Resumos
Em Pterodon pubescens, os pulvinos primário e secundário, órgãos responsáveis pelos movimentos foliares, apresentam peculiaridades estruturais em relação ao pecíolo e raque. Estas peculiaridades incluem cutícula mais espessa, córtex mais desenvolvido formado por células parenquimáticas de formas variáveis, sistema vascular em posição central, floema circundado por fibras septadas não lignificadas e medula parenquimática reduzida ou ausente. Pecíolo e raque apresentam córtex reduzido constituído por células parenquimáticas isodiamétricas, sistema vascular periférico, floema envolvido por calota de fibras gelatinosas e medula ampla. Compostos fenólicos foram detectados no vacúolo das células corticais dos pulvinos, enquanto que no córtex e medula do pecíolo e da raque foram observados amiloplastos em abundância e ausência de compostos fenólicos. Em geral, as características estruturais do pulvino de P. pubescens são comuns aos pulvinos das demais espécies de Fabaceae e têm sido relacionadas com a intensificação de trocas laterais de íons e água entre células do córtex e sistema vascular, além de proporcionar maior flexibilidade e capacidade de movimento desta região. Já as características do pecíolo e da raque conferem maior rigidez a estas estruturas foliares.
anatomia; pecíolo; Pterodon pubescens; pulvino; raque
In Pterodon pubescens the primary and secondary pulvini show structural peculiarities in relations of petiole and rachis. These peculiarities include thicker cuticule, more developed cortex with only parenchyma cells with irregular shape, central vascular tissues, presence of septate fibers around the phloem, and absent or reduced pith. Petiole and rachis show reduced cortex constituted by parenchyma isodiameters cells, peripheric vascular tissues surrounded by gelatinous fibers and pith. Phenolic compounds were detected inside vacuole of parenchyma cells of the pulvinus. Starch grains were observed in the parenchyma cells of petiole and rachis. The features of P. pubescens pulvinus are common to pulvini of others species of Fabaceae, and have been associated to the intensification of lateral changes of ion and water between cortical cells and vascular tissues, besides to provide more flexibility and movement capacity of this organ. On the other hand, the features of petiole and rachis give more rigidity to these leaves structures.
anatomy; petiole; Pterodon pubescens; pulvinus; rachis
Untitled1
Anatomia comparada do pulvino, pecíolo e raque de Pterodon pubescens Benth. (Fabaceae - Faboideae)
Comparative anatomy of pulvinus, petiole and rachis of Pterodon pubescens Benth. (Fabaceae - Faboideae)
Tatiane Maria RodriguesI; Sílvia Rodrigues MachadoII,1 1 Autor para correspondência: smachado@ibb.unesp.br
IBolsista de Iniciação Científica, FAPESP (01/01588-4)
IIDepartamento de Botânica, Instituto de Biociências, UNESP, Campus de Botucatu, C. Postal 510, CEP 18618-000, Botucatu, SP, Brasil
RESUMO
Em Pterodon pubescens, os pulvinos primário e secundário, órgãos responsáveis pelos movimentos foliares, apresentam peculiaridades estruturais em relação ao pecíolo e raque. Estas peculiaridades incluem cutícula mais espessa, córtex mais desenvolvido formado por células parenquimáticas de formas variáveis, sistema vascular em posição central, floema circundado por fibras septadas não lignificadas e medula parenquimática reduzida ou ausente. Pecíolo e raque apresentam córtex reduzido constituído por células parenquimáticas isodiamétricas, sistema vascular periférico, floema envolvido por calota de fibras gelatinosas e medula ampla. Compostos fenólicos foram detectados no vacúolo das células corticais dos pulvinos, enquanto que no córtex e medula do pecíolo e da raque foram observados amiloplastos em abundância e ausência de compostos fenólicos. Em geral, as características estruturais do pulvino de P. pubescens são comuns aos pulvinos das demais espécies de Fabaceae e têm sido relacionadas com a intensificação de trocas laterais de íons e água entre células do córtex e sistema vascular, além de proporcionar maior flexibilidade e capacidade de movimento desta região. Já as características do pecíolo e da raque conferem maior rigidez a estas estruturas foliares.
