





Abstract
Angiogenesis is essential for normal mammalian development and is controlled by the local balance of pro- and antiangiogenic factors. Here we describe a novel mouse cDNA sequence encoding sFLT-1 that is a potent antagonist to vascular endothelial growth factor (VEGF) and show for the first time its in vivo production. In situ hybridization and Northern blot analysis with probes specific for sFLT-1 or FLT-1 showed that the relative abundance of their mRNAs changed markedly in spongiotrophoblast cells in the placenta as gestation progressed. On day 11 of pregnancy, sFLT-1 mRNA was undetectable but FLT-1 readily apparent, and by day 17 sFLT-1 mRNA was abundant but FLT-1 barely detectable. sFLT-1 was identified in conditioned medium of cultured placenta from day 17 pregnant mice and likely to be present in the circulation, as there is a substantial increase of VEGF-binding activity in the serum from day 13 of pregnancy, which coincides with the abundant sFLT-1 expression in placenta. Expression of sFLT-1 was also observed in adult lung, kidney, liver, and uterus. These data suggest a novel mechanism of regulation of angiogenesis by alternative splicing of FLT-1 pre-mRNA. Treatment of pregnant mice with exogenous VEGF from day 9 to 17 of pregnancy, which alters the ratio of VEGF to sFLT-1, resulted in an increase in the number of resorption sites and fibrin deposition in the placenta of ongoing pregnancies. These findings have important implications for understanding placental function and may be relevant in a range of disease states.
INTRODUCTION
Angiogenesis, the sprouting of new capillaries, is rare in the adult with exceptions in the female reproductive tract and in pathological conditions (1–3). Mammalian placentation requires extensive angiogenesis to establish a suitable vascular network for the supply of oxygen and nutrients to the fetus (4). A variety of angiogenic growth factors including fibroblast growth factors (5–7) and some of the vascular endothelial growth factor (VEGF) family (8–12) are expressed in placenta. However, rapid growth of placenta in early pregnancy is accomplished in an unusually hypoxic environment (13), suggesting that hypoxia-induced angiogenesis mediated by angiogenic activators such as VEGF and its receptors may be of particular importance (14, 15). In the placenta, VEGF transcripts were detected in maternal and labyrinthine layers and also in trophoblast giant cells (10, 11, 16). VEGFR1 (FLT-1) is found in the spongiotrophoblast layer and VEGFR2 (FLK) in labyrinthine layer (10, 11). In tissues where blood vessel growth is occurring, the net angiogenic effect is controlled by the balance between angiogenic inducers and inhibitors (17). Thus the identification of specific antiangiogenic agents in the placenta, such as PRP (18) and sFLT-1, described in this paper, is of considerable importance for understanding placental and fetal growth.
Vascular endothelial growth factor (VEGF), a potent and endothelial-specific mitogen, has been demonstrated to have a pivotal role in vasculogenesis and angiogenesis (19–22). This protein was independently isolated as vascular permeability factor (23) and is a potent stimulator of this process (24, 25). Alternative splicing of the pre-mRNA encoding one of the VEGF receptors (FLT-1) results in the production of a soluble form comprising the ligand-binding domain of this receptor (sFLT-1), which is a potent antagonist of VEGF (26). In addition to inhibiting VEGF binding to cell surface receptors, sFLT-1 also forms heterodimers with the other VEGF receptor, KDR (27). This would account for the efficient inhibition of VEGF function by sFLT-1. Soluble FLT-1 chimeric proteins have also been shown to suppress retinal neovascularization (28) and corpus luteum angiogenesis (29). However, until now there are no data available showing whether sFLT-1 is present in vivo, or when and where it may be produced. We and others have previously shown the expression of FLT-1 by human trophoblast (30, 31) and mouse spongiotrophoblast (10, 11). In this study we cloned a 3′-end cDNA fragment of mouse sFLT-1 and showed stage-dependent expression of FLT-1 and sFLT-1 in placenta, suggesting a novel mechanism of regulation of angiogenesis by alternative splicing of FLT-1 pre-mRNA. We also explored the effect of exogenous VEGF on the developing placenta in mice, which showed that perturbation of the VEGF to sFLT-1 ratio by administration of exogenous VEGF led to fibrin deposition in the placenta and a reduction in embryo weight. Angiogenesis is currently the target for therapies in a wide range of diseases, including cancer, retinopathy, and heart disease. These findings have important implications for the regulation of angiogenesis in mammals.
