Mesenchymal stromal cells, metabolism, and mitochondrial transfer in bone marrow normal and malignant hematopoiesis
 Abhishek K. Singh
Abhishek K. Singh Parash Prasad
Parash Prasad Jose A. Cancelas
Jose A. Cancelas- 1Division of Experimental Hematology and Cancer Biology, Cincinnati Children’s Hospital Medical Center, Cincinnati, OH, United States
- 2Hoxworth Blood Center, University of Cincinnati College of Medicine, Cincinnati, OH, United States
Hematopoietic stem cell (HSC) transplantation-based treatments are in different phases of clinical development, ranging from current therapies to a promise in the repair and regeneration of diseased tissues and organs. Mesenchymal stromal/stem cells (MSCs), which are fibroblast-like heterogeneous progenitors with multilineage differentiation (osteogenic, chondrogenic, and adipogenic) and self-renewal potential, and exist in the bone marrow (BM), adipose, and synovium, among other tissues, represent one of the most widely used sources of stem cells in regenerative medicine. MSCs derived from bone marrow (BM-MSCs) exhibit a variety of traits, including the potential to drive HSC fate and anti-inflammatory and immunosuppressive capabilities via paracrine activities and interactions with the innate and adaptive immune systems. The role of BM-MSC-derived adipocytes is more controversial and may act as positive or negative regulators of benign or malignant hematopoiesis based on their anatomical location and functional crosstalk with surrounding cells in the BM microenvironment. This review highlights the most recent clinical and pre-clinical findings on how BM-MSCs interact with the surrounding HSCs, progenitors, and immune cells, and address some recent insights on the mechanisms that mediate MSCs and adipocyte metabolic control through a metabolic crosstalk between BM microenvironment cells and intercellular mitochondrial transfer in normal and malignant hematopoiesis.
1 Introduction
Stem cell transplantation-based treatments have demonstrated considerable promise in the repair and regeneration of damaged tissues and organs in a variety of degenerative disorders. Mesenchymal stromal/stem cells (MSCs), defined as non-hematopoietic, plastic-adherent, fibroblastic colony-forming cells, are among the most frequently used cell types in translational medicine. These cells are multipotent heterogeneous progenitors, able to self-renew, and differentiate into multiple cell lineages, such as chondrocytes, osteoblasts, and adipocytes. MSCs reside in the bone marrow (BM) and in a variety of other tissues and organs, including the adipose tissue, placenta, skin, fallopian tubes, cord blood, liver, and lungs (Pittenger et al., 1999). MSCs can engraft after intrafemoral administration (Abbuehl et al., 2017) but seem to be short-lived after intravenous administration due to apoptosis (Preda et al., 2021), which hampers their ability to engraft robustly in the adult BM (Poloni et al., 2006; Eggenhofer et al., 2012). Despite these limitations, numerous studies have showcased the broad anti-inflammatory and immunosuppressive properties of infused MSCs even if undergoing apoptosis (Abkowitz et al., 1992; Luk et al., 2016; Galleu et al., 2017).
MSCs, through their interaction with the innate and adaptive immune systems and tissue regeneration capacity, emerged in the last 2 decades as a promising tool in regenerative medicine and some contexts of immunosuppression (Keating, 2012; Bernardo and Fibbe, 2013). Ex vivo expanded human MSCs have been demonstrated to be efficacious in pre-clinical models of hematopoietic stem cell (HSC) transplantation (HSCT) and have also been employed in phase I/II clinical trials to facilitate the hematopoietic engraftment and alleviate acute graft-versus-host disease (GvHD) (Noort et al., 2002; Le Blanc et al., 2004; Bernardo and Fibbe, 2015; Burnham et al., 2020; Ringden et al., 2022). In a recent phase III clinical trial, remestemcel-L, an ex vivo culture-expanded allogeneic adult human MSC, has been developed as a first-line therapy in pediatric patients with steroid-refractory acute GvHD (Kurtzberg et al., 2020). Ex vivo expansion of cord blood-derived HSC and progenitor cells (HSPCs) cultured on BM mesenchymal lineage stroma maintains the long-term primitive HSC (CD34+CD38−) immunophenotype and improves the engraftment kinetics (Robinson et al., 2006; de Lima et al., 2012; Mehta et al., 2017).
In BM, MSCs are the critical components of the HSC niche, regulating the composition of the niche environment and the hematopoietic process by secreting paracrine factors. Over the last decade, there has been a surge of interest in BM-MSC metabolism, which plays a crucial role in the regulation of surrounding niche architecture and HSCT outcomes in both normal physiology and pathological conditions. A growing body of research describes a link between MSC metabolic reprogramming via mitochondria transfer and tissue repair/regeneration (Islam et al., 2012; Mistry et al., 2019; Golan et al., 2020) (Figure 1). This mitochondrion-mediated MSC metabolic reprogramming, on the other hand, becomes poised in malignancy, modifies leukemia cell metabolism by mitochondrial transfer, and provides a survival advantage, following chemotherapy (Moschoi et al., 2016; Marlein et al., 2017; Burt et al., 2019; Forte et al., 2020; Saito et al., 2021). This review highlights new concepts on the mechanisms through which BM-MSCs interact with the surrounding HSPCs and address the mechanism that mediates MSC metabolic reprogramming to provide a better understanding of MSC-based therapies in normal and malignant hematopoiesis.
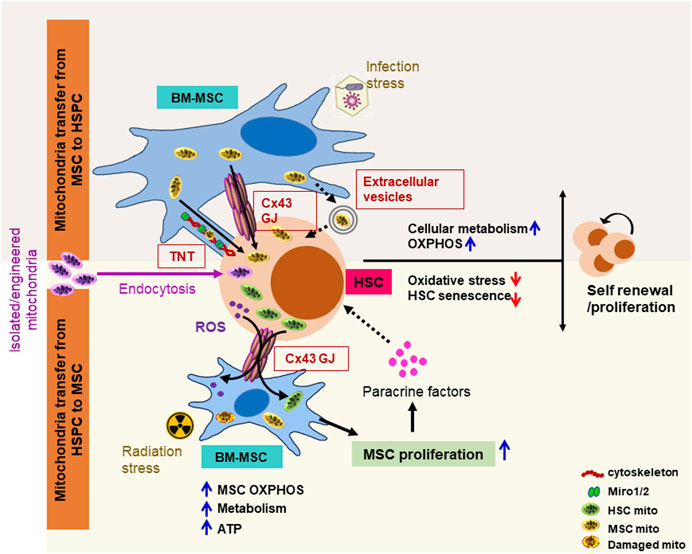
FIGURE 1. Schematic representation of mitochondria transfer mechanisms between BM-MSCs and HSCs in response to blood formation emergencies. Intercellular mitochondrial exchange between BM-MSCs and HSPCs occurs through (1) tunneling nano-tubules (TNTs) via stabilization of cytoskeleton element (F-actin, myosin, and tubulin) and membrane adapter proteins (miro1/2); (2) gap junction (GJ) channels such as connexin 43 (Cx43), where two juxtapositioned connexin hemichannels form pores connecting two neighboring cells, facilitating bidirectional mitochondrial exchange; and (3) extracellular vesicles ranging from 0.1 to 1 μm formed by blebbing of the plasma membrane. Different stress signals differentially regulate the intercellular mitochondrial exchange between MSCs and HSCs to improve the hematopoietic outcome. Following myeloablative irradiation and HSPC transplantation, transplanted HSPCs transfer functional mitochondria to BM-MSC via the gap junction protein connexin 43 in a cell contact-dependent manner, enhances MSC metabolic recovery, ATP levels, and proliferation, which, in turn, by regulating the BM niche factors (CXCL12, SCF, IL-7, osteopontin, and angiopoietin-1) improves HSPC proliferation/survival. By contrast, in infection stress hematopoiesis, BM-MSCs facilitate the transfer of mitochondria to HSPCs via TNTs and/or Cx43 GJ, improve HSPC metabolism by switching from glycolysis to oxidative phosphorylation, and subsequently stimulate hematopoietic recovery. Exogeneous-isolated functional mitochondria can be efficiently transferred into HSPCs through macropicnocytosis/endocytosis. These transferred mitochondria metabolically upgrade HSPC and boost long-term engraftment potential. ATP, adenosine triphosphate; Miro1/2, mitochondrial RhoGTPase 1/2; mito, mitochondria.
2 Identity and characterization of BM-MSCs
BM-MSCs are often perivascular, wrapped tightly around arterioles and more loosely around the sinusoidal blood vessels (Crisan et al., 2008; Crane et al., 2017; Wei and Frenette, 2018; Pinho and Frenette, 2019). The varied nature of the MSC population and the requirement to identify functionally distinct subgroups present a challenge to understanding the function of MSCs in the HSC niche. In human tissue, MSCs are recognized based on the presence of CD44, CD90, CD105, CD73, and CD106 and the lack of hematopoietic and endothelial (CD31) cell markers (Dominici et al., 2006; Schwab and Gargett, 2007; Crisan et al., 2008; Ouzin and Kogler, 2023). However, the MSCs isolated from different sources exhibit differential gene expression patterns, and over the years, a significant number of markers have been proposed to aid in the isolation of MSCs from their surroundings. BM-MSCs can be enriched using a combination of stemness markers such as Stro-1, SSEA-4, CD271, and CD146 (Buhring et al., 2007; Lv et al., 2014). MSCs expressing CD146 persist around the sinusoidal blood vessels in the ossicles, highly express the HSC niche factor angiopoietin-1, and regenerate bone and stroma to establish a hematopoietic milieu and favor HSPC proliferation (Sacchetti et al., 2007; Schwab and Gargett, 2007; Tormin et al., 2011). In contrast, CD271+ MSC localized in the trabecular region of human BM associated with the maintenance of long-term HSPCs in low-oxygen areas. These cells lack the ability to express CD90 and CD73, and when combined with minimal or no expression of PDGFRα, define the MSC subgroup promoting ex vivo expansion of transplantable CD34+ HSC (Tormin et al., 2011; Li et al., 2014; Crippa et al., 2021). Both CD271+CD146−/low and CD271+CD146+ cells were observed to generate colony-forming unit–fibroblasts (CFU-Fs) in vitro. However, MSCs could only self-renew in serial transplantation in vivo when grown as tridimensional mesenspheres and not as plastic-adherent CFU-Fs, demonstrating that plastic-adherent CFU-Fs contain very few primitive MSCs (Ghazanfari et al., 2016). In a recent study, Zhang et al. (2023), by performing a comprehensive screening of human fetal BM-nucleated cells at single-cell resolution, discovered LIFR+PDGFRB+CD45−CD31−CD235a− (LIFR+PDGFRB+) and TM4SF1+CD44+CD73+CD45−CD31−CD235a− (TM4SF1+CD44+CD73+) mesenchymal progenitors. By performing in vivo transplantation, they showed that only the LIFR+PDGFRB+ MSC subset can effectively restore the hematopoietic microenvironment in vivo, whereas TM4SF1+CD44+CD73+ cells are preferentially committed to osteogenic differentiation (Zhang et al., 2023). RNA velocity and cell–cell communication analysis further identified six transcriptionally and functionally distinct stromal cell populations, demonstrating that CD45low/−CD235a−CD71−CD271+NCAM1−CD52−CD81++ MSCs have the highest CFU-F potential and tri-lineage differentiation capacity and interact with essentially all hematopoietic cell types through CXCL12 pathways (Li et al., 2023).
Mouse BM-MSCs are identified by the presence of PDGFRα, CD29, CD49e, CD44, CD73, CD105, and Sca-1, with the absence of hematopoietic and endothelial cell markers (Morikawa et al., 2009; Breitbach et al., 2018). Self-renewing mouse MSCs are highly concentrated in the PDGFRα+Sca-1+CD45−Ter119− (PαS) population, preferentially reside in the arterial perivascular space near the inner surface of the cortical bone, and differentiate into osteogenic, chondrogenic, and adipogenic populations. These cells selectively express HSC niche factor angiopoietin-1. On the other hand, the CXCL12-abundant reticular (CAR) cells, phenotypically identified as PDGFRα+Sca-1−CD45-Ter119− MSCs, primarily found surrounding sinusoids induce adipo-osteogenic bipotential progenitors and express high levels of CXCL12 to maintain hematopoiesis (Morikawa et al., 2009; Omatsu et al., 2010). Lineage tracing and fate mapping studies further identified nestin and leptin receptor (leptinR+), expressing perivascular MSCs in BM (Mendez-Ferrer et al., 2010; Ding et al., 2012; Greenbaum et al., 2013; Kunisaki et al., 2013; Zhou et al., 2014; Wei and Frenette, 2018; Pinho and Frenette, 2019; Severe et al., 2019; Kara et al., 2023). Nestin+ cells have been shown to originate from the neural crest and together with an increased capacity to differentiate into mesodermal cells, release of colony-stimulating factor-1 (CSF-1; OR macrophage-stimulating-factor; M-CSF), and the tissue inhibitors of metalloproteinase (TIMP)-1 and -2 support the HSC function (Mendez-Ferrer et al., 2010; Kunisaki et al., 2013). LeptinR+ cells are heterogenous, with peri-arteriolar leptinR+ cells poised to undergo osteogenic differentiation, while peri-sinusoidal leptinR+ cells poised to undergo adipogenic differentiation and support hematopoiesis in vivo by secreting several niche factors, such as CXCL12, SCF, and angiopoietin-1 (Zhou et al., 2014; Shen et al., 2021). LeptinR+CD45-Ter119− represent 0.2%–0.3% of enzymatically dissociated BM cells and are highly overlapping with PDGFRα+CD45−Ter119−CD31− MSCs. Additionally, MSC markers CD51 and PDGFRβ are consistently expressed by these LepR+CD45−Ter119− cells (Zhou et al., 2014; Shen et al., 2021). Contrarily, nestin-GFP+CD45−Ter119−CD31− MSCs represent a small subset (0.08%) of BM-nucleated cells and uniformly express PDGFRα and CD51. Other mesenchymal lineage markers were either heterogeneously expressed (CD29, CD44, CD61, CD130, and P75) or restricted to a small subset of nestin+ cells (CD10, CD90, CD166, and CD133) (Mendez-Ferrer et al., 2010; Pinho et al., 2013). Nestin transgenes display different expression patterns in BM, and by using reporter and conditional gene-targeted mice, Kunisaki et al., identified a rare (0.002% of total BM) sympathetic nervous system-innervated nestin-GFPbright subset that is positive for the pericyte marker NG2 (chondroitin sulfate proteoglycan-4, Cspg4) and α-smooth muscle actin (α-SMA) and regulates HSC quiescence (Kunisaki et al., 2013). In a recent study, Kara et al. (2023), employing scRNA seq analysis, showed that leptinR+ MSCs are the primary sources of SCF and CXCL12 in early postnatal and adult BM. Intriguingly, within nestin+ MSCs, SCF and CXCL12 clusters only coincide with nestin+SMA+ cells (Kara et al., 2023).
