Mechanosensory feedback loops during chronic inflammation
 Sarbari Saha
Sarbari Saha Dafne Müller
Dafne Müller Andrew G. Clark
Andrew G. Clark- 1University of Stuttgart, Institute of Cell Biology and Immunology, Stuttgart, Germany
- 2University of Stuttgart, Stuttgart Research Center Systems Biology, Stuttgart, Germany
- 3University of Tübingen, Center for Personalized Medicine, Tübingen, Germany
Epithelial tissues are crucial to maintaining healthy organization and compartmentalization in various organs and act as a first line of defense against infection in barrier organs such as the skin, lungs and intestine. Disruption or injury to these barriers can lead to infiltration of resident or foreign microbes, initiating local inflammation. One often overlooked aspect of this response is local changes in tissue mechanics during inflammation. In this mini-review, we summarize known molecular mechanisms linking disruption of epithelial barrier function to mechanical changes in epithelial tissues. We consider direct mechanisms, such as changes in the secretion of extracellular matrix (ECM)-modulating enzymes by immune cells as well as indirect mechanisms including local activation of fibroblasts. We discuss how these mechanical changes can modulate local immune cell activity and inflammation and perturb epithelial homeostasis, further dysregulating epithelial barrier function. We propose that this two-way relationship between loss of barrier function and altered tissue mechanics can lead to a positive feedback loop that further perpetuates inflammation. We discuss this cycle in the context of several chronic inflammatory diseases, including inflammatory bowel disease (IBD), liver disease and cancer, and we present the modulation of tissue mechanics as a new framework for combating chronic inflammation.
Introduction
Epithelial barrier tissues maintain a tight seal between the outside environment and the inside of the body. Loss of barrier integrity leads to local activation of immune cells and fibroblasts, which can remodel local ECM networks, the major determinants of tissue mechanics. Over time, these structural and molecular changes result in tissue stiffening (Barron and Wynn, 2011; Chrysanthopoulou et al, 2014; Curaj et al, 2020). During acute inflammation, increased tissue stiffness can be beneficial for regeneration and wound healing, for example, by enhancing immune cell activity and stimulating immune cell migration and infiltration (Sridharan et al, 2019; Gaertner et al, 2022; Millán-Salanova and Vicente-Manzanares, 2022; Nalkurthi et al, 2022). However, during chronic inflammation, modifications in local ECM networks can become permanent, leading to irreversible stiffening of the tissue and culminating in fibrosis (Jeljeli et al, 2019; Velotti et al, 2020).
Pathologically stiff tissue can promote immune cell recruitment and activation via mechanosensing pathways, leading to increased immune cell migration and differentiation and activation of fibroblasts (Chen et al, 2020; Atcha et al, 2021; Chirivì et al, 2021; Jiang et al, 2022). Increased tissue stiffness also results in epithelial cell depolarization, reduced cell-cell junctions and increased migration (Discher et al, 2005; Aparicio-Yuste et al, 2022). While in the short-term this may aid in wound resealing, epithelial cells on stiff environments are less able to maintain a tight barrier, creating a feedback loop between increased barrier permeability and inflammation mediated by changes in tissue mechanics (Figure 1). Such mechanical feedback can ultimately disrupt organ function and presents a major risk factor for cancer development. Here, we discuss the molecular mechanisms that contribute to these feedback loops as well as pathologies where such mechanical feedback can play a role in disease progression.
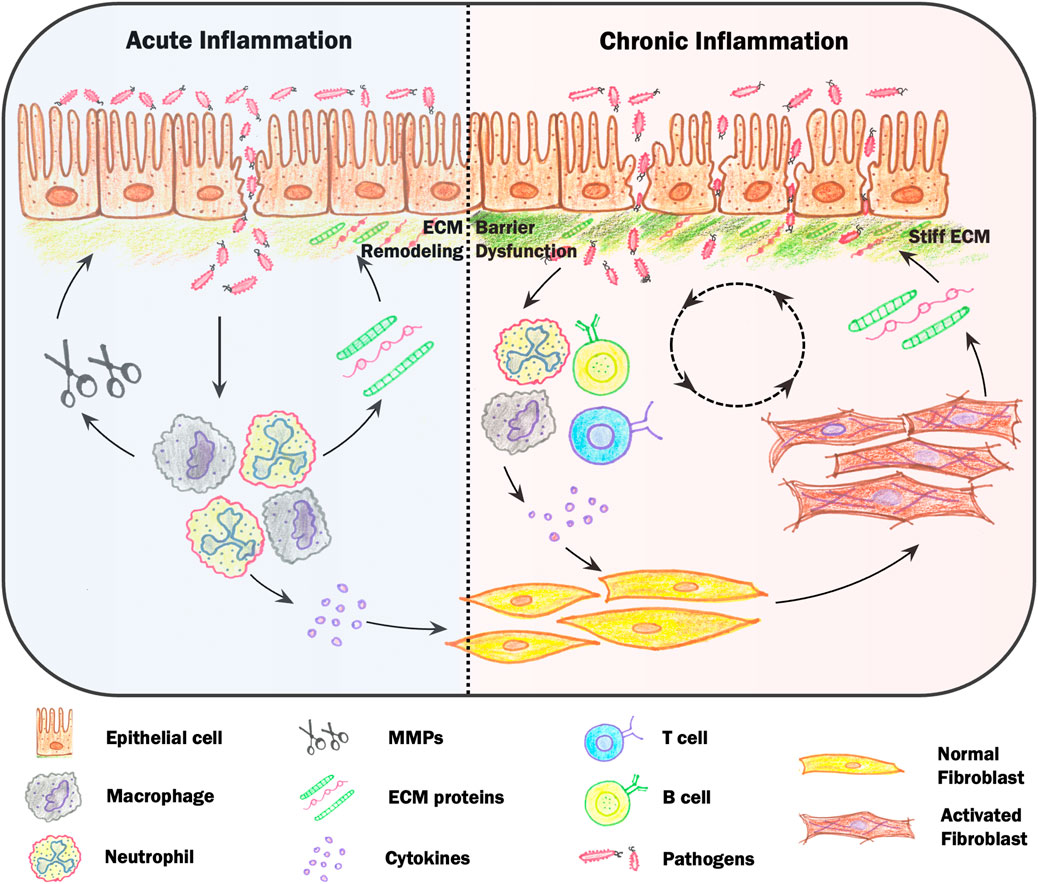
FIGURE 1. Mechanosensory feedback loops during inflammation. Loss of barrier integrity results in infiltration of microbes that initiates acute inflammation. Inflammation is led by neutrophils and macrophages, which release cytokines and chemokines and modify local extracellular matrix (ECM) structures by secretion of ECM proteins and matrix metalloproteinases (MMPs). Activated immune cells also stimulate fibroblasts, which secrete, assemble and physically remodel ECM networks, resulting in a stiffening of ECM networks. During chronic inflammation, pathologically stiff tissue can lead to over-activation of immune cells via mechanosensing pathways, resulting in increased immune cell migration and differentiation. Increases in tissue stiffness can also lead to epithelial cell depolarization, reduced cell-cell junctions and increased migration. While in the short-term this may aid in wound resealing, epithelial cells on stiff environments are less able to maintain a tight barrier, creating a positive feedback loop between increased barrier permeability and inflammation mediated by changes in tissue mechanics.
