 Katherine Shaw
Katherine Shaw Mylène Therrien
Mylène Therrien Chunyu Lu1
Chunyu Lu1 Xiaochun Liu
Xiaochun Liu Vance L. Trudeau
Vance L. Trudeau- 1Department of Biology, University of Ottawa, Ottawa, ON, Canada
- 2State Key Laboratory of Biocontrol, Institute of Aquatic Economic Animals and Guangdong Province Key Laboratory for Aquatic Economic Animals, School of Life Sciences, Sun Yat-Sen University, Guangzhou, China
Aromatase (Cyp19a1) is the steroidogenic enzyme that converts androgens into bioactive estrogens, and hence is in a pivotal position to mediate reproduction and sexual behavior. In teleosts, there are two aromatase paralogs: cyp19a1a that is highly expressed in granulosa and Leydig cells in the gonads with critical function in sexual differentiation of the ovary, and cyp19a1b that is highly expressed in radial glial cells in the brain with unknown roles in reproduction. Cyp19a1-/- mutant zebrafish lines were used to investigate the importance of the cyp19a1 paralogs for spawning behavior and offspring survival and early development. Mutation of cyp19a1b was found to increase the latency to the first oviposition in females. Mutation of cyp19a1b in females also increased the number of eggs spawned; however, significantly more progeny died during early development resulting in no net increase in female fecundity. This finding suggests a higher metabolic cost of reproduction in cyp19a1b-/- mutant females. In males, the combined mutation of both cyp19a1 paralogs resulted in significantly lower progeny survival rates, indicating a critical function of cyp19a1 during early larval development. These data establish the specific importance of cyp19a1b for female spawning behavior and the importance of the cyp19a1 paralogs for early larval survival.
1 Introduction
Androgens and estrogens are two major groups of sex steroids that play critical roles in vertebrates to coordinate the physiology and behavior of an individual with its environment. Aromatase (Cyp19a1) is the terminal steroidogenic enzyme that converts the aromatizable androgens, testosterone and androstenedione, into estradiol (E2) and estrone, respectively. In birds and most mammals, there is only one aromatase gene, cyp19a1, whose differential tissue distribution is attributed to differences in splicing of 5’ untranslated promoter regions across tissues (1). In contrast, teleosts possess two distinct cyp19a1 genes, cyp19a1a and cyp19a1b, with differential tissue distribution. This is due to distinct regulatory elements in their promoter regions and differences in the presence of required transcription factors in specific cell types (2). In many teleosts, cyp19a1a is the most highly expressed aromatase in granulosa and Leydig cells in the gonads; while cyp19a1b is expressed at much higher levels in the brain, and specifically in radial glial cells (RGCs; 3).
The ovaries are a major source of estrogens that are released into systemic circulation to prepare the body for reproduction (4). Loss of aromatase expression significantly impairs female fertility and fecundity, at least in part, through loss of ovarian estrogen production. For example, total aromatase knockout (tAroKO) female mice, which lack whole body aromatase expression, are infertile due to disrupted follilculogenesis and ovulation failure (5). In teleosts, short term chemical inhibition of aromatase reduces female fertility and fecundity through impaired oocyte development and reduces plasma vitellogenin levels (6–8). Long term aromatase inhibitor treatment induces more dramatic effects such as ovarian retraction followed by testis formation resulting in female-to-male sex change (9, 10). Since chemical inhibitors are non-selective for the two Cyp19a1 isoforms, it only recently became possible to begin identifying their independent contributions to reproduction via the creation of zebrafish cyp19a1-/- mutant lines. It was discovered that cyp19a1a expression is critical for sexual differentiation of the ovaries while cyp19a1b expression is not required for this process (11–13). Though cyp19a1a is not required for testis differentiation, it is expressed at low levels and differences have been observed in the importance of its expression for male fertility in mice and teleosts. For example, the testis of male tAroKO mice were found to have arrested germ cell development at the spermatid stage as well as impaired sperm motility (14–17). In contrast, there were no developmental abnormalities observed in the testis of zebrafish cyp19a1a-/- mutants compared to wild-type (WT) males (18). Rather, the testis of cyp19a1a-/- mutants had more spermatozoa and higher levels of spermatogenesis-related genes, and these mutant males displayed normal fertility levels. These findings reveal critical roles of ovarian aromatase expression for female fertility and fecundity in mice and teleosts, whilst differences have been observed for testicular aromatase importance in male fertility, with cyp19a1a being dispensable for male teleost fertility.
A second major source of estrogens is the brain. In birds and mammals, the cyp19a1 gene contains a brain-specific promoter region that specifies constitutive neuronal expression (19–22). Teleosts have a second paralog, cyp19a1b, that is expressed exclusively in RGCs due to the presence of G x RE, the glial x responsive element, in the promoter region (2). Increasing evidence has identified important roles for brain-derived estrogens in reproduction. For example, both female and male total tAroKO mice display reduced sexual behavior (23–26) and impaired olfactory discrimination (25, 27) that together suggests an involvement of brain estrogens in social recognition. The recent creation of a male brain aromatase knockout (bAroKO) mouse line identified the important role of brain-derived estrogens in male sexual behavior. Male bAroKO mice were found to have a significantly longer latency to the first mount event and a trend, though not significant, towards a greater latency to the first intromission event when paired with a hormonally primed female compared to WT males in sexual behavior assays (28). Female bAroKO mice have not yet been studied. In teleosts, studies have identified impairments in social recognition following chemical aromatase inhibition, such as reduced dominant male aggression in social behavior assays (29, 30). There has been comparatively less study, however, of effects on sexual behavior. This is likely due, at least in part, to the inability to identify specific Cyp19a1b- versus Cyp19a1a-induced effects on reproduction with the use of chemical aromatase inhibitors. There is strong evidence, however, to suggest that brain-derived estrogens likely play an important role in teleost sexual behaviors. Firstly, cyp19a1b is expressed in numerous brain regions important for sexual behavior (31). Secondly, cyp19a1b is a known estrogen-regulated gene due to the presence of an estrogen response element in its promoter region (32). This observation suggests that increased systemic estrogen levels, via high ovarian Cyp19a1a activity, likely drive increased cyp19a1b expression to prepare the brain for sexual behavior at the time in which the ovaries are prepared for reproduction.
Here we report that mutant cyp19a1b-/- females exhibit a longer latency to oviposition and release a significantly higher number of eggs during spawning compared to WT females. However, there was a higher larval mortality, resulting in no net fecundity differences between female cyp19a1b-/- mutant and WT pairings. A significantly higher larval mortality rate was found in progeny from cyp19a1a-/-;cyp19a1b-/- mutant male pairings. These data reveal the importance of cyp19a1b for zebrafish reproduction.
2 Materials and methods
2.1 Experimental animals
Procedures used in this study were approved by the University of Ottawa Animal Care Committee and follow the guidelines of the Canadian Council on Animal Care for the use of animals in research. All fish were reared at the University of Ottawa Aquatics Facility according to standard housing procedures. The cyp19a1-/- mutant lines and WT zebrafish used in the experiments were all derived from a parental zebrafish AB strain to ensure identical genetic backgrounds amongst the groups for assessing the effects of cyp19a1 mutation on spawning behavior. The cyp19a1-/- mutant lines were generated using the transcription activator-like effector nucleases (TALEN) genome editing system to create indel mutations at target sites in each of the cyp19a1 paralogs producing frame-shift mutations (13). Mutation of cyp19a1a impairs sexual differentiation of the ovary resulting in all male populations of cyp19a1a-/- and cyp19a1a-/-;cyp19a1b-/- mutant lines, i.e., only male mutants can be tested in these lines. These males have significantly lower serum E2 levels compared to WT males due to the contributions of gonadal aromatase expression to circulating estrogen levels. Mutation of cyp19a1b does not affect the sex ratio of the mutant line allowing testing of cyp19a1b-/- mutant effects on both males and females, and serum E2 levels in these fish are not significantly different from their WT counterparts. Fish were housed in 10-L tanks with dechloraminated water at 28°C and maintained on a 14:10 light-dark cycle and fed twice daily. All fish tested in this experiment were between the ages of 5-11 months post-fertilization and had no previous experience in the sexual behavior assay; with males and females separated into same-sex tanks at sexual maturation to prevent sexual interactions.