Palavras-chave: anatomia, pecíolo, Pterodon pubescens, pulvino, raque
ABSTRACT
In Pterodon pubescens the primary and secondary pulvini show structural peculiarities in relations of petiole and rachis. These peculiarities include thicker cuticule, more developed cortex with only parenchyma cells with irregular shape, central vascular tissues, presence of septate fibers around the phloem, and absent or reduced pith. Petiole and rachis show reduced cortex constituted by parenchyma isodiameters cells, peripheric vascular tissues surrounded by gelatinous fibers and pith. Phenolic compounds were detected inside vacuole of parenchyma cells of the pulvinus. Starch grains were observed in the parenchyma cells of petiole and rachis. The features of P. pubescens pulvinus are common to pulvini of others species of Fabaceae, and have been associated to the intensification of lateral changes of ion and water between cortical cells and vascular tissues, besides to provide more flexibility and movement capacity of this organ. On the other hand, the features of petiole and rachis give more rigidity to these leaves structures.
Key words: anatomy, petiole, Pterodon pubescens, pulvinus, rachis
Introdução
Os eventos fisiológicos responsáveis pelos movimentos foliares em diferentes espécies de leguminosas têm sido correlacionados com características estruturais do pulvino, principalmente a existência de um arranjo estrutural que assegura mudanças celulares rápidas, além de características peculiares das células motoras e do sistema vascular que facilitam tanto a sua deformação, quanto o fluxo lateral de íons K+ e de água (Satter et al. 1970; Morse & Satter 1979; Campbell & Garber 1980; Fleurat-Lessard 1981; Satter & Galston 1981; Fleurat-Lessard & Roblin 1982; Fleurat-Lessard & Satter 1985; Moysset & Simón 1991).
Informações sobre a histologia comparada do pulvino, pecíolo e raque são restritas a poucas espécies de leguminosas, especialmente Mimosa pudica que se caracteriza por apresentar movimentos foliares rápidos (Fleurat-Lessard & Bonnemain 1978; Fleurat-Lessard & Roblin 1982). Nesta espécie, pecíolo e pulvino diferem principalmente quanto a posição e composição do cilindro vascular e, segundo os autores, essas diferenças seriam as principais responsáveis pela grande flexibilidade do pulvino e maior rigidez do pecíolo.
De acordo com Fleurat-Lessard & Bonnemain (1978), uma característica histológica particular de pulvinos em relação ao pecíolo e raque, é a substituição das fibras floemáticas por colênquima e a redução ou completa substituição das células do parênquima xilemático por elementos fibriformes vivos. Esses autores ressaltam a falta de estudos comparativos sobre a histologia dessas estruturas foliares para verificar se as analogias e homologias observadas em M. pudica ocorrem em outras espécies de leguminosas com movimentos foliares lentos ou intermediários.
Pterodon pubescens, leguminosa arbórea nativa de cerrados, apresenta movimentos heliotrópicos lentos e reversíveis resultantes da curvatura dos pulvinos primário e secundário (Caldas et al. 1997). Nesta espécie, os autores constataram que o ajuste dos folíolos mantém as reações fotoquímicas em taxas elevadas, maximizando a eficiência fotossintética em condições adversas. Dados sobre a estrutura do pulvino e demais regiões foliares de P. pubescens não foram encontradas na literatura.
Visando fornecer dados que possibilitem relacionar os eventos fisiológicos responsáveis pelos movimentos foliares lentos de P. pubescens com características estruturais da folha, neste trabalho foi realizada uma análise anatômica comparativa dos pulvinos, pecíolo e raque.
Material e métodos
Folhas, com o limbo completamente expandido, foram coletadas de ramos localizados na periferia da copa de exemplares adultos de P. pubescens ocorrentes em cerrado no município de Botucatu, SP (22º55'S, 48º30'W). As amostras foram retiradas da região mediana dos pulvinos primário e secundário, pecíolo e raque e processadas conforme descrito a seguir. Ramos férteis foram herborizados e depositados como documento no Herbário BOTU, do Instituto de Biociências, UNESP, Campus de Botucatu.