Epithelial barrier disruption leads to inflammation and local ECM remodelling
Loss of barrier integrity leads to infiltration of microbes, initiating a cascade of immune reactions whereby neutrophils and monocytes are first recruited to the site of infection (Jenne et al, 2018; Herrero-Cervera et al, 2022). These first responders not only trigger inflammation by releasing cytokines and chemokines, but also modify local extracellular matrix (ECM) structures by secretion of neutrophil elastase (NE), cathepsins, gelatinases and matrix metalloproteinases (MMPs; Delclaux et al, 1996; Ong et al, 2015; 2017; Medeiros et al, 2017). These enzymes promote the degradation of ECM components such as collagen, laminin, elastin, fibronectin and matrix bound glycoproteins (Ong et al, 2015; Xu et al, 2020). Activated neutrophils release exosomes and Neutrophil Extracellular Traps (NETs) rich in NE. NE-rich exosomes can bind to the ECM via the integrin Mac1 and degrade Collagen-I (Genschmer et al, 2019). NEs found in NETs have been shown to degrade cartilage matrix synovium, resulting in synovial joint injury (Carmona-Rivera et al, 2020). Collagenase and elastase treatment in lung tissues causes a loss and shortening of ECM fibers and decreases mechanical tissue stiffness by up to 50% (Mariano et al, 2023). In addition to degrading local ECM networks, neutrophils are also involved in tissue repair and scar formation. In response to liver injury, neutrophils physically transport existing ECM fibers to the wound site, leading to ECM accumulation at the site of damage (Fischer et al, 2022).
Similar to neutrophils, macrophages also produce and secrete various ECM-degrading enzymes (Sutherland et al, 2023). In addition, macrophages ingest and degrade ECM structures by integrin-mediated phagocytosis and receptor-mediated endocytosis (McKleroy et al, 2013; Zhao et al, 2022). On the other hand, macrophages also secrete ECM proteins including fibronectin, laminin and versican, which can help to provide a mechanical scaffold following injury and aid in the renewal of tissue architecture (Tomlin and Piccinini, 2018). Exposure to inflammatory cytokines including Transforming growth factor beta (TGF-β), Interleukin (IL)-10 and IL-13 can stimulate secretion of collagen-IV in macrophages (Schnoor et al, 2008). Differentiation of macrophages to myofibroblasts results in the production of fibrillar collagen during scar formation and ECM remodelling (Simões et al, 2020). Both macrophages and neutrophils are thus involved in degradation, production and remodeling of ECM networks and are crucial to maintaining a proper balance during homeostasis and regeneration.
When this balance is disturbed, for example, during chronic inflammation, macrophages and neutrophils can activate fibroblasts, which secrete, assemble and physically remodel ECM networks (Jeljeli et al, 2019). Culturing fibroblasts in conditioned medium from M2-like macrophages causes an increase in Col5a1 and Col6a1 production, leading to the production of thinner and more aligned collagen matrices. On the other hand, treating fibroblasts with hybrid M1/M2-conditioned medium results in the production of thicker, randomly oriented collagen networks. This suggests that shifting the phenotype of macrophages can promote architectural changes in the ECM via modulation of fibroblast activity (Witherel et al, 2021). In addition to molecular signals, physical cues from the microenvironment can also influence fibroblast-mediated ECM remodeling. When fibroblasts treated with M1/M2 conditioned medium are cultured on stiff substrates, they produce more aligned collagen networks compared to when they are cultured on softer hydrogels (Li and Bratlie, 2021). Fibroblasts also regulate their own activity via autocrine signaling. For example, during the inflammatory phase of myocardial infarction, activated fibroblasts produce and assemble fibrin and fibronectin and begin secreting TGF-β1, leading to a positive feedback loop of enhanced fibroblast differentiation, collagen synthesis and macrophage polarization. After reaching a stable state, a negative feedback loop is initiated, reducing TGF-β1 expression and resulting in completion of the mature scar (Curaj et al, 2020). Repeated injury and scarring can lead to a build-up of stiff fibrotic tissue that triggers fibroblasts to secrete more collagen, further driving the cycle of ECM deposition (Liu et al, 2010). Interestingly, a number of inflammatory conditions can also lead to tissue hypoxia, which, at least in tumors, can stimulate fibroblast-mediated collagen deposition and secretion of collagen-modifying enzymes including prolyl and lysyl hydroxylases (Gilkes et al, 2013). Together, these studies suggest that in various inflammatory conditions, activation of immune cells and fibroblasts leads to the reorganization of local ECM structures. During chronic inflammation, this results in a build-up of ECM and stiffening of the tissue, which can in turn stimulate immune cell activity via various mechanosensitive pathways.
Immune cell activation by mechanosensing pathways
The innate immune system forms the first line of defense against pathogens entering the body. Leucocytes involved in the innate immune response, or myeloid cells, including macrophages, dendritic cells and mast cells, are adherent and contact-dependent, making them sensitive to changes in tissue mechanics. In particular, increased substrate stiffness, which is a result of long-term chronic inflammation, leads to increased immune cell activation and secretion of inflammatory cytokines. Lipopolysaccharide (LPS)-Activated macrophages and bone-marrow derived dendritic cells (DCs) both display enhanced production of inflammatory cytokines when cultured on mechanically stiff substrates as compared to soft hydrogels (Meli et al, 2023). DCs cultured on stiff substrates also show increased expression of glucose metabolism genes and an overall increase in their glycolytic rate, suggesting that DCs are more metabolically active on stiff substrates (Chakraborty et al, 2021). Mast cells, which are implicated in pulmonary fibrosis, are also mechanosensitive. Reseeding of healthy mast cells onto decellularized fibrotic lung tissue leads to increased degranulation and secretion of histamine and TGF-β1 compared to mast cells reseeded on healthy decellularized lung. Mechanical stretching of mast cells can produce a similar phenotype, further implicating mechanosensing in this response (Shimbori et al, 2019). The regulation of immune cell activity by increased substrate stiffness and mechanical stress is mediated by various mechanosensitive pathways including Yes-associated protein 1 (YAP) and Transcriptional coactivator with PDZ-binding motif (TAZ). High substrate stiffness leads to increased stress on the nuclear envelope, resulting in the accumulation of nuclear YAP and activation of downstream targets (Elosegui-Artola et al, 2017). In addition to YAP/TAZ signaling, stretch-activated ion channels such as piezo type mechanosensitive ion channel component 1 (PIEZO1) and Transient Receptor Potential Cation Channel Subfamily V Member 4 (TRPV4) are also involved in mechanosensing responses (reviewed in Du et al, 2022). Together, these studies suggest that immune cells involved in the innate immune response are mechanosensitive and display pro-inflammatory phenotypes in response to increased mechanical stiffness.
