specificity to viral antigens, are polyclonal in nature, and exhibit stem cell like memory phenotypes (Figure 1) (8). Apart from enhanced memory potential, low levels of PD-1 (exhaustion), CD161 expressing CD8+ T cells are also known to be tissue homing and demonstrate high levels of cytotoxicity against infectious agents, making them suitable therapeutic candidates against a wide range of infectious diseases and tumors (4, 7, 20). This review will focus on the expression of CD161 and the mouse homolog NK1.1 on polyclonal CD8+ T cells and discuss the biological significance of the expression of this receptor in disease.
Cytotoxicity
CD8+NK1.1+ T Cells in Mice Share Cytolytic Functions of Conventional CD8+ T Cells
Effective cellular immune responses rely substantially on the cytolytic ability of the responding immune cells and first line defenders. In this context, cytotoxic CD8+ T cells and NK cells play a critical role in providing durable and protective immune responses. Accordingly, research efforts are focused on the therapeutic potential of subsets that combine characteristics of both CD8+ T cells and NK cells (8). In mice, CD8+NK1.1+ T cells have been shown to share cytolytic functions of conventional CD8+ T cells with a TH1 skewed cytokine profile and lytic activity upon CD3 stimulation (21).
Cytotoxicity of CD8+CD161high Effector Memory T Cells Is Primarily Mediated Through Granzymes
In humans, the cytotoxic potential of different subsets of CD161+ expressing CD8+ T cells has not been clearly defined with reports of both high and low cytotoxic potential. CD8+CD161high cells were once described as anergic subset of T cells with high levels of intracellular granzyme and perforin and no killing activity (5). In another report, CD161high CD8+ T cells with a TH17 phenotype (Tc17 cells) displayed reduced cytotoxic potential compared to conventional effector CD8+ cells and were associated with low expression of granzyme B and perforin (8). Tc17 populations in mice also displayed limited cytolytic ability (22, 23). In contrast, another study reported that 29% of CD8+CD161neg, 56% of CD8+CD161int, and 88% of CD8+CD161high cells stained positive for granzyme A suggesting high cytotoxic potential (5). CD161high CD8+ T cells have also been reported to upregulate granzyme B and perforin and become highly cytotoxic upon activation (20) suggesting these are not similar to Tc17 cells but are more analogous to effector memory cells with stem cell like characteristics (3). We and others have also shown that, compared to the CD8+CD161neg and CD8+CD161int subsets, CD8+CD161high cells produced significantly lower amounts of IFN-γ (5) suggesting that cytotoxicity may be mediated primarily through granzyme pathways.
Tissue Homing
Preprogrammed Expression of Tissue Resident Markers Enable Migration and Homing of CD8+CD161high T Cells
Tissue resident innate and adaptive immune cells that do not recirculate in blood or lymphatics adopt a unique phenotype and contribute to barrier immunity, tissue homeostasis, and immune regulation (24). Studies on central and effector memory CD8+ T cell subsets distinguished by homing markers such as CCR7 and CD62L and detected in peripheral non-lymphoid organs have largely contributed to the concept of tissue-resident lymphocytes (16). Cytotoxic cells with tissue homing properties, especially to the mucosal tissues of the gastrointestinal tract, can act as a first line of defense at pathogen entry points and provide protective immune responses (18). Within the CD4+ and CD8+ T cell subsets, CD161+ cells make up more than half of T cells in the intestine (25). Among CD8+ T cells, tissue homing properties of subsets expressing CD161 have been well defined (7). Transendothelial migration of CD161+ CD8+ T cells without chemotactic stimuli suggests a tissue homing preference for these cells (14). CD8+ T cells expressing intermediate and high levels of CD161 are reported to also secrete high levels of IL-22, a cytokine involved in tissue repair and epithelial defense. Although MAIT cells were reported to be involved in tissue repair, recent studies on transcriptomic analysis of human and mouse MAIT cells identified a distinct tissue homing gene signature. Scratches on monolayers of colonic Caco2 cells were successfully closed by applying culture supernatants from MAIT cells activated by E. coli. MR1 blockade abrogated the effect confirming the TCR-dependent tissue repair potential of MAIT cells (26–30). CD161high CD8+ MAIT cells are highly enriched in mucosal tissues and significantly upregulate chemokine receptors such as CXCR6 and CCR6 (31). In peripheral blood CD8+ cells, CD161 expression is associated with CXCR6, a chemokine receptor that binds CXCL16 constitutively expressed by the liver and respiratory tract (4, 32). CCR6+CD161highCD8+ cells have been detected in the naïve T cell population of cord blood suggesting pre-programmed expression of tissue resident markers enabling these cells to also home to non-lymphoid organs such as liver and gut (7). Previous studies have also shown that CD161 and CCR6 alone favor T cell migration and tissue homing (14, 33). Compared to the levels found in peripheral blood, CD161highCD8+ T cells are highly enriched in the gut and liver, while CD161intCD8+ cells are enriched in the colon. CD161intCD8+ cells also expressed CD103 and CD69, markers consistent with a tissue resident phenotype (18). CD161high CD8+ MAIT cells with high expression of the multidrug resistance transporter ABCB1 are reported to preferentially home to intestine and liver and display an effector phenotype. Representing up to 45% of the liver lymphocytes, MAIT cells produce IFN-γ and granzyme B and upon stimulation with PMA/Ionomycin and secrete high levels of IL-17 (31). Thus, CD161 expressing CD8 T cells represent subsets of immune cells with therapeutic potential against tissue specific infections and diseases. Although there exists an overlap in the properties of CD161 expressing CD8+ T cell subsets, enhanced cytotoxicity, effector functions, tissue homing properties, survival and resistance to xenobiotics, make these cells key therapeutic effectors.
CD8+NK1.1+/CD8+CD161+ T Cell Gene Expression Studies
CD8+NK1.1+ Cells Express Innate Stress Receptors and Cytotoxic Granzymes
Gene expression profile analysis of CD1d independent, polyclonal CD8+NK1.1+ NKT-like cells, CD8+NK1.1- cells, and conventional NK cells indicated that CD8+NK1.1+ cells showed a combination of T cell and NK cell markers, suggesting their potential to function as a kind of hybrid CTL/NK cell (34). Significantly elevated expression of pro-inflammatory cytokines, chemokines, chemokine receptors and other adhesion molecules indicate a distinct capacity to migrate, and elevated levels of cytotoxicity molecules involved in granule exocytosis suggest elevated killing potential (34). Gene expression analysis of antigen experienced CD8+NK1.1+ cells revealed a significant upregulation of cytotoxic granzymes F, D, G, C, B, A, and N and innate like stress receptors in comparison to the CD8+NK1.1- subset.