2.2 Behavioral tests
For all trials, mutant fish were size-matched within 2 mm body length (<10% body length difference) to an opposite sex WT fish. The evening before each experiment, paired fish were transferred to a 1-L testing tank containing an insert at the bottom for egg collection and a divider in the middle of the tank to keep the male and female separate before testing. The testing pair was allowed to acclimate overnight in a ZebraCube (Viewpoint Behavior Technology, Inc., Lyon, France) with the camera (either a Panasonic 16GB HC-V700M Full HD camcorder, Osaka, Japan or a Canon VIXIA HF R800 camcorder, Tokyo, Japan) present. The next morning, the pair was transferred to a new 1-L testing tank containing clean system water and the divider was removed and video recording started at 0900h (lights on). The fish pair was allowed to interact for 150 min, which was the most appropriate time determined in preliminary trials to capture the full timing of spawning behavior, particularly in the cyp19a1b-/- mutant lines. Videos were coded by date to ensure that the observer was blind to the treatment groups during viewing and video analysis using VLC media player (https://www.videolan.org/). Oviposition events, which represent spawning behavior, were selected as the most appropriate measure to identify changes in sexual behavior in this study. This decision was made based on the similar descriptive measures for the ethograms of sexual and aggressive behaviors in zebrafish (33, 34) that prevented unambiguous identification of motivational state during initial interactions in the video recordings. Since there is strong evidence identifying roles of brain-derived E2, and therefore, brain aromatase (i.e., Cyp19a1b), in both sexual and aggressive behaviors in vertebrates (29–31, 35), it was important to select a definitive measure of sexual behavior for analysis in this study. Oviposition events were characterized and were identified in the video recordings as the timing of gamete release during physical interaction. The oviposition times were manually recorded in an Excel spreadsheet for the later determination of the time to first and last spawning events, as well as the total number of spawning events in a trial.
2.3 Egg collection and eleutheroembryo rearing
Eggs were obtained from natural mating of a cyp19a1-/- mutant or WT fish paired with an opposite sex WT fish using the methods described above. Following the careful removal of the eggs from the bottom of the tank, they were then rinsed with clean system water and transferred to a Petri dish containing E3 medium at a density of 3 embryos/mL (36). At 4 h post-fertilization, unfertilized eggs were counted and removed from the Petri dish for the assessment of fertilization rates. All embryos were raised in an incubator at 28°C for the duration of the experiment. The following day (1-day post-fertilization; 1 dpf), any dead embryos were counted and recorded, then removed, following which E3 medium was replaced, and dishes were returned to the incubator. On 2, 3, and 4 dpf, any dead eggs were again counted and removed. Eleutheroembryos (i.e., hatched embryos) were counted and placed into separate Petri dishes containing E3 medium for measurements, and the medium was then replaced for the incubation of the remaining live eggs.
2.4 Zebrafish eleutheroembryo morphometrics
On day 2 post-fertilization, five eleutheroembryos were randomly selected from each clutch for measurement. The five measurements were later averaged to obtain a single value to represent the clutch at the given time. The eleutheroembryos were anesthetized with tricaine methanesulfonate (100 mg/L) and positioned under a dissecting microscope (Wild M10, Leica, Wetzlar, Germany) fitted with a phone mount and iPhone XS. Eleutheroembryos were gently positioned on their side to produce a lateral view for measurement of body length, eye area, and yolk sac area in photographs. A ruler was positioned under the microscope and photographed at the same magnification as used for Petri dishes to serve as a scale bar for image analysis. Following completion of the measurements, the eleutheroembryos were euthanized by immersion in an ice water bath.
2.5 Enzyme-linked immunosorbent assay
Female cyp19a1b-/- and WT fish were euthanized between 9h00 – 10h00 in an ice water bath, and brains and ovaries were immediately dissected, placed into individual labelled tubes, weighed, and tubes were then placed on ice. Homogenization buffer (90% methanol) was added to each tube, 150 µL per brain and 1000 µL per ovary sample, and samples were then sonicated for tissue dispersion and steroid release into solution. Samples were centrifuged (4°C, 13,200 rpm) for 10 min, and the supernatant was carefully removed and transferred to a new labelled tube. The samples were then evaporated to dryness (Labconco Centrivap Centrifugal Vacuum Concentrator, Model #7810014, 45°C, 1 h), and stored at 4°C overnight. The following day, 100 µL resuspension buffer (0.2% formic acid, 5% acetonitrile in water) was added to each sample tube, vortexed, and the tubes were then placed in a sonic water bath for 15 min to resuspend the samples. Following resuspension, the samples were run in C-18 solid-phase extraction columns (Catalog #r10.aq, Dr. Maisch HPLC GmbH, Ammerbuch-Entringen, Germany; >85% recovery rate in liquid chromatography tandem mass spectrometry) and the eluted samples were then evaporated to dryness (45°C, 3 h). The evaporated steroid residue was then resuspended in ELISA buffer, 200 µL for brain samples and 1000 µL for ovary samples, for 24 h at 4°C with intermittent vortexing prior to testing. Estradiol levels were measured using enzyme-linked immunoassay test kits (ELISA; Catalog #501890, Cayman Chemical, Ann Arbor, MI, USA) according to the manufacturer’s instructions. This assay has been extensively tested in teleost species (37–42) with an assay range of 0.61 – 10,000 pg/mL and sensitivity limit of 20 pg/mL. This E2 assay has very low levels of cross reactivity to other steroids such as cortisol, progesterone and testosterone (< 0.01%). All samples were run in duplicate on a single assay plate and only samples with intra-assay CVs < 10% were used in analyses.
2.6 Image analysis
Photos were analyzed using ImageJ (v1.53) software. All images were analyzed by the same observer to ensure consistent measurements (i.e., to prevent inter-observer variability), and each measurement was repeated three times and the average value was calculated. Each image was measured for body length, eye area, and yolk sac area. Body length consisted of a straight-line measurement from the posterior tip of the notochord to the most anterior tip of the head passing through the eye (Figure 1). Eye and yolk sac areas were measured according to Martínez et al. (36) and traced with the freehand tracing tool in ImageJ (Figure 1).
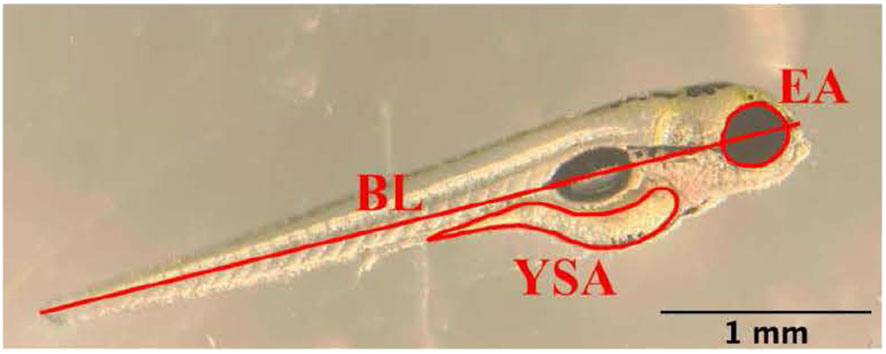
Figure 1 Depiction of the measurement of body length (BL), eye area (EA), and yolk sac area (YSA) in an eleutheroembryo on day 2 post-fertilization. Scale bar = 1 mm.