Cells involved in the adaptive immune response are also mechanosensitive. In order to carry out their effector functions, naïve B cells and T cells must first be activated, or “primed,” by antigen presenting cells (APCs) such as DCs. Increased stiffness of substrates designed to mimic the APC cell surface has been shown to facilitate the activation of B cells, T cells and Natural Killer (NK) cells (Judokusumo et al, 2012; Comrie et al, 2015; Meng et al, 2020). Similarly, increased stiffness of the actomyosin cortex of antigen presenting DCs enhances T cell activation (Blumenthal et al, 2020). Experiments using optical tweezers or fluid flow have demonstrated that direct application of mechanical force on T cell receptors (TCRs) can induce T cell activation (Kim et al, 2009; Li et al, 2010). Although adaptive immune cell activation is clearly mechanosensitive, it is not clear how tissue stiffness influences adaptive immune cell activity. Furthermore, B cell and T cell priming typically occurs in lymph nodes, not in the inflamed tissue. The relationship between tissue stiffness and adaptive immune priming therefore remains an open question. However, recent studies have suggested that T cell migration, along with the migration of DCs and mast cells is increased on stiff environments (Meng et al, 2020; Yu et al, 2021). This suggests that increased tissue stiffness may enhance local immune activity by stimulating both innate and adaptive immune cell migration. Increased mechanical stiffness during inflammation not only affects immune cell activity but can also have an impact on epithelial barrier integrity by directly regulating epithelial cells.
Modulation of epithelial cell behavior by mechanical cues
The maintenance of epithelial barrier integrity is most commonly associated with tight junctions (TJs), which provide a tight seal at cell-cell boundaries and prevent the passage of materials across the epithelial layer. Recent work also suggests that adherens junctions (AJs) play a major role in epithelial integrity, either directly through mechanosensing pathways or by mediating TJ stability (Yap et al, 2018). A number of studies have demonstrated that both AJs and TJs are mechanosensitive in response to in-plane stresses arising from actomyosin contraction or external stretch, whereby moderate amounts of tensile stress led to junction reinforcement, while very high stresses cause epithelial tearing and rupture (Spadaro et al, 2017; Acharya et al, 2018; Schwayer et al, 2019). In addition to in-plane stresses, mechanosensing at cell-substrate adhesions can also affect cell-cell junction integrity. The balance between cell-cell and cell-substrate adhesions has been described as an “active wetting” phenomenon (Gonzalez-Rodriguez et al, 2012; Beaune et al, 2014; Pérez-González et al, 2019). For surfaces where cell-substrate adhesion is low, for example, very soft substrates, cell-cell adhesions dominate, leading to rounding and aggregation. This is analogous to water droplet formation on a hydrophobic surface, where liquid-substrate interactions are unfavorable and the surface tension of the droplet dominates. On substrates where cell-substrate adhesions are high, for example, on very stiff substrates, cell-substrate adhesions dominate, causing the multicellular structure to spread, or “wet” (Gonzalez-Rodriguez et al, 2012). Softer substrates therefore favor stable junctions and a tight barrier, whereas a stiff substrate favors more loosely attached cells and can also lead to dispersal into individual cells (Gonzalez-Rodriguez et al, 2012; Pérez-González et al, 2019; Ilina et al, 2020). In addition to mechanical wetting/dewetting resulting from the balance between cell-cell and cell-substrate adhesions, molecular cross-talk between different adhesion structures has also been shown to regulate cell-cell junction integrity in a substrate stiffness-dependent manner (Haas et al, 2020).
In addition to stabilization of junction proteins, efficient wound healing is a crucial aspect of tissue barrier maintenance. Wound healing requires cell migration to rapidly infiltrate the wound and actomyosin contraction to reseal the damaged area (Martin and Leibovich, 2005; Rodrigues et al, 2019). Higher substrate stiffness leads to faster wound closure mediated by increased collective migration speed and more coordinated cell movements. On stiffer substrates, actomyosin contraction slows down due to increased drag from the substrate, while crawling migration is independent of the substrate mechanics (Staddon et al, 2018; Ajeti et al, 2019). Other reports have suggested that higher stiffness can increase collective migration speeds and correlation in wound healing assays (Ng et al, 2012). It is likely that the dependence on stiffness is biphasic and highly cell-type dependent. In addition to elastic stiffness of tissues and cellular substrates, viscoelastic properties of ECM networks also influence coordinated cell movements. Crosslinking of collagen networks leads to increased network stiffness and reduces viscoelasticity, resulting in reduced collective migration (Murrell et al, 2011; Clark et al, 2022). Interestingly, changes in tissue viscoelasticity have also recently been shown to regulate collective cell behavior during development and cell invasion (Barriga and Mayor, 2019; Elosegui-Artola, 2021; Elosegui-Artola et al, 2023). Along with cell rearrangements to seal the wound, increased cell division is required to repopulate the wounded area. Substrate mechanics also regulates this process by modulating in-plane stresses generated during the resealing response, which can stimulate cell division (Zhang et al, 2003; Gudipaty et al, 2017; Donker et al, 2022). The mechanisms underlying the regulation of epithelial cell division and turnover in response to in-plane forces has been studied in several contexts (reviewed in Ragkousi and Gibson, 2014). Taken together, these studies indicate that increased substrate stiffness can perturb cell-cell junctions and cell polarity and impair the wound healing response. This suggests that the mechanical changes induced during inflammation can feed back onto epithelial cell function, resulting in further loss of barrier integrity.
Mechanosensing feedback loops in chronic inflammatory diseases
Mechanical feedback loops are likely to play a role in a number of chronic inflammatory diseases including IBD, liver disease and cancer. IBD is characterized by a cycle of increased intestinal barrier permeability and inflammation. Both immune cells and fibroblasts participate in ECM deposition and reorganization in IBD, leading to the onset of pathological tissue stiffening (Wang et al, 2022). Once tissue stiffening has begun, additional feedback mechanisms drive further tissue stiffening, leading to fibrosis and stricture formation (Figure 2A). During intestinal fibrosis, mast cell infiltration and degranulation leads to the release of large amounts of tryptase through the PAR-2/Akt/mTOR pathway, which converts fibroblasts into activated myofibroblasts. This results in deposition of collagen and fibronectin to promote intestinal fibrosis (Liu et al, 2021). Other recent work has suggested that ubiquitin-specific protease 2 (USP2), which is upregulated in intestinal myeloid cells during IBD and mouse models of colitis, increases the expression of collagen and alpha smooth muscle actin (αSMA), leading to further ECM remodeling and tissue stiffening (An et al, 2022). Collagen-I deposition in the intestine also activates the YAP/TAZ pathway in epithelial cells through Fak/Src signaling to initiate a regenerative cascade to induce a fetal-like state in the colonic epithelium, where cells become more motile and prone to reorganization compared to homeostatic conditions (Yui et al, 2018). Downstream effects of YAP/TAZ also induce the secretion of IL-33 and IL-18 and lead to cytoskeletal re-organization (Kobayashi et al, 2022). Together, these studies suggest that mechanical reorganization of ECM networks during IBD can drive further tissue stiffening, prolonged inflammation and reduced barrier function.