Single-cell protein expression mapping has demonstrated that leptinR+ and nestin+ BM-MSCs are highly susceptible to myeloablative conditioning, and the stromal cell subset defined as CD105−CD73+NGFRhigh participates in HSPC engraftment and initial stages of hematological recovery post-irradiation (Severe et al., 2019). Of note, the radio-tolerant CD105−CD73+NGFRhigh stromal cells express a high level of CXCL12 with minimal or no overlap with leptinR+ and nestin+ MSCs (Severe et al., 2019). Further studies employing in vivo ectopic bone-forming assay and differential expression of cell surface markers revealed MSC hierarchy in BM, where CD45−Ter119−CD31−CD166−CD146−Sca1+ (Sca1+) cells are most primitive, giving rise to CD45−Ter119−CD31−CD166−CD146+ (CD146+) intermediate progenitors and mature CD45−Ter119−CD31−CD166+CD146−(CD166+) osteo-progenitors (Hu et al., 2016). All these mesenchymal progenitors preserve in vitro HSC long-term multi-lineage reconstitution potential. However, their differentiation potential differs dramatically in vivo, and only most primitive Sca1+ mesenchymal stromal progenitors, which coincide with leptin receptor-expressing MSCs, differentiate into CXCL12-producing stromal cells to support hematopoiesis (Hu et al., 2016). Considering the fact that MSCs are highly heterogenous and differentially regulate HSC composition, the ex vivo expansion of MSCs on the plastic surface induces phenotypic and functional changes, and myeloablative conditioning differentially impairs BM-resident MSC fate and function, further studies are required to identify the clinically relevant MSC subsets to improve the hematopoietic outcome.
3 Role of BM-MSC and its adipocytic lineage in hematopoiesis
Inadequate engraftment of HSCs is a major cause of morbidity after clinical HSCT. Notwithstanding, myelosuppression, which lasts for several weeks in post-transplant patients, deteriorates the adaptive immunity and raises the risk of opportunistic infections (Storek et al., 2008). Although multiple studies have focused on understanding the HSC biology that sustains native hematopoiesis, therapeutic strategies promoting BM niche reconstitution after myelosuppressive injury are essential to improve clinical HSCT outcomes. Multiple BM niche cells differentially support and preserve the integrity of hematopoiesis. MSCs constitute an essential component of the BM niche and is one of the main regulators of HSPC homeostasis by secreting paracrine factors like CXCL12, SCF, IL-7, osteopontin, angiopoietin-1, and VCAM-1, as well as by establishing a cell-to-cell contact (Hooper et al., 2009; Butler et al., 2010; Schajnovitz et al., 2011; Ding et al., 2012; Mendelson and Frenette, 2014; Ziegler et al., 2016; Guo et al., 2017; Himburg et al., 2018; Chen et al., 2019; Batsivari et al., 2020; Singh and Cancelas, 2020; Yin et al., 2020). The potent immunomodulatory properties of MSCs offer therapeutic benefits in the management and prevention of GvHD and in the promotion of tissue regeneration and engraftment, following HSCT.
BM adipocytes (BMATs) play opposing roles in the process of hematopoiesis. Different cytokines, including CXCL12, IL-8, CSF3, and LIF, which positively control HSC survival and provide essential niche factors, are generated from BM adipocytes (Mattiucci et al., 2018; Wilson et al., 2018). These cells also produce several hematopoietic supportive factors and regulate HSC homing. In contrast, others observed that BM adipocytes function as a negative regulator of hematopoiesis (Naveiras et al., 2009; Zhu et al., 2013; Lu et al., 2016), and depending on the anatomic location (proximal-regulated-rBMAT vs. distal-constitutive-cBMAT) within the BM, they act differently (Tratwal et al., 2021). In comparison to cBMATs, rBMATs may be the primary regulator of hematopoiesis; however, further research with appropriate origin information on these different types of BMATs is needed to justify their involvement in hematopoiesis.
3.1 Mesenchymal stem cells facilitate hematopoietic engraftment
HSC mainly resides in the specialized niche of the BM, and myeloablative preconditioning for HSCT by total body irradiation or chemotherapy not only depletes the recipient’s HSCs but also permanently incapacitates the sinusoidal blood vessels and BM stromal cells (Abbuehl et al., 2017; Tikhonova et al., 2019). In the past decade, several preclinical and clinical investigations have shown that co-transplantation of MSCs with HSCs facilitates the migration and homing of the HSCs to recipient BM niches and improves hematopoietic recovery (Noort et al., 2002; Maitra et al., 2004; Muguruma et al., 2006; Le Blanc et al., 2007; Abbuehl et al., 2017; Tikhonova et al., 2019; Zhang et al., 2023). Transplantation of adult BM cells from EGFP transgenic animals can generate a population of Lin−/Sca-1+/c-kit− BM cells, albeit at low levels, in the BM of osteogenesis imperfecta (OI) recipients when transplanted intra-utero (Panaroni et al., 2009). Unfortunately, this study did not report engraftment of purified, fresh, or expanded MSCs into the same OI mice. Transplantation of a freshly isolated primary BM-MSC demonstrated that only a rare CD73+/CD105−/Sca1+ population within the BM-MSCs engrafts long-term, possesses self-renewal potential, and regenerates multilineage BM niche cells to support hematopoiesis, when co-transplanted intrafemorally with HSC (Abbuehl et al., 2017). Furthermore, following 3D whole-mount imaging by light-sheet microscopy, they observed that the engrafted MSC primarily localizes to the metaphysis and cortical region of bone close to HSCs (Abbuehl et al., 2017).
The current view is that infused BM-MSCs are, in general, short-lived (Poloni et al., 2006; Eggenhofer et al., 2012; Preda et al., 2021). However, a non-apoptotic subpopulation of BM-MSCs may survive and remain dormant during homeostasis. In response to stress/injury, these MSCs may proliferate and differentiate to support emergency hematopoiesis. Epidermal growth factor (EGF), fibroblast growth factor-2 (FGF-2), and PDGFβ are amongst the BM environment-derived cytokines shown to promote long-term ex vivo expansion and differentiation of MSCs (Ng et al., 2008; Tamama et al., 2010; Itkin et al., 2012; Doan et al., 2013; Chen et al., 2015; Han et al., 2022). Of note, MSCs engineered to overexpress PDGFβ exhibit improved survival and growth after transplantation and dramatically promote the engraftment of the human HSC in immunodeficient mice (Yin et al., 2020). Due to the limited availability of human tissues, MSCs must be expanded ex vivo to meet the cellular demand and transplantable HSPC expansion. A large cohort of studies have suggested that following the ex vivo culture, BM-MSCs gradually lose their proliferative and secretory capabilities and alter the expression of key regulators of HSC self-renewal and maintenance (Abbuehl et al., 2017; Nakahara et al., 2019; Zhang et al., 2023). The expression of Spp1 (osteopontin), a negative regulatory element of the BM niche that limits the size of the stem cell pool, is substantially upregulated (Abbuehl et al., 2017), while the expression of early growth response 1 (Erg1) and nestin was substantially downregulated (Nakahara et al., 2019; Li et al., 2020), which is correlated with the loss of proliferative and secretory activities of the cultured MSCs. The genetic alteration in BM niche cells leads to long-term functional changes in BM HSCs as overexpression of the transcription factor EGR1 in BM-MSCs improves the hematological stroma support via CCL28 and VCAM1 induction, and promoted the ex vivo expansion of transplantable CD34+CD90+ HSC (Li et al., 2020). Nakahara et al. (2019), following RNA sequencing screen, identified five genes encoding transcription factors (Klf7, Ostf1, Xbp1, Irf3, and Irf7) that fully restored the HSC niche function in the cultured BM-derived MSCs. Notably, the expression of the PαS cell-specific early B-cell factor (EBF) family of transcription factors, specifically Ebf1 and Ebf3, is essential for the preservation of HSCs and myeloid/lymphoid lineage output, and exhibits the memory phenotype that endures post-transplants (Derecka et al., 2020; Nakatani et al., 2023).
Clinically, MSC infusions are, in general, safe, with no adverse effects (Gao et al., 2016). Co-transplantation of BM-MSCs in the context of cord blood HSC transplantation accelerates short-term hematopoietic engraftment (de Lima et al., 2012). A positive impact of the MSC co-transplant has also been observed in lymphoma and myeloma patients with an increased production in memory and naïve T cells (Batorov et al., 2015) and reducing GvHD after allogeneic HSCT (Burnham et al., 2020; Macías-Sánchez et al., 2022), possibly through cytokine-mediated promotion of CD5+ B-cell production and downregulation of NK cells, CD4+ T cells, and/or macrophages (Lin et al., 2023). MSCs can also compromise the immune cell activity in leukemia patients, which may associate with disease recurrence after HSCT and MSC co-transplantation (Ning et al., 2008).
3.2 MSC mitochondrial transfer in normal hematopoiesis
BM niche cells form a heterogeneous population of cells that, upon stress activation, can exhibit specific single-cell responses which differentially affect the hematopoietic outcomes of surrounding HSPCs (Vannini et al., 2016; Vannini et al., 2019; Hinge et al., 2020; Liang et al., 2020). The crosstalk between the HSPC and BM microenvironment is critical for homeostasis and hematopoietic regeneration in response to blood formation emergencies. BM niche cells undergo massive damage to their mitochondrial function after myeloablative irradiation and/or pathogen infection, which eventually impairs hematopoietic regeneration (Figure 1). HSCs are exquisitely sensitive to irradiation, and even a low dose of ionizing radiation can increase the levels of reactive oxygen species (ROS) enough to activate p38MAPK-dependent senescence and apoptosis programs or even just impair their self-renewal capacity (Henry et al., 2020). LeptinR+ and nestin+ stromal cell subsets are also irradiation-sensitive, albeit at a much lesser degree (Severe et al., 2019), where moderate ROS levels are tolerated and even exert positive effects (Sheppard et al., 2022). At higher levels, ROS accumulation associated with inflammatory stress through the alteration of the RIG-I-Trim25-Keap1-NRF2 complex, which impairs the clonogenic capacity and bone-forming ability of BM-MSCs (Lou et al., 2022).
Intercellular mitochondrial transport, which refers to the transfer of either mitochondrial DNA or the entire organelle to supplement cellular energy demand in metabolically compromised recipient cells, has been documented both in vitro and in vivo in a variety of cells under physiological and pathophysiological conditions (Moschoi et al., 2016; Chandran et al., 2017; Marlein et al., 2017; Burt et al., 2019; Golan et al., 2020; Shanmughapriya et al., 2020; Brestoff et al., 2021; Saito et al., 2021; Borcherding et al., 2022). This intercellular mitochondrial transfer occurs through different means like intercellular tunneling nanotube (TnT) bridges or secreted small vesicles and frequently requires connexin 43 (Cx43)-containing gap junction (GJ) channels (Qin et al., 2021; Singh and Cancelas, 2021) (Figure 1).
Our previous research has demonstrated that hematopoietic regeneration and efficient blood formation after myeloablation depend on the transfer of damaging ROS from HSPC to the BM microenvironment, and hematopoietic Cx43 facilitates this transfer and prevents ROS-p38MAPK-p16/INK4a-mediated HSC senescence (Taniguchi Ishikawa et al., 2012) (Figure 1). Mitochondria are the major source of cellular bioenergetics and an important regulator of HSC fate decisions and energy homeostasis (Ito et al., 2004; Liu et al., 2015; Ito et al., 2016; Khacho et al., 2016; Luchsinger et al., 2016; Hu et al., 2018; Jin et al., 2018; Umemoto et al., 2018; Hinge et al., 2020). Under steady-state conditions, HSCs are primarily quiescent, rely on anaerobic glycolysis, and exhibit low mitochondrial ROS and mitochondrial membrane potential. However, upon regenerative stress, as found after myeloablative or infection stress, HSC undergoes rapid division and switches to mitochondrial oxidative phosphorylation to meet the energy demands (Ito et al., 2004; Maryanovich et al., 2015; Ito et al., 2016; Khacho et al., 2016; Luchsinger et al., 2016; Hu et al., 2018; Umemoto et al., 2018). Our group identified a new approach of hematopoietic recovery post-conditioning and demonstrated that following whole-body irradiation and HSPC transplantation, transplanted HSPCs through the transfer of mitochondria to the irradiated host MSC improve the metabolic recovery and proliferation of recipient BM stromal cells, which, in turn, increases the hematopoietic reconstitution (Golan et al., 2020) (Figure 1). We further identified that expression of Cx43 and low adenosine 5′-monophosphate-activated protein kinase (AMPK) activity in the HSPC compartment is crucial for rapid hematopoietic reconstitution after transplantation (Golan et al., 2020).