2.7 Statistical analyses
Statistical analyses were conducted using GraphPad Prism v9 (GraphPad Software, Inc., La Jolla, CA, USA) with normality and homoscedasticity assessed using Shapiro-Wilk and Levene’s tests, respectively. For the behavior and eleutheroembryo analyses, normally distributed data were analyzed by either Student’s T test or One-Way ANOVA followed by Dunnett’s multiple comparisons tests for pairwise comparisons. Data that were not normally distributed were analyzed using either Mann-Whitney U or Kruskal-Wallis tests followed by Dunn’s multiple comparisons tests for pairwise comparisons. Data are presented as boxplots with the horizontal lines representing mean or median values, boxes representing interquartile ranges, and whiskers representing min-max values. For the E2 measurements, data were log-transformed and analyzed using a Two-Way ANOVA followed by Tukey’s multiple comparisons tests. For all data, significance is defined at p < 0.05 and all tests were assessed as two-tailed.
3 Results
3.1 Time to the first oviposition event
Female cyp19a1b-/- mutant fish took significantly more time to the first oviposition event compared to WT females (U(17,17)=70.50, p=0.0096; Figure 2A). On average, cyp19a1b-/- females took 4.1 times longer to the first oviposition event compared to WT females. There were no significant differences in the time to the first oviposition event among any of the male genotypes (H(3)=7.005, p=0.0717; Figure 2B).
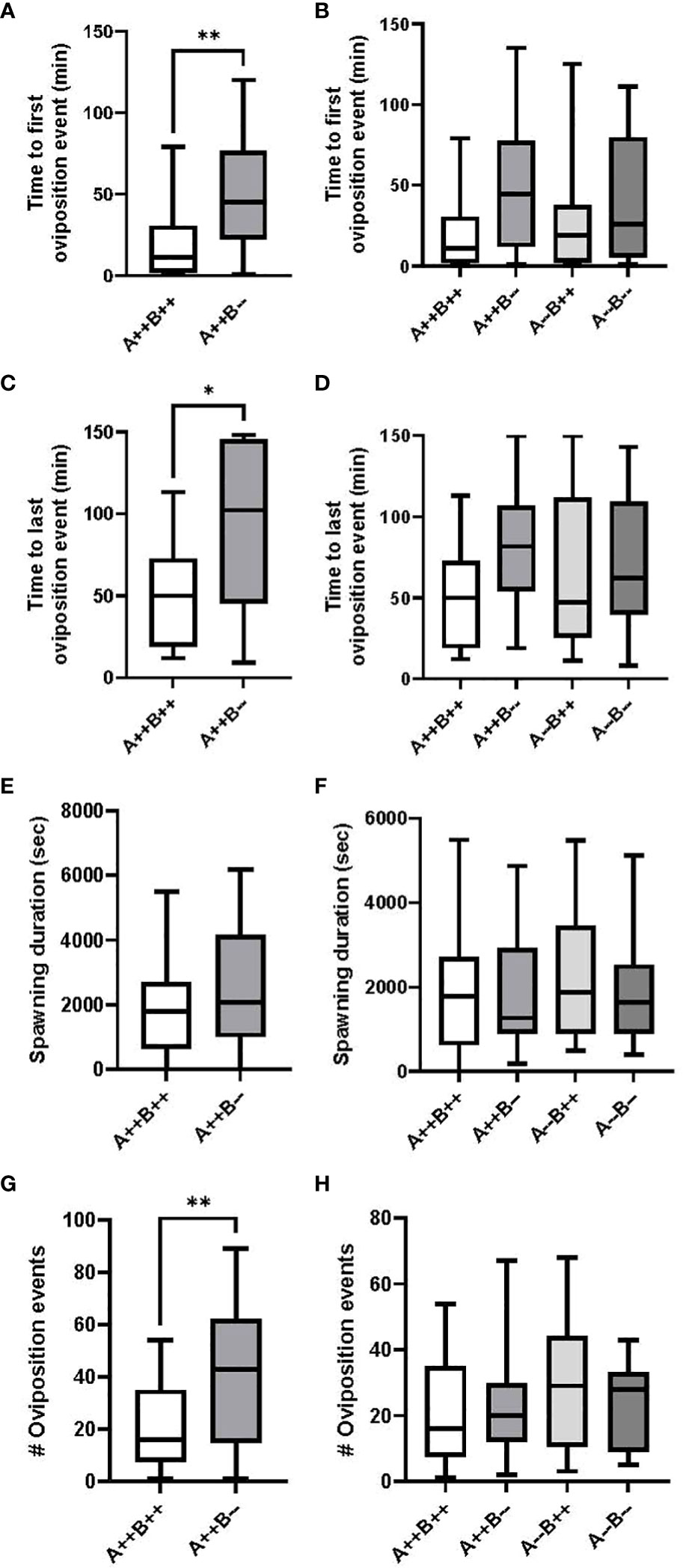
Figure 2 Timing and number of oviposition events during zebrafish pairwise mating trials. Test females (n=17 pairs) are displayed on the left (A, C, E, G) and test males (n=17-18 pairs) are displayed on the right (B, D, F, H). Mann-Whitney U (A, C, G), Student’s T (E), Kruskal-Wallis (B) and One-Way ANOVA (D, F, H) tests were performed. Horizontal lines represent mean or median values, boxes represent interquartile ranges, and whiskers represent min-max values. Key to genotypes: A=cyp19a1a, B=cyp19a1b. Asterisks denote p<0.05 (*) and p<0.01 (**).
3.2 Time to the last oviposition event
Female cyp19a1b-/- mutant fish took significantly more time to the last oviposition event compared to WT females (U(17,17)=81.50, p=0.0293; Figure 2C). On average, the last oviposition event in cyp19a1b-/- female pairings occurred 2 times later than in WT pairings. There were no significant differences in the time to the last oviposition event among any of the male genotypes (F(3)=1.657, p=0.1850; Figure 2D).
3.3 Spawning duration
There were no significant differences in the spawning duration between the cyp19a1b-/- mutant and WT female groups (T(32)=1.130, p=0.2668; Figure 2E) or between any of the mutant and WT male groups (F(3)=0.3184, p=0.8120; Figure 2F).
3.4 Number of oviposition events
Female cyp19a1b-/- mutant fish had significantly more oviposition events compared to WT females (U(17,17)=68, p=0.0074; Figure 2G). On average, cyp19a1b-/- mutant females exhibited 2.7 times more oviposition events than WT females. There were no significant differences in the number of oviposition events among any of the male genotypes (F(3)=0.7744, p=0.5125; Figure 2H).
3.5 Fecundity
There were no significant differences in fertilization rates between female cyp19a1b-/- mutant and WT female pairings (U(14,16)=90, p=0.3557, data not shown), with average fertilization rates of 99% and 93%, respectively. Female cyp19a1b-/- fish spawned significantly more eggs per clutch compared to WT females (U(14,17)=44, p=0.0022, Figure 3A); however, significantly more eleutheroembryos died by Day 4 from the female cyp19a1b-/- clutches compared to the WT clutches (U(14,16)=45, p=0.0043, Figure 3C), resulting in no net difference in fecundity between the female groups (T(28)=0.1303, p=0.8973, Figure 3E). On average, there were 2.3 times more eggs spawned by cyp19a1b-/- females and 4.2 times more progeny died by Day 4 from these mutant females compared to WT females.