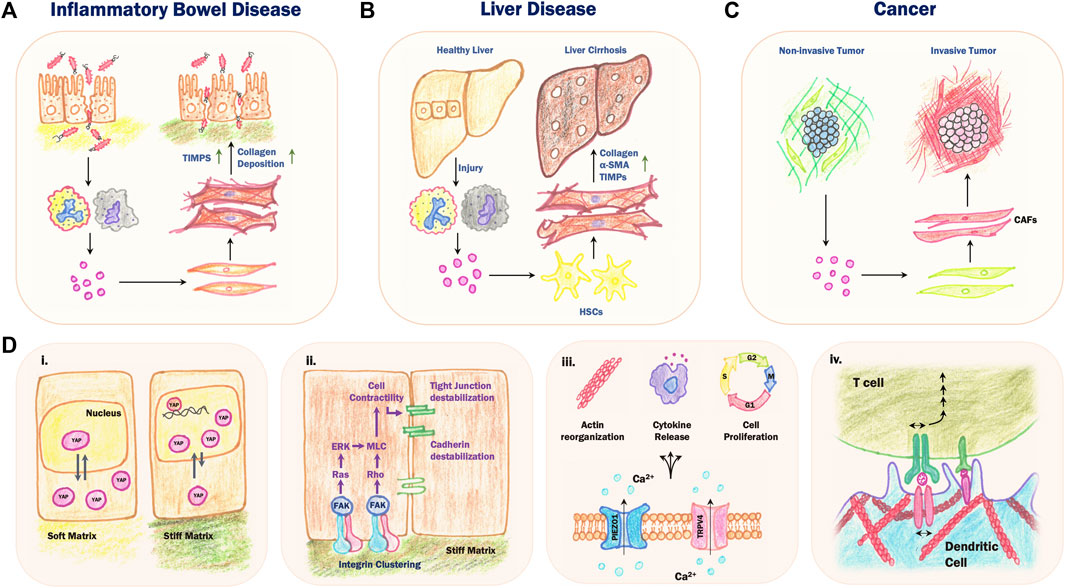
FIGURE 2. Mechanosensory feedback loops in chronic inflammatory diseases. (A) During Inflammatory Bowel Disease (IBD), a cycle of reduced intestinal barrier function and chronic inflammation results in increased collagen deposition and secretion of tissue inhibitors of metalloproteinases (TIMPs) by local activated fibroblasts, leading to stiffening of the underlying ECM and tissue fibrosis. (B) Persistent injury and inflammation in the liver results in the differentiation of hepatic stellate cells (HSCs) into activated fibroblast-like cells which secrete collagen and TIMPs and express α-SMA. This results in a replacement of the normal liver parenchyma with fibrotic scar tissue, eventually leading to fibrosis and cirrhosis. (C) The cross-talk between cancer cells and stromal cells leads to the activation of cancer-associated fibroblasts (CAFs), which secrete ECM proteins and matrix remodelling enzymes that contribute to increased stromal stiffness and invasion and metastasis. (D) Mechanochemical feedback during chronic inflammation involves various mechanosensing pathways. (i) Increased substrate stiffness results in mechanical stress on the nucleus, which inhibits nuclear export of yes-associated protein (YAP). In the nucleus, YAP acts as transcriptional coactivator to increase expression of downstream genes involved in cell proliferation and migration. (ii) High substrate stiffness leads to increased integrin clustering, which activates downstream signal transduction pathways leading to destabilization of cell-cell junctions. (iii) Mechanical stretching of the plasma membrane opens stretch-activated channels including PIEZO1 and TRPV4, leading to an influx of Ca2+ ions and several downstream effects including changes in actin dynamics, cytokine release and cell proliferation. (iv) Activation, or “priming”, of B cells and T cells involves heterotypic binding of membrane receptors between the B or T cell and an antigen presenting cell such as a dendritic cell. Increased stiffness of the actomyosin cortex in the dendritic cell limits mobility of the membrane receptors, resulting in increased mechanical stress on the B or T cell receptor, which enhances the activation process.
The liver is also exposed to various external stresses arising from dietary factors, exposure to gut microbe metabolites and alcohol and drug use, leading to tissue damage and inflammation (Lang and Schnabl, 2020; Yahoo et al, 2023). In the case of persistent inflammation, increased accumulation of ECM results in the replacement of healthy liver parenchyma with fibrotic scar tissue, further driving mechanosensitive feedback pathways (Figure 2B; Dhar et al, 2020). In mouse models of liver fibrosis, excess deposition of collagen and fibronectin along with accumulation of αSMA-expressing myofibroblasts leads to cirrhosis and increased expression of ECM genes, which correlates with poor patient prognosis (Wu et al, 2021). During this process, hepatic stellate cells (HSCs) transdifferentiate into fibroblast-like cells that express αSMA and secrete ECM components such as collagen-I and-III, fibronectin and laminin, contributing to the development of fibrosis (Friedman, 2008). HSCs also produce MMPs and Tissue Inhibitors of Metalloproteinases (TIMPs) which are the major drivers of ECM remodelling during hepatic fibrosis (Duarte et al, 2015). Chronic overexpression of TIMPs prevents normal collagen remodeling, leading to an increased collagen build-up that drives liver fibrosis (Benyon and Arthur, 2021). The resulting altered biomechanical environment can also drive liver tumorigenesis by activation of integrin-β1 and focal adhesion kinase, leading to increased cell proliferation (Schrader et al, 2011).
Chronic inflammation is a risk factor for tumorigenesis and cancer not only in the liver, but also in other tissues. The evolution of the tumor microenvironment shares many similarities with chronic inflammation, and tumors have been notably characterized as “wounds that never heal” (Dvorak, 1986; Hua and Bergers, 2019). One prominent feature of tumor progression is the cross-talk between tumor cell behavior and the increased stiffening of connective tissue surrounding the tumor (the “stroma”; Figure 2C). High stromal stiffness can lead to increased cytoskeletal activity and migration, reduced polarity and epithelial-mesenchymal transition (EMT; Clark and Vignjevic, 2015). Changes in stromal network architecture and mechanics are mediated primarily by cancer-associated fibroblasts (CAFs), which share many common features with activated fibroblasts during chronic inflammation. CAFs display increased secretion of cytokines, growth factors and matrix remodeling enzymes as well as increased mechanical force production (Sahai et al, 2020). Together, these factors drive changes in ECM organization that contribute to increased stromal stiffness, tumor invasion and metastasis. In addition, CAFs secrete proteases that cleave and activate ECM-bound cytokines and cell adhesion molecules, promoting increased migration of cancer cells and EMT (Fiori et al, 2019). The mechanical properties of the tumor stroma are also thought to contribute to immune escape mechanisms during cancer and could interfere with cancer immunotherapy (Denton et al, 2018; Ollauri-Ibáñez et al, 2021). Together, these studies suggest that similar to chronic inflammatory diseases, mechanosensory feedback loops can drive local tissue stiffening and cancer progression.