Parte das amostras foi estocada em álcool 70%, seccionada com o auxílio de micrótomo de Ranvier, sendo as secções, com 9µm de espessura, clarificadas em hipoclorito de sódio a 20%, lavadas em água acética a 1%, coradas com Azul de Astra e Fucsina básica e montadas entre lâmina e lamínula com glicerina (Roeser 1972). Outra parte foi fixada em FAA 50 (Johansen 1940), desidratada em série etílica, incluída em resina glicol-metacrilato e seccionada em micrótomo rotativo; as secções, com 6µm a 8µm de espessura, foram coradas em Azul de Toluidina 0,05%, pH 4,7 (O'Brien et al. 1964) e montadas entre lâmina e lamínula com Permount. Secções de material fresco, obtidas com auxílio de lâminas de barbear, foram tratadas com Sudan IV para detecção de substâncias lipídicas (Johansen 1940); Vermelho de Rutênio para detecção de substâncias pécticas (Jensen 1962); Reagente de Lugol, para amido e alcalóides (Johansen 1940) e Cloreto Férrico, para substâncias fenólicas (Johansen 1940). Os aspectos relevantes foram fotografados em fotomicroscópio Zeiss, utilizando-se filme Kodak Asa 100.
Resultados
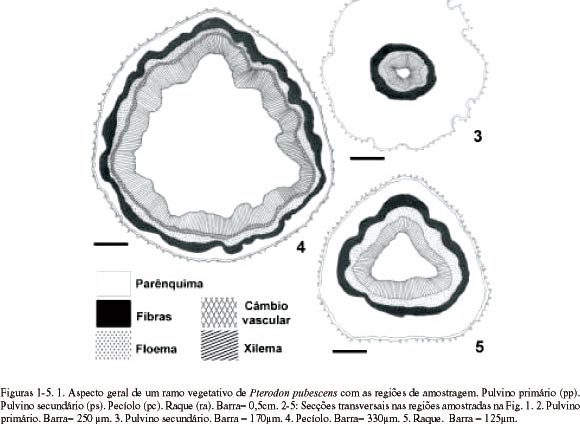
Organização geral do pulvino, pecíolo e raque (Fig. 1) Os pulvinos primário (Fig. 2) e secundário (Fig. 3), em secção transversal, são dorsiventrais a cilíndricos e apresentam contorno irregular devido às reentrâncias e protuberâncias, as quais são mais desenvolvidas no pulvino secundário. Possuem epiderme com tricomas tectores, córtex desenvolvido, sistema vascular central e medula reduzida ou ausente.
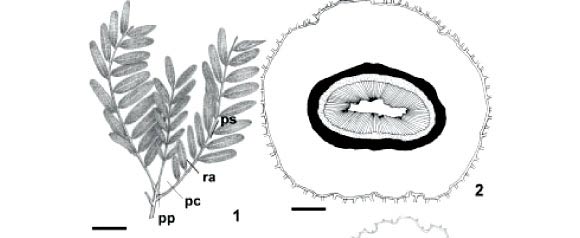
O pecíolo (Fig. 4) e a raque (Fig. 5), em secção transversal, são dorsiventrais com formato triangular e contorno regular. O pecíolo apresenta córtex reduzido, sistema vascular periférico com câmbio ativo e medula ampla. A raque possui córtex relativamente amplo, sistema vascular em posição mediana e medula pouco desenvolvida. Uma bainha de fibras, de espessura e contorno variáveis, envolve o floema das quatro regiões analisadas (Fig. 2-5), sendo mais espessa nos pulvinos.
Epiderme - Os pulvinos apresentam epiderme revestida por cutícula espessa e constituída por células papilosas (Fig. 13); no pecíolo (Fig. 19) e raque (Fig. 23), a cutícula é relativamente mais delgada e as células epidérmicas são retangulares e de tamanhos variáveis. Acúmulo de composto fenólicos foi detectado nos vacúolos das células epidérmicas do pecíolo (Fig. 19) e raque (Fig. 22).