RESULTS
Cloning of 3′-End of Mouse sFLT-1
The 3′-end of mouse sFLT-1 was cloned using 3′-RACE (rapid amplification of cDNA ends) and characterized by DNA sequencing. The cloned fragment includes 150 bp of coding sequence, an inframe stop codon, and 18 bp of noncoding sequence followed by a poly (A)+ tail (EMBL accession No. AJ001177). This 171-bp fragment shares 85.5% nucleotide sequence identity to the corresponding region of human sFLT-1 as determined by the alignment program GAP (GCG, Madison, WI). However, the 3′-noncoding sequence of mouse sFLT-1 is much shorter than that of human sFLT-1 (338 bp) (26). The nucleotides around the site of FLT-1 and sFLT-1 divergence are identical in human and mouse, suggesting that splice site skipping occurs in the mouse as well as in the human, as suggested by Kendall and Thomas (26). Alignment of the 3′-end cDNA (which is not present in FLT-1) and the deduced C-terminal amino acid sequence of mouse sFLT-1 with those of human sFLT-1 is shown in Fig. 1.

Alignment of the 3′-End of cDNA and C-Terminal Amino Acid Sequence of Mouse sFLT-1 (m) with Those of Human sFLT-1 (h) The deduced C-terminal amino acid sequence of mouse and human sFLT-1 are above or below their cDNA sequence, respectively. Arrowhead, Divergent site of membrane-bound FLT-1 and sFLT-1.
sFLT-1 Expression Increased, but FLT-1 Decreased, in Placenta in Late Gestation
To investigate the sites and relative expression levels of FLT-1 and sFLT-1 in the placenta, in situ hybridization using probes specific for the two forms was performed on sections prepared from placentas at different stages of gestation. The specificity of the probes demonstrated by Northern blot is shown in Fig. 2. sFLT-1 transcripts were detected in the placental spongiotrophoblast cells of days 13 (Fig. 3E), 15 (Fig. 3H), and 17 (Fig. 3I). No hybridization signals were observed on day 11. Serial sections were also hybridized with the antisense FLT-1 probe. FLT-1 expression was observed in the spongiotrophoblast cells of days 11 (Fig. 3A), 13 (Fig. 3D), and 15 (Fig. 3G), but only weak signals were found on day 17. Serial sections hybridized with the sense sFLT-1 (Fig. 3B) and FLT-1 (data not shown) probes did not show signal above the background.

Northern Blot Demonstrating the Specificity of the Probes for FLT-1 (A), or sFLT-1 (B) Transcripts
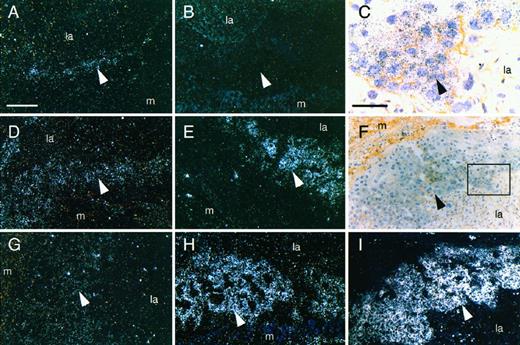
In Situ Hybridization Analysis of FLT-1 and sFLT-1 Expression in Placenta at Different Stages of Pregnancy At each stage, serial sections of the same placenta were used for a comparative analysis using specific FLT-1 and sFLT-1 probes. FLT-1 expression was detected in the spongiotrophoblast cells of days 11 (A), 13 (D), and 15 (G). sFLT-1 transcripts were localized in the spongiotrophoblast cells of days 13 (E), 15 (H), and 17 (I). F, Bright field image of panel I; C, higher magnification of panel F. B, Hybridization of serial sections with sense sFLT-1 probe did not produce any signal above the background. la, Labyrinthine layer; arrowhead, spongiotrophoblast layer; m, maternal layer. White scale bar, 200 μm (A, B, D, E, F, G, H, and I); black scale bar, 60 μm (C).
Total RNA from days 13, 15, or 17 placenta was also analyzed by Northern blot using a cDNA probe detecting both sFLT-1 and FLT-1 transcripts, and two bands were detected in each sample (Fig. 4). The level of sFLT-1 transcripts was much higher than that of FLT-1, and the ratio of sFLT-1 to FLT-1 changed markedly as gestation progressed. This result is consistent with the observation from in situ hybridization.