Conclusion and outlook
Disruption of epithelial barrier tissues leads to local inflammation and activation of immune cells and fibroblasts that modify local ECM structures. Repeated injury or chronic inflammation can lead to permanent ECM remodeling and tissue stiffening, which can further exacerbate inflammation, excess fibroblast activity and barrier disruption via various mechanosensing pathways (Figure 2D). Altered tissue mechanics represents a common and general feature of chronic inflammatory diseases, despite differences in the molecular profiles of these pathologies. Future translational studies aimed at modulating tissue mechanics therefore have the potential to identify exciting new therapeutic approaches with broad applications from chronic inflammation to cancer.
Author contributions
All authors listed have made a substantial, direct, and intellectual contribution to the work and approved it for publication.
Funding
AGC and SS are funded by the Federal Ministry of Education and Research (BMBF) and the Baden-Württemberg Ministry of Science (MWK BW) as part of the Excellence Strategy of the German Federal and State Governments (NWG-GastroTumors to AGC). DM is supported by the German Cancer Aid (Grant 70114233). This work was also supported by the Biomedical Systems Seed Funding from the University of Stuttgart.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Acharya, B. R., Nestor-Bergmann, A., Liang, X., Gupta, S., Duszyc, K., Gauquelin, E., et al. (2018). A Mechanosensitive RhoA Pathway that Protects Epithelia against Acute Tensile Stress. Dev. Cell 47, 439–452.e6. doi:10.1016/j.devcel.2018.09.016
Ajeti, V., Tabatabai, A. P., Fleszar, A. J., Staddon, M. F., Seara, D. S., Suarez, C., et al. (2019). Wound Healing Coordinates Actin Architectures to Regulate Mechanical Work. Nat. Phys. 15, 696–705. doi:10.1038/S41567-019-0485-9
An, R., Wang, P., Guo, H., Liuyu, T., Zhong, B., and Zhang, Z.-D. (2022). USP2 promotes experimental colitis and bacterial infections by inhibiting the proliferation of myeloid cells and remodeling the extracellular matrix network. Cell Insight 1, 100047. doi:10.1016/j.cellin.2022.100047
Aparicio-Yuste, R., Muenkel, M., Clark, A. G., Gómez-Benito, M. J., and Bastounis, E. E. (2022). A Stiff Extracellular Matrix Favors the Mechanical Cell Competition that Leads to Extrusion of Bacterially-Infected Epithelial Cells. Front. Cell Dev. Biol. 10, 912318. doi:10.3389/fcell.2022.912318
Atcha, H., Jairaman, A., Holt, J. R., Meli, V. S., Nagalla, R. R., Veerasubramanian, P. K., et al. (2021). Mechanically activated ion channel Piezo1 modulates macrophage polarization and stiffness sensing. Nat. Commun. 12, 3256. doi:10.1038/s41467-021-23482-5
Barriga, E. H., and Mayor, R. (2019). Adjustable viscoelasticity allows for efficient collective cell migration. Semin. Cell Dev. Biol. 93, 55–68. doi:10.1016/j.semcdb.2018.05.027
Barron, L., and Wynn, T. A. (2011). Fibrosis is regulated by Th2 and Th17 responses and by dynamic interactions between fibroblasts and macrophages. Am. J. Physiol. Gastrointest. Liver Physiol. 300, 723–728. doi:10.1152/ajpgi.00414.2010
Beaune, G., Stirbat, T. V., Khalifat, N., Cochet-Escartin, O., Garcia, S., Gurchenkov, V. V., et al. (2014). How cells flow in the spreading of cellular aggregates. Proc. Natl. Acad. Sci. 111, 8055–8060. doi:10.1073/pnas.1323788111
Benyon, R. C., and Arthur, M. J. P. (2001). Extracellular Matrix Degradation and the Role of Hepatic Stellate Cells. Semin. Liver Dis. 21 (3), 373–384. doi:10.1055/s-2001-17552
Blumenthal, D., Chandra, V., Avery, L., and Burkhardt, J. K. (2020). Mouse T cell priming is enhanced by maturation-dependent stiffening of the dendritic cell cortex. Elife 9, e55995. doi:10.7554/eLife.55995
Carmona-Rivera, C., Carlucci, P. M., Goel, R. R., James, E., Brooks, S. R., Rims, C., et al. (2020). Neutrophil extracellular traps mediate articular cartilage damage and enhance cartilage component immunogenicity in rheumatoid arthritis. JCI Insight 5, e139388. doi:10.1172/jci.insight.139388
Chakraborty, M., Chu, K., Shrestha, A., Revelo, X. S., Zhang, X., Gold, M. J., et al. (2021). Mechanical Stiffness Controls Dendritic Cell Metabolism and Function. Cell Rep. 34, 108609. doi:10.1016/j.celrep.2020.108609
Chen, M., Zhang, Y., Zhou, P., Liu, X., Zhao, H., Zhou, X., et al. (2020). Substrate stiffness modulates bone marrow-derived macrophage polarization through NF-κB signaling pathway. Bioact. Mater 5, 880–890. doi:10.1016/j.bioactmat.2020.05.004
Chirivì, M., Maiullari, F., Milan, M., Presutti, D., Cordiglieri, C., Crosti, M., et al. (2021). Tumor extracellular matrix stiffness promptly modulates the phenotype and gene expression of infiltrating T lymphocytes. Int. J. Mol. Sci. 22, 5862. doi:10.3390/ijms22115862
Chrysanthopoulou, A., Mitroulis, I., Apostolidou, E., Arelaki, S., Mikroulis, D., Konstantinidis, T., et al. (2014). Neutrophil extracellular traps promote differentiation and function of fibroblasts. J. Pathol. 233, 294–307. doi:10.1002/path.4359
Clark, A. G., Maitra, A., Jacques, C., Bergert, M., Pérez-González, C., Simon, A., et al. (2022). Self-generated gradients steer collective migration on viscoelastic collagen networks. Nat. Mater 21 (10), 1200–1210. doi:10.1038/s41563-022-01259-5
Clark, A. G., and Vignjevic, D. M. (2015). Modes of cancer cell invasion and the role of the microenvironment. Curr. Opin. Cell Biol. 36, 13–22. doi:10.1016/j.ceb.2015.06.004
Comrie, W. A., Li, S., Boyle, S., and Burkhardt, J. K. (2015). The dendritic cell cytoskeleton promotes T cell adhesion and activation by constraining ICAM-1 mobility. J. Cell Biol. 208, 457–473. doi:10.1083/jcb.201406120