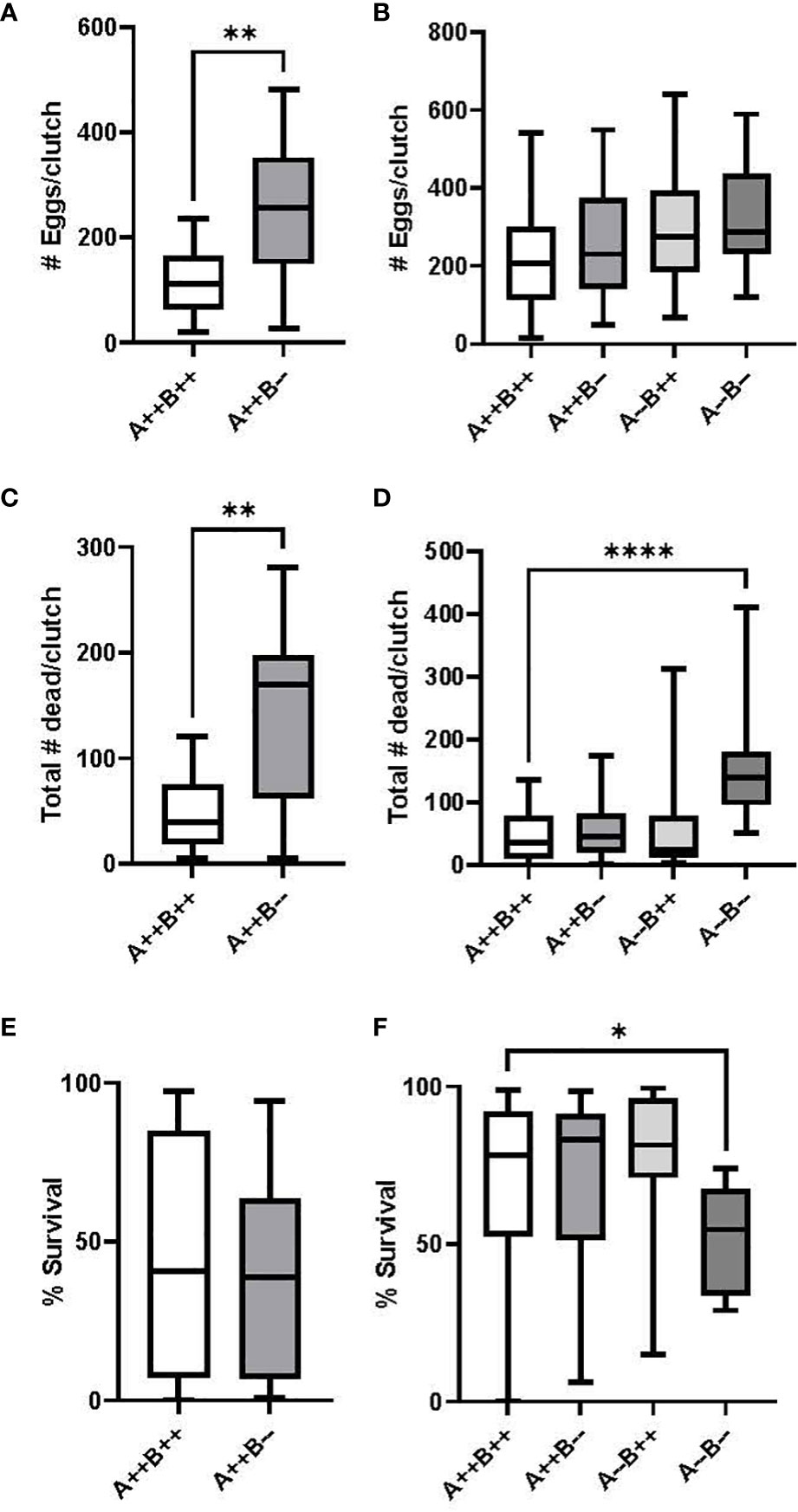
Figure 3 Number of eggs spawned per clutch and egg survival rate during zebrafish pairwise mating trials. Test females (n=14-16 pairs) are displayed on the left (A, C, E) and test males (n=16-23 pairs) are displayed on the right (B, D, F). Mann-Whitney U (A, C), Student’s T (E), One-Way ANOVA (B) and Kruskal Wallis (D, F) tests were performed. Horizontal lines represent mean or median values, boxes represent interquartile ranges, and whiskers represent min-max values. Key to genotypes: A=cyp19a1a, B=cyp19a1b. Asterisks denote p<0.05 (*), p<0.01 (**), p<0.0001 (****).
Fertilization rates for WT, cyp19a1b-/-, cyp19a1a-/-, and cyp19a1a-/-; cyp19a1b-/- group pairings were 99%, 98%, 95%, and 99%, respectively, with no significant differences observed among the male groups (H(3)=3.039, p=0.3856, data not shown). There were no significant differences in the number of eggs per clutch between any of the male group pairings (F(3)=1.649, p=0.1857, Figure 3B); however, there was a significant difference in the number of dead eleutheroembryos per clutch amongst the male genotypes (H(3)=23.91, p<0.0001). Pairwise comparisons revealed that significantly more progeny died by Day 4 from the male cyp19a1a-/-; cyp19a1b-/- group pairings compared to the WT male pairings (p<0.0001, Figure 3D). There was also a significant difference in the survival rate of eleutheroembryos from the male group pairings (H(3)=14.13, p=0.0027). Pairwise comparisons revealed that there was a significantly lower survival rate in the double mutant male offspring compared to the WT male offspring (p=0.0203, Figure 3F). The survival rate of larvae from male cyp19a1a-/-; cyp19a1b-/- pairings was 24% lower than WT pairings.
3.6 Eleutheroembryo morphometrics
There were no significant differences on day 2 between the body length (T(21)=0.1487, p=0.8832; Figure 4A), eye area (T(21)=1.647, p=0.1144; Figure 4C), or yolk sac area (T(21)= 0.09161, p=0.9279, Figure 4E) of offspring from female cyp19a1b-/- and WT females. There were also no significant differences on day 2 between the body length (F(3)=1.657, p=0.1844; Figure 4B), eye area (H(3)=5.172, p=0.1596, Figure 4D), or yolk sac area (H(3)=2.561, p=0.4644, Figure 4F) of offspring from any of the male genotypes.