CD8+CD161+ Cells Express Chemokine, Tissue Homing, Cytotoxic, and Innate Stress Receptors
Previous gene expression studies of CD8+CD161+ Tc17 cells revealed upregulation of several TH17 related genes including RORγt, CYP1B1, and the chemokine receptors IL23R, CXCR6, and IL8R (7, 35). CCR7 was downregulated among the CD161+ cells (7). Gene expression analysis of CD161int cells from adult circulating peripheral blood and naïve umbilical cord blood revealed significant upregulation of IL18R, CXCR6, MDR1, and PLZF indicating a preprogrammed phenotype with tissue homing properties (18). Expression of MDR1 on the CD161high and CD161int subsets enables survival of these subsets in gastrointestinal tissues exposed to xenobiotics (18). CD161int subsets also showed significantly elevated levels of granzyme B and perforin compared to the CD161neg subsets. CD161high MAIT cells on the other hand are potent producers of IFN-γ (9, 18). Different subsets of CD8+CD161+ T cells including MAIT, Tc17, and non-MAIT effectors show overlap in gene expression profiles suggesting enhanced tissue homing and migratory potential of these cells is facilitated by the combined action of different subsets.
Molecular Mechanisms of NK1.1/CD161 Ligand Interactions
In mice, NK1.1 belongs to the family of NKRP1 receptors, a family of disulfide linked homodimers that interact with C-type lectin related (Clr) molecules (13). NKRP1B and NKRP1D are inhibitory receptors and recognize Clr-b (encoded by Clec2d) while the activating receptor NKRP1F binds to Clr-g (36, 37).
CD161 Engagement With Its Ligand LLT1 Can Be Both Costimulatory and Inhibitory
In humans, the CD161 receptor KLRB1 is expressed on NK and T cells and interacts with the ligands Clec2/Lectin like transcript-1 (LLT1) and the lesser characterized PILAR (proliferation induced lymphocyte receptor) (38–40). PILAR is known to modulate T cell expansion and acts as a survival signal for CD161-expressing naïve and activated T cells (41). Restricted to hematopoietic cells, LLT1 is not expressed on the surface of resting PBMCs but can be transiently expressed on activated B cells, dendritic cells, T cells and NK cells (42). Elevated expression of LLT1 has been reported on germinal center B cells in tonsils and lymph nodes and B cell derived lymphomas (43). In NK cells, the interaction of LLT1 with the CD161 receptor is described as inhibitory (13, 39, 44) while in T cells, CD161 engagement with LLT1 is considered both costimulatory (39, 45) and inhibitory (38, 46). CD161 receptor engagement with the ligand LLT1 was not sufficient to trigger IFN-γ production among T cells unless simultaneously engaged with CD3 (39). LLT1 interaction with CD161 did not modulate degranulation in CD8 T cells but partially inhibited TNF-α production (38). Since CD161 is not known to possess any characterized signaling motifs, additional studies will be required to better understand the true consequences of this ligand/receptor interaction (39).
CD161 Intracellular Signaling in CD8+ T Cells Is Not Well Defined
In NK cells, CD161 has been shown to directly interact with acid sphingomyelinase (ASM) resulting in intracellular AKT signaling and regulation of NK cell function (47). In T cells however, especially CD8+ T cells, CD161 signaling is not very well defined. In CD4+ CD161+ Th17 cells, CD161 is shown to interact with the surface receptor CD39 to further amplify ASM-mediated mTOR and STAT3 signals driving Th17 expansion (48, 49). Further studies are warranted in understanding the signaling events among non-Th17 CD8+ CD161+ T cells.
Role of CD8+NK1.1+/CD8+CD161+ in Infection and Immunity
In the infectious disease literature, CD8+NK1.1+ cells and the analogous human CD8+CD161+ cells are described as highly cytotoxic memory T cells with antiviral specificity (6–8, 11, 18).
CD8+NK1.1+ T Cells Are Protective Against Viral Infections and Intracellular Pathogens
In previous studies, we observed a significant upregulation in the number of peripheral CD8+NK1.1+ cells following administration of a TH1 polarized dendritic cell vaccine (50–53). In mice, TGF-β is reported to repress the differentiation of NK1.1+ T cells from CD8+ T cells. However, upon bacterial or viral infection a fraction of CD8+ T cells have been shown to escape TGF-β control during priming and acquire NK1.1 expression. In an LCMV infection model, unlike the CD8+NK1.1neg subset, the CD8+NK1.1+ cells underwent delayed contraction and apoptosis and provided prolonged pathogen-specific reactivity by producing IFN-γ and granzyme B (54). Endowed with innate immunity features and contributing to the adaptive immune responses, these CD8+NK1.1+ cells not only cleared the initial microbial and viral infections but also offered protection against reinfection. In a study involving lethal Listeria infection, CD8+NK1.1+ cells provided rapid innate immune responses characterized by early, antigen-independent IFN-γ production, granzyme B expression, degranulation, and protection against re-exposure (55). In a separate study, CD8+NK1.1+ T cells were shown to comprise 10% of total CD8+ T cells in the lungs and offer durable protection at ten days after primary influenza infection (11). These cells were elevated in number in CD1d-/- mice suggesting they are not NKT cells but a distinct population in which NK1.1 may modulate effector functions of activated antigen experienced CD8+ T cells. CD8+NK1.1+ cells described as Tc17 cells were also highly protective against lethal influenza infection (23). In intracellular parasite infection models, CD1d-independent CD8+NK1.1+ T cells have been shown to play a protective role against the liver stage of Plasmodium yoelii infection (56). A significant increase in the number of splenic antigen experienced, activated CD8+NK1.1+ T cells was also seen during the acute stage of Plasmodium chabaudi infection (57). These studies suggest that in murine models, CD8+NK1.1+ T cells are protective against viral infections and intracellular pathogens. Antigen dependent activation leads to an enhanced proliferation of these cells and upregulation of innate stress receptors, cytotoxic molecules resulting in durable protective responses against reinfection and improved disease-free survival.