Furthermore, there is much evidence that BM-MSCs transfer mitochondria to HSPC to resist the oxidative stress via metabolic reprogramming and improve the hematopoietic reconstitution (Marlein et al., 2017; Mistry et al., 2019) (Figure 1). In infection stress hematopoiesis, BM-MSCs facilitate the transfer of mitochondria to HSPCs, which improves cellular metabolism by switching from glycolysis to oxidative phosphorylation and subsequently stimulates granulopoiesis (Mistry et al., 2019). This study further investigated the BM microenvironment heterogeneity in terms of mitochondria transfer and demonstrated that in response to infection, ROS-induced oxidative stress promotes phosp-protein kinase B (AKT) signaling, which drives Cx43 channel-mediated mitochondria transfer from BM-MSCs but not from other niche cells to HSPCs (Mistry et al., 2019). Recently, Jacoby et al. (2021), following ex vivo supplementation of functional mitochondria in human cord blood CD34+ cells, showed that mitochondrial augmentation boosts HSPC bioenergetics, enables T-cell expansion, and improves long-term engraftment in the NSGS mice (Figure 1). Although HSPCs and preclinical models highlight the beneficial effect of HSPC and BM-MSC mitochondrial exchange and the cellular bioenergetic shift in emergency hematopoiesis, the potential therapeutic implications of mitochondrial transfer in steady-state and stress hematopoiesis, the specific interaction between the HSPC and BM stromal cells, and mechanisms regulating intercellular mitochondrial exchange require further investigation.
4 Role of MSCs and their adipocytic lineage in hematological malignancies
Despite the therapeutic benefit of BM-MSCs, it has become poised in malignancy, and it has been observed that in hematological malignancies, remodeling of the BM niche creates an environment that favors malignant stem cells and hinders normal hematopoiesis (Forte et al., 2020). Additionally, through direct cell-to-cell contacts that frequently change the metabolism of cancer cells, dysregulation of extracellular matrix deposition, and modification of soluble factors/metabolites, BM-MSCs offer a survival advantage post-chemotherapy. Substantial evidence suggests that acute myelogenous leukemia (AML)-associated BM-MSCs show reduced CXCL12 and SCF expression with reduced sinusoidal and increased arterial cell populations (Arranz et al., 2014; Baryawno et al., 2019). Furthermore, an imbalanced BM-MSC/endothelial cell ratio in B-cell precursor acute lymphoblastic leukemia (BCP-ALL) post-therapy associated with a shorter disease-free survival, irrespective of minimal residual disease status (Oliveira et al., 2022). Likewise, myelodysplastic syndrome (MDS)-driving mutations require the presence of altered stromal cells and MSCs in an aged milieu, where they express senescence markers, elevated inflammatory molecules, and reduced levels of cytokines required for normal hematopoiesis (Huang et al., 2015; Fernando et al., 2019; Plakhova et al., 2023).
Myeloproliferative neoplasms have also been linked to increased BM-MSC apoptosis (Arranz et al., 2014; Corradi et al., 2018; Genitsari et al., 2018). Sympathetic nerve cells maintain BM Schwann cells and nestin+ cells, whereas leukemia progression is facilitated by a reduction in nerve fibers, which results in the apoptosis of nestin+ MSCs and a subsequent drop in CXCL12 levels. Treatment with β3-adrenergic agonists effectively restores the BM-MSC pool, followed by leukemia inhibition (Arranz et al., 2014). There are no notable alterations to the cytomorphology of BM-MSC or marker-based discrepancies during the progression of AML or B-ALL however, a substantial and varied change in the gene/protein expression patterns are observed, resulting in decreased cell proliferation and CFU-F-forming ability similar to that of senescent BM-MSCs (Le et al., 2016; Desbourdes et al., 2017; Genitsari et al., 2018; Baryawno et al., 2019; Fernando et al., 2019; Zhang et al., 2021).
Metabolically, proliferating transformed leukemic cells also consume glutamine and asparagine from their surrounding BM niches (Samudio and Konopleva, 2013; Bolzoni et al., 2016; Jiang et al., 2021). As glutamine and, to a lesser extent, asparagine are important for BM-MSC proliferation, lineage allocation, stemness, and osteogenesis (Zhou et al., 2019; Zhou et al., 2019; Sharma et al., 2021; Lungu et al., 2023), extracellular depletion of glutamine and asparagine, by proliferating leukemic cells, may impair MSC functionality and induce MSC senescence and osteopenia (Fernando et al., 2019; Gooding et al., 2019) (Figure 2).
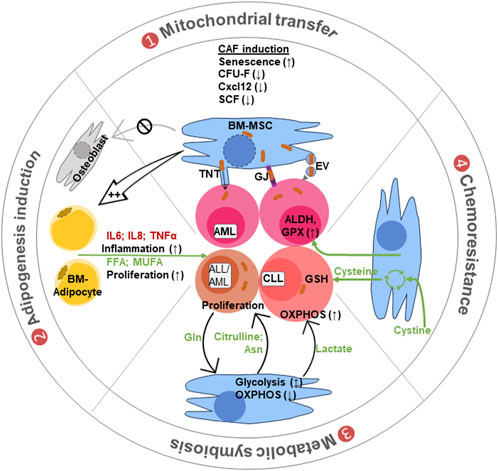
FIGURE 2. Different modes of BM-MSC crosstalk with leukemic cells. (1) Mitochondrial transfer: leukemic cell interaction converts the BM-MSCs into cancer-associated fibroblasts (CAFs) with altered features to support leukemia. Mitochondrial transfer from MSCs to leukemic cells occurs through tunneling nanotubes (TnTs), gap junctions (GJs), or via mitochondria carrying extracellular vesicles (EVs). (2) Adipogenic induction: these CAFs are biased toward adipocyte production with reduced osteoblast differentiation. Adipocytes support leukemia progression by secreting inflammatory cytokines and lipid molecules like free fatty acid (FFA) and monounsaturated fatty acids (MUFA) for cell growth and proliferation. (3) Metabolic symbiosis: the BM stromal cells supply different metabolites like lactate to maintain OXPHOS and amino acids like asparagine and glutamine (also citrulline), for biosynthetic needs of leukemic cells. (4) Chemoresistance: BM-MSCs also provide chemoresistance by providing cysteine for glutathione (GSH) synthesis. They also induce aldehyde dehydrogenase (ALDH) and glutathione peroxidase (GPX) in leukemic cells to metabolize chemotherapeutic drugs and reduce their effects.
4.1 BM-MSC-derived adipocytes positively regulate proliferation and survival of hematological malignant cells
BM stromal cells, especially adipocytes, are the source of different inflammatory molecules like IL-6, IL-8, IL-1B, IL-15, IL-17, and TNF-α; they can support clonal hematopoiesis and leukemia progression (Figure 2). The majority of studies have shown a positive correlation between adipogenicity with reduced osteogenic potential of BM-MSC and BM leukemia progression (Le et al., 2016; Fernando et al., 2019; Azadniv et al., 2020; Weickert et al., 2021; Kobari et al., 2022). The co-culture of mixed lineage leukemia (MLL) with BM-MSC results in downregulation of the expression of the early adipogenic cell fate inhibitor DLK1 in BM-MSCs (Weickert et al., 2021). Likewise, patients with multiple myeloma (MM) display higher levels of cytokines (ANG1, ENA-78, EGF, PDGF-AA/AB/BB, and TARC) that were linked to decreased osteoblastic differentiation and increased expression of CD36 and PPARγ with a skewing toward adipogenesis (Kobari et al., 2022). In contrast, Boyd et al. (2017), by performing in vitro co-culture and in vivo xenograft modeling, revealed that leukemia cells disrupt the adipocytic niche in BM, which led to the imbalanced regulation of endogenous HSPCs and hamper myelo-erythroid maturation. An in vivo administration of PPARγ agonists increases the BM adipocyte content and rescues healthy hematopoiesis (Boyd et al., 2017).
Proliferating leukemic cells consume large amounts of glucose (Warburg et al., 1927) for biosynthetic material and adenosine-5′-triphosphate (ATP) production, exceeding the biosynthetic needs (Vander Heiden et al., 2009). Continuous enhanced glycolysis faces a limitation of nicotine adenine dinucleotide, oxidized (NAD+). A sustained high rate of glycolysis results in rapid consumption of NAD+, which obliges tumor cells to consume extracellular pyruvate. To recover pyruvate, tumor cells use alternative fuels including fatty acids, glutamine, asparagine, serine, or aspartate (Sullivan et al., 2018; Li et al., 2022). Adipocytes represented the most important source of BM mono-unsaturated fatty acids, a preferred substrate for cancer cell membrane synthesis, which are less prone to fatty acid oxidation and ferroptosis than poly-unsaturated fatty acids (Hargrove et al., 2001; Gan, 2021; Mao et al., 2021). The differential effect of the BM microenvironment on leukemia metabolism is evident in B-cell acute lymphoblastic leukemia (B-ALL). The expression of stearoyl-coenzymeA-desaturase in BM B-ALL cells is reduced, making leukemic cells dependent on BM adipocytes to supply the mono-unsaturated free fatty acids for their survival and proliferation (Savino et al., 2020), illustrating the metabolic coupling of leukemic cells with the BM microenvironment.
4.2 BM-MSCs in chemotherapy resistance
BM-MSCs provide chemoresistance against retinoic acid (R)+, the proteasome inhibitor bortezomib (B), and the oxidative stress inducer arsenic trioxide (A) RBA (Liccardo et al., 2023). Nestin+ BM-MSCs support survival and chemotherapy relapse of AML through increased oxidative phosphorylation, tricarboxylic acid (TCA) cycle activity, and glutathione (GSH)-mediated antioxidant defense (Forte et al., 2020) with increased GPX activity. Interestingly, some reports suggest that the inflamed BM stromal cell microenvironment may increase the vulnerability of AML toward chemotherapies (Chen et al., 2023; Lisi-Vega and Méndez-Ferrer, 2023).
Zhang et al. (2012) demonstrated that chronic lymphocytic leukemia (CLL) cells express very low levels of the cysteine/glutamate antiporter xCT, preventing them from uptaking extracellular cystine for GSH synthesis. BM-MSCs actively uptake cystine and release cysteine for CLL uptake, thus helping in GSH synthesis and protection against ROS and drug-induced cytotoxicity (Figure 2). xCT inhibitors like (S)-4-carboxyphenylglycine and sulfasalazine induced chemotherapy sensitivity and cell death in CLLs. The anti-CD44 monoclonal antibody induces apoptosis and/or differentiation of AML cells via PI3K/AKT-p27 pathway upregulation in culture systems. Interestingly, BM stromal cells provide chemoresistance against anti-CD44 therapy by downregulating the AKT pathway in AML cells (Chen et al., 2015). Additionally, BM-MSCs activate the ALDH enzyme, which, through the TGFb-p38-ALDH pathway, converts xenobiotic aldehydes into less toxic carboxylic acid that is essential for drug metabolism and chemoresistance in AML (Sládek, 2003; Januchowski et al., 2013; Yuan et al., 2020) (Figure 2).
4.3 Metabolic crosstalk between leukemic cells and BM-MSCs
A stromal cell interaction with CLL causes a shift from mitochondrial to glycolysis-dependent bioenergetics through the NOTCH-cMYC signaling pathway that incurs increased proliferation and chemoresistance properties in CLL cells (Koppenol et al., 2011; Jitschin et al., 2015). Leukemic cells (ALL and BCP-ALL) secrete leukemic extracellular vesicles (EVs) that are consumed by the BM-MSCs (Johnson et al., 2016). Following EV ingestion, these BM-MSCs exhibit increased lactate generation, glycolysis, and mitochondrial metabolism. Similar improvements in BM-MSC glycolysis and mitochondrial metabolism by the CLL secretome improve stromal cell fitness, which aids in tumor cell survival (Lazarian et al., 2022). Vangapandu et al. (2017) have shown that the oxidative phosphorylation (OXPHOS) and ribonucleotide synthesis pathway are elevated in patient-derived CLL cells. The increase in OXPHOS and ribonucleotide was successfully decreased by the small-molecule complex I inhibitor IACS-010759; nevertheless, as a compensatory strategy, glycolysis increased. Concurrent administration of 2-deoxy-glucose and IACS-010759 suppressed this compensatory glycolysis and induced CLL cells apoptosis (Vangapandu et al., 2018). Protein kinase C-β (PKCβ) also plays a critical role in the BM stroma and CLL cell interaction, where they provide a cancer-supportive inflammatory microenvironment (vom Stein et al., 2023), chemoresistance (Amigo-Jiménez et al., 2015), and establish metabolic symbiosis (von Heydebrand et al., 2021). An interaction through PKCβ reduced glycolysis and insulin signaling in the BM-stromal cells and simultaneously increased the glycolysis/lactate production in CLL cells. It has been proposed that an elevated lactate environment aids in immunosuppression (Chen et al., 2022), and the leukemic cells can utilize it to run the TCA cycle or vice versa (Saulle et al., 2021). Extracellular lactate binds to the lactate receptor GPCR81 and takes part in the cellular metabolism after being taken up through MCT4 channels. Furthermore, intracellular lactate-mediated histone-lysine lactylation and protein lactylation are crucial for lactate-mediated signaling (Chen et al., 2022; Liu et al., 2022; Zhang et al., 2023). Thus, targeting lactate consumption is also helpful in reducing cell proliferation and leukemic burden (Yu et al., 2020; Saulle et al., 2021; Wang et al., 2021) (Figure 2).