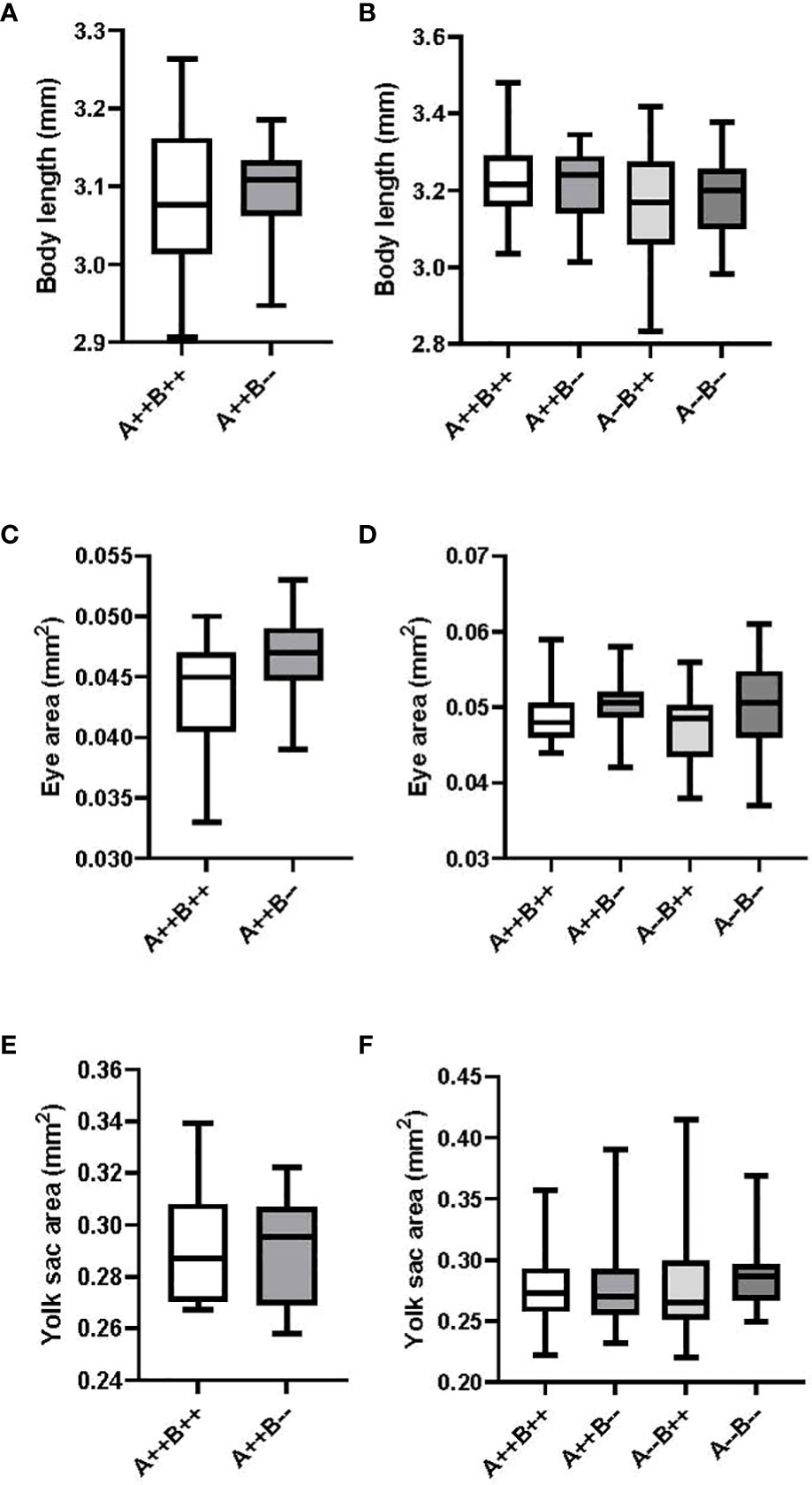
Figure 4 Body length, eye area, and yolk sac area measurements of eleutheroembryos from zebrafish pairwise mating trials on day 2 post-fertilization. Eleutheroembryos from test females (n=9-14) are displayed on the left (A, C, E) and from test males (n=16-21 pairs) are displayed on the right (B, D, F). Student’s T (A, C, E), One-Way ANOVA (B) and Kruskal-Wallis (D, F) tests were performed. Horizontal lines represent mean or median values, boxes represent interquartile ranges, and whiskers represent min-max values. No statistical differences between genotypes were evident. Key to genotypes: A=cyp19a1a, B=cyp19a1b.
3.7 Estradiol content in female brain and ovary
There was a significant main effect of tissue type on E2 levels (F(1,21)=11.40, p=0.0029). Ovarian tissue had 1.6 times higher E2 levels compared to brain. There was no significant main effect of genotype on E2 levels (F(1,21)=3.295, p=0.0838). There was a significant tissue type X genotype interaction (F(1,21)=15.36, p=0.0008). The brains of cyp19a1b-/- mutant females had 2.3 times lower E2 levels compared to the brains of WT females (p=0.0066; Figure 5). The brains of cyp19a1b-/- mutant females also had 3 and 2.2 times lower E2 levels compared to their ovaries (p=0.0002) and to the ovaries of WT females (p=0.0080), respectively (Figure 5).
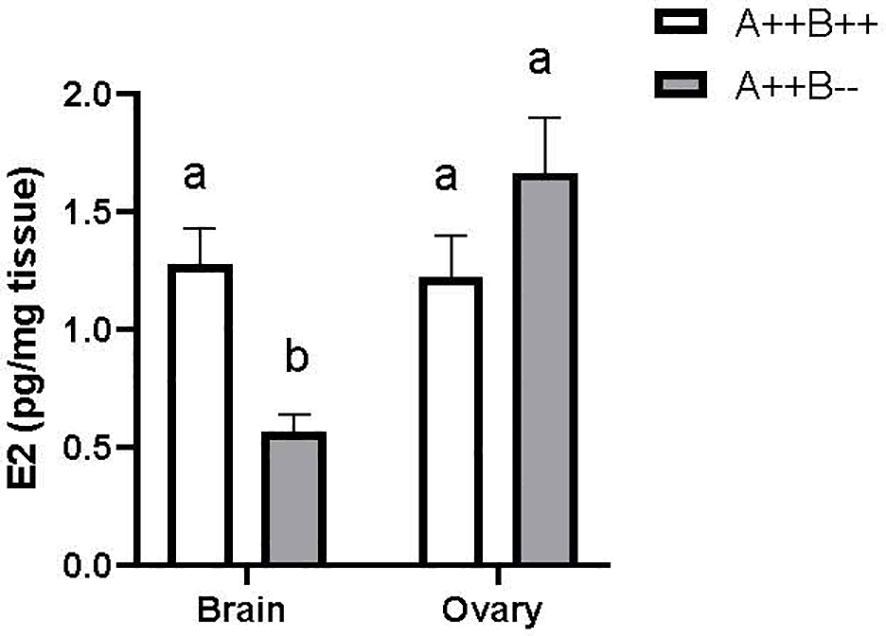
Figure 5 Estradiol (E2) levels in the brains and ovaries of adult WT (A++B++) and cyp19a1b-/- (A++B–) female zebrafish (n=5-8 per group). Data were log-transformed and analyzed using a Two-Way ANOVA followed by Tukey’s multiple comparisons tests. Data are plotted as means + SEM. Means with different letters a-b represent statistically significant differences (p<0.05). For clarity, the significant main effect of tissue type is not displayed. Key to genotypes: A=cyp19a1a, B=cyp19a1b.
4 Discussion
This study is the first assessment of the independent contribution of cyp19a1b and by implication, brain estrogen production, to female spawning behavior in a teleost species. It was found that cyp19a1b-/- mutant female zebrafish exhibited an increased latency to initiate spawning and released higher numbers of eggs compared to WT females. These findings represent an important difference from the observed impairment of female fertility in chemical inhibitor studies in which both aromatase isoforms are inhibited in the body. For example, total aromatase inhibition reduced female fathead minnow (Pimephales promelas; 6), medaka (Oryzias latipes; 7) and zebrafish (8) fecundity. These differences are likely due to the contribution of circulating estrogens produced by the ovaries via Cyp19a1a that function to maintain the integrity of the ovarian state and the reproductive capacity (i.e., fertility) of female zebrafish (10). The observed effects on female spawning behavior in the current study are linked to altered local brain estrogen production via Cyp19a1b, because brain E2 levels are significantly lower in cyp19a1b-/- mutant females compared to WT females, whilst ovarian E2 levels are similar between the groups. The time to the last spawning event, but not the spawning duration, was also longer in the cyp19a1b-/- mutant females. This suggests that the mutation of cyp19a1b affects the perception of reproductive cues important for timely mate identification and assessment, since zebrafish rely heavily on visual and pheromonal cues for reproductive behavior (43). We note that cyp19a1b and estrogen receptors (esr1, esr2a, esr2b) are highly expressed at multiple levels of these sensory pathways including from the peripheral level of sensory nerve fibres to the levels of primary targets and sensory integration centres in the brain (31). Future study of cyp19a1b-/- mutant females will need to account for these potential multiple levels of sensory impairments as well as to identify downstream neuronal mediators of RGC-derived estrogens.