Curaj, A., Schumacher, D., Rusu, M., Staudt, M., Li, X., Simsekyilmaz, S., et al. (2020). Neutrophils modulate fibroblast function and promote healing and scar formation after murine myocardial infarction. Int. J. Mol. Sci. 21 (10), 3685. doi:10.3390/ijms21103685
Delclaux, C., Delacourt, C., D’Ortho, M. P., Boyer, V., Lafuma, C., and Harf, A. (1996). Role of Gelatinase B and Elastase in Human Polymorphonuclear Neutrophil Migration across Basement Membrane. Am. J. Respir. Cell Mol. Biol. 14, 288–295. doi:10.1165/ajrcmb/.14.3.8845180
Denton, A. E., Roberts, E. W., and Fearon, D. T. (2018). “Stromal Cells in the Tumor Microenvironment,” in Stromal Immunology. Editors B. M. J. Owens, and M. A. Lakins (Cham: Springer International Publishing), 99–114. doi:10.1007/978-3-319-78127-3_6
Dhar, D., Baglieri, J., Kisseleva, T., and Brenner, D. A. (2020). Mechanisms of liver fibrosis and its role in liver cancer. Exp. Biol. Med. 245, 96–108. doi:10.1177/1535370219898141
Discher, D. E., Janmey, P., and Wang, Y.-L. (2005). Tissue cells feel and respond to the stiffness of their substrate. Science 310, 1139–1143. doi:10.1126/science.1116995
Donker, L., Houtekamer, R., Vliem, M., Trepat, X., Borghi, N., Gloerich, M., et al. (2022). A mechanical G2 checkpoint controls epithelial cell division through E-cadherin-mediated regulation of Wee1-Cdk1. Cell Rep. 41, 111475. doi:10.1016/j.celrep.2022.111475
Du, H., Bartleson, J. M., Butenko, S., Alonso, V., Liu, W. F., Winer, D. A., et al. (2022). Tuning immunity through tissue mechanotransduction. Nat. Rev. Immunol. 23, 174–188. doi:10.1038/s41577-022-00761-w
Duarte, S., Baber, J., Fujii, T., and Coito, A. J. (2015). Matrix metalloproteinases in liver injury, repair and fibrosis. Matrix Biol. 44-46, 147–156. doi:10.1016/j.matbio.2015.01.004
Dvorak, H. F. (1986). Tumors: wounds that do not heal. Similarities between tumor stroma generation and wound healing. N. Engl. J. Med. 315, 1650–1659. doi:10.1056/NEJM198612253152606
Elosegui-Artola, A., Andreu, I., Beedle, A. E. M., Lezamiz, A., Uroz, M., Kosmalska, A. J., et al. (2017). Force triggers YAP nuclear entry by regulating transport across nuclear pores. Cell 171, 1397–1410.e14. doi:10.1016/j.cell.2017.10.008
Elosegui-Artola, A., Gupta, A., Najibi, A. J., Seo, B. R., Garry, R., Tringides, C. M., et al. (2023). Matrix viscoelasticity controls spatiotemporal tissue organization. Nat. Mater 22, 117–127. doi:10.1038/s41563-022-01400-4
Elosegui-Artola, A. (2021). The extracellular matrix viscoelasticity as a regulator of cell and tissue dynamics. Curr. Opin. Cell Biol. 72, 10–18. doi:10.1016/j.ceb.2021.04.002
Fiori, M. E., Di Franco, S., Villanova, L., Bianca, P., Stassi, G., and De Maria, R. (2019). Cancer-associated fibroblasts as abettors of tumor progression at the crossroads of EMT and therapy resistance. Mol. Cancer 18, 70. doi:10.1186/s12943-019-0994-2
Fischer, A., Wannemacher, J., Christ, S., Koopmans, T., Kadri, S., Zhao, J., et al. (2022). Neutrophils direct preexisting matrix to initiate repair in damaged tissues. Nat. Immunol. 23, 518–531. doi:10.1038/s41590-022-01166-6
Friedman, S. L. (2008). Hepatic stellate cells: Protean, multifunctional, and enigmatic cells of the liver. Physiol. Rev. 88, 125–172. doi:10.1152/physrev.00013.2007
Gaertner, F., Reis-Rodrigues, P., de Vries, I., Hons, M., Aguilera, J., Riedl, M., et al. (2022). WASp triggers mechanosensitive actin patches to facilitate immune cell migration in dense tissues. Dev. Cell 57, 47–62.e9. doi:10.1016/j.devcel.2021.11.024
Genschmer, K. R., Russell, D. W., Lal, C., Szul, T., Bratcher, P. E., Noerager, B. D., et al. (2019). Activated PMN Exosomes: Pathogenic Entities Causing Matrix Destruction and Disease in the Lung. Cell 176, 113–126.e15. doi:10.1016/j.cell.2018.12.002
Gilkes, D. M., Bajpai, S., Chaturvedi, P., Wirtz, D., and Semenza, G. L. (2013). Hypoxia-inducible Factor 1 (HIF-1) Promotes Extracellular Matrix Remodeling under Hypoxic Conditions by Inducing P4HA1, P4HA2, and PLOD2 Expression in Fibroblasts. J. Biol. Chem. 288, 10819–10829. doi:10.1074/jbc.M112.442939
Gonzalez-Rodriguez, D., Guevorkian, K., Douezan, S., and Brochard-Wyart, F. (2012). Soft Matter Models of Developing Tissues and Tumors. Science 338, 910–917. doi:10.1126/science.1226418
Gudipaty, S. A., Lindblom, J., Loftus, P. D., Redd, M. J., Edes, K., Davey, C. F., et al. (2017). Mechanical stretch triggers rapid epithelial cell division through Piezo1. Nature 543, 118–121. doi:10.1038/nature21407
Haas, A. J., Zihni, C., Ruppel, A., Hartmann, C., Ebnet, K., Tada, M., et al. (2020). Interplay between Extracellular Matrix Stiffness and JAM-A Regulates Mechanical Load on ZO-1 and Tight Junction Assembly. Cell Rep. 32, 107924. doi:10.1016/j.celrep.2020.107924
Herrero-Cervera, A., Soehnlein, O., and Kenne, E. (2022). Neutrophils in chronic inflammatory diseases. Cell Mol. Immunol. 19, 177–191. doi:10.1038/s41423-021-00832-3
Hua, Y., and Bergers, G. (2019). Tumors vs. Chronic Wounds: An Immune Cell’s Perspective. Front. Immunol. 10, 2178. doi:10.3389/fimmu.2019.02178
Ilina, O., Gritsenko, P. G., Syga, S., Lippoldt, J., La Porta, C. A. M., Chepizhko, O., et al. (2020). Cell–cell adhesion and 3D matrix confinement determine jamming transitions in breast cancer invasion. Nat. Cell Biol. 22 (9), 1103–1115. doi:10.1038/s41556-020-0552-6
Jeljeli, M., Riccio, L. G. C., Doridot, L., Chêne, C., Nicco, C., Chouzenoux, S., et al. (2019). Trained immunity modulates inflammation-induced fibrosis. Nat. Commun. 10, 5670. doi:10.1038/s41467-019-13636-x
Jenne, C. N., Liao, S., and Singh, B. (2018). Neutrophils: multitasking first responders of immunity and tissue homeostasis. Cell Tissue Res. 371, 395–397. doi:10.1007/s00441-018-2802-5