L-asparaginase therapy achieves approximately 90% remission rate in B-ALL when combined with doxorubicin, vincristine, and prednisone (Parmentier et al., 2015; Tabe et al., 2019; Wang et al., 2023). BM-MSCs contribute to B-ALL resistance against L-asparaginase therapy by supplying asparagine to leukemic cells (Iwamoto et al., 2007). L-asparaginase-treated leukemic blasts synthesize glutamine to be uptaken by BM-MSCs, and in return, BM-MSCs convert glutamine into asparagine that cycles back to leukemic cells (Chiu et al., 2021). BM adipocytes can also secrete glutamine and asparagine to prevent leukemic cell death by L-asparaginase, which may be at the root of the poor response of obese patients to conventional L-asparaginase containing B-ALL therapy (Samudio and Konopleva, 2013) (Figure 2).
T-ALL cells are specifically dependent on arginine levels and consume arginine produced by the BM microenvironment (Chiu et al., 2021). Based on this fact, the PEGylated arginase (pegargiminase) has been used for clinical trials (De Santo et al., 2018). However, some studies also show that BM-MSCs synthesize and secrete citrulline, which is consumed by T-ALL cells to gain chemoresistance against pegargiminase (Sugimura et al., 1990; Kwong-Lam and Chi-Fung, 2013).
Finally, extracellular glutamine is specifically utilized by AML, MM, and CLL cells for anaplerosis and amino acid production. It also aids in mTOR activation, permitting enhanced protein synthesis and leukemia burden (Samudio and Konopleva, 2013; Bolzoni et al., 2016; Chiu et al., 2021) (Figure 2).
4.4 BM mitochondrial transfer and leukemic chemoresistance
AML cells prefer OXPHOS and, in many situations, fatty acid oxidation (FAO) over glycolysis (Tabe and Konopleva, 2023). High aerobic glycolysis or “Warburg effect” in AML is related to better treatment efficacy and increased lifespan (Herst et al., 2011). In contrast, increased OXPHOS, mitochondrial mass, ROS, and FAO are associated with greater chemoresistance against medications such as cytarabine in AML (Moschoi et al., 2016; Farge et al., 2017; de Beauchamp et al., 2022). Conventional chemotherapy-resistant leukemic cells typically rely on a very active mitochondrial metabolism (de Beauchamp et al., 2022). These resistant cells modify their surrounding BM niches and reprogram them into cancer-associated fibroblasts (CAFs), which, in turn, help the leukemic cells by transferring functional mitochondria via tunneling nanotubes (Burt et al., 2019). Nonetheless, inhibiting CAF production with corticosteroids, intercellular communication with vincristine, and ROS detoxification with N-acetyl cysteine, glutathione, and diphenyleneiodonium selectively reduced the leukemia burden and improve survival (Burt et al., 2019).
A growing body of data indicate that leukemia cell metabolism is especially increased by mitochondrial donation from BM-MSCs, which also increases the likelihood of leukemia cell survival after chemotherapy (Moschoi et al., 2016; Marlein et al., 2017; Burt et al., 2019; Forte et al., 2020; Saito et al., 2021). The increased frequency of nestin+ MSCs in AML patients’ BM and in ALL murine xenograft models represents a cell-to-cell contact-dependent ROS-detoxifying mechanism that permits mitochondrial transfer to promote chemoresistance and recurrence (Wang et al., 2018; Burt et al., 2019; Moore et al., 2019) (Figure 2). In response to chemotherapy, such as daunorubicin or cytarabine treatment, which caused ROS and damaged mitochondria accumulation, formation of TnT in B-ALL patients provides a survival advantage through mitochondrial transfer from BM-MSCs. Inhibiting TnT formation with vincristine, however, limits mitochondrial transport from BM-MSCs and sensitizes leukemic cells against chemotherapy (Burt et al., 2019). Similarly, healthy mitochondria are acquired by AML and MM cells from BM-MSCs via TnT, and this acquisition is further improved, following chemotherapy (Moschoi et al., 2016; Matula et al., 2021; Saito et al., 2021). Following transfer of mitochondria, AML cell-derived NADPH oxidase 2 increases ROS accumulation in the surrounding BM-MSCs and activates their mitochondrial biogenesis through the peroxisome proliferator-activated receptor-gamma coactivator-1 alpha (PGC1α). These extra mitochondria are transferred from BM-MSCs to AML cells, trigger compensatory activation of mitochondrial OXPHOS and ATP generation, and, therefore, improve chemoresistance (Marlein et al., 2017; Shafat et al., 2017; Marlein et al., 2018). Inhibition of electron transport complex I by metformin reduces mitochondrial transfer from BM-MSC to AML, resulting in enhanced sensitivity of AML cells to cytarabine treatment (You et al., 2022), while IACS-010759, a small complex I inhibitor, enhances mitochondrial trafficking from BM-MSC, triggers compensatory adaptation of leukemia cells to energetic stress via endogenous mitochondrial fission and mitophagy, and facilitates AML resistance (Saito et al., 2021). As leukemia-initiating AML cells can reside in the CD34+CD38+ compartment, treatment with the recombinant antibody daratumumab against CD38 inhibits mitochondrial transfer to AML blasts, inhibiting the leukemia growth (Mistry et al., 2019). ATRA treatment has been shown to increase the CD38 expression in multiple myeloma, where it helps in mitochondrial transfer from BM-MSCs to myeloma cells (Marlein et al., 2019). Extracellular vesicles (EVs) released by MSCs have also been demonstrated to transfer mitochondria to neighboring cells, albeit the precise role of EV-mediated mitochondrial transport in leukemia remains unclear (Phinney et al., 2015; Griessinger et al., 2017) and necessitates more investigation to determine the precise mechanism for which BM-MSC mitotransfer may be involved in elimination of minimal residual leukemia.
5 Concluding remarks
BM-MSCs represent a powerful tool due to their regenerative potential and their immunomodulatory characteristics. Although challenges are associated with MSC isolation, limited ex vivo expansion, short life after transplantation, and the mode of in vivo administration must be addressed in order to improve hematopoietic outcomes. Metabolic coupling between BM-MSCs and HSCs is a crucial mechanism of fine-tuning of regeneration hematopoiesis, frequently hijacked by hematological malignancies. Coupling is highly dependent on the metabolic state of both cell compartments, and a major mediator of such metabolic coupling resides in their ability to exchange mitochondria and potentially other organelles. Identification of the molecular signals controlling the mitochondria exchange and overall metabolic coupling opens significant vulnerability opportunities for intervention and therapy in hematological malignancies.
Author contributions
AS: conceptualization, investigation, and writing–original draft. PP: conceptualization, investigation, and writing–original draft. JC: conceptualization, funding acquisition, investigation, supervision, writing–original draft, and writing–review and editing.
Funding
The author(s) declare that no financial support was received for the research, authorship, and/or publication of this article.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors, and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abbuehl, J. P., Tatarova, Z., Held, W., and Huelsken, J. (2017). Long-term engraftment of primary bone marrow stromal cells repairs niche damage and improves hematopoietic stem cell transplantation. Cell Stem Cell 21 (2), 241–255. doi:10.1016/j.stem.2017.07.004
Abkowitz, J. L., Broudy, V. C., Bennett, L. G., Zsebo, K. M., and Martin, F. H. (1992). Absence of abnormalities of c-kit or its ligand in two patients with Diamond-Blackfan anemia. Blood 79 (1), 25–28. doi:10.1182/blood.v79.1.25.bloodjournal79125
Amigo-Jiménez, I., Bailón, E., Aguilera-Montilla, N., Terol, M. J., García-Marco, J. A., and García-Pardo, A. (2015). Bone marrow stroma-induced resistance of chronic lymphocytic leukemia cells to arsenic trioxide involves Mcl-1 upregulation and is overcome by inhibiting the PI3Kδ or PKCβ signaling pathways. Oncotarget 6 (42), 44832–44848. doi:10.18632/oncotarget.6265
Arranz, L., Sánchez-Aguilera, A., Martín-Pérez, D., Isern, J., Langa, X., Tzankov, A., et al. (2014). Neuropathy of haematopoietic stem cell niche is essential for myeloproliferative neoplasms. Nature 512 (7512), 78–81. doi:10.1038/nature13383
Azadniv, M., Myers, J. R., McMurray, H. R., Guo, N., Rock, P., Coppage, M. L., et al. (2020). Bone marrow mesenchymal stromal cells from acute myelogenous leukemia patients demonstrate adipogenic differentiation propensity with implications for leukemia cell support. Leukemia 34 (2), 391–403. doi:10.1038/s41375-019-0568-8
Baryawno, N., Przybylski, D., Kowalczyk, M. S., Kfoury, Y., Severe, N., Gustafsson, K., et al. (2019). A cellular taxonomy of the bone marrow stroma in homeostasis and leukemia. Cell 177 (7), 1915–1932. doi:10.1016/j.cell.2019.04.040
Batorov, E. V., Shevela, E. Y., Tikhonova, M. A., Batorova, D. S., Ushakova, G. Y., Sizikova, S. A., et al. (2015). Mesenchymal stromal cells improve early lymphocyte recovery and T cell reconstitution after autologous hematopoietic stem cell transplantation in patients with malignant lymphomas. Cell. Immunol. 297 (2), 80–86. doi:10.1016/j.cellimm.2015.07.001
Batsivari, A., Haltalli, M. L. R., Passaro, D., Pospori, C., Lo Celso, C., and Bonnet, D. (2020). Dynamic responses of the haematopoietic stem cell niche to diverse stresses. Nat. Cell Biol. 22 (1), 7–17. doi:10.1038/s41556-019-0444-9
Bernardo, M. E., and Fibbe, W. E. (2013). Mesenchymal stromal cells: sensors and switchers of inflammation. Cell Stem Cell 13 (4), 392–402. doi:10.1016/j.stem.2013.09.006
Bernardo, M. E., and Fibbe, W. E. (2015). Mesenchymal stromal cells and hematopoietic stem cell transplantation. Immunol. Lett. 168 (2), 215–221. doi:10.1016/j.imlet.2015.06.013
Bolzoni, M., Chiu, M., Accardi, F., Vescovini, R., Airoldi, I., Storti, P., et al. (2016). Dependence on glutamine uptake and glutamine addiction characterize myeloma cells: a new attractive target. Blood 128 (5), 667–679. doi:10.1182/blood-2016-01-690743
Borcherding, N., Jia, W., Giwa, R., Field, R. L., Moley, J. R., Kopecky, B. J., et al. (2022). Dietary lipids inhibit mitochondria transfer to macrophages to divert adipocyte-derived mitochondria into the blood. Cell Metab. 34 (10), 1499–1513.e8. doi:10.1016/j.cmet.2022.08.010
Boyd, A. L., Reid, J. C., Salci, K. R., Aslostovar, L., Benoit, Y. D., Shapovalova, Z., et al. (2017). Acute myeloid leukaemia disrupts endogenous myelo-erythropoiesis by compromising the adipocyte bone marrow niche. Nat. Cell Biol. 19 (11), 1336–1347. doi:10.1038/ncb3625
Breitbach, M., Kimura, K., Luis, T. C., Fuegemann, C. J., Woll, P. S., Hesse, M., et al. (2018). Vivo labeling by CD73 marks multipotent stromal cells and highlights endothelial heterogeneity in the bone marrow niche. Cell Stem Cell 22 (2), 262–276 e267. doi:10.1016/j.stem.2018.01.008
Brestoff, J. R., Wilen, C. B., Moley, J. R., Li, Y., Zou, W., Malvin, N. P., et al. (2021). Intercellular mitochondria transfer to macrophages regulates white adipose tissue homeostasis and is impaired in obesity. Cell Metab. 33 (2), 270–282.e8. doi:10.1016/j.cmet.2020.11.008