CD8+CD161+ Cells Offer Pathogen Immunity, Specifically to Viral Infection
In humans, CD161 has been reported as a marker for long lived memory CD4+ T cells. It was reported that the proportion of influenza specific CD4+CD161+ T cells was more highly elevated at two years post immunization than four weeks post immunization, suggesting that CD161 is a marker of long-term memory among T cells (58). Several groups have reported the role of CD8+CD161+ cells in pathogen immunity, specifically immunity to viral infection. Enrichment of CD161+ cells was seen in the liver in response to infection and non-alcoholic steatohepatitis (7). CD8+CD161+ cells specific for hepatitis C virus (HCV) and hepatitis B virus (HBV) were reported earlier (4). TH17 cells responding to HCV specific peptides have been reported (7, 59). CD161+ MAIT cells on the other hand are reported to be responsive to bacterial infections (19). CD161highCD8+ T cells specific for EBV, CMV, or influenza encompassed IL18Rahigh, IL7, and IL15 responding memory cells expressing higher levels of anti-apoptotic molecules and high drug efflux capacity (3) suggesting these cells can survive hostile inflammatory conditions leading to pathogenesis of tissues such as in inflamed CNS.
IL-17 Producing CD161+ T Cells Implicated in Auto-Immune Diseases
CD161 expressing T cells, specifically the IL-17 producing subset, have been implicated in auto-immune diseases like psoriasis, Crohn’s disease, rheumatoid arthritis, and multiple sclerosis (7, 60–64). A subset of CD8+CD161int cells with elevated expression of granzyme B and perforin have been shown to cross the blood brain barrier and are enriched in MS lesions (17). While enriched in the CNS, CD8+CD161+ cells were reduced in number in the peripheral blood in MS patients in comparison to healthy adults. In MS brain infiltrates, 10% of all CD8+ T cells were IFN-γ producing CD161+ cells that also secreted IL-17 and IL-22 and contributed to the pathogenesis of the disease (64). Activation induced expansion of CD161 cells and the implication of CD161 polymorphism in MS suggests potential therapeutic modulation of these cells in disease conditions mediated or ameliorated by CD8+ T-cells (40, 65, 66). In SLE, a disease in which CD8+ cells play a relatively minor role in pathogenesis, reduced CD161 expression on CD8+ T cells and NKT cells was noted in patients with advanced disease (67).
Roles of CD8+NK1.1+/CD8+CD161+ in Tumor Immunology
CD8+NK1.1+ T Cells Offer Protection and Improve Survival Against Multiple Murine Tumor Malignancies
Although the role of NK1.1+ T cells is defined in the context of viral infection, little is known about any potential role for this receptor on CD8+ T cells in the context of cancer. In mice, in vitro expansion of highly cytotoxic CD8+NK1.1+ T cells derived from bone marrow, spleen, and thymus have been shown to mediate strong anti-leukemia effects without GVHD after allogeneic transplantation (68, 69). In a model of murine pancreatic cancer, NKT cells have been shown to offer protection by modulating tumor associated macrophages to drive a TH1 adaptive immune response (70). In another study, the tumoricidal effects of the CD8+NK1.1+ cells derived from OT-1 mice were significantly higher than that of the CD8+NK1.1neg equivalent population (71). In a B16 lung metastatic in vivo model, adoptive transfer of CD8+NK1.1+ NKT-like cells significantly inhibited metastasis and improved survival in comparison to adoptive transfer of CD8+NK1.1neg cells or conventional NK cells (34). In in vitro killing assays, CD8+NK1.1+ NKT-like cells exerted cytotoxicity against tumor cells and MDSCs (myeloid derived suppressor cells) through a granzyme B-mediated granule exocytosis pathway. Granzyme B inhibitors suppressed the cytotoxic effects while treatment of the cells with anti-FasL, anti-TRAIL, or anti-IFN-γ antibodies did not (34). In a previous study, we demonstrated that adoptively transferred CD8+NK1.1+ cells offered durable protection against murine PDAC (pancreatic ductal adenocarcinoma) and improved survival (53). These cells were present nine months after initial antigen exposure and were potent in clearing tumors. Gene expression analysis revealed that these CD8+NK1.1+ cells exhibited significantly elevated levels of cytotoxic molecules and stress receptors. These studies highlight the role of CD8+NK1.1+ cells in generating durable anti-tumor responses against murine tumors suggesting their potential in cell based therapeutics. Enhanced cytotoxicity and memory characteristics of these cells make them ideal candidates for cell based therapies against aggressive malignancies.
CD161 Expression on T Cells Results in Both Favorable and Unfavorable Outcomes Against Tumors
In humans, there are limited studies defining the role of CD161 expression on CD8+ T cells in tumor immunology with reports suggesting both favorable and unfavorable outcomes. In a cancer- wide genome analysis of prognostic gene signatures, KLRB1, the gene encoding CD161, was identified as most frequently associated with favorable outcomes against several indications including bladder, breast, colon, prostate cancers, melanoma, lung adenocarcinoma, multiple myeloma, glioma, and neuroblastoma to name a few (72, 73). Transcription of KLRB1 was suppressed in 68% of NSCLC (non-small cell lung cancer) and 57% of esophageal squamous-cell carcinoma patients indicating that CD161 can be a predictive marker in these indications (74). In NSCLC, interaction of CD161 expressing tumor infiltrating CD4+ and CD8+ T cells with LLT1-expressing germinal center B cells within tumor microenvironment tertiary lymphoid structures resulted in improved survival (73). In the CD4+ T cells, genes associated with KLRB1 expression were CCR2, CCL4, and GZMA, markers of a TH1 polarized effector phenotype (73). Similarly, in oropharyngeal cancer, IFN-γ and IL-17 producing CD161+ tumor infiltrating T cells were associated with better tumor control (75). In head and neck cancer, CD161 expression was significantly down modulated in the peripheral blood compared to peripheral blood from healthy controls and in tumor infiltrating TH17 cells as a postulated immune escape mechanism induced by the tumor milieu (41). In a recent study of hepatocellular carcinoma (HCC), infiltration of CD8+ PD-1+ CD161+T cells into the tumor microenvironment represented reactivated cytotoxic cells with proliferative and not exhaustive characteristics and correlated with better prognosis. Co-expression of IL-7R and enhanced expressions of IL-2, TNF- α and perforin mediated the maintenance of proliferative phenotype (76). These studies highlight the therapeutic potential of CD8+CD161+ T cells as prognostic biomarkers and/or candidates for cell based therapies. Enhanced cytotoxicity, tissue homing and memory characteristics make them suitable candidates against aggressive malignancies (Table 1).