Jiang, Y., Zhang, H., Wang, J., Liu, Y., Luo, T., and Hua, H. (2022). Targeting extracellular matrix stiffness and mechanotransducers to improve cancer therapy. J. Hematol. Oncol. 15, 34. doi:10.1186/s13045-022-01252-0
Judokusumo, E., Tabdanov, E., Kumari, S., Dustin, M. L., and Kam, L. C. (2012). Mechanosensing in T Lymphocyte Activation. Biophys. J. 102, L5–L7. doi:10.1016/J.BPJ.2011.12.011
Kim, S. T., Takeuchi, K., Sun, Z. Y. J., Touma, M., Castro, C. E., Fahmy, A., et al. (2009). The alphabeta T cell receptor is an anisotropic mechanosensor. J. Biol. Chem. 284, 31028–31037. doi:10.1074/jbc.M109.052712
Kobayashi, S., Ogasawara, N., Watanabe, S., Yoneyama, Y., Kirino, S., Hiraguri, Y., et al. (2022). Collagen type I-mediated mechanotransduction controls epithelial cell fate conversion during intestinal inflammation. Inflamm. Regen. 42, 49. doi:10.1186/S41232-022-00237-3
Lang, S., and Schnabl, B. (2020). Microbiota and Fatty Liver Disease—the Known, the Unknown, and the Future. Cell Host Microbe 28, 233–244. doi:10.1016/j.chom.2020.07.007
Li, Y.-C., Chen, B.-M., Wu, P.-C., Cheng, T.-L., Kao, L.-S., Tao, M.-H., et al. (2010). Cutting Edge: Mechanical Forces Acting on T Cells Immobilized via the TCR Complex Can Trigger TCR Signaling. J. Immunol. 184, 5959–5963. doi:10.4049/jimmunol.0900775
Li, Z., and Bratlie, K. M. (2021). Fibroblasts treated with macrophage conditioned medium results in phenotypic shifts and changes in collagen organization. Mater Sci. Eng. C 122, 111915. doi:10.1016/j.msec.2021.111915
Liu, B., Yang, M. Q., Yu, T. Y., Yin, Y. Y., Liu, Y., Wang, X. D., et al. (2021). Mast Cell Tryptase Promotes Inflammatory Bowel Disease-Induced Intestinal Fibrosis. Inflamm. Bowel Dis. 27, 242–255. doi:10.1093/ibd/izaa125
Liu, F., Mih, J. D., Shea, B. S., Kho, A. T., Sharif, A. S., Tager, A. M., et al. (2010). Feedback amplification of fibrosis through matrix stiffening and COX-2 suppression. J. Cell Biol. 190, 693–706. doi:10.1083/jcb.201004082
Mariano, C. A., Sattari, S., Ramirez, G. O., and Eskandari, M. (2023). Effects of tissue degradation by collagenase and elastase on the biaxial mechanics of porcine airways. Respir. Res. 24, 105. doi:10.1186/s12931-023-02376-8
Martin, P., and Leibovich, S. J. (2005). Inflammatory cells during wound repair: The good, the bad and the ugly. Trends Cell Biol. 15, 599–607. doi:10.1016/j.tcb.2005.09.002
McKleroy, W., Lee, T. H., and Atabai, K. (2013). Always cleave up your mess: targeting collagen degradation to treat tissue fibrosis. Am. J. Physiol. Lung Cell Mol. Physiol. 304, L709–L721. doi:10.1152/ajplung.00418.2012
Medeiros, N. I., Fares, R. C. G., Franco, E. P., Sousa, G. R., Mattos, R. T., Chaves, A. T., et al. (2017). Differential Expression of Matrix Metalloproteinases 2, 9 and Cytokines by Neutrophils and Monocytes in the Clinical Forms of Chagas Disease. PLoS Negl. Trop. Dis. 11, e0005284. doi:10.1371/journal.pntd.0005284
Meli, V. S., Atcha, H., Veerasubramanian, P. K., Nagalla, R. R., Luu, T. U., Chen, E. Y., et al. (2023). YAP-mediated mechanotransduction tunes the macrophage inflammatory response. Sci. Adv. 6, eabb8471. doi:10.1126/sciadv.abb8471
Meng, K. P., Majedi, F. S., Thauland, T. J., and Butte, M. J. (2020). Mechanosensing through YAP controls T cell activation and metabolism. J. Exp. Med. 217, e20200053. doi:10.1084/jem.20200053
Millán-Salanova, M., and Vicente-Manzanares, M. (2022). The interface between biochemical signaling and cell mechanics shapes T lymphocyte migration and activation. Eur. J. Cell Biol. 101, 151236. doi:10.1016/j.ejcb.2022.151236
Murrell, M., Kamm, R., and Matsudaira, P. (2011). Substrate Viscosity Enhances Correlation in Epithelial Sheet Movement. Biophys. J. 101, 297–306. doi:10.1016/j.bpj.2011.05.048
Nalkurthi, C., Schroder, W. A., Melino, M., Irvine, K. M., Nyuydzefe, M., Chen, W., et al. (2022). ROCK2 inhibition attenuates profibrogenic immune cell function to reverse thioacetamide-induced liver fibrosis. JHEP Rep. 4, 100386. doi:10.1016/j.jhepr.2021.100386
Ng, M. R., Besser, A., Danuser, G., and Brugge, J. S. (2012). Substrate stiffness regulates cadherin-dependent collective migration through myosin-II contractility. J. Cell Biol. 199, 545–563. doi:10.1083/jcb.201207148
Ollauri-Ibáñez, C., Ayuso-íñigo, B., and Pericacho, M. (2021). Hot and Cold Tumors: Is Endoglin (CD105) a Potential Target for Vessel Normalization? Cancers (Basel) 13, 1552. doi:10.3390/cancers13071552
Ong, C. W. M., Elkington, P. T., Brilha, S., Ugarte-Gil, C., Tome-Esteban, M. T., Tezera, L. B., et al. (2015). Neutrophil-Derived MMP-8 Drives AMPK-Dependent Matrix Destruction in Human Pulmonary Tuberculosis. PLoS Pathog. 11, e1004917. doi:10.1371/journal.ppat.1004917
Ong, C. W. M., Pabisiak, P. J., Brilha, S., Singh, P., Roncaroli, F., Elkington, P. T., et al. (2017). Complex regulation of neutrophil-derived MMP-9 secretion in central nervous system tuberculosis. J. Neuroinflammation 14, 31. doi:10.1186/s12974-017-0801-1
Pérez-González, C., Alert, R., Blanch-Mercader, C., Gómez-González, M., Kolodziej, T., Bazellieres, E., et al. (2019). Active wetting of epithelial tissues. Nat. Phys. 15, 79–88. doi:10.1038/s41567-018-0279-5
Ragkousi, K., and Gibson, M. C. (2014). Cell division and the maintenance of epithelial order. J. Cell Biol. 207, 181–188. doi:10.1083/jcb.201408044
Rodrigues, M., Kosaric, N., Bonham, C. A., and Gurtner, G. C. (2019). Wound Healing: A Cellular Perspective. Physiol. Rev. 99, 665–706. doi:10.1152/physrev.00067.2017