Buhring, H. J., Battula, V. L., Treml, S., Schewe, B., Kanz, L., and Vogel, W. (2007). Novel markers for the prospective isolation of human MSC. Ann. N. Y. Acad. Sci. 1106, 262–271. doi:10.1196/annals.1392.000
Burnham, A. J., Daley-Bauer, L. P., and Horwitz, E. M. (2020). Mesenchymal stromal cells in hematopoietic cell transplantation. Blood Adv. 4 (22), 5877–5887. doi:10.1182/bloodadvances.2020002646
Burt, R., Dey, A., Aref, S., Aguiar, M., Akarca, A., Bailey, K., et al. (2019). Activated stromal cells transfer mitochondria to rescue acute lymphoblastic leukemia cells from oxidative stress. Blood 134 (17), 1415–1429. doi:10.1182/blood.2019001398
Butler, J. M., Nolan, D. J., Vertes, E. L., Varnum-Finney, B., Kobayashi, H., Hooper, A. T., et al. (2010). Endothelial cells are essential for the self-renewal and repopulation of Notch-dependent hematopoietic stem cells. Cell Stem Cell 6 (3), 251–264. doi:10.1016/j.stem.2010.02.001
Chandran, V., Gao, K., Swarup, V., Versano, R., Dong, H., Jordan, M. C., et al. (2017). Inducible and reversible phenotypes in a novel mouse model of Friedreich's Ataxia. Elife 6, e30054. doi:10.7554/eLife.30054
Chen, L., Huang, L., Gu, Y., Cang, W., Sun, P., and Xiang, Y. (2022). Lactate-lactylation hands between metabolic reprogramming and immunosuppression. Int. J. Mol. Sci. 23 (19), 11943. doi:10.3390/ijms231911943
Chen, L., Pronk, E., van Dijk, C., Bian, Y., Feyen, J., van Tienhoven, T., et al. (2023). A single-cell taxonomy predicts inflammatory niche remodeling to drive tissue failure and outcome in human AML. Blood Cancer Discov. 4 (5), 394–417. doi:10.1158/2643-3230.BCD-23-0043
Chen, P., Huang, H., Wu, J., Lu, R., Wu, Y., Jiang, X., et al. (2015). Bone marrow stromal cells protect acute myeloid leukemia cells from anti-CD44 therapy partly through regulating PI3K/Akt-p27(Kip1) axis. Mol. Carcinog. 54 (12), 1678–1685. doi:10.1002/mc.22239
Chen, Q., Liu, Y., Jeong, H. W., Stehling, M., Dinh, V. V., Zhou, B., et al. (2019). Apelin(+) endothelial niche cells control hematopoiesis and mediate vascular regeneration after myeloablative injury. Cell Stem Cell 25 (6), 768–783. doi:10.1016/j.stem.2019.10.006
Chen, W., Baylink, D. J., Brier-Jones, J., Neises, A., Kiroyan, J. B., Rundle, C. H., et al. (2015). PDGFB-based stem cell gene therapy increases bone strength in the mouse. Proc. Natl. Acad. Sci. U. S. A. 112 (29), E3893–E3900. doi:10.1073/pnas.1501759112
Chiu, M., Taurino, G., Bianchi, M. G., and Bussolati, O. (2021). The role of amino acids in the crosstalk between mesenchymal stromal cells and neoplastic cells in the hematopoietic niche. Front. Cell Dev. Biol. 9, 714755. doi:10.3389/fcell.2021.714755
Chiu, M., Taurino, G., Dander, E., Bardelli, D., Fallati, A., Andreoli, R., et al. (2021). ALL blasts drive primary mesenchymal stromal cells to increase asparagine availability during asparaginase treatment. Blood Adv. 5 (23), 5164–5178. doi:10.1182/bloodadvances.2020004041
Corradi, G., Baldazzi, C., Očadlíková, D., Marconi, G., Parisi, S., Testoni, N., et al. (2018). Mesenchymal stromal cells from myelodysplastic and acute myeloid leukemia patients display in vitro reduced proliferative potential and similar capacity to support leukemia cell survival. Stem Cell Res. Ther. 9 (1), 271. doi:10.1186/s13287-018-1013-z
Crane, G. M., Jeffery, E., and Morrison, S. J. (2017). Adult haematopoietic stem cell niches. Nat. Rev. Immunol. 17 (9), 573–590. doi:10.1038/nri.2017.53
Crippa, S., Santi, L., Berti, M., De Ponti, G., and Bernardo, M. E. (2021). Role of ex vivo expanded mesenchymal stromal cells in determining hematopoietic stem cell transplantation outcome. Front. Cell Dev. Biol. 9, 663316. doi:10.3389/fcell.2021.663316
Crisan, M., Yap, S., Casteilla, L., Chen, C. W., Corselli, M., Park, T. S., et al. (2008). A perivascular origin for mesenchymal stem cells in multiple human organs. Cell Stem Cell 3 (3), 301–313. doi:10.1016/j.stem.2008.07.003
de Beauchamp, L., Himonas, E., and Helgason, G. V. (2022). Mitochondrial metabolism as a potential therapeutic target in myeloid leukaemia. Leukemia 36 (1), 1–12. doi:10.1038/s41375-021-01416-w
de Lima, M., McNiece, I., Robinson, S. N., Munsell, M., Eapen, M., Horowitz, M., et al. (2012). Cord-blood engraftment with ex vivo mesenchymal-cell coculture. N. Engl. J. Med. 367 (24), 2305–2315. doi:10.1056/NEJMoa1207285
Derecka, M., Herman, J. S., Cauchy, P., Ramamoorthy, S., Lupar, E., Grun, D., et al. (2020). EBF1-deficient bone marrow stroma elicits persistent changes in HSC potential. Nat. Immunol. 21 (3), 261–273. doi:10.1038/s41590-020-0595-7
De Santo, C., Cheng, P., Beggs, A., Egan, S., Bessudo, A., and Mussai, F. (2018). Metabolic therapy with PEG-arginase induces a sustained complete remission in immunotherapy-resistant melanoma. J. Hematol. Oncol. 11 (1), 68. doi:10.1186/s13045-018-0612-6
Desbourdes, L., Javary, J., Charbonnier, T., Ishac, N., Bourgeais, J., Iltis, A., et al. (2017). Alteration analysis of bone marrow mesenchymal stromal cells from de novo acute myeloid leukemia patients at diagnosis. Stem Cells Dev. 26 (10), 709–722. doi:10.1089/scd.2016.0295
Ding, L., Saunders, T. L., Enikolopov, G., and Morrison, S. J. (2012). Endothelial and perivascular cells maintain haematopoietic stem cells. Nature 481 (7382), 457–462. doi:10.1038/nature10783
Doan, P. L., Himburg, H. A., Helms, K., Russell, J. L., Fixsen, E., Quarmyne, M., et al. (2013). Epidermal growth factor regulates hematopoietic regeneration after radiation injury. Nat. Med. 19 (3), 295–304. doi:10.1038/nm.3070
Dominici, M., Le Blanc, K., Mueller, I., Slaper-Cortenbach, I., Marini, F., Krause, D., et al. (2006). Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 8 (4), 315–317. doi:10.1080/14653240600855905
Eggenhofer, E., Benseler, V., Kroemer, A., Popp, F. C., Geissler, E. K., Schlitt, H. J., et al. (2012). Mesenchymal stem cells are short-lived and do not migrate beyond the lungs after intravenous infusion. Front. Immunol. 3, 297. doi:10.3389/fimmu.2012.00297
Farge, T., Saland, E., de Toni, F., Aroua, N., Hosseini, M., Perry, R., et al. (2017). Chemotherapy-resistant human acute myeloid leukemia cells are not enriched for leukemic stem cells but require oxidative metabolism. Cancer Discov. 7 (7), 716–735. doi:10.1158/2159-8290.CD-16-0441
Fernando, R. C., Mazzotti, D. R., Azevedo, H., Sandes, A. F., Rizzatti, E. G., de Oliveira, M. B., et al. (2019). Transcriptome analysis of mesenchymal stem cells from multiple myeloma patients reveals downregulation of genes involved in cell cycle progression, immune response, and bone metabolism. Sci. Rep. 9 (1), 1056. doi:10.1038/s41598-018-38314-8
Forte, D., Garcia-Fernandez, M., Sanchez-Aguilera, A., Stavropoulou, V., Fielding, C., Martin-Perez, D., et al. (2020). Bone marrow mesenchymal stem cells support acute myeloid leukemia bioenergetics and enhance antioxidant defense and escape from chemotherapy. Cell Metab. 32 (5), 829–843. doi:10.1016/j.cmet.2020.09.001
Galleu, A., Riffo-Vasquez, Y., Trento, C., Lomas, C., Dolcetti, L., Cheung, T. S., et al. (2017). Apoptosis in mesenchymal stromal cells induces in vivo recipient-mediated immunomodulation. Sci. Transl. Med. 9 (416), eaam7828. doi:10.1126/scitranslmed.aam7828
Gan, B. (2021). Mitochondrial regulation of ferroptosis. J. Cell Biol. 220 (9), e202105043. doi:10.1083/jcb.202105043
Gao, L., Zhang, Y., Hu, B., Liu, J., Kong, P., Lou, S., et al. (2016). Phase II multicenter, randomized, double-blind controlled study of efficacy and safety of umbilical cord–derived mesenchymal stromal cells in the prophylaxis of chronic graft-versus-host disease after HLA-haploidentical stem-cell transplantation. J. Clin. Oncol. 34 (24), 2843–2850. doi:10.1200/JCO.2015.65.3642
Genitsari, S., Stiakaki, E., Perdikogianni, C., Martimianaki, G., Pelagiadis, I., Pesmatzoglou, M., et al. (2018). Biological features of bone marrow mesenchymal stromal cells in childhood acute lymphoblastic leukemia. Turk J. Haematol. 35 (1), 19–26. doi:10.4274/tjh.2017.0209
Ghazanfari, R., Li, H., Zacharaki, D., Lim, H. C., and Scheding, S. (2016). Human non-hematopoietic cd271(pos)/CD140a(low/neg) bone marrow stroma cells fulfill stringent stem cell criteria in serial transplantations. Stem Cells Dev. 25 (21), 1652–1658. doi:10.1089/scd.2016.0169
Golan, K., Singh, A. K., Kollet, O., Bertagna, M., Althoff, M. J., Khatib-Massalha, E., et al. (2020). Bone marrow regeneration requires mitochondrial transfer from donor Cx43-expressing hematopoietic progenitors to stroma. Blood 136 (23), 2607–2619. doi:10.1182/blood.2020005399
Gooding, S., Olechnowicz, S. W. Z., Morris, E. V., Armitage, A. E., Arezes, J., Frost, J., et al. (2019). Transcriptomic profiling of the myeloma bone-lining niche reveals BMP signalling inhibition to improve bone disease. Nat. Commun. 10 (1), 4533. doi:10.1038/s41467-019-12296-1
Greenbaum, A., Hsu, Y. M., Day, R. B., Schuettpelz, L. G., Christopher, M. J., Borgerding, J. N., et al. (2013). CXCL12 in early mesenchymal progenitors is required for haematopoietic stem-cell maintenance. Nature 495 (7440), 227–230. doi:10.1038/nature11926
Griessinger, E., Moschoi, R., Biondani, G., and Peyron, J.-F. (2017). Mitochondrial transfer in the leukemia microenvironment. Trends Cancer 3 (12), 828–839. doi:10.1016/j.trecan.2017.10.003
Guo, P., Poulos, M. G., Palikuqi, B., Badwe, C. R., Lis, R., Kunar, B., et al. (2017). Endothelial jagged-2 sustains hematopoietic stem and progenitor reconstitution after myelosuppression. J. Clin. Invest. 127 (12), 4242–4256. doi:10.1172/JCI92309
Han, Y., Yang, J., Fang, J., Zhou, Y., Candi, E., Wang, J., et al. (2022). The secretion profile of mesenchymal stem cells and potential applications in treating human diseases. Signal Transduct. Target Ther. 7 (1), 92. doi:10.1038/s41392-022-00932-0
Hargrove, R. L., Etherton, T. D., Pearson, T. A., Harrison, E. H., and Kris-Etherton, P. M. (2001). Low fat and high monounsaturated fat diets decrease human low density lipoprotein oxidative susceptibility in vitro. J. Nutr. 131 (6), 1758–1763. doi:10.1093/jn/131.6.1758
Henry, E., Souissi-Sahraoui, I., Deynoux, M., Lefevre, A., Barroca, V., Campalans, A., et al. (2020). Human hematopoietic stem/progenitor cells display reactive oxygen species-dependent long-term hematopoietic defects after exposure to low doses of ionizing radiations. Haematologica 105 (8), 2044–2055. doi:10.3324/haematol.2019.226936
Herst, P. M., Howman, R. A., Neeson, P. J., Berridge, M. V., and Ritchie, D. S. (2011). The level of glycolytic metabolism in acute myeloid leukemia blasts at diagnosis is prognostic for clinical outcome. J. Leukoc. Biol. 89 (1), 51–55. doi:10.1189/jlb.0710417
Himburg, H. A., Termini, C. M., Schlussel, L., Kan, J., Li, M., Zhao, L., et al. (2018). Distinct bone marrow sources of pleiotrophin control hematopoietic stem cell maintenance and regeneration. Cell Stem Cell 23 (3), 370–381. doi:10.1016/j.stem.2018.07.003
Hinge, A., He, J., Bartram, J., Javier, J., Xu, J., Fjellman, E., et al. (2020). Asymmetrically segregated mitochondria provide cellular memory of hematopoietic stem cell replicative history and drive HSC attrition. Cell Stem Cell 26 (3), 420–430. doi:10.1016/j.stem.2020.01.016
Hooper, A. T., Butler, J. M., Nolan, D. J., Kranz, A., Iida, K., Kobayashi, M., et al. (2009). Engraftment and reconstitution of hematopoiesis is dependent on VEGFR2-mediated regeneration of sinusoidal endothelial cells. Cell Stem Cell 4 (3), 263–274. doi:10.1016/j.stem.2009.01.006