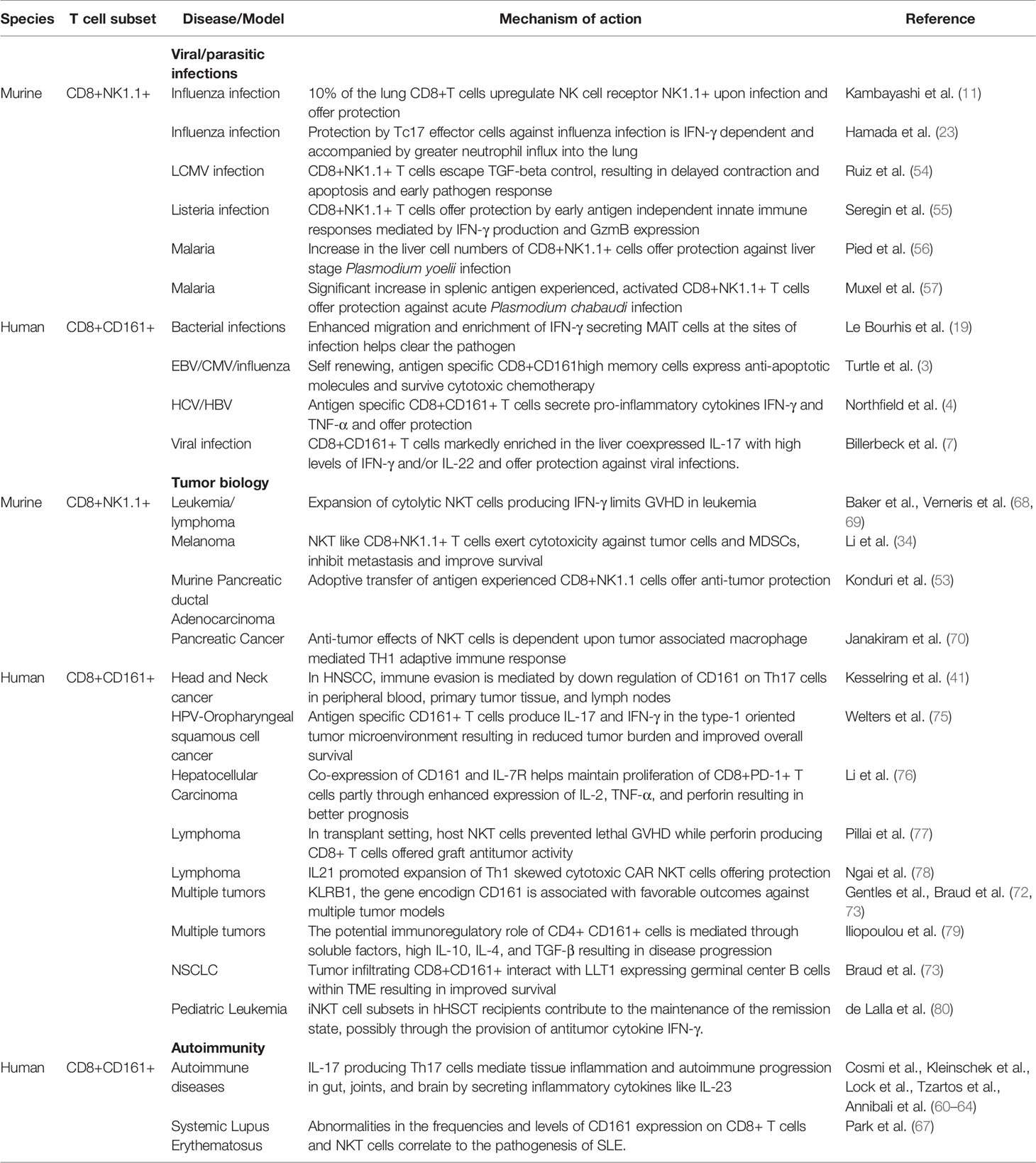
Table 1 Roles of CD8+NK1.1+/CD8+CD161+ Cells in Infection and Immunity.
CD161 Expression Correlates With Immune Regulatory Functions
In other studies, CD161 expression has been described as a negative prognostic marker. In a previous study, analysis of several different malignancies showed a significant increase in the number of CD161 expressing CD4+ T cells in the peripheral blood of cancer patients compared to healthy individuals, and this increase was positively correlated with disease stage. The potential immunoregulatory role of CD4+ CD161+ cells was mediated through soluble factors, mainly through high IL-10, IL-4, and TGF- β (79). In T cell lymphoblastic leukemia, aberrant expression of CD161 and other NK cell markers like CD56 and CD16 on T cells was correlated with disease progression (81). In T cell pro-lymphocytic leukemia (T-PLL), aberrant CD161 expression on T cells was not associated with other NK cell markers suggesting the usefulness of this unique phenotype as a diagnostic marker (82). These studies suggest that the expression of CD161 on T cells can serve both activating and inhibitory roles in tumor progression.
The natural anti-tumor effector properties of CD1d restricted NKT cells have been effectively used to treat lymphomas without associated GVHD. Peripheral blood NKTs also offered long term remission against pediatric leukemia (77, 80). Ex vivo expanded CAR NKTs in the presence of IL-21 promoted expansion of highly cytotoxic TH1 polarized cells that enabled long term survival of lymphoma bearing mice (78). However, the potential applicability of CD161 expressing CD8+ CAR T cells in the solid tumor setting has not yet been evaluated.
Conclusions and Future Work
CD8+CD161+ T cells and their murine counterparts CD8+NK1.1+ T cells exhibit elevated cytotoxic potential, characteristics of long-term memory, drug–effluxing capacity, and extended survival and, as such, are potential candidates for adoptive cell therapy to treat pathologic indications (3, 58). Tissue homing properties of these cells to mucosal surfaces may mediate protection at pathogen sites of entry (18). Gene expression analysis indicates enhanced expression levels of granzyme, perforin, and innate-like receptors among these cells when activated in comparison to NK1.1neg/CD161neg counterparts. Although studied extensively in the context of viral infection, there is limited information on the homeostatic function of this cell subset in cancer progression. With confounding reports of protective and inhibitory potential, it is imperative that they are tested for their efficacy under specific conditions. The effector functions of these cells operate by both cytolytic and noncytolytic mechanisms. In the case of LCMV infection, viral load is controlled by cytotoxic granzymes and perforin secreted by these cells, while antiviral activity against HBV and HCV is regulated principally by IFN-γ secretion. Protective effects of these cells against murine influenza is also driven by IFN-γ secretion. Thus, cytolytic and non-cytolytic protective effects of these cells depend upon the pathogenicity of infection and the tissue environment (Table 1). Little is known about the cascade of signaling events that lead to the effector functions of CD161 activation. Further work is needed to delineate the role of costimulatory and signaling molecules during CD161 engagement. When simultaneously stimulated with CD3, ligation of CD161 on CD8+ T cells serves as a co-stimulatory molecule for IFN-γ production as seen during pathogenesis of multiple sclerosis. Ligand interactions play an important role in the effector functions of this subset, and it will be interesting to test the cytotoxic efficacy of these cells against tumors expressing LLT1. Nonetheless, the sum total of the data indicated that CD8+CD161+ T cells are critical effector memory cytolytic effectors that are TH1 polarized, tissue homing, and primarily of anti-viral specificity. Future investigations into the biology of these cells will focus on signaling events downstream of CD161 ligation but will also necessarily look upstream at the signals, environmental conditions, and APC subsets present at the time of priming.