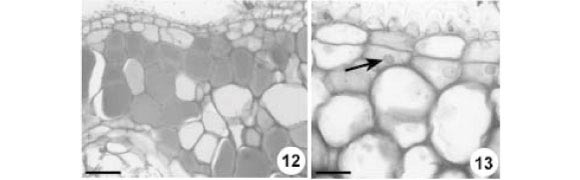




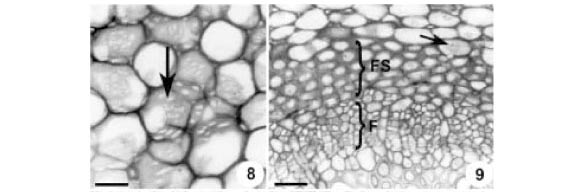
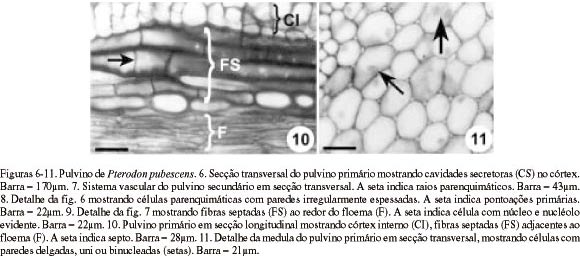
Córtex - No pulvino primário, o córtex é constituído por cerca de 30 camadas de células parenquimáticas com formas e tamanhos irregulares, de disposição compacta (Fig. 6) e paredes irregularmente espessadas, mostrando numerosos campos de pontoação primários amplos (Fig. 8). No pulvino secundário, o córtex possui cerca de 10 camadas celulares, com o mesmo arranjo e disposição que no primário. No pecíolo (Fig. 18, 19) e na raque (Fig. 22), o córtex é formado por seis a oito camadas de células parenquimáticas de paredes delgadas.
Em ambos os pulvinos, chama a atenção a ocorrência de divisões periclinais nas células corticais subjacentes à epiderme (Fig. 13), cujas resultantes, com núcleo esférico e nucléolo evidente, originam protuberâncias em forma de almofadas na superfície do pulvino (Fig. 6, 14). A quantidade de células em divisão e o grau de desenvolvimento das protuberâncias variam ao longo da circunferência do pulvino.
Na região cortical mais externa do pulvino primário (Fig. 6, 12), ocorrem agrupamentos de células que se caracterizam pelo maior tamanho, acúmulo de substâncias fenólicas no vacúolo e ausência de grãos de amido. Não se notam diferenças no tamanho e formato entre as células corticais do pulvino secundário. As camadas corticais mais internas de ambos os pulvinos caracterizam-se pela presença de grãos de amido (Fig. 16, 17) e ausência de compostos fenólicos; na endoderme, além de grãos de amido ocorrem cristais prismáticos de oxalato de cálcio (Fig. 15, 16).
No pecíolo (Fig. 19) e na raque (Fig. 22), as células corticais mais periféricas são menos volumosas que as mais internas e apresentam-se repletas de grãos de amido (Fig. 23), sem evidências de acúmulo de substâncias fenólicas; as células da endoderme, entretanto, apresentam compostos fenólicos (Fig. 19, 20) e ausência de grãos de amido.
Cavidades secretoras são observadas no córtex de todas as regiões analisadas, sendo mais desenvolvidas no pulvino primário (Fig. 6).
Sistema Vascular - Os pulvinos primário (Fig. 2, 6) e secundário (Fig. 3), em secção transversal, apresentam sistema vascular de posição central e formado por um feixe concêntrico. Circundando o floema, observam-se cinco a sete camadas de células pequenas, com citoplasma denso, núcleo volumoso com nucléolo evidente e paredes pecto-celulósicas espessadas (Fig. 6, 9). Em secção longitudinal (Fig. 10), estas células são fusiformes, apresentam crescimento intrusivo e possuem septos, sendo, portanto, fibras septadas. Em suas paredes não lignificadas (Fig. 15, 16) são visíveis pontoações simples amplas. Por entre estas células, ocorrem fibras gelatinosas esparsas que se caracterizam por apresentar a camada parietal interna não lignificada (Fig. 16).