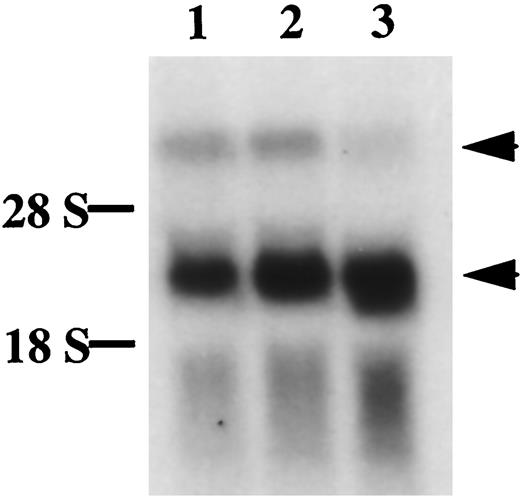
Northern Blot Analysis of Total RNAs from Placenta of Days 13 (lane 1), 15 (lane 2), and 17 (lane 3) Using a cDNA Probe for Detecting Both FLT-1 and sFLT-1 Transcripts Upper arrowhead, FLT-1 transcripts; lower arrowhead, sFLT-1 transcripts.
sFLT-1 Is Also Expressed in Adult Organs
Sections from adult lung, liver, kidney, and uterus were also analyzed by in situ hybridization, but no signals above background were detected. However, RT-PCR for 28 cycles using specific primers demonstrated that sFLT-1 transcripts could be detected in adult kidney, lung, and uterus (Fig. 5) and also in liver if 35 cycles were used (data not shown). The same amount of RNA without RT was used as control and no band was detected.
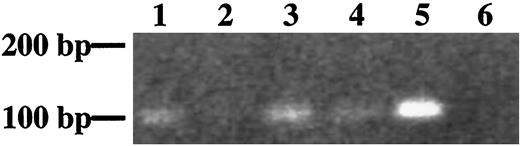
RT-PCR to Specifically Detect sFLT-1 mRNA in Different Adult Tissues Lane 1, kidney; lane 2, liver; lane 3, lung; lane 4, uterus; lane 5, placenta; lane 6, negative control.
Characterization of the Placental VEGF-Binding Factor
Serum-free conditioned medium of mouse placenta contains VEGF-binding activity. To determine whether this VEGF-binding activity is related to sFLT-1, a fraction of heparin-Sepharose partially purified protein from placenta- conditioned medium was further analyzed by Western blot. A protein band, approximately 111 kDa, was detected by antihuman FLT-1 antibody (Fig. 6, lane 2) and also by anti-FLT-1 N-terminal antibody (data not shown). This is in good agreement with Kendall et al. (27), who found that endothelial cells produced a sFLT-1 of 110 kDa. These authors (26) also showed that baculovirus-expressed sFLT-1 had an approximate molecular mass of 90 kDa. This is consistent with our results (Fig. 6, lane 1). The size difference between the endogenously encoded and recombinant sFLT-1 is probably due to the differential glycosylation of protein in mammalian and insect cells. In control immunoblots, rabbit IgG instead of FLT-1 antibody was used, and no band was detected (data not shown).