Author Contributions
VK, DO-S, JV-P, MH, JL, and WD all collaborated in the writing and editing of this manuscript. SW provided the original artwork. All authors contributed to the article and approved the submitted version.
Funding
This work was funded in part by NIH R01 AI127387 (to WD).
Conflict of Interest
Institutional policy requires VK, MH, and WD to declare their ownership stakes in Diakonos Research, Ltd.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. Seaman MS, Peyerl FW, Jackson SS, Lifton MA, Gorgone DA, Schmitz JE, et al. Subsets of memory cytotoxic T lymphocytes elicited by vaccination influence the efficiency of secondary expansion in vivo. J Virol (2004) 78(1):206–15. doi: 10.1128/JVI.78.1.206-215.2004
2. Martin E, Treiner E, Duban L, Guerri L, Laude H, Toly C, et al. Stepwise development of MAIT cells in mouse and human. PloS Biol (2009) 7(3):e54. doi: 10.1371/journal.pbio.1000054
3. Turtle CJ, Swanson HM, Fujii N, Estey EH, Riddell SR. A distinct subset of self-renewing human memory CD8+ T cells survives cytotoxic chemotherapy. Immunity (2009) 31(5):834–44. doi: 10.1016/j.immuni.2009.09.015
4. Northfield JW, Kasprowicz V, Lucas M, Kersting N, Bengsch B, Kim A, et al. CD161 expression on hepatitis C virus-specific CD8+ T cells suggests a distinct pathway of T cell differentiation. Hepatology (2008) 47(2):396–406. doi: 10.1002/hep.22040
5. Takahashi T, Dejbakhsh-Jones S, Strober S. Expression of CD161 (NKR-P1A) defines subsets of human CD4 and CD8 T cells with different functional activities. J Immunol (2006) 176(1):211–6. doi: 10.4049/jimmunol.176.1.211
6. Assarsson E, Kambayashi T, Sandberg JK, Hong S, Taniguchi M, Van Kaer L, et al. CD8+ T cells rapidly acquire NK1.1 and NK cell-associated molecules upon stimulation in vitro and in vivo. J Immunol (2000) 165(7):3673–9. doi: 10.4049/jimmunol.165.7.3673
7. Billerbeck E, Kang YH, Walker L, Lockstone H, Grafmueller S, Fleming V, et al. Analysis of CD161 expression on human CD8+ T cells defines a distinct functional subset with tissue-homing properties. Proc Natl Acad Sci USA (2010) 107(7):3006–11. doi: 10.1073/pnas.0914839107
8. Fergusson JR, Fleming VM, Klenerman P. CD161-expressing human T cells. Front Immunol (2011) 2:36. doi: 10.3389/fimmu.2011.00036
9. Fergusson JR, Smith KE, Fleming VM, Rajoriya N, Newell EW, Simmons R, et al. CD161 defines a transcriptional and functional phenotype across distinct human T cell lineages. Cell Rep (2014) 9(3):1075–88. doi: 10.1016/j.celrep.2014.09.045
10. Slifka MK, Pagarigan RR, Whitton JL. NK markers are expressed on a high percentage of virus-specific CD8+ and CD4+ T cells. J Immunol (2000) 164(4):2009–15. doi: 10.4049/jimmunol.164.4.2009
11. Kambayashi T, Assarsson E, Michaelsson J, Berglund P, Diehl AD, Chambers BJ, et al. Emergence of CD8+ T cells expressing NK cell receptors in influenza A virus-infected mice. J Immunol (2000) 165(9):4964–9. doi: 10.4049/jimmunol.165.9.4964
12. Wang Y, Wang H, Xia J, Liang T, Wang G, Li X, et al. Activated CD8 T cells acquire NK1.1 expression and preferentially locate in the liver in mice after allogeneic hematopoietic cell transplantation. Immunol Lett (2013) 150(1-2):75–8. doi: 10.1016/j.imlet.2012.12.009
13. Lanier LL, Chang C, Phillips JH. Human NKR-P1A. A disulfide-linked homodimer of the C-type lectin superfamily expressed by a subset of NK and. J Immunol (1994) 153(6):2417–28.