No pecíolo (Fig. 18) e raque (Fig. 22), o sistema vascular é formado por vários feixes colaterais contíguos dispostos em um círculo periférico.
O floema no pecíolo é envolvido por uma calota de fibras de paredes espessas e lignificadas (Fig. 20), enquanto que na raque, é circundado por fibras gelatinosas (Fig. 24 e 25); nas quatro regiões foliares estudadas, é constituído por elementos de tubo crivado, células companheiras e células parenquimáticas (Fig. 9, 19, 20, 26). Na raque, o floema apresenta células parenquimáticas alongadas, volumosas, preenchidas por substâncias fenólicas (Fig. 26) e com parede relativamente espessada, algumas em processo de esclerificação.
O xilema, nos pulvinos primário (Fig. 7) e secundário, pecíolo (Fig. 19) e raque (Fig. 26) é constituído por elementos de vaso associados com células de parênquima e elementos fibriformes. Raios parenquimáticos com número variável de células são nítidos no xilema do pulvino primário (Fig. 7) e pecíolo (Fig. 20). No pecíolo (Fig. 19, 20) nota-se câmbio ativo na produção de xilema e floema secundários.
A região medular, em ambos os pulvinos é constituída por um pequeno número de células de paredes delgadas, com núcleo e nucléolo evidentes (Fig. 11); no pecíolo (Fig. 21) e raque (Fig. 27), é formada por numerosas células parenquimáticas volumosas, repletas de grãos de amido, além de esclereídes esparsas.
Discussão
As diferenças anatômicas observadas entre pulvino, pecíolo e raque de P. pubescens são, de modo geral, similares às relatadas para Mimosa pudica (Fleurat-Lessard & Roblin 1982).
O contorno irregular do pulvino, com protuberâncias em forma de "almofadas", é uma das características que o diferencia do pecíolo e raque e, segundo estudos realizados em M. pudica por Toriyama (1953), a presença dessas protuberâncias está associada à capacidade de movimentação do pulvino. A cutícula mais espessa no pulvino pode funcionar como uma barreira eficiente à perda de água, mantendo esta região distendida mesmo em condições de estresse hídrico (Kolattukudy 1984).
Outra característica dos pulvinos primário e secundário de P. pubescens em relação ao pecíolo e à raque é o grande desenvolvimento da região cortical, de natureza exclusivamente parenquimática, cujas células situadas na porção mais periférica do córtex apresentam conteúdo fenólico, ausência de grãos de amido e paredes sinuosas ou dobradas, com campos de pontoação primários amplos. Estas células foram denominadas células motoras por Toriyama (1953), sendo consideradas as principais responsáveis pelos movimentos foliares. Paredes sinuosas ou dobradas são evidências de modificações no compartimento apoplástico (Fleurat-Lessard 1988; Moysset & Simón 1991). Segundo Fleurat-Lessard & Millet (1984), mudanças na extensibilidade da parede das células motoras poderiam estar associadas com o AIA, cuja concentração é três vezes maior no pulvino em relação às outras partes da planta de M. pudica.
Abundância de compostos fenólicos e escassez de grãos de amido nas células motoras, conforme observado em P. pubescens é característica marcante em pulvinos (Esau 1965), sendo esta relação inversa nas células da endoderme, as quais apresentam grãos de amido e ausência de compostos fenólicos, como também observado em M. pudica por Fleurat-Lessard (1981).
A presença de muitos amiloplastos nas células corticais e medulares do pecíolo e raque, juntamente com ausência de modificações no contorno das paredes celulares, indica que, nestas regiões, a principal função do parênquima é a de reserva.
A ocorrência de compostos fenólicos, principalmente taninos, no vacúolo das células motoras é característica comum em pulvinos de diferentes espécies (Fleurat-Lessard 1988; Moysset & Simón 1991). Taninos intervêm na estocagem e liberação de íons, especialmente do cálcio, o qual está envolvido na agregação dos componentes do citoesqueleto, particularmente dos microfilamentos de actina em células motoras (Toriyama & Satô 1971; Fleurat-Lessard & Millet 1984; Fleurat-Lessard 1988; Kameyama et al. 2000; Yamashiro et al. 2001).