Hu, M., Zeng, H., Chen, S., Xu, Y., Wang, S., Tang, Y., et al. (2018). SRC-3 is involved in maintaining hematopoietic stem cell quiescence by regulation of mitochondrial metabolism in mice. Blood 132 (9), 911–923. doi:10.1182/blood-2018-02-831669
Hu, X., Garcia, M., Weng, L., Jung, X., Murakami, J. L., Kumar, B., et al. (2016). Identification of a common mesenchymal stromal progenitor for the adult haematopoietic niche. Nat. Commun. 7, 13095. doi:10.1038/ncomms13095
Huang, J. C., Basu, S. K., Zhao, X., Chien, S., Fang, M., Oehler, V. G., et al. (2015). Mesenchymal stromal cells derived from acute myeloid leukemia bone marrow exhibit aberrant cytogenetics and cytokine elaboration. Blood Cancer J. 5 (4), e302. doi:10.1038/bcj.2015.17
Islam, M. N., Das, S. R., Emin, M. T., Wei, M., Sun, L., Westphalen, K., et al. (2012). Mitochondrial transfer from bone-marrow-derived stromal cells to pulmonary alveoli protects against acute lung injury. Nat. Med. 18 (5), 759–765. doi:10.1038/nm.2736
Itkin, T., Ludin, A., Gradus, B., Gur-Cohen, S., Kalinkovich, A., Schajnovitz, A., et al. (2012). FGF-2 expands murine hematopoietic stem and progenitor cells via proliferation of stromal cells, c-Kit activation, and CXCL12 down-regulation. Blood 120 (9), 1843–1855. doi:10.1182/blood-2011-11-394692
Ito, K., Hirao, A., Arai, F., Matsuoka, S., Takubo, K., Hamaguchi, I., et al. (2004). Regulation of oxidative stress by ATM is required for self-renewal of haematopoietic stem cells. Nature 431 (7011), 997–1002. doi:10.1038/nature02989
Ito, K., Turcotte, R., Cui, J., Zimmerman, S. E., Pinho, S., Mizoguchi, T., et al. (2016). Self-renewal of a purified Tie2+ hematopoietic stem cell population relies on mitochondrial clearance. Science 354 (6316), 1156–1160. doi:10.1126/science.aaf5530
Iwamoto, S., Mihara, K., Downing, J. R., Pui, C. H., and Campana, D. (2007). Mesenchymal cells regulate the response of acute lymphoblastic leukemia cells to asparaginase. J. Clin. Invest. 117 (4), 1049–1057. doi:10.1172/JCI30235
Jacoby, E., Ben Yakir-Blumkin, M., Blumenfeld-Kan, S., Brody, Y., Meir, A., Melamed-Book, N., et al. (2021). Mitochondrial augmentation of CD34(+) cells from healthy donors and patients with mitochondrial DNA disorders confers functional benefit. NPJ Regen. Med. 6 (1), 58. doi:10.1038/s41536-021-00167-7
Januchowski, R., Wojtowicz, K., and Zabel, M. (2013). The role of aldehyde dehydrogenase (ALDH) in cancer drug resistance. Biomed. Pharmacother. 67 (7), 669–680. doi:10.1016/j.biopha.2013.04.005
Jiang, J., Batra, S., and Zhang, J. (2021). Asparagine: a metabolite to Be targeted in cancers. Metabolites 11 (6), 402. doi:10.3390/metabo11060402
Jin, G., Xu, C., Zhang, X., Long, J., Rezaeian, A. H., Liu, C., et al. (2018). Atad3a suppresses Pink1-dependent mitophagy to maintain homeostasis of hematopoietic progenitor cells. Nat. Immunol. 19 (1), 29–40. doi:10.1038/s41590-017-0002-1
Jitschin, R., Braun, M., Qorraj, M., Saul, D., Le Blanc, K., Zenz, T., et al. (2015). Stromal cell–mediated glycolytic switch in CLL cells involves Notch-c-Myc signaling. Blood 125 (22), 3432–3436. doi:10.1182/blood-2014-10-607036
Johnson, S. M., Dempsey, C., Chadwick, A., Harrison, S., Liu, J., Di, Y., et al. (2016). Metabolic reprogramming of bone marrow stromal cells by leukemic extracellular vesicles in acute lymphoblastic leukemia. Blood 128 (3), 453–456. doi:10.1182/blood-2015-12-688051
Kara, N., Xue, Y., Zhao, Z., Murphy, M. M., Comazzetto, S., Lesser, A., et al. (2023). Endothelial and Leptin Receptor(+) cells promote the maintenance of stem cells and hematopoiesis in early postnatal murine bone marrow. Dev. Cell 58 (5), 348–360.e6. doi:10.1016/j.devcel.2023.02.003
Keating, A. (2012). Mesenchymal stromal cells: new directions. Cell Stem Cell 10 (6), 709–716. doi:10.1016/j.stem.2012.05.015
Khacho, M., Clark, A., Svoboda, D. S., Azzi, J., MacLaurin, J. G., Meghaizel, C., et al. (2016). Mitochondrial dynamics impacts stem cell identity and fate decisions by regulating a nuclear transcriptional program. Cell Stem Cell 19 (2), 232–247. doi:10.1016/j.stem.2016.04.015
Kobari, L., Auclair, M., Piau, O., Ferrand, N., Zaoui, M., Delhommeau, F., et al. (2022). Circulating cytokines present in multiple myeloma patients inhibit the osteoblastic differentiation of adipose stem cells. Leukemia 36 (2), 540–548. doi:10.1038/s41375-021-01428-6
Koppenol, W. H., Bounds, P. L., and Dang, C. V. (2011). Otto Warburg's contributions to current concepts of cancer metabolism. Nat. Rev. Cancer 11 (5), 325–337. doi:10.1038/nrc3038
Kunisaki, Y., Bruns, I., Scheiermann, C., Ahmed, J., Pinho, S., Zhang, D., et al. (2013). Arteriolar niches maintain haematopoietic stem cell quiescence. Nature 502 (7473), 637–643. doi:10.1038/nature12612
Kurtzberg, J., Abdel-Azim, H., Carpenter, P., Chaudhury, S., Horn, B., Mahadeo, K., et al. (2020). A phase 3, single-arm, prospective study of remestemcel-L, ex vivo culture-expanded adult human mesenchymal stromal cells for the treatment of pediatric patients who failed to respond to steroid treatment for acute graft-versus-host disease. Biol. Blood Marrow Transpl. 26 (5), 845–854. doi:10.1016/j.bbmt.2020.01.018
Kwong-Lam, F., and Chi-Fung, C. G. (2013). Vincristine could partly suppress stromal support to T-ALL blasts during pegylated arginase I treatment. Exp. Hematol. Oncol. 2 (1), 11. doi:10.1186/2162-3619-2-11
Lazarian, G., Ferreira, J., Morilla, I., Saindoy, E., Bisio, V., Dulphy, N., et al. (2022). Metabolic reprogramming of bone marrow stromal cells in chronic lymphocytic leukemia. Blood 140 (Suppl. 1), 6996–6997. doi:10.1182/blood-2022-158450
Le, Y., Fraineau, S., Chandran, P., Sabloff, M., Brand, M., Lavoie, J. R., et al. (2016). Adipogenic mesenchymal stromal cells from bone marrow and their hematopoietic supportive role: towards understanding the permissive marrow microenvironment in acute myeloid leukemia. Stem Cell Rev. Rep. 12 (2), 235–244. doi:10.1007/s12015-015-9639-z
Le Blanc, K., Rasmusson, I., Sundberg, B., Gotherstrom, C., Hassan, M., Uzunel, M., et al. (2004). Treatment of severe acute graft-versus-host disease with third party haploidentical mesenchymal stem cells. Lancet 363 (9419), 1439–1441. doi:10.1016/S0140-6736(04)16104-7
Le Blanc, K., Samuelsson, H., Gustafsson, B., Remberger, M., Sundberg, B., Arvidson, J., et al. (2007). Transplantation of mesenchymal stem cells to enhance engraftment of hematopoietic stem cells. Leukemia 21 (8), 1733–1738. doi:10.1038/sj.leu.2404777
Li, H., Braunig, S., Dhapolar, P., Karlsson, G., Lang, S., and Scheding, S. (2023). Identification of phenotypically, functionally, and anatomically distinct stromal niche populations in human bone marrow based on single-cell RNA sequencing. Elife 12. doi:10.7554/elife.81656
Li, H., Ghazanfari, R., Zacharaki, D., Ditzel, N., Isern, J., Ekblom, M., et al. (2014). Low/negative expression of PDGFR-alpha identifies the candidate primary mesenchymal stromal cells in adult human bone marrow. Stem Cell Rep. 3 (6), 965–974. doi:10.1016/j.stemcr.2014.09.018
Li, H., Lim, H. C., Zacharaki, D., Xian, X., Kenswil, K. J. G., Braunig, S., et al. (2020). Early growth response 1 regulates hematopoietic support and proliferation in human primary bone marrow stromal cells. Haematologica 105 (5), 1206–1215. doi:10.3324/haematol.2019.216648
Li, Z., Ji, B. W., Dixit, P. D., Tchourine, K., Lien, E. C., Hosios, A. M., et al. (2022). Cancer cells depend on environmental lipids for proliferation when electron acceptors are limited. Nat. Metab. 4 (6), 711–723. doi:10.1038/s42255-022-00588-8
Liang, R., Arif, T., Kalmykova, S., Kasianov, A., Lin, M., Menon, V., et al. (2020). Restraining lysosomal activity preserves hematopoietic stem cell quiescence and potency. Cell Stem Cell 26 (3), 359–376. doi:10.1016/j.stem.2020.01.013
Liccardo, F., Śniegocka, M., Tito, C., Iaiza, A., Ottone, T., Divona, M., et al. (2023). Retinoic acid and proteotoxic stress induce AML cell death overcoming stromal cell protection. J. Exp. Clin. Cancer Res. 42 (1), 223. doi:10.1186/s13046-023-02793-z
Lin, T., Yang, Y., and Chen, X. (2023). A review of the application of mesenchymal stem cells in the field of hematopoietic stem cell transplantation. Eur. J. Med. Res. 28 (1), 268. doi:10.1186/s40001-023-01244-x
Lisi-Vega, L. E., and Méndez-Ferrer, S. (2023). The inflamed niche: a double-edged sword in AML? Blood Cancer Discov. 4 (5), 349–351. doi:10.1158/2643-3230.BCD-23-0125
Liu, X., Zhang, Y., Li, W., and Zhou, X. (2022). Lactylation, an emerging hallmark of metabolic reprogramming: current progress and open challenges. Front. Cell Dev. Biol. 10, 972020. doi:10.3389/fcell.2022.972020
Liu, X., Zheng, H., Yu, W. M., Cooper, T. M., Bunting, K. D., and Qu, C. K. (2015). Maintenance of mouse hematopoietic stem cells ex vivo by reprogramming cellular metabolism. Blood 125 (10), 1562–1565. doi:10.1182/blood-2014-04-568949
Lou, Q., Jiang, K., Xu, Q., Yuan, L., Xie, S., Pan, Y., et al. (2022). The RIG-I-NRF2 axis regulates the mesenchymal stromal niche for bone marrow transplantation. Blood 139 (21), 3204–3221. doi:10.1182/blood.2021013048
Lu, W., Wang, W., Wang, S., Feng, Y., and Liu, K. (2016). Rosiglitazone promotes bone marrow adipogenesis to impair myelopoiesis under stress. PLoS One 11 (2), e0149543. doi:10.1371/journal.pone.0149543
Luchsinger, L. L., de Almeida, M. J., Corrigan, D. J., Mumau, M., and Snoeck, H. W. (2016). Mitofusin 2 maintains haematopoietic stem cells with extensive lymphoid potential. Nature 529 (7587), 528–531. doi:10.1038/nature16500
Luk, F., de Witte, S. F., Korevaar, S. S., Roemeling-van Rhijn, M., Franquesa, M., Strini, T., et al. (2016). Inactivated mesenchymal stem cells maintain immunomodulatory capacity. Stem Cells Dev. 25 (18), 1342–1354. doi:10.1089/scd.2016.0068
Lungu, O., Toscani, D., Burroughs-Garcia, J., and Giuliani, N. (2023). The metabolic features of osteoblasts: implications for multiple myeloma (MM) bone disease. Int. J. Mol. Sci. 24 (5), 4893. doi:10.3390/ijms24054893
Lv, F. J., Tuan, R. S., Cheung, K. M., and Leung, V. Y. (2014). Concise review: the surface markers and identity of human mesenchymal stem cells. Stem Cells 32 (6), 1408–1419. doi:10.1002/stem.1681
Macías-Sánchez, M. d. M., Morata-Tarifa, C., Cuende, N., Cardesa-Gil, A., Cuesta-Casas, M. Á., Pascual-Cascon, M. J., et al. (2022). Mesenchymal stromal cells for treating steroid-resistant acute and chronic graft versus host disease: a multicenter compassionate use experience. Stem Cells Transl. Med. 11 (4), 343–355. doi:10.1093/stcltm/szac003
Maitra, B., Szekely, E., Gjini, K., Laughlin, M. J., Dennis, J., Haynesworth, S. E., et al. (2004). Human mesenchymal stem cells support unrelated donor hematopoietic stem cells and suppress T-cell activation. Bone Marrow Transpl. 33 (6), 597–604. doi:10.1038/sj.bmt.1704400
Mao, C., Liu, X., Zhang, Y., Lei, G., Yan, Y., Lee, H., et al. (2021). DHODH-mediated ferroptosis defence is a targetable vulnerability in cancer. Nature 593 (7860), 586–590. doi:10.1038/s41586-021-03539-7
Marlein, C. R., Piddock, R. E., Mistry, J. J., Zaitseva, L., Hellmich, C., Horton, R. H., et al. (2019). CD38-Driven mitochondrial trafficking promotes bioenergetic plasticity in multiple myeloma. Cancer Res. 79 (9), 2285–2297. doi:10.1158/0008-5472.CAN-18-0773