14. Poggi A, Rubartelli A, Moretta L, Zocchi MR. Expression and function of NKRP1A molecule on human monocytes and dendritic cells. Eur J Immunol (1997) 27(11):2965–70. doi: 10.1002/eji.1830271132
15. Kurioka A, Klenerman P, Willberg CB. Innate-like CD8+ T-cells and NK cells: converging functions and phenotypes. Immunology (2018) 154(4):547–56. doi: 10.1111/imm.12925
16. Sallusto F, Lenig D, Forster R, Lipp M, Lanzavecchia A. Two subsets of memory T lymphocytes with distinct homing potentials and effector functions. Nature (1999) 401(6754):708–12. doi: 10.1038/44385
17. Nicol B, Salou M, Vogel I, Garcia A, Dugast E, Morille J, et al. An intermediate level of CD161 expression defines a novel activated, inflammatory, and pathogenic subset of CD8(+) T cells involved in multiple sclerosis. J Autoimmun (2018) 88:61–74. doi: 10.1016/j.jaut.2017.10.005
18. Fergusson JR, Huhn MH, Swadling L, Walker LJ, Kurioka A, Llibre A, et al. CD161(int)CD8+ T cells: a novel population of highly functional, memory CD8+ T cells enriched within the gut. Mucosal Immunol (2016) 9(2):401–13. doi: 10.1038/mi.2015.69
19. Le Bourhis L, Martin E, Peguillet I, Guihot A, Froux N, Core M, et al. Antimicrobial activity of mucosal-associated invariant T cells. Nat Immunol (2010) 11(8):701–8. doi: 10.1038/ni.1890
20. Kurioka A, Ussher JE, Cosgrove C, Clough C, Fergusson JR, Smith K, et al. MAIT cells are licensed through granzyme exchange to kill bacterially sensitized targets. Mucosal Immunol (2015) 8(2):429–40. doi: 10.1038/mi.2014.81
21. Terabe M, Tagaya Y, Zhu Q, Granger L, Roederer M, Waldmann TA, et al. IL-15 expands unconventional CD8alphaalphaNK1.1+ T cells but not Valpha14Jalpha18+ NKT cells. J Immunol (2008) 180(11):7276–86. doi: 10.4049/jimmunol.180.11.7276
22. Huber M, Heink S, Grothe H, Guralnik A, Reinhard K, Elflein K, et al. Th17-like developmental process leads to CD8(+) Tc17 cells with reduced cytotoxic activity. A Eur J Immunol (2009) 39(7):1716–25. doi: 10.1002/eji.200939412
23. Hamada H, Garcia-Hernandez Mde L, Reome JB, Misra SK, Strutt TM, McKinstry KK, et al. Tc17, a unique subset of CD8 T cells that can protect against lethal influenza challenge. J Immunol (2009) 182(6):3469–81. doi: 10.4049/jimmunol.0801814
24. Mackay LK, Kallies A. Transcriptional Regulation of Tissue-Resident Lymphocytes. Trends Immunol (2017) 38(2):94–103. doi: 10.1016/j.it.2016.11.004
25. O’Keeffe J, Doherty DG, Kenna T, Sheahan K, O’Donoghue DP, Hyland JM, et al. Diverse populations of T cells with NK cell receptors accumulate in the human intestine in health and in colorectal cancer. Eur J Immunol (2004) 34(8):2110–9. doi: 10.1002/eji.200424958
26. Wolk K, Kunz S, Witte E, Friedrich M, Asadullah K, Sabat R. IL-22 increases the innate immunity of tissues. Immunity (2004) 21(2):241–54. doi: 10.1016/j.immuni.2004.07.007
27. Salou M, Franciszkiewicz K, Lantz O. MAIT cells in infectious diseases. Curr Opin Immunol (2017) 48:7–14. doi: 10.1016/j.coi.2017.07.009
28. Leng T, Akther HD, Hackstein CP, Powell K, King T, Friedrich M, et al. TCR and Inflammatory Signals Tune Human MAIT Cells to Exert Specific Tissue Repair and Effector Functions. Cell Rep (2019) 28(12):3077–91.e5. doi: 10.1016/j.celrep.2019.08.050
29. Lamichhane R, Schneider M, de la Harpe SM, Harrop TWR, Hannaway RF, Dearden PK, et al. TCR- or Cytokine-Activated CD8(+) Mucosal-Associated Invariant T Cells Are Rapid Polyfunctional Effectors That Can Coordinate Immune Responses. Cell Rep (2019) 28(12):3061–76. doi: 10.1016/j.celrep.2019.08.054
30. Hinks TSC, Marchi E, Jabeen M, Olshansky M, Kurioka A, Pediongco TJ, et al. Activation and In Vivo Evolution of the MAIT Cell Transcriptome in Mice and Humans Reveals Tissue Repair Functionality. Cell Rep (2019) 28(12):3249–62.e5. doi: 10.1016/j.celrep.2019.07.039
31. Dusseaux M, Martin E, Serriari N, Peguillet I, Premel V, Louis D, et al. Human MAIT cells are xenobiotic-resistant, tissue-targeted, CD161hi IL-17-secreting T cells. Blood (2011) 117(4):1250–9. doi: 10.1182/blood-2010-08-303339
32. Freeman CM, Curtis JL, Chensue SW. CC chemokine receptor 5 and CXC chemokine receptor 6 expression by lung CD8+ cells correlates with chronic obstructive pulmonary disease severity. Am J Pathol (2007) 171(3):767–76. doi: 10.2353/ajpath.2007.061177
33. Reboldi A, Coisne C, Baumjohann D, Benvenuto F, Bottinelli D, Lira S, et al. C-C chemokine receptor 6-regulated entry of TH-17 cells into the CNS through the choroid plexus is required for the initiation of EAE. Nat Immunol (2009) 10(5):514–23. doi: 10.1038/ni.1716
34. Li Z, Wu Y, Wang C, Zhang M. Mouse CD8(+)NKT-like cells exert dual cytotoxicity against mouse tumor cells and myeloid-derived suppressor cells. Cancer Immunol Immunother (2019) 68(8):1303–15. doi: 10.1007/s00262-019-02363-3
35. Veldhoen M, Hirota K, Westendorf AM, Buer J, Dumoutier L, Renauld JC, et al. The aryl hydrocarbon receptor links TH17-cell-mediated autoimmunity to environmental toxins. Nature (2008) 453(7191):106–9. doi: 10.1038/nature06881
36. Iizuka K, Naidenko OV, Plougastel BF, Fremont DH, Yokoyama WM. Genetically linked C-type lectin-related ligands for the NKRP1 family of natural killer cell receptors. Nat Immunol (2003) 4(8):801–7. doi: 10.1038/ni954
37. Carlyle JR, Jamieson AM, Gasser S, Clingan CS, Arase H, Raulet DH. Missing self-recognition of Ocil/Clr-b by inhibitory NKR-P1 natural killer cell receptors. Proc Natl Acad Sci U S A (2004) 101(10):3527–32. doi: 10.1073/pnas.0308304101
38. Rosen DB, Cao W, Avery DT, Tangye SG, Liu YJ, Houchins JP, et al. Functional consequences of interactions between human NKR-P1A and its ligand LLT1 expressed on activated dendritic cells and B cells. J Immunol (2008) 180(10):6508–17. doi: 10.4049/jimmunol.180.10.6508