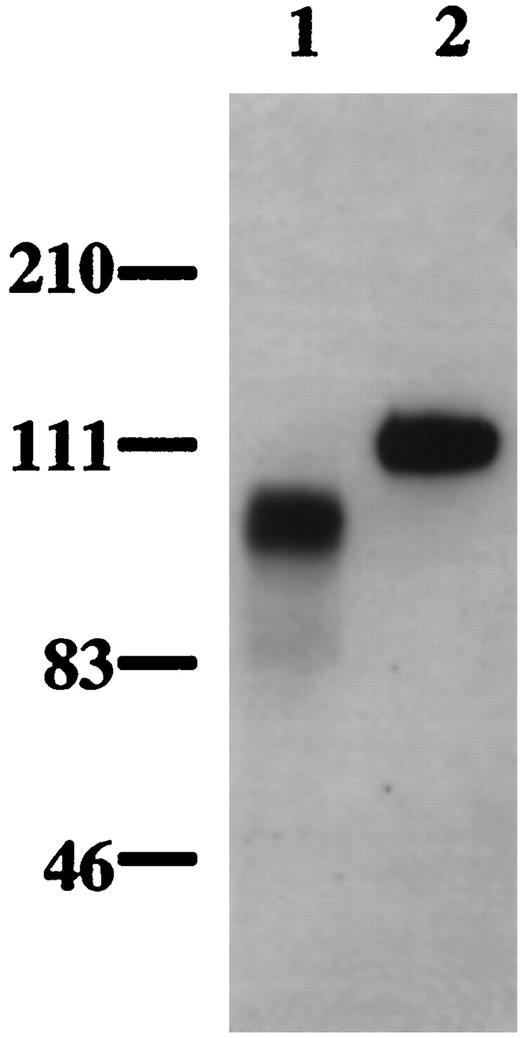
The VEGF-Binding Activity from Conditioned Medium of Mouse Placenta (lane 2) and Baculovirus-Expressed Human Recombinant sFLT-1 (lane 1) Were Separated by 4–12% Bis-Tris NuPAGE and Analyzed by a Western Blot Probed with the Biotinylated anti-Human FLT-1 Antibody
The VEGF-Binding Activity Increases in the Serum of Mice in Late Gestation
Since sFLT-1 transcripts became much more abundant as gestation progressed, we investigated whether sFLT-1 protein was detectable in the maternal circulation at this time. After incubation of serum with[ 125I]VEGF, complexes between [125I]VEGF and the VEGF-binding activity were formed and separated using a Sephacryl S-200 gel filtration column. Fractions eluted in PBS were collected and the radioactivity in each was determined. Serum from nonpregnant and day 11 pregnant mice produced a similar pattern of peaks (Fig. 7), with one small peak at fraction 10 and two additional peaks around fractions 14 and 28. However, serum from days 13, 15, or 17 pregnant mice showed a different peak pattern, with a major peak at fraction 10 (Fig. 7). The peak around fraction 28 remained in all samples. When 100-fold excess of unlabeled recombinant human VEGF was coincubated with serum from day 17 pregnant mice and [125I]VEGF, the major peak at fraction 10 disappeared, indicating specific binding between VEGF and the VEGF-binding activity. When serum from a nonpregnant mouse, to which recombinant human sFLT-1 had been added, was coincubated with[ 125I]VEGF, a major peak appeared at fraction 10, indicating the chromatographic similarity between recombinant sFLT-1 and the serum VEGF-binding factor. Incubation of[ 125I]VEGF with PBS also produced two peaks, one at fraction 14 and the other at 28. These correspond to the complexes of[ 125I]VEGF and BSA and free [125I]VEGF.[ 125I]VEGF is supplied with BSA carrier, and ligand blots show VEGF interacts weakly with BSA (data not shown).
![Sephacryl S-200 Gel Filtration Chromatography The peak at fraction 10 is the complexes of [125I]VEGF and the VEGF-binding activity, and the peak around fraction 14 is[ 125I]VEGF and BSA complexes. Free VEGF produces a peak around fraction 28. np, Serum from nonpregnant mice.](https://oup.silverchair-cdn.com/oup/backfile/Content_public/Journal/mend/13/4/10.1210_mend.13.4.0265/3/m_mg0490265007.jpeg?Expires=1716316350&Signature=jnegMCEcR~SmE~ereTcmlfwxTwl0ipGHNWQX5fGPy5GaDFa-X6KORdI5fPDhZHnKNeeVekHaPrdoOVmXhn~~5hzSoFEERtgvDktgRP43vzxCWRzwdK~xxYqtNLmAlSgcAZUzwkYTNAGdX73a0S73b8OFpZkHJWxNwbsMBpggMb2LUT687Os~a2F4wo6LLea5dDS3~4oJKqXwf1ScblRr9IrCle6AdbyJB3EdsZlz5hlFlFplgmmxws3kKUQmaR8G74ddDFVRAh~25~s4lP0dE6rHHXPOJyRkMrgjONS5EwxPuomudNck2G8hKU16MtmDYm8YzScBLo6AEVslsqz~Gg__&Key-Pair-Id=APKAIE5G5CRDK6RD3PGA)
Sephacryl S-200 Gel Filtration Chromatography The peak at fraction 10 is the complexes of [125I]VEGF and the VEGF-binding activity, and the peak around fraction 14 is[ 125I]VEGF and BSA complexes. Free VEGF produces a peak around fraction 28. np, Serum from nonpregnant mice.