Our results resemble those observed in the tAroKO female mice that display reduced sexual behavior when paired with WT males (25), likely due to impaired brain estrogen signalling (44). There has been no study to date of specific bAroKO effects on female sexual behavior in mammalian models. However, male bAroKO male mice have impaired social recognition and a significantly longer latency to initiate mounting behavior with hormonally primed female mice compared to WT males (28). Our data for male zebrafish contrast those in mice as we observed no significant differences in spawning success between male cyp19a1b-/- mutants and WT fish. Rather, increasing evidence indicates that brain androgen signalling is critical for male teleost sexual behavior. It was recently discovered that cyp17a1-/- male zebrafish display reduced mating behaviors with WT females compared to WT males, which is likely a result of lower brain levels of testosterone and 11-ketotestosterone (45). The reduced contact time of cyp17a1-/- male mutants with WT females could be rescued following 11-ketotestosterone administration, indicating that non-aromatizable androgens regulate male sexual behavior (45). Evidence from androgen receptor gene editing studies also demonstrate the important role for androgen signalling in male teleost sexual behavior (46–48).
An important observation in the current study is that female cyp19a1b-/- mutants paired with WT males spawned significantly more eggs compared to WT females; however, there was a concomitant decrease in larval survival so that the total number of viable larvae produced was similar in female mutants and WT fish. One possible but unlikely explanation for the observed higher larval mortality is reduced circulating E2 and vitellogenin egg deposition in female cyp19a1b-/- mutants. Vitellogenin is a classical estrogen-regulated hepatic protein that nourishes the embryo during early development and is critical for embryo survival (49). Vitellogenin levels correlate with serum E2 and are significantly lower in females following administration of chemical aromatase inhibitors (6–8). The higher larval mortality in the current study is not due to changes in ovary-derived E2 in cyp19a1b-/- mutant females since the cyp19a1b-/- mutant and WT females have similar ovarian tissue (this study) and circulating E2 (13) levels. Moreover, there were no significant differences in yolk sac volume or body length at 2 dpf in progeny from cyp19a1b-/- mutant female and WT pairings. These two morphometrics are positively correlated in embryos and directly linked to maternal vitellogenin levels that are driven by systemic estrogens which bind to activate hepatic estrogen receptors (50, 51).
The cyp19a1b-/- mutant females released more than twice the number of eggs than WT females. This suggests that the mutation of Cyp19a1b may increase the energetic cost of reproduction due to the substantial metabolic investment in large quantities of larvae that do not survive to adulthood. It is well known that spawning is one of the most metabolically demanding activities for a fish (52). For example, the rate of gamete biomass production is roughly proportional to whole-organism metabolic rate, with female fishes allocating approximately half of their energy reserves towards reproductive function (53). It will be important for future studies to determine the contribution of Cyp19a1b to long-term fitness costs associated with higher reproductive investment by mutant females.
Higher mortality rates in larvae from cyp19a1a-/-;cyp19a1b-/- pairings compared to WT pairings among the male genotypes also reveals a role for aromatase in males. While larval survival was similar between WT, cyp19a1a-/- and cyp19a1b-/- males, survival of offspring from the double mutant males was 24% lower than WT offspring. Thus, total aromatase activity of the father contributes to offspring survival. This idea is supported by previous studies reporting increased mortality in larval zebrafish during acute chemical aromatase inhibition (36, 54–57). Moreover, both cyp19a1 transcripts are expressed during the first 48 h post-fertilization in zebrafish embryos, which suggests a role in early development and survival (58). While overall survival rate was lower in larvae from cyp19a1a-/-;cyp19a1b-/- males, there were no obvious larval abnormalities observed and the size of the remaining survivors appeared relatively normal. Yolk sac area, eye area, and body length at 2 dpf were similar across all genotypes. These observations are similar to those of Gould et al. (59), reporting no effect of aromatase inhibition on larval zebrafish development. However, our study contrasts those demonstrating chemical inhibition of both aromatases affect one or multiple of these morphometrics (36, 56, 57, 60, 61). It is possible that significant differences in these morphometrics might emerge during later development after 2 dpf.
5 Conclusions
We have demonstrated a role of brain aromatase in female spawning behavior. Female zebrafish carrying a frameshift mutation in cyp19a1b had a longer latency to initiate spawning behavior and had higher numbers of eggs spawned compared to WT females. The importance of cyp19a1b for embryo survival was demonstrated by the increased mortality of progeny from female cyp19a1b-/- mutants and cyp19a1a-/-;cyp19a1b-/- mutants compared to WT pairings. Further study will be needed to determine the downstream neuronal pathways through which brain estrogens produced in RGCs lead to the observed changes in female spawning behavior. It will also be important to determine the causes of increased mortality in eleutheroembryos from cyp19a1b-/- mutant females and cyp19a1a-/-;cyp19a1b-/- mutant males.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The animal study was reviewed and approved by University of Ottawa Animal Care Committee.
Author contributions
KS, MT, and VLT contributed to conception and design of the study. XL contributed the cyp19a1-/- mutant lines for study. KS and MT conducted data collection and performed statistical analyses. KS wrote the first draft of the manuscript. MT and CL wrote sections of the manuscript. All co-authors contributed to manuscript revision, read, and approved the submitted version.
Funding
This work was supported by an Ontario Graduate Scholarship (KS), University of Ottawa Research Chair in Neuroendocrinology (VLT), and the Natural Sciences and Engineering Research Council of Canada Graduate Scholarship (KS) and Discovery Grant (VLT) Programs.
Acknowledgments
We acknowledge that this research was conducted at the University of Ottawa, which is on unceded Algonquin territory of the Three Fire Confederacy, Anishinaabewaki.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Simpson ER, Mahendroo MS, Means GD, Kilgore MW, Hinshelwood MM, Graham-Lorence S, et al. Aromatase cytochrome P450, the enzyme responsible for estrogen biosynthesis. Endocr Rev (1994) 15(3):342–55. doi: 10.1210/edrv-15-3-342