39. Aldemir H, Prod’homme V, Dumaurier MJ, Retiere C, Poupon G, Cazareth J, et al. Cutting edge: lectin-like transcript 1 is a ligand for the CD161 receptor. J Immunol (2005) 175(12):7791–5. doi: 10.4049/jimmunol.175.12.7791
40. Huarte E, Cubillos-Ruiz JR, Nesbeth YC, Scarlett UK, Martinez DG, Engle XA, et al. PILAR is a novel modulator of human T-cell expansion. Blood (2008) 112(4):1259–68. doi: 10.1182/blood-2007-12-130773
41. Kesselring R, Thiel A, Pries R, Wollenberg B. The number of CD161 positive Th17 cells are decreased in head and neck cancer patients. Cell Immunol (2011) 269(2):74–7. doi: 10.1016/j.cellimm.2011.03.026
42. Germain C, Meier A, Jensen T, Knapnougel P, Poupon G, Lazzari A, et al. Induction of lectin-like transcript 1 (LLT1) protein cell surface expression by pathogens and interferon-gamma contributes to modulate immune responses. J Biol Chem (2011) 286(44):37964–75. doi: 10.1074/jbc.M111.285312
43. Germain C, Guillaudeux T, Galsgaard ED, Hervouet C, Tekaya N, Gallouet AS, et al. Lectin-like transcript 1 is a marker of germinal center-derived B-cell non-Hodgkin’s lymphomas dampening natural killer cell functions. Oncoimmunology (2015) 4(8):e1026503. doi: 10.1080/2162402X.2015.1026503
44. Rosen DB, Bettadapura J, Alsharifi M, Mathew PA, Warren HS, Lanier LL. Cutting edge: lectin-like transcript-1 is a ligand for the inhibitory human NKR-P1A receptor. J Immunol (2005) 175(12):7796–9. doi: 10.4049/jimmunol.175.12.7796
45. Exley M, Porcelli S, Furman M, Garcia J, Balk S. CD161 (NKR-P1A) costimulation of CD1d-dependent activation of human T cells expressing invariant V alpha 24 J alpha Q T cell receptor alpha chains. J Exp Med (1998) 188(5):867–76. doi: 10.1084/jem.188.5.867
46. Le Bourhis L, Mburu YK, Lantz O. MAIT cells, surveyors of a new class of antigen: development and functions. Curr Opin Immunol (2013) 25(2):174–80. doi: 10.1016/j.coi.2013.01.005
47. Pozo D, Vales-Gomez M, Mavaddat N, Williamson SC, Chisholm SE, Reyburn H. CD161 (human NKR-P1A) signaling in NK cells involves the activation of acid sphingomyelinase. J Immunol (2006) 176(4):2397–406. doi: 10.4049/jimmunol.176.4.2397
48. Bai A, Robson S. Beyond ecto-nucleotidase: CD39 defines human Th17 cells with CD161. Purinerg Signal (2015) 11(3):317–9. doi: 10.1007/s11302-015-9457-4
49. Bai A, Moss A, Kokkotou E, Usheva A, Sun X, Cheifetz A, et al. CD39 and CD161 modulate Th17 responses in Crohn’s disease. J Immunol (2014) 193(7):3366–77. doi: 10.4049/jimmunol.1400346
50. Decker WK, Xing D, Li S, Robinson SN, Yang H, Yao X, et al. Double loading of dendritic cell MHC class I and MHC class II with an AML antigen repertoire enhances correlates of T-cell immunity in vitro via amplification of T-cell help. Vaccine (2006) 24(16):3203–16. doi: 10.1016/j.vaccine.2006.01.029
51. Decker WK, Shpall EJ. Progress in dendritic cell immunotherapy: elucidating the enigma of Th-1 polarization. Hum Vaccin (2008) 4(2):162–4. doi: 10.4161/hv.4.2.5091
52. Halpert MM, Konduri V, Liang D, Vazquez-Perez J, Hofferek CJ, Weldon SA, et al. MHC class I and II peptide homology regulates the cellular immune response. FASEB J (2020) 34(6):8082-101. doi: 10.1096/fj.201903002R
53. Konduri V, Li D, Halpert MM, Liang D, Liang Z, Chen Y, et al. Chemo-immunotherapy mediates durable cure of orthotopic Kras(G12D)/p53(-/-) pancreatic ductal adenocarcinoma. Oncoimmunology (2016) 5(9):e1213933. doi: 10.1080/2162402X.2016.1213933
54. Ruiz AL, Soudja SM, Deceneux C, Lauvau G, Marie JC. NK1.1+ CD8+ T cells escape TGF-beta control and contribute to early microbial pathogen response. Nat Commun (2014) 5:5150. doi: 10.1038/ncomms6150
55. Seregin SS, Chen GY, Laouar Y. Dissecting CD8+ NKT Cell Responses to Listeria Infection Reveals a Component of Innate Resistance. J Immunol (2015) 195(3):1112–20. doi: 10.4049/jimmunol.1500084
56. Pied S, Roland J, Louise A, Voegtle D, Soulard V, Mazier D, et al. Liver CD4-CD8- NK1.1+ TCR alpha beta intermediate cells increase during experimental malaria infection and are able to exhibit inhibitory activity against the parasite liver stage in vitro. J Immunol (2000) 164(3):1463–9. doi: 10.4049/jimmunol.164.3.1463
57. Muxel SM, Freitas do Rosario AP, Sardinha LR, Castillo-Mendez SI, Zago CA, Rodriguez-Malaga SM, et al. Comparative analysis of activation phenotype, proliferation, and IFN-gamma production by spleen NK1.1(+) and NK1.1(-) T cells during Plasmodium chabaudi AS malaria. J Interferon Cytokine Res (2010) 30(6):417–26. doi: 10.1089/jir.2009.0095
58. Alsuliman A, Muftuoglu M, Khoder A, Ahn YO, Basar R, Verneris MR, et al. A subset of virus-specific CD161(+) T cells selectively express the multidrug transporter MDR1 and are resistant to chemotherapy in AML. Blood (2017) 129(6):740–58. doi: 10.1182/blood-2016-05-713347
59. Rowan AG, Fletcher JM, Ryan EJ, Moran B, Hegarty JE, O’Farrelly C, et al. Hepatitis C virus-specific Th17 cells are suppressed by virus-induced TGF-beta. J Immunol (2008) 181(7):4485–94. doi: 10.4049/jimmunol.181.7.4485
60. Cosmi L, De Palma R, Santarlasci V, Maggi L, Capone M, Frosali F, et al. Human interleukin 17-producing cells originate from a CD161+CD4+ T cell precursor. J Exp Med (2008) 205(8):1903–16. doi: 10.1084/jem.20080397
61. Kleinschek MA, Boniface K, Sadekova S, Grein J, Murphy EE, Turner SP, et al. Circulating and gut-resident human Th17 cells express CD161 and promote intestinal inflammation. J Exp Med (2009) 206(3):525–34. doi: 10.1084/jem.20081712