Sahai, E., Astsaturov, I., Cukierman, E., DeNardo, D. G., Egeblad, M., Evans, R. M., et al. (2020). A framework for advancing our understanding of cancer-associated fibroblasts. Nat. Rev. Cancer 20 (3 20), 174–186. doi:10.1038/s41568-019-0238-1
Schnoor, M., Cullen, P., Lorkowski, J., Stolle, K., Robenek, H., Troyer, D., et al. (2008). Production of type VI collagen by human macrophages: a new dimension in macrophage functional heterogeneity. J Immunol 180, 5707–5719. doi:10.4049/jimmunol.180.8.5707
Schrader, J., Gordon-Walker, T. T., Aucott, R. L., van Deemter, M., Quaas, A., Walsh, S., et al. (2011). Matrix Stiffness Modulates Proliferation, Chemotherapeutic Response and Dormancy in Hepatocellular Carcinoma Cells. Hepatology 53, 1192–1205. doi:10.1002/hep.24108
Schwayer, C., Shamipour, S., Pranjic-Ferscha, K., Schauer, A., Balda, M., Tada, M., et al. (2019). Mechanosensation of Tight Junctions Depends on ZO-1 Phase Separation and Flow. Cell 179, 937–952.e18. doi:10.1016/j.cell.2019.10.006
Shimbori, C., Upagupta, C., Bellaye, P.-S., Ayaub, E. A., Sato, S., Yanagihara, T., et al. (2019). Mechanical stress-induced mast cell degranulation activates TGF-β1 signalling pathway in pulmonary fibrosis. Thorax 74, 455–465. doi:10.1136/thoraxjnl-2018-211516
Simões, F. C., Cahill, T. J., Kenyon, A., Gavriouchkina, D., Vieira, J. M., Sun, X., et al. (2020). Macrophages directly contribute collagen to scar formation during zebrafish heart regeneration and mouse heart repair. Nat. Commun. 11 (1), 600–617. doi:10.1038/s41467-019-14263-2
Spadaro, D., Le, S., Laroche, T., Mean, I., Jond, L., Yan, J., et al. (2017). Tension-Dependent Stretching Activates ZO-1 to Control the Junctional Localization of Its Interactors. Curr. Biol. 27, 3783–3795.e8. doi:10.1016/j.cub.2017.11.014
Sridharan, R., Cavanagh, B., Cameron, A. R., Kelly, D. J., and O’Brien, F. J. (2019). Material stiffness influences the polarization state, function and migration mode of macrophages. Acta Biomater. 89, 47–59. doi:10.1016/j.actbio.2019.02.048
Staddon, M. F., Bi, D., Tabatabai, A. P., Ajeti, V., Murrell, M. P., and Banerjee, S. (2018). Cooperation of dual modes of cell motility promotes epithelial stress relaxation to accelerate wound healing. PLoS Comput. Biol. 14, e1006502. doi:10.1371/journal.pcbi.1006502
Sutherland, T. E., Dyer, D. P., and Allen, J. E. (2023). The extracellular matrix and the immune system: A mutually dependent relationship. Science 379, eabp8964. doi:10.1126/science.abp8964
Tomlin, H., and Piccinini, A. M. (2018). A complex interplay between the extracellular matrix and the innate immune response to microbial pathogens. Immunology 155, 186–201. doi:10.1111/imm.12972
Velotti, F., Barchetta, I., Cimini, F. A., and Cavallo, M. G. (2020). Granzyme B in Inflammatory Diseases: Apoptosis, Inflammation, Extracellular Matrix Remodeling, Epithelial-to-Mesenchymal Transition and Fibrosis. Front. Immunol. 11, 587581. doi:10.3389/fimmu.2020.587581
Wang, Y., Huang, B., Jin, T., Ocansey, D. K. W., Jiang, J., and Mao, F. (2022). Intestinal Fibrosis in Inflammatory Bowel Disease and the Prospects of Mesenchymal Stem Cell Therapy. Front. Immunol. 13, 835005. doi:10.3389/fimmu.2022.835005
Witherel, C. E., Sao, K., Brisson, B. K., Han, B., Volk, S. W., Petrie, R. J., et al. (2021). Regulation of extracellular matrix assembly and structure by hybrid M1/M2 macrophages. Biomaterials 269, 120667. doi:10.1016/j.biomaterials.2021.120667
Wu, Y., Cao, Y., Xu, K., Zhu, Y., Qiao, Y., Wu, Y., et al. (2021). Dynamically remodeled hepatic extracellular matrix predicts prognosis of early-stage cirrhosis. Cell Death Dis. 12, 163. doi:10.1038/s41419-021-03443-y
Xu, J., Mao, X., Jin, R., Yin, J., Lu, K., Guo, Y., et al. (2020). Neutrophil extracellular traps degrade fibronectin in a rat model of bronchopulmonary dysplasia induced by perinatal exposure to lipopolysaccharide. J. Cell Mol. Med. 24, 14645–14649. doi:10.1111/jcmm.15842
Yahoo, N., Dudek, M., Knolle, P., and Heikenwälder, M. (2023). Role of immune responses in the development of NAFLD-associated liver cancer and prospects for therapeutic modulation. J. Hepatol. doi:10.1016/j.jhep.2023.02.033
Yap, A. S., Duszyc, K., and Viasnoff, V. (2018). Mechanosensing and mechanotransduction at cell –cell junctions. Cold Spring Harb. Perspect. Biol. 10, a028761. doi:10.1101/cshperspect.a028761
Yu, Y., Ren, L.-J., Liu, X.-Y., Gong, X.-B., and Yao, W. (2021). Effects of substrate stiffness on mast cell migration. Eur. J. Cell Biol. 100, 151178. doi:10.1016/j.ejcb.2021.151178
Yui, S., Azzolin, L., Maimets, M., Pedersen, M. T., Fordham, R. P., Hansen, S. L., et al. (2018). YAP/TAZ-Dependent Reprogramming of Colonic Epithelium Links ECM Remodeling to Tissue Regeneration. Cell Stem Cell 22, 35–49.e7. doi:10.1016/j.stem.2017.11.001
Zhang, J., Li, W., Sanders, M. A., Sumpio, B. E., Panja, A., and Basson, M. D. (2003). Regulation of the intestinal epithelial response to cyclic strain by extracellular matrix proteins. FASEB J. 17, 926–928. doi:10.1096/fj.02-0663fje
Keywords: epithelial barrier, inflammation, immune cells, extracellular matrix, tissue mechanics, chronic inflammatory diseases, immuno-biophysics, immuno-mechanobiology
Citation: Saha S, Müller D and Clark AG (2023) Mechanosensory feedback loops during chronic inflammation. Front. Cell Dev. Biol. 11:1225677. doi: 10.3389/fcell.2023.1225677
Received: 19 May 2023; Accepted: 27 June 2023;
Published: 10 July 2023.
Edited by:
Carlos Pardo-Pastor, King’s College London, United KingdomReviewed by:
Sharon Celeste Morley, Washington University in St. Louis, United StatesCopyright © 2023 Saha, Müller and Clark. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Andrew G. Clark, andrew.clark@srcsb.uni-stuttgart.de