2. Le Page Y, Menuet A, Kah O, Pakdel F. Characterization of a cis-acting element involved in cell-specific expression of the zebrafish brain aromatase gene. Mol Reprod Dev (2008) 75(10):1549–57. doi: 10.1002/mrd.20892
3. Diotel N, Charlier TD, Lefebvre d'Hellencourt C, Couret D, Trudeau VL, Nicolau JC, et al. Steroid transport, local synthesis, and signaling within the brain: roles in neurogenesis, neuroprotection, and sexual behaviors. Front Neurosci (2018) 12:84. doi: 10.3389/fnins.2018.00084
4. Findlay JK, Liew SH, Simpson ER, Korach KS. Estrogen signaling in the regulation of female reproductive functions. Fertil. Control (2010) 198:29–35. doi: 10.1007/978-3-642-02062-9_2
5. Britt KL, Drummond AE, Dyson M, Wreford NG, Jones ME, Simpson ER, et al. The ovarian phenotype of the aromatase knockout (ArKO) mouse. J Steroid Biochem Mol Biol (2001) 79(1-5):181–5. doi: 10.1016/S0960-0760(01)00158-3
6. Ankley GT, Kahl MD, Jensen KM, Hornung MW, Korte JJ, Makynen EA, et al. Evaluation of the aromatase inhibitor fadrozole in a short-term reproduction assay with the fathead minnow (Pimephales promelas). Toxicol Sci (2002) 67(1):121–30. doi: 10.1093/toxsci/67.1.121
7. Sun L, Zha J, Spear PA, Wang Z. Toxicity of the aromatase inhibitor letrozole to Japanese medaka (Oryzias latipes) eggs, larvae and breeding adults. Comp Biochem Physiol Part C: Toxicol Pharmacol (2007) 145(4):533–41. doi: 10.1016/j.cbpc.2007.01.017
8. Ayobahan SU, Eilebrecht E, Kotthoff M, Baumann L, Eilebrecht S, Teigeler M, et al. A combined FSTRA-shotgun proteomics approach to identify molecular changes in zebrafish upon chemical exposure. Sci Rep (2019) 9(1):6599. doi: 10.1038/s41598-019-43089-7
9. Takatsu K, Miyaoku K, Roy SR, Murono Y, Sago T, Itagaki H, et al. Induction of female-to-male sex change in adult zebrafish by aromatase inhibitor treatment. Sci Rep (2013) 3(1):1–7. doi: 10.1038/srep03400
10. Rahaman MM, Kumagai RI, Tokumoto T. Rapid induction of female-to-male sex change in adult zebrafish by injection of an aromatase inhibitor. Zebrafish (2020) 17(4):261–7. doi: 10.1089/zeb.2020.1864
11. Dranow DB, Hu K, Bird AM, Lawry ST, Adams MT, Sanchez A, et al. Bmp15 is an oocyte-produced signal required for maintenance of the adult female sexual phenotype in zebrafish. PloS Genet (2016) 12(9):e1006323. doi: 10.1371/journal.pgen.1006323
12. Lau ESW, Zhang Z, Qin M, Ge W. Knockout of zebrafish ovarian aromatase gene (cyp19a1a) by TALEN and CRISPR/Cas9 leads to all-male offspring due to failed ovarian differentiation. Sci Rep (2016) 6(1):1–14. doi: 10.1038/srep37357
13. Yin Y, Tang H, Liu Y, Chen Y, Li G, Liu X, et al. Targeted disruption of aromatase reveals dual functions of cyp19a1a during sex differentiation in zebrafish. Endocrinology (2017) 158(9):3030–41. doi: 10.1210/en.2016-1865
14. Fisher CR, Graves KH, Parlow AF, Simpson ER. Characterization of mice deficient in aromatase (ArKO) because of targeted disruption of the cyp19 gene. Proc Natl Acad Sci USA (1998) 95(12):6965–70. doi: 10.1073/pnas.95.12.6965
15. Robertson KM, O’Donnell L, Jones ME, Meachem SJ, Boon WC, Fisher CR, et al. Impairment of spermatogenesis in mice lacking a functional aromatase (cyp 19) gene. Proc Natl Acad Sci USA (1999) 96(14):7986–91. doi: 10.1073/pnas.96.14.7986
16. O’Donnell L, Robertson KM, Jones ME, Simpson ER. Estrogen and spermatogenesis. Endocr. Rev (2001) 22(3):289–318. doi: 10.1210/edrv.22.3.0431
17. Robertson KM, Simpson ER, Lacham-Kaplan ORLY, Jones ME. Characterization of the fertility of male aromatase knockout mice. J Androl (2001) 22(5):825–30. doi: 10.1002/j.1939-4640.2001.tb02587.x
18. Tang H, Chen Y, Liu Y, Yin Y, Li G, Guo Y, et al. New insights into the role of estrogens in male fertility based on findings in aromatase-deficient zebrafish. Endocrinology (2017) 158(9):3042–54. doi: 10.1210/en.2017-00156
19. Honda SI, Harada N, Takagi Y. Novel exon 1 of the aromatase gene specific for aromatase transcripts in human brain. Biochem Biophys Res Commun (1994) 198(3):1153–60. doi: 10.1006/bbrc.1994.1163
20. Honda SI, Harada N, Abe-Dohmae S, Takagi Y. Identification of cis-acting elements in the proximal promoter region for brain-specific exon 1 of the mouse aromatase gene. Mol Brain Res (1999) 66(1-2):122–32. doi: 10.1016/S0169-328X(99)00017-0
21. Ramachandran B, Schlinger BA, Arnold AP, Campagnoni AT. Zebra finch aromatase gene expression is regulated in the brain through an alternate promoter. Gene (1999) 240(1):209–16. doi: 10.1016/S0378-1119(99)00399-6
22. Yilmaz MB, Wolfe A, Cheng YH, Glidewell-Kenney C, Jameson JL, Bulun SE. Aromatase promoter if is regulated by estrogen receptor alpha (ESR1) in mouse hypothalamic neuronal cell lines. Biol Reprod (2009) 81(5):956–65. doi: 10.1095/biolreprod.109.077206
23. Honda SI, Harada N, Ito S, Takagi Y, Maeda S. Disruption of sexual behavior in male aromatase-deficient mice lacking exons 1 and 2 of the cyp19 gene. Biochem Biophys Res Commun (1998) 252(2):445–9. doi: 10.1006/bbrc.1998.9672
24. Bakker J, Honda S, Harada N, Balthazart J. Sexual partner preference requires a functional aromatase (cyp19) gene in male mice. Hormones Behav (2002) 42(2):158–71. doi: 10.1006/hbeh.2002.1805
25. Bakker J, Honda SI, Harada N, Balthazart J. The aromatase knock-out mouse provides new evidence that estradiol is required during development in the female for the expression of sociosexual behaviors in adulthood. J Neurosci (2002) 22(20):pp.9104–9112. doi: 10.1523/JNEUROSCI.22-20-09104.2002
26. Matsumoto T, Honda SI, Harada N. Alteration in sex-specific behaviors in male mice lacking the aromatase gene. Neuroendocrinology (2003) 77(6):416–24. doi: 10.1159/000071313
27. Pierman S, Sica M, Allieri F, Viglietti-Panzica C, Panzica GC, Bakker J. Activational effects of estradiol and dihydrotestosterone on social recognition and the arginine-vasopressin immunoreactive system in male mice lacking a functional aromatase gene. Hormones Behav (2008) 54(1):98–106. doi: 10.1016/j.yhbeh.2008.02.001
28. Brooks DC, Coon V JS, Ercan CM, Xu X, Dong H, Levine JE, et al. Brain aromatase and the regulation of sexual activity in male mice. Endocrinology (2020) 161(10):bqaa137. doi: 10.1210/endocr/bqaa137
29. Huffman LS, O’Connell LA, Hofmann HA. Aromatase regulates aggression in the African cichlid fish astatotilapia burtoni. Physiol Behav (2013) 112:77–83. doi: 10.1016/j.physbeh.2013.02.004
30. Jalabert C, Quintana L, Pessina P, Silva A. Extra-gonadal steroids modulate non-breeding territorial aggression in weakly electric fish. Hormones Behav (2015) 72:60–7. doi: 10.1016/j.yhbeh.2015.05.003
31. Shaw K. Aromatase expression and function in the brain and behavior: a comparison across communication systems in teleosts. J Chem Neuroanat (2018) 94:139–53. doi: 10.1016/j.jchemneu.2018.10.004
32. Menuet A, Pellegrini E, Brion F, Gueguen MM, Anglade I, Pakdel F, et al. Expression and estrogen-dependent regulation of the zebrafish brain aromatase gene. J Comp Neurol (2005) 485(4):304–20. doi: 10.1002/cne.20497