62. Lock C, Hermans G, Pedotti R, Brendolan A, Schadt E, Garren H, et al. Gene-microarray analysis of multiple sclerosis lesions yields new targets validated in autoimmune encephalomyelitis. Nat Med (2002) 8(5):500–8. doi: 10.1038/nm0502-500
63. Tzartos JS, Friese MA, Craner MJ, Palace J, Newcombe J, Esiri MM, et al. Interleukin-17 production in central nervous system-infiltrating T cells and glial cells is associated with active disease in multiple sclerosis. Am J Pathol (2008) 172(1):146–55. doi: 10.2353/ajpath.2008.070690
64. Annibali V, Ristori G, Angelini DF, Serafini B, Mechelli R, Cannoni S, et al. CD161(high)CD8+T cells bear pathogenetic potential in multiple sclerosis. Brain (2011) 134(Pt 2):542–54. doi: 10.1093/brain/awq354
65. International Multiple Sclerosis Genetics C, Hafler DA, Compston A, Sawcer S, Lander ES, Daly MJ, et al. Risk alleles for multiple sclerosis identified by a genomewide study. N Engl J Med (2007) 357(9):851–62. doi: 10.1056/NEJMoa073493
66. Acquaviva M, Bassani C, Sarno N, Dalla Costa G, Romeo M, Sangalli F, et al. Loss of Circulating CD8+ CD161(high) T Cells in Primary Progressive Multiple Sclerosis. Front Immunol (2019) 10:1922. doi: 10.3389/fimmu.2019.01922
67. Park Y, Lim J, Kim SY, Kwon GC, Koo SH, Kim J. Changes of frequency and expression level of CD161 in CD8(+) T cells and Natural killer T cells in Peripheral Blood of Patients with Systemic Lupus Erythematosus. Microbiol Immunol (2020) 64(7):532–9. doi: 10.1111/1348-0421.12798
68. Baker J, Verneris MR, Ito M, Shizuru JA, Negrin RS. Expansion of cytolytic CD8(+) natural killer T cells with limited capacity for graft-versus-host disease induction due to interferon gamma production. Blood (2001) 97(10):2923–31. doi: 10.1182/blood.V97.10.2923
69. Verneris MR, Ito M, Baker J, Arshi A, Negrin RS, Shizuru JA. Engineering hematopoietic grafts: purified allogeneic hematopoietic stem cells plus expanded CD8+ NK-T cells in the treatment of lymphoma. Biol Blood Marrow Transpl (2001) 7(10):532–42. doi: 10.1016/S1083-8791(01)70014-6
70. Janakiram NB, Mohammed A, Bryant T, Ritchie R, Stratton N, Jackson L, et al. Loss of natural killer T cells promotes pancreatic cancer in LSL-Kras(G12D/+) mice. Immunology (2017) 152(1):36–51. doi: 10.1111/imm.12746
71. Wingender G, Berg M, Jungerkes F, Diehl L, Sullivan BA, Kronenberg M, et al. Immediate antigen-specific effector functions by TCR-transgenic CD8+ NKT cells. Eur J Immunol (2006) 36(3):570–82. doi: 10.1002/eji.200535461
72. Gentles AJ, Newman AM, Liu CL, Bratman SV, Feng W, Kim D, et al. The prognostic landscape of genes and infiltrating immune cells across human cancers. Nat Med (2015) 21(8):938–45. doi: 10.1038/nm.3909
73. Braud VM, Biton J, Becht E, Knockaert S, Mansuet-Lupo A, Cosson E, et al. Expression of LLT1 and its receptor CD161 in lung cancer is associated with better clinical outcome. Oncoimmunology (2018) 7(5):e1423184. doi: 10.1080/2162402X.2017.1423184
74. Pleshkan VV, Zinov’eva MV, Vinogradova TV, Sverdlov ED. Transcription of the KLRB1 gene is suppressed in human cancer tissues. Mol Gen Mikrobiol Virusol (2007) (4):3–7. doi: 10.3103/S0891416807040015
75. Welters MJP, Ma W, Santegoets S, Goedemans R, Ehsan I, Jordanova ES, et al. Intratumoral HPV16-Specific T Cells Constitute a Type I-Oriented Tumor Microenvironment to Improve Survival in HPV16-Driven Oropharyngeal Cancer. Clin Cancer Res (2018) 24(3):634–47. doi: 10.1158/1078-0432.CCR-17-2140
76. Li Z, Zheng B, Qiu X, Wu R, Wu T, Yang S, et al. The identification and functional analysis of CD8+PD-1+CD161+ T cells in hepatocellular carcinoma. NPJ Precis Oncol (2020) 4:28. doi: 10.1038/s41698-020-00133-4
77. Pillai AB, George TI, Dutt S, Teo P, Strober S. Host NKT cells can prevent graft-versus-host disease and permit graft antitumor activity after bone marrow transplantation. J Immunol (2007) 178(10):6242–51. doi: 10.4049/jimmunol.178.10.6242
78. Ngai H, Tian G, Courtney AN, Ravari SB, Guo L, Liu B, et al. IL-21 Selectively Protects CD62L(+) NKT Cells and Enhances Their Effector Functions for Adoptive Immunotherapy. J Immunol (2018) 201(7):2141–53. doi: 10.4049/jimmunol.1800429
79. Iliopoulou EG, Karamouzis MV, Missitzis I, Ardavanis A, Sotiriadou NN, Baxevanis CN, et al. Increased frequency of CD4+ cells expressing CD161 in cancer patients. Clin Cancer Res (2006) 12(23):6901–9. doi: 10.1158/1078-0432.CCR-06-0977
80. de Lalla C, Rinaldi A, Montagna D, Azzimonti L, Bernardo ME, Sangalli LM, et al. Invariant NKT cell reconstitution in pediatric leukemia patients given HLA-haploidentical stem cell transplantation defines distinct CD4+ and CD4- subset dynamics and correlates with remission state. J Immunol (2011) 186(7):4490–9. doi: 10.4049/jimmunol.1003748
81. Morice WG, Kurtin PJ, Leibson PJ, Tefferi A, Hanson CA. Demonstration of aberrant T-cell and natural killer-cell antigen expression in all cases of granular lymphocytic leukaemia. Br J Haematol (2003) 120(6):1026–36. doi: 10.1046/j.1365-2141.2003.04201.x
Keywords: KLRB1, CD161, effector memory, T-cell, TH1 polarization
Citation: Konduri V, Oyewole-Said D, Vazquez-Perez J, Weldon SA, Halpert MM, Levitt JM and Decker WK (2021) CD8+CD161+ T-Cells: Cytotoxic Memory Cells With High Therapeutic Potential. Front. Immunol. 11:613204. doi: 10.3389/fimmu.2020.613204
Received: 01 October 2020; Accepted: 09 December 2020;
Published: 01 February 2021.
Edited by:
Luca Gattinoni, Regensburg Center for Interventional Immunology (RCI), GermanyReviewed by:
Edwin Leeansyah, Tsinghua-Berkeley Shenzhen Institute, ChinaVijayakumar Velu, Emory University, United States
Copyright © 2021 Konduri, Oyewole-Said, Vazquez-Perez, Weldon, Halpert, Levitt and Decker. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: William K. Decker, decker@bcm.edu