A camada cortical mais interna dos pulvinos é uma endoderme típica (Schrempf et al. 1976; Morse & Satter 1979; Fleurat-Lessard & Roblin 1982; Moysset & Simón 1991), e segundo alguns autores (Fleurat-Lessard 1988, Chang et al. 2001) não intervém diretamente no movimento do pulvino, mas sim na percepção da gravidade. Presença unicamente de amiloplastos nesta camada é característica de pulvinos com movimentos rápidos, como os de M. pudica; já, a ocorrência simultânea de cristais de oxalato de cálcio e de amiloplastos, conforme observado em P. pubescens, é característica de pulvinos de plantas com movimentos mais lentos ou imperceptíveis (Schrempf et al. 1976; Morse & Satter 1979).
Peculiaridade do sistema vascular de pulvinos, relatada por diferentes autores (Moysset & Simón 1991; Fleurat-Lessard & Bonnemain 1978; Fleurat-Lessard & Roblin 1982), é a substituição de fibras lignificadas do floema por uma bainha de células vivas, a qual tem sido interpretada como sendo um colênquima ou um parênquima colenquimatoso. Deve-se salientar que nos trabalhos consultados não foram apresentadas ilustrações ou descrições desta região em secções longitudinais. Em P. pubescens verificou-se que esta região do pulvino é constituída por fibras septadas de paredes celulósicas espessas não lignificadas e com campos de pontoação amplos, os quais garantem continuidade simplástica entre a região cortical e o cilindro vascular. Além disso, a ausência de lignina em suas paredes proporciona maior flexibilidade a esta região da folha.
No pecíolo de P. pubescens a presença de fibras lignificadas desprovidas de conteúdo vivo ao redor do floema indica descontinuidade simplástica entre o córtex e o sistema vascular, além de conferir rigidez a essa região da folha.
A presença de fibras gelatinosas em folhas de P. pubescens, especialmente mais abundantes no pecíolo e raque em relação ao pulvino, é característica já relatada para outras plantas de cerrado, tais como Plathymenia reticulata (Paviani 1974) e Brasilia sickii (Paviani 1972;1977), e pode estar associada com reserva de água, uma vez que a parede interna é altamente higroscópica (Paviani 1978).
Paredes celulares dobradas ou sinuosas, escassez ou ausência de lignificação e ocorrência generalizada de campos de pontoação primários, indicando conexão simplástica entre todos os tecidos, são características que diferenciam o pulvino das demais regiões foliares de P. pubescens.
Agradecimentos
À FAPESP, pela concessão de bolsa de Iniciação Científica à primeira autora (Processo 01/01588-4) e apoio financeiro ao desenvolvimento deste trabalho (BIOTA-Processo 00/12469-3).
Recebido em 16/05/2003. Aceito em 22/10/2003
- Caldas, L.S.; Lüttge, U.; Franco, A.C. & Haridasan, M. 1997. Leaf heliotropism in Pterodon pubescens, a woody legume from the brazilian cerrado. Revista Brasileira de Fisiologia Vegetal 9: 1-7.
- Campbell, N.A. & Garber, R.C. 1980. Vacuolar reorganization in motor cells of Albizzia during leaf movement. Planta 148: 251-255.
- Chang, S-C.; Cho, M.H.; Kang, B.G. & Kaufman, P.B. 2001. Changes in starch content in oat (Avena sativa) shoot pulvini during the gravitropic response. Journal of Experimental Botany 52: 1029-1040.
- Esau, K. 1965. Plant anatomy 2nd Ed., J. Wiley and Sons, New York, London, Sydney.
- Fleurat-Lessard, P. 1981. Ultrastructural features of the starch sheat cells of the primary pulvinus after gravistimulation of the sensitive plant (Mimosa pudica L.). Protoplasma 105: 177-184.
- Fleurat-Lessard, P. 1988. Structural and ultrastructural features of cortical cells in motor organs of sensitive plants. Biological Review 63: 1-22.