Marlein, C. R., Zaitseva, L., Piddock, R. E., Raso-Barnett, L., Scott, M. A., Ingham, C. J., et al. (2018). PGC-1α driven mitochondrial biogenesis in stromal cells underpins mitochondrial trafficking to leukemic blasts. Leukemia 32 (9), 2073–2077. doi:10.1038/s41375-018-0221-y
Marlein, C. R., Zaitseva, L., Piddock, R. E., Robinson, S. D., Edwards, D. R., Shafat, M. S., et al. (2017). NADPH oxidase-2 derived superoxide drives mitochondrial transfer from bone marrow stromal cells to leukemic blasts. Blood 130 (14), 1649–1660. doi:10.1182/blood-2017-03-772939
Maryanovich, M., Zaltsman, Y., Ruggiero, A., Goldman, A., Shachnai, L., Zaidman, S. L., et al. (2015). An MTCH2 pathway repressing mitochondria metabolism regulates haematopoietic stem cell fate. Nat. Commun. 6, 7901. doi:10.1038/ncomms8901
Mattiucci, D., Maurizi, G., Izzi, V., Cenci, L., Ciarlantini, M., Mancini, S., et al. (2018). Bone marrow adipocytes support hematopoietic stem cell survival. J. Cell Physiol. 233 (2), 1500–1511. doi:10.1002/jcp.26037
Matula, Z., Mikala, G., Lukácsi, S., Matkó, J., Kovács, T., Monostori, É., et al. (2021). Stromal cells serve drug resistance for multiple myeloma via mitochondrial transfer: a study on primary myeloma and stromal cells. Cancers 13 (14), 3461. doi:10.3390/cancers13143461
Mehta, R. S., Saliba, R. M., Cao, K., Kaur, I., Rezvani, K., Chen, J., et al. (2017). Ex vivo mesenchymal precursor cell-expanded cord blood transplantation after reduced-intensity conditioning regimens improves time to neutrophil recovery. Biol. Blood Marrow Transpl. 23 (8), 1359–1366. doi:10.1016/j.bbmt.2017.05.002
Mendelson, A., and Frenette, P. S. (2014). Hematopoietic stem cell niche maintenance during homeostasis and regeneration. Nat. Med. 20 (8), 833–846. doi:10.1038/nm.3647
Mendez-Ferrer, S., Michurina, T. V., Ferraro, F., Mazloom, A. R., Macarthur, B. D., Lira, S. A., et al. (2010). Mesenchymal and haematopoietic stem cells form a unique bone marrow niche. Nature 466 (7308), 829–834. doi:10.1038/nature09262
Mistry, J. J., Hellmich, C., Moore, J. A., Marlein, C. R., Pillinger, G., Collins, A., et al. (2019). Daratumumab inhibits AML metabolic capacity and tumor growth through inhibition of CD38 mediated mitochondrial transfer from bone marrow stromal cells to blasts in the leukemic microenvironment. Blood 134 (Suppl. ment_1), 1385. doi:10.1182/blood-2019-128592
Mistry, J. J., Marlein, C. R., Moore, J. A., Hellmich, C., Wojtowicz, E. E., Smith, J. G. W., et al. (2019). ROS-mediated PI3K activation drives mitochondrial transfer from stromal cells to hematopoietic stem cells in response to infection. Proc. Natl. Acad. Sci. U. S. A. 116, 24610–24619. doi:10.1073/pnas.1913278116
Moore, J. A., Mistry, J. J., Horton, R. H., Hellmich, C., Pattinson, A., Morfakis, A., et al. (2019). Acute myeloid leukemia export mitochondria in extracellular vesicles which induces pro-tumoral changes in bone marrow macrophages. Blood 134 (Suppl. ment_1), 1427. doi:10.1182/blood-2019-128447
Morikawa, S., Mabuchi, Y., Kubota, Y., Nagai, Y., Niibe, K., Hiratsu, E., et al. (2009). Prospective identification, isolation, and systemic transplantation of multipotent mesenchymal stem cells in murine bone marrow. J. Exp. Med. 206 (11), 2483–2496. doi:10.1084/jem.20091046
Moschoi, R., Imbert, V., Nebout, M., Chiche, J., Mary, D., Prebet, T., et al. (2016). Protective mitochondrial transfer from bone marrow stromal cells to acute myeloid leukemic cells during chemotherapy. Blood 128 (2), 253–264. doi:10.1182/blood-2015-07-655860
Muguruma, Y., Yahata, T., Miyatake, H., Sato, T., Uno, T., Itoh, J., et al. (2006). Reconstitution of the functional human hematopoietic microenvironment derived from human mesenchymal stem cells in the murine bone marrow compartment. Blood 107 (5), 1878–1887. doi:10.1182/blood-2005-06-2211
Nakahara, F., Borger, D. K., Wei, Q., Pinho, S., Maryanovich, M., Zahalka, A. H., et al. (2019). Engineering a haematopoietic stem cell niche by revitalizing mesenchymal stromal cells. Nat. Cell Biol. 21 (5), 560–567. doi:10.1038/s41556-019-0308-3
Nakatani, T., Sugiyama, T., Omatsu, Y., Watanabe, H., Kondoh, G., and Nagasawa, T. (2023). Ebf3(+) niche-derived CXCL12 is required for the localization and maintenance of hematopoietic stem cells. Nat. Commun. 14 (1), 6402. doi:10.1038/s41467-023-42047-2
Naveiras, O., Nardi, V., Wenzel, P. L., Hauschka, P. V., Fahey, F., and Daley, G. Q. (2009). Bone-marrow adipocytes as negative regulators of the haematopoietic microenvironment. Nature 460 (7252), 259–263. doi:10.1038/nature08099
Ng, F., Boucher, S., Koh, S., Sastry, K. S., Chase, L., Lakshmipathy, U., et al. (2008). PDGF, TGF-beta, and FGF signaling is important for differentiation and growth of mesenchymal stem cells (MSCs): transcriptional profiling can identify markers and signaling pathways important in differentiation of MSCs into adipogenic, chondrogenic, and osteogenic lineages. Blood 112 (2), 295–307. doi:10.1182/blood-2007-07-103697
Ning, H., Yang, F., Jiang, M., Hu, L., Feng, K., Zhang, J., et al. (2008). The correlation between cotransplantation of mesenchymal stem cells and higher recurrence rate in hematologic malignancy patients: outcome of a pilot clinical study. Leukemia 22 (3), 593–599. doi:10.1038/sj.leu.2405090
Noort, W. A., Kruisselbrink, A. B., Kruger, M., van Bezooijen, R. L., de Paus, R. A., Heemskerk, M. H., et al. (2002). Mesenchymal stem cells promote engraftment of human umbilical cord blood-derived CD34(+) cells in NOD/SCID mice. Exp. Hematol. 30 (8), 870–878. doi:10.1016/s0301-472x(02)00820-2
Oliveira, E., Costa, E. S., Ciudad, J., Gaipa, G., Sedek, Ł., Barrena, S., et al. (2022). Bone marrow stromal cell regeneration profile in treated B-cell precursor acute lymphoblastic leukemia patients: association with MRD status and patient outcome. Cancers 14 (13), 3088. doi:10.3390/cancers14133088
Omatsu, Y., Sugiyama, T., Kohara, H., Kondoh, G., Fujii, N., Kohno, K., et al. (2010). The essential functions of adipo-osteogenic progenitors as the hematopoietic stem and progenitor cell niche. Immunity 33 (3), 387–399. doi:10.1016/j.immuni.2010.08.017
Ouzin, M., and Kogler, G. (2023). Mesenchymal stromal cells: heterogeneity and therapeutical applications. Cells 12 (16), 2039. doi:10.3390/cells12162039
Panaroni, C., Gioia, R., Lupi, A., Besio, R., Goldstein, S. A., Kreider, J., et al. (2009). In utero transplantation of adult bone marrow decreases perinatal lethality and rescues the bone phenotype in the knockin murine model for classical, dominant osteogenesis imperfecta. Blood 114 (2), 459–468. doi:10.1182/blood-2008-12-195859
Parmentier, J. H., Maggi, M., Tarasco, E., Scotti, C., Avramis, V. I., and Mittelman, S. D. (2015). Glutaminase activity determines cytotoxicity of l-asparaginases on most leukemia cell lines. Leukemia Res. 39 (7), 757–762. doi:10.1016/j.leukres.2015.04.008
Phinney, D. G., Di Giuseppe, M., Njah, J., Sala, E., Shiva, S., St Croix, C. M., et al. (2015). Mesenchymal stem cells use extracellular vesicles to outsource mitophagy and shuttle microRNAs. Nat. Commun. 6, 8472. doi:10.1038/ncomms9472
Pinho, S., and Frenette, P. S. (2019). Haematopoietic stem cell activity and interactions with the niche. Nat. Rev. Mol. Cell Biol. 20 (5), 303–320. doi:10.1038/s41580-019-0103-9
Pinho, S., Lacombe, J., Hanoun, M., Mizoguchi, T., Bruns, I., Kunisaki, Y., et al. (2013). PDGFRα and CD51 mark human nestin+ sphere-forming mesenchymal stem cells capable of hematopoietic progenitor cell expansion. J. Exp. Med. 210 (7), 1351–1367. doi:10.1084/jem.20122252
Pittenger, M. F., Mackay, A. M., Beck, S. C., Jaiswal, R. K., Douglas, R., Mosca, J. D., et al. (1999). Multilineage potential of adult human mesenchymal stem cells. Science 284 (5411), 143–147. doi:10.1126/science.284.5411.143
Plakhova, N., Panagopoulos, V., Vandyke, K., Zannettino, A. C. W., and Mrozik, K. M. (2023). Mesenchymal stromal cell senescence in haematological malignancies. Cancer Metastasis Rev. 42 (1), 277–296. doi:10.1007/s10555-022-10069-9
Poloni, A., Leoni, P., Buscemi, L., Balducci, F., Pasquini, R., Masia, M. C., et al. (2006). Engraftment capacity of mesenchymal cells following hematopoietic stem cell transplantation in patients receiving reduced-intensity conditioning regimen. Leukemia 20 (2), 329–335. doi:10.1038/sj.leu.2404018
Preda, M. B., Neculachi, C. A., Fenyo, I. M., Vacaru, A. M., Publik, M. A., Simionescu, M., et al. (2021). Short lifespan of syngeneic transplanted MSC is a consequence of in vivo apoptosis and immune cell recruitment in mice. Cell Death Dis. 12 (6), 566. doi:10.1038/s41419-021-03839-w
Qin, Y., Jiang, X., Yang, Q., Zhao, J., Zhou, Q., and Zhou, Y. (2021). The functions, methods, and mobility of mitochondrial transfer between cells. Front. Oncol. 11, 672781. doi:10.3389/fonc.2021.672781
Ringden, O., Moll, G., Gustafsson, B., and Sadeghi, B. (2022). Mesenchymal stromal cells for enhancing hematopoietic engraftment and treatment of graft-versus-host disease, hemorrhages and acute respiratory distress syndrome. Front. Immunol. 13, 839844. doi:10.3389/fimmu.2022.839844
Robinson, S. N., Ng, J., Niu, T., Yang, H., McMannis, J. D., Karandish, S., et al. (2006). Superior ex vivo cord blood expansion following co-culture with bone marrow-derived mesenchymal stem cells. Bone Marrow Transpl. 37 (4), 359–366. doi:10.1038/sj.bmt.1705258
Sacchetti, B., Funari, A., Michienzi, S., Di Cesare, S., Piersanti, S., Saggio, I., et al. (2007). Self-renewing osteoprogenitors in bone marrow sinusoids can organize a hematopoietic microenvironment. Cell 131 (2), 324–336. doi:10.1016/j.cell.2007.08.025
Saito, K., Zhang, Q., Yang, H., Yamatani, K., Ai, T., Ruvolo, V., et al. (2021). Exogenous mitochondrial transfer and endogenous mitochondrial fission facilitate AML resistance to OxPhos inhibition. Blood Adv. 5 (20), 4233–4255. doi:10.1182/bloodadvances.2020003661
Samudio, I., and Konopleva, M. (2013). Asparaginase unveils glutamine-addicted AML. Blood 122 (20), 3398–3400. doi:10.1182/blood-2013-09-526392
Saulle, E., Spinello, I., Quaranta, M. T., Pasquini, L., Pelosi, E., Iorio, E., et al. (2021). Targeting lactate metabolism by inhibiting MCT1 or MCT4 impairs leukemic cell proliferation, induces two different related death-pathways and increases chemotherapeutic sensitivity of acute myeloid leukemia cells. Front. Oncol. 10, 621458. doi:10.3389/fonc.2020.621458
Savino, A. M., Fernandes, S. I., Olivares, O., Zemlyansky, A., Cousins, A., Markert, E. K., et al. (2020). Metabolic adaptation of acute lymphoblastic leukemia to the central nervous system microenvironment is dependent on Stearoyl CoA desaturase. Nat. Cancer 1 (10), 998–1009. doi:10.1038/s43018-020-00115-2
Schajnovitz, A., Itkin, T., D'Uva, G., Kalinkovich, A., Golan, K., Ludin, A., et al. (2011). CXCL12 secretion by bone marrow stromal cells is dependent on cell contact and mediated by connexin-43 and connexin-45 gap junctions. Nat. Immunol. 12 (5), 391–398. doi:10.1038/ni.2017
Schwab, K. E., and Gargett, C. E. (2007). Co-expression of two perivascular cell markers isolates mesenchymal stem-like cells from human endometrium. Hum. Reprod. 22 (11), 2903–2911. doi:10.1093/humrep/dem265
Severe, N., Karabacak, N. M., Gustafsson, K., Baryawno, N., Courties, G., Kfoury, Y., et al. (2019). Stress-induced changes in bone marrow stromal cell populations revealed through single-cell protein expression mapping. Cell Stem Cell 25 (4), 570–583. doi:10.1016/j.stem.2019.06.003