33. Darrow KO, Harris WA. Characterization and development of courtship in zebrafish, Danio rerio. Zebrafish (2004) 1(1):40–5. doi: 10.1089/154585404774101662
34. Oliveira RF, Silva JF, Simoes JM. Fighting zebrafish: characterization of aggressive behavior and winner–loser effects. Zebrafish (2011) 8(2):73–81. doi: 10.1089/zeb.2011.0690
35. Trainor BC, Kyomen HH, Marler CA. Estrogenic encounters: how interactions between aromatase and the environment modulate aggression. Front Neuroendocrinol (2006) 27(2):170–9. doi: 10.1016/j.yfrne.2005.11.001
36. Martínez R, Navarro-Martín L, Luccarelli C, Codina AE, Raldúa D, Barata C, et al. Unravelling the mechanisms of PFOS toxicity by combining morphological and transcriptomic analyses in zebrafish embryos. Sci Total Environ (2019) 674:462–71. doi: 10.1016/j.scitotenv.2019.04.200
37. Chen Y, Tang H, Wang L, He J, Guo Y, Liu Y, et al. Fertility enhancement but premature ovarian failure in esr1-deficient female zebrafish. Front Endocrinol (2018) 9:567. doi: 10.3389/fendo.2018.00567
38. Converse A, Thomas P. Androgens regulate follicle stage-dependent pro-and anti-apoptosis in teleost ovaries through ZIP9 activation of different G proteins. Biol Reprod (2019) 101(2):377–91. doi: 10.1093/biolre/ioz086
39. Giroux M, Vliet SM, Volz DC, Gan J, Schlenk D. Mechanisms behind interactive effects of temperature and bifenthrin on the predator avoidance behaviors in parr of Chinook salmon (Oncorhynchus tshawytscha). Aquat Toxicol (2019) 216:105312. doi: 10.1016/j.aquatox.2019.105312
40. Saravanan M, Nam SE, Eom HJ, Lee DH, Rhee JS. Long-term exposure to waterborne nonylphenol alters reproductive physiological parameters in economically important marine fish. Comp Biochem Physiol Part C: Toxicol Pharmacol (2019) 216:10–8. doi: 10.1016/j.cbpc.2018.11.009
41. Kayo D, Oka Y, Kanda S. Examination of methods for manipulating serum 17β-estradiol (E2) levels by analysis of blood E2 concentration in medaka (Oryzias latipes). Gen Comp Endocrinol (2020) 285:113272. doi: 10.1016/j.ygcen.2019.113272
42. Nishiike Y, Miyazoe D, Togawa R, Yokoyama K, Nakasone K, Miyata M, et al. Estrogen receptor 2b is the major determinant of sex-typical mating behavior and sexual preference in medaka. Curr Biol (2021) 31(8):1699–710. doi: 10.1016/j.cub.2021.01.089
43. Li J, Hubbard PC, Canário AV. Male Zebrafish (Danio rerio) odorants attract females and induce spawning. Aquacult Fish (2018) 3(4):139–44. doi: 10.1016/j.aaf.2018.06.004
44. Cornil CA. On the role of brain aromatase in females: why are estrogens produced locally when they are available systemically? J Comp Physiol A (2018) 204(1):31–49. doi: 10.1007/s00359-017-1224-2
45. Shu T, Zhai G, Pradhan A, Olsson PE, Yin Z. Zebrafish cyp17a1 knockout reveals that androgen-mediated signaling is important for male brain sex differentiation. Gen Comp Endocrinol (2020) 295:113490. doi: 10.1016/j.ygcen.2020.113490
46. Yong L, Thet Z, Zhu Y. Genetic editing of the androgen receptor contributes to impaired male courtship behavior in zebrafish. J Exp Biol (2017) 220(17):3017–21. doi: 10.1242/jeb.161596
47. Alward BA, Laud VA, Skalnik CJ, York RA, Juntti SA, Fernald RD. Modular genetic control of social status in a cichlid fish. Proc Natl Acad Sci USA (2020) 117(45):28167–74. doi: 10.1073/pnas.2008925117
48. Ogino Y, Ansai S, Watanabe E, Yasugi M, Katayama Y, Sakamoto H, et al. Evolutionary differentiation of androgen receptor is responsible for sexual characteristic development in a teleost fish. Nat Commun (2023) 14(1):1428. doi: 10.1038/s41467-023-37026-6
49. Hara A, Hiramatsu N, Fujita T. Vitellogenesis and choriogenesis in fishes. Fish. Sci (2016) 82:187–202. doi: 10.1007/s12562-015-0957-5
50. Jardine D, Litvak MK. Direct yolk sac volume manipulation of zebrafish embryos and the relationship between offspring size and yolk sac volume. J Fish Biol (2003) 63(2):388–97. doi: 10.1046/j.1095-8649.2003.00161.x
51. Bugel SM, White LA, Cooper KR. Decreased vitellogenin inducibility and 17β-estradiol levels correlated with reduced egg production in killifish (Fundulus heteroclitus) from Newark bay, NJ. Aquat Toxicol (2011) 105(1-2):1–12. doi: 10.1016/j.aquatox.2011.03.013
52. McBride RS, Somarakis S, Fitzhugh GR, Albert A, Yaragina NA, Wuenschel MJ, et al. Energy acquisition and allocation to egg production in relation to fish reproductive strategies. Fish Fish. (2015) 16(1):23–57. doi: 10.1111/faf.12043
53. Hayward A, Gillooly JF. The cost of sex: quantifying energetic investment in gamete production by males and females. PloS One (2011) 6(1):e16557. doi: 10.1371/journal.pone.0016557
54. Allgood OE Jr., Hamad A, Fox J, DeFrank A, Gilley R, Dawson F, et al. Estrogen prevents cardiac and vascular failure in the ‘listless’ zebrafish (Danio rerio) developmental model. Gen Comp Endocrinol (2013) 189:33–42. doi: 10.1016/j.ygcen.2013.04.016
55. Santos D, Matos M, Coimbra AM. Developmental toxicity of endocrine disruptors in early life stages of zebrafish, a genetic and embryogenesis study. Neurotoxicol. Teratol. (2014) 46:18–25. doi: 10.1016/j.ntt.2014.08.002
56. Alharthy KM, Albaqami FF, Thornton C, Corrales J, Willett KL. Mechanistic evaluation of benzo [a] pyrene’s developmental toxicities mediated by reduced Cyp19a1b activity. Toxicol. Sci (2017) 155(1):135–47. doi: 10.1093/toxsci/kfw182
57. Bernardo RC, Connaughton VP. Transient developmental exposure to tributyltin reduces optomotor responses in larval zebrafish (Danio rerio). Neurotoxicol. Teratol. (2022) 89:107055. doi: 10.1016/j.ntt.2021.107055
58. Sawyer SJ, Gerstner KA, Callard GV. Real-time PCR analysis of cytochrome P450 aromatase expression in zebrafish: gene specific tissue distribution, sex differences, developmental programming, and estrogen regulation. Gen Comp Endocrinol (2006) 147(2):108–17. doi: 10.1016/j.ygcen.2005.12.010
59. Gould CJ, Saldanha CJ, Connaughton VP. Acute exposure to 4-OH-A, not PCB1254, alters brain aromatase activity but does not adversely affect growth in zebrafish. Environ Toxicol Pharmacol (2019) 68:133–40. doi: 10.1016/j.etap.2019.02.010
60. Hamad A, Kluk M, Fox J, Park M, Turner JE. The effects of aromatase inhibitors and selective estrogen receptor modulators on eye development in the zebrafish (Danio rerio). Curr Eye Res (2007) 32(10):819–27. doi: 10.1080/02713680701573712
Keywords: aromatase, brain, cyp19a1b, estrogen, hormone, neuroendocrine, sexual behavior, zebrafish
Citation: Shaw K, Therrien M, Lu C, Liu X and Trudeau VL (2023) Mutation of brain aromatase disrupts spawning behavior and reproductive health in female zebrafish. Front. Endocrinol. 14:1225199. doi: 10.3389/fendo.2023.1225199
Received: 18 May 2023; Accepted: 13 June 2023;
Published: 23 June 2023.
Edited by:
Paula Gabriela Vissio, University of Buenos Aires, ArgentinaReviewed by:
Toshinobu Tokumoto, Shizuoka University, JapanTaisen Iguchi, Graduate University for Advanced Studies (Sokendai), Japan
Copyright © 2023 Shaw, Therrien, Lu, Liu and Trudeau. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Katherine Shaw, kshaw032@uottawa.ca; Vance L. Trudeau, trudeauv@uottawa.ca