Administration of Exogenous VEGF Led to Fibrin Deposition in the Placenta
The presence of a potent VEGF antagonist suggests that the action of this growth factor is regulated. We investigated this by perturbing the ratio of VEGF to sFLT-1 by administration of exogenous recombinant VEGF to pregnant mice. In animals treated with recombinant VEGF, there was an increase in the number of resorption sites. There were 18 resorption sites present in the treated mice (n = 5) and only 1 in the control mice (n = 4) (P < 0.05, Wilcoxon rank sum test). Embryos from these mice, which appeared macroscopically normal, weighed less than those from the control group (mean ± sd: control, 0.53 ± 0.05 g; treated, 0.46± 0.09 g P < 0.001, Student’s t test). There was no difference in the placental weight between these two groups. However, histological examination of placenta from these two groups did reveal differences. In the treated group there was an increase in fibrin deposition apparent within the labyrinthine layer (Fig. 8). The deposition was widespread as shown by the red/purple stain around many of the cells (Fig. 8, A and B). There were also occasional areas of intense staining (Fig. 8, C and D). Such staining was absent from the placentas of the vehicle-treated mice (Fig. 8, E and F).
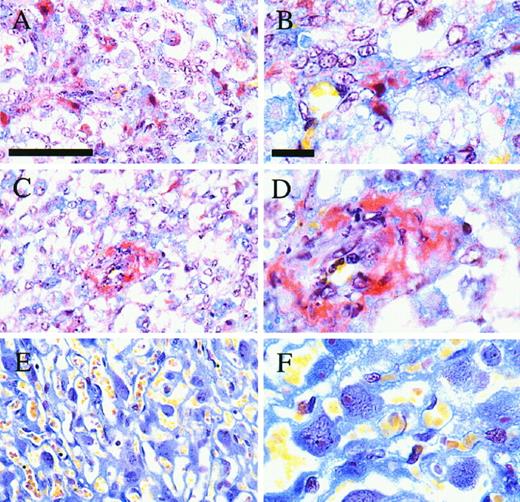
Histology of Mouse Placental Sections Stained with MSB to Reveal Fibrin Deposition Obtained from Mice Treated with VEGF (A, B, C, and D) or Vehicle (E and F) during Pregnancy Fibrin deposits are bright red, red blood cells are stained yellow, and other cells are stained blue. Scale bar, 50 μm in panels A, C, and E; 10 μm in panels B, D, and F.
DISCUSSION
In this study we have shown the stage-dependent expression of FLT-1 and sFLT-1 in placental spongiotrophoblast during pregnancy in the mouse. This suggests a novel mechanism for the regulation of VEGF activity by alternative splicing of FLT-1 pre-mRNA.
The mRNA encoding full-length membrane-bound FLT-1 was detected in placental spongiotrophoblast cells on day 11 of pregnancy, but sFLT-1 mRNA was undetectable by in situ hybridization at this time. However, high levels of sFLT-1 transcripts were observed on day 13 and rose as gestation progressed. In contrast, FLT-1 transcripts declined in late gestation such that by day 17 they were almost undetectable by in situ hybridization. This suggests that there may be regulation of the splice site selection in these cells leading to a marked shift in the ratio of their mature mRNAs. The change of the sFLT-1 to FLT-1 ratio was further confirmed by Northern blot analysis (Fig. 4). sFLT-1 was identified in conditioned medium of cultured mouse placenta. These results indicate that sFLT-1 is produced in the placenta. Thus, it is likely that the VEGF-binding activity found in the serum of mice in late pregnancy is sFLT-1. VEGF-binding activity was also detected in the serum of pregnant women (data not shown), suggesting that sFLT-1 may have a systemic role in antagonizing increasing VEGF during pregnancy (32). Recent data have confirmed the role of sFLT-1 as an inhibitor of angiogenesis. Intravitreal injection of soluble VEGF-receptor chimeric proteins is able to suppress retinal neovascularization in a murine model of ischemic retinopathy (28). Furthermore, treatment of super-ovulated rats with truncated soluble FLT-1 receptors resulted in the complete suppression of corpus luteal angiogenesis and a failure of endometrial maturation (29). These studies show that sFLT-1 is a potent antagonist of VEGF in vivo. Thus, it is likely that the physiological alternative splicing of FLT-1 pre-mRNA to generate FLT-1 and/or sFLT-1, described in this paper, will be important for the regulation of placental angiogenesis.