Shafat, M. S., Oellerich, T., Mohr, S., Robinson, S. D., Edwards, D. R., Marlein, C. R., et al. (2017). Leukemic blasts program bone marrow adipocytes to generate a protumoral microenvironment. Blood 129 (10), 1320–1332. doi:10.1182/blood-2016-08-734798
Shanmughapriya, S., Langford, D., and Natarajaseenivasan, K. (2020). Inter and Intracellular mitochondrial trafficking in health and disease. Ageing Res. Rev. 62, 101128. doi:10.1016/j.arr.2020.101128
Sharma, D., Yu, Y., Shen, L., Zhang, G.-F., and Karner, C. M. (2021). SLC1A5 provides glutamine and asparagine necessary for bone development in mice. eLife 10, e71595. doi:10.7554/eLife.71595
Shen, B., Tasdogan, A., Ubellacker, J. M., Zhang, J., Nosyreva, E. D., Du, L., et al. (2021). A mechanosensitive peri-arteriolar niche for osteogenesis and lymphopoiesis. Nature 591 (7850), 438–444. doi:10.1038/s41586-021-03298-5
Sheppard, A. J., Barfield, A. M., Barton, S., and Dong, Y. (2022). Understanding reactive oxygen species in bone regeneration: a glance at potential therapeutics and bioengineering applications. Front. Bioeng. Biotechnol. 10, 836764. doi:10.3389/fbioe.2022.836764
Singh, A. K., and Cancelas, J. A. (2020). Gap junctions in the bone marrow lympho-hematopoietic stem cell niche, leukemia progression, and chemoresistance. Int. J. Mol. Sci. 21 (3), 796. doi:10.3390/ijms21030796
Singh, A. K., and Cancelas, J. A. (2021). Mitochondria transfer in bone marrow hematopoietic activity. Curr. Stem Cell Rep. 7 (1), 1–12. doi:10.1007/s40778-020-00185-z
Sládek, N. E. (2003). Human aldehyde dehydrogenases: potential pathological, pharmacological, and toxicological impact. J. Biochem. Mol. Toxicol. 17 (1), 7–23. doi:10.1002/jbt.10057
Storek, J., Geddes, M., Khan, F., Huard, B., Helg, C., Chalandon, Y., et al. (2008). Reconstitution of the immune system after hematopoietic stem cell transplantation in humans. Semin. Immunopathol. 30 (4), 425–437. doi:10.1007/s00281-008-0132-5
Sugimura, K., Kimura, T., Arakawa, H., Ohno, T., Wada, Y., Kimura, Y., et al. (1990). Elevated argininosuccinate synthetase activity in adult T leukemia cell lines. Leukemia Res. 14 (10), 931–934. doi:10.1016/0145-2126(90)90184-b
Sullivan, L. B., Luengo, A., Danai, L. V., Bush, L. N., Diehl, F. F., Hosios, A. M., et al. (2018). Aspartate is an endogenous metabolic limitation for tumour growth. Nat. Cell Biol. 20 (7), 782–788. doi:10.1038/s41556-018-0125-0
Tabe, Y., and Konopleva, M. (2023). Resistance to energy metabolism - targeted therapy of AML cells residual in the bone marrow microenvironment. Cancer Drug Resist 6 (1), 138–150. doi:10.20517/cdr.2022.133
Tabe, Y., Lorenzi, P. L., and Konopleva, M. (2019). Amino acid metabolism in hematologic malignancies and the era of targeted therapy. Blood 134 (13), 1014–1023. doi:10.1182/blood.2019001034
Tamama, K., Kawasaki, H., and Wells, A. (2010). Epidermal growth factor (EGF) treatment on multipotential stromal cells (MSCs). Possible enhancement of therapeutic potential of MSC. J. Biomed. Biotechnol. 2010, 795385. doi:10.1155/2010/795385
Taniguchi Ishikawa, E., Gonzalez-Nieto, D., Ghiaur, G., Dunn, S. K., Ficker, A. M., Murali, B., et al. (2012). Connexin-43 prevents hematopoietic stem cell senescence through transfer of reactive oxygen species to bone marrow stromal cells. Proc. Natl. Acad. Sci. U. S. A. 109 (23), 9071–9076. doi:10.1073/pnas.1120358109
Tikhonova, A. N., Dolgalev, I., Hu, H., Sivaraj, K. K., Hoxha, E., Cuesta-Dominguez, A., et al. (2019). The bone marrow microenvironment at single-cell resolution. Nature 569 (7755), 222–228. doi:10.1038/s41586-019-1104-8
Tormin, A., Li, O., Brune, J. C., Walsh, S., Schutz, B., Ehinger, M., et al. (2011). CD146 expression on primary nonhematopoietic bone marrow stem cells is correlated with in situ localization. Blood 117 (19), 5067–5077. doi:10.1182/blood-2010-08-304287
Tratwal, J., Rojas-Sutterlin, S., Bataclan, C., Blum, S., and Naveiras, O. (2021). Bone marrow adiposity and the hematopoietic niche: a historical perspective of reciprocity, heterogeneity, and lineage commitment. Best. Pract. Res. Clin. Endocrinol. Metab. 35 (4), 101564. doi:10.1016/j.beem.2021.101564
Umemoto, T., Hashimoto, M., Matsumura, T., Nakamura-Ishizu, A., and Suda, T. (2018). Ca(2+)-mitochondria axis drives cell division in hematopoietic stem cells. J. Exp. Med. 215 (8), 2097–2113. doi:10.1084/jem.20180421
Vander Heiden, M. G., Cantley, L. C., and Thompson, C. B. (2009). Understanding the Warburg effect: the metabolic requirements of cell proliferation. Science 324 (5930), 1029–1033. doi:10.1126/science.1160809
Vangapandu, H. V., Alston, B., Morse, J., Ayres, M. L., Wierda, W. G., Keating, M. J., et al. (2018). Biological and metabolic effects of IACS-010759, an OxPhos inhibitor, on chronic lymphocytic leukemia cells. Oncotarget 9 (38), 24980–24991. doi:10.18632/oncotarget.25166
Vangapandu, H. V., Ayres, M. L., Bristow, C. A., Wierda, W. G., Keating, M. J., Balakrishnan, K., et al. (2017). The stromal microenvironment modulates mitochondrial oxidative phosphorylation in chronic lymphocytic leukemia cells. Neoplasia 19 (10), 762–771. doi:10.1016/j.neo.2017.07.004
Vannini, N., Campos, V., Girotra, M., Trachsel, V., Rojas-Sutterlin, S., Tratwal, J., et al. (2019). The NAD-booster nicotinamide riboside potently stimulates hematopoiesis through increased mitochondrial clearance. Cell Stem Cell 24 (3), 405–418. doi:10.1016/j.stem.2019.02.012
Vannini, N., Girotra, M., Naveiras, O., Nikitin, G., Campos, V., Giger, S., et al. (2016). Specification of haematopoietic stem cell fate via modulation of mitochondrial activity. Nat. Commun. 7, 13125. doi:10.1038/ncomms13125
vom Stein, A. F., Rebollido-Rios, R., Lukas, A., Koch, M., von Lom, A., Reinartz, S., et al. (2023). LYN kinase programs stromal fibroblasts to facilitate leukemic survival via regulation of c-JUN and THBS1. Nat. Commun. 14 (1), 1330. doi:10.1038/s41467-023-36824-2
von Heydebrand, F., Fuchs, M., Kunz, M., Voelkl, S., Kremer, A. N., Oostendorp, R. A. J., et al. (2021). Protein kinase C-β-dependent changes in the glucose metabolism of bone marrow stromal cells of chronic lymphocytic leukemia. Stem Cells 39 (6), 819–830. doi:10.1002/stem.3352
Wang, J., Liu, X., Qiu, Y., Shi, Y., Cai, J., Wang, B., et al. (2018). Cell adhesion-mediated mitochondria transfer contributes to mesenchymal stem cell-induced chemoresistance on T cell acute lymphoblastic leukemia cells. J. Hematol. Oncol. 11 (1), 11. doi:10.1186/s13045-018-0554-z
Wang, M., Zhao, A., Li, M., and Niu, T. (2023). Amino acids in hematologic malignancies: current status and future perspective. Front. Nutr. 10, 1113228. doi:10.3389/fnut.2023.1113228
Wang, Z.-H., Peng, W.-B., Zhang, P., Yang, X.-P., and Zhou, Q. (2021). Lactate in the tumour microenvironment: from immune modulation to therapy. EBioMedicine 73, 103627. doi:10.1016/j.ebiom.2021.103627
Warburg, O., Wind, F., and Negelein, E. (1927). The metabolism of tumors in the body. J. Gen. Physiol. 8 (6), 519–530. doi:10.1085/jgp.8.6.519
Wei, Q., and Frenette, P. S. (2018). Niches for hematopoietic stem cells and their progeny. Immunity 48 (4), 632–648. doi:10.1016/j.immuni.2018.03.024
Weickert, M.-T., Hecker, J. S., Buck, M. C., Schreck, C., Rivière, J., Schiemann, M., et al. (2021). Bone marrow stromal cells from MDS and AML patients show increased adipogenic potential with reduced Delta-like-1 expression. Sci. Rep. 11 (1), 5944. doi:10.1038/s41598-021-85122-8
Wilson, A., Fu, H., Schiffrin, M., Winkler, C., Koufany, M., Jouzeau, J. Y., et al. (2018). Lack of adipocytes alters hematopoiesis in lipodystrophic mice. Front. Immunol. 9, 2573. doi:10.3389/fimmu.2018.02573
Yin, X., Hu, L., Zhang, Y., Zhu, C., Cheng, H., Xie, X., et al. (2020). PDGFB-expressing mesenchymal stem cells improve human hematopoietic stem cell engraftment in immunodeficient mice. Bone Marrow Transpl. 55 (6), 1029–1040. doi:10.1038/s41409-019-0766-z
You, R., Wang, B., Chen, P., Zheng, X., Hou, D., Wang, X., et al. (2022). Metformin sensitizes AML cells to chemotherapy through blocking mitochondrial transfer from stromal cells to AML cells. Cancer Lett. 532, 215582. doi:10.1016/j.canlet.2022.215582
Yu, H., Yin, Y., Yi, Y., Cheng, Z., Kuang, W., Li, R., et al. (2020). Targeting lactate dehydrogenase A (LDHA) exerts antileukemic effects on T-cell acute lymphoblastic leukemia. Cancer Commun. 40 (10), 501–517. doi:10.1002/cac2.12080
Yuan, B., El Dana, F., Ly, S., Yan, Y., Ruvolo, V., Shpall, E. J., et al. (2020). Bone marrow stromal cells induce an ALDH+ stem cell-like phenotype and enhance therapy resistance in AML through a TGF-β-p38-ALDH2 pathway. PLoS One 15 (11), e0242809. doi:10.1371/journal.pone.0242809
Zhang, L., Chi, Y., Wei, Y., Zhang, W., Wang, F., Zhang, L., et al. (2021). Bone marrow-derived mesenchymal stem/stromal cells in patients with acute myeloid leukemia reveal transcriptome alterations and deficiency in cellular vitality. Stem Cell Res. Ther. 12 (1), 365. doi:10.1186/s13287-021-02444-0
Zhang, P., Dong, J., Fan, X., Yong, J., Yang, M., Liu, Y., et al. (2023). Characterization of mesenchymal stem cells in human fetal bone marrow by single-cell transcriptomic and functional analysis. Signal Transduct. Target Ther. 8 (1), 126. doi:10.1038/s41392-023-01338-2
Zhang, W., Trachootham, D., Liu, J., Chen, G., Pelicano, H., Garcia-Prieto, C., et al. (2012). Stromal control of cystine metabolism promotes cancer cell survival in chronic lymphocytic leukaemia. Nat. Cell Biol. 14 (3), 276–286. doi:10.1038/ncb2432
Zhang, Y., Peng, Q., Zheng, J., Yang, Y., Zhang, X., Ma, A., et al. (2023). The function and mechanism of lactate and lactylation in tumor metabolism and microenvironment. Genes and Dis. 10 (5), 2029–2037. doi:10.1016/j.gendis.2022.10.006
Zhou, B. O., Yue, R., Murphy, M. M., Peyer, J. G., and Morrison, S. J. (2014). Leptin-receptor-expressing mesenchymal stromal cells represent the main source of bone formed by adult bone marrow. Cell Stem Cell 15 (2), 154–168. doi:10.1016/j.stem.2014.06.008
Zhou, T., Yang, Y., Chen, Q., and Xie, L. (2019). Glutamine metabolism is essential for stemness of bone marrow mesenchymal stem cells and bone homeostasis. Stem Cells Int. 2019, 8928934. doi:10.1155/2019/8928934
Zhu, R. J., Wu, M. Q., Li, Z. J., Zhang, Y., and Liu, K. Y. (2013). Hematopoietic recovery following chemotherapy is improved by BADGE-induced inhibition of adipogenesis. Int. J. Hematol. 97 (1), 58–72. doi:10.1007/s12185-012-1233-4
Keywords: bone marrow, mesenchymal stem cells, adipocytes, hematopoiesis, hematological malignancies, metabolism, mitochondrial transfer
Citation: Singh AK, Prasad P and Cancelas JA (2023) Mesenchymal stromal cells, metabolism, and mitochondrial transfer in bone marrow normal and malignant hematopoiesis. Front. Cell Dev. Biol. 11:1325291. doi: 10.3389/fcell.2023.1325291
Received: 21 October 2023; Accepted: 23 November 2023;
Published: 18 December 2023.
Edited by:
Pamela L. Wenzel, University of Texas Health Science Center at Houston, United StatesReviewed by:
Graca Almeida-Porada, Wake Forest University, United StatesSimon Mendez-Ferrer, University of Cambridge, United Kingdom
Copyright © 2023 Singh, Prasad and Cancelas. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jose A. Cancelas, jose.cancelas@cchmc.org, jose.cancelas@uc.edu