- Fleurat-Lessard, P. & Bonnemain, J-L. 1978. Structural and ultrastructural characteristics of the vascular apparatus of the sensitive plant (Mimosa pudica L.). Protoplasma 94: 127-143.
- Fleurat-Lessard, P. & Roblin, G. 1982. Comparative histocitology of the petiole and the main pulvinus in Mimosa pudica L. Annals of Botany 50: 83-92.
- Fleurat-Lessard, P. & Millet, B. 1984. Ultrastructural features of cortical parenchyma cells ("motor cells") in stamen filaments of Berberis canadensis Mill. and tertiary pulvini of Mimosa pudica L. Journal of Experimental Botany 35: 1332-1341.
- Fleurat-Lessard, P. & Satter, R.L. 1985. Relationship between structure and motility of Albizzia motor organs: changes in ultrastructure of cortical cells during dark-induced closure. Protoplasma 128: 72-79.
- Jensen, W.A. 1962. Botanical histochemistry: principle and practice W.H. Freeman, San Francisco.
- Johansen, D.A. 1940. Plant michrotechnique McGraw-Hill Book, New York.
- Kameyama, K.; Kishi, Y.; Yoshimura, M.; Kanzawa, N.; Sameshima, M. & Tsuchiya, T. 2000. Tyrosine phosphorylation in plant bending. Nature 407: 37.
- Kolattukudy, P.E. 1984. Biochemistry and function of cutin and suberin. Canadian Journal of Botany 62: 2918-2933.
- Morse, M.J. & Satter, R.L. 1979. Relationships between motor cell ultrastructure and leaf movements in Samanea saman Physiologia Plantarum 46: 338-346.
- Moysset, L. & Simón, E. 1991. Secondary pulvinus of Robinia pseudoacacia (Leguminosae): structural and ultrastructural features. American Journal of Botany 78: 1467-1486.
- O'Brien, T.P.; Feder, N. & Mccully, M.E. 1964. Polychromatic staining of plant cell walls by toluidine blue O. Protoplasma 59: 368-373.
- Paviani, T.I. 1972. Estudo morfológico e anatômico de Brasilia sickii G.M. Barroso. Revista Brasileira de Biologia 32: 451-472.
- Paviani, T.I. 1974. Sobre a ocorrência de fibras gelatinosas em Plathymenia reticulata Benth. Ciência & Cultura26: 783-786.
- Paviani, T.I. 1977. Estudo morfológico e anatômico de Brasilia sickii G.M. Barroso. II. Anatomia da raiz, do xilopódio e do caule. Revista Brasileira de Biologia 37: 307-324.
- Paviani, T.I. 1978. Anatomia vegetal e cerrado. Ciência & Cultura30: 1076-1086.
- Roeser, K.R. 1972. Die nadel der schwarzkiefer - massenprodikt und kunstwerk der nautr. Mikrokosmos 61(2): 33-36.
- Satter, R.L.; Sabinis, D.D. & Galston, A.W. 1970. Phytochrome controlled nyctinasty in Albizzia julibrissin I. Anatomy and fine structure of the pulvinule. American Journal of Botany 57(4): 374-381.
- Satter, R.L. & Galston, A.W. 1981. Mechanisms of control of leaf movements. Annual Review of Plant Physiology 32: 83-110.
- Schrempf, M.; Satter, R.L. & Galston, A.W. 1976. Potassium-linked chloride fluxes during rhythmic leaf movement of Albizzia julibrissin Plant Physiology 58: 190-192.
- Toriyama, H. 1953. Observational and experimental studies of sensitive plants. I. The structure of parenchymatous cells of pulvinus. Cytology 18: 283-292.
- Toriyama, H. & Sato, S. 1971. On the contents of the central vacuole in the Mimosa motor cell. Cytology 36(2): 359-375.
- Yamashiro, S.; Kameyama, K.; Kanzawa, N.; Tamuya, T.; Mabuchi, I. & Tsuchiya, T. 2001. The gelsolin/fragmin family protein identified in the higher plant Mimosa pudica Journal of Biochemistry 130(2): 243-249.
Datas de Publicação
-
Publicação nesta coleção
23 Set 2004 -
Data do Fascículo
Jun 2004