RT-PCR analysis showed that sFLT-1 mRNA is also present in adult lung, liver, kidney, and uterus, suggesting that sFLT-1 may have a role in maintaining endothelial cells in a quiescent state in the adult. sFLT-1 is also produced by human umbilical vein endothelial cells in vitro (27). Thus, a similar alternative splicing mechanism for FLT-1 pre-mRNA also exists in endothelial cells. Whether the FLT-1/sFLT-1 switch is involved in the modulation of pathological angiogenesis remains to be investigated.
It has been shown that the migration of monocytes/macrophages in response to VEGF is mediated by FLT-1 (33, 34). Since FLT-1 mRNA was detected in the trophoblast before day 13, it is possible that FLT-1 may regulate the migration of trophoblast cells.
The spongiotrophoblast cell layer where sFLT-1 transcripts are localized is located between the maternal and labyrinthine layers where VEGF is expressed (10, 11). The balance of the locally expressed VEGF and sFLT-1 could be important in the regulation of placental endothelial cell function. Administration of exogenous VEGF to pregnant mice led to fibrin deposition in the placenta, an increase in resorption sites, and a reduction in embryonic weight. This suggests that the exogenous VEGF circumvents the regulatory control of the angiogenic events in the placenta, and that a balance of angiogenic inducers and inhibitors is critical for normal placental function. The enhanced vascular permeability and the induction of tissue factor synthesis by endothelium and monocytes in response to exogenous VEGF (24, 25, 35) may contribute to fibrin deposition in the placenta of VEGF-treated mice. The placental abnormalities may account for the significant reduction of the embryo weight in these mice.
However, no abnormalities were observed in other organs of VEGF-treated mice (data not shown). This may be a reflection of the presence of locally acting inhibitors or phenotypic differences in mature endothelium. It is not known whether the exogenous VEGF crossed the placenta to reach the developing fetus, but no vascular defects were observed in the embryos from VEGF-treated mice. However, direct injection of exogenous VEGF into Quail embryos induced malformed and hyperfused vessels during embryonic neovascularization (36). These data suggest that developing organs are more sensitive to the change of balance of angiogenic inducers and inhibitors.
The expression of FLT-1 and/or sFLT-1 by alternative splicing of FLT-1 pre-mRNA in the spongiotrophoblast cells provides a means by which they can regulate the local response to VEGF. How the switch between FLT-1 and sFLT-1 is regulated needs to be further elucidated. Understanding this will provide a fuller understanding of physiological angiogenesis and may lead to novel means for the modulation of pathological angiogenesis.
MATERIALS AND METHODS
Tissue and Serum Collection
All procedures and care of animals were in accordance with the regulations laid down by the UK Home Office. BALB/c females (20–25 g) were mated with males of the same stock, and the day the vaginal plug was detected was designated day 1 of pregnancy. Once pregnant, the mice were housed individually. Placentas were collected on days 11, 13, 15, and 17 of pregnancy. Organs from adult mice (10–12 weeks old), including lung, liver, kidney, and uterus, were also collected. Tissues were fixed overnight in 10% formalin in PBS and processed for routine histology or snap-frozen in liquid nitrogen and stored at −70 C.
Twenty placentas from day 17 pregnant mice were minced, washed three times in DMEM/nut mix F-12 medium (GIBCO/BRL), and cultured in the same medium at 37 C in 5% CO2. Conditioned medium was harvested 24 h later and stored at -20 C for further analysis.
VEGF treatment commenced on day 9 of gestation and continued until day 17. Recombinant human VEGF (Amgen, endotoxin <0.006 pg/μg VEGF) was dissolved in PBS and 1.5 μg were injected ip into the pregnant mice (n = 5) daily. Control mice (n = 4) were treated with vehicle alone. At autopsy on day 18, the number of resorption sites per mouse was determined and the embryo and placenta were weighed. Tissue was collected and processed as above and 5-μm sections were cut and stained for fibrin using the specific histological stain Martius Scarlet Blue (MSB) (37) in which fibrin is stained bright red.
Blood from normal pregnant and nonpregnant mice was also collected. Serum was recovered and stored at −70 C until required for analysis.
3′-RACE
Total RNA was isolated from placenta of day 17 pregnant mice using the method of Chomczynski and Sacchi (38). Poly (A)+ RNA was purified using oligo (dT) cellulose (Pharmacia, Piscataway, NJ) and cDNA was prepared using (dT) 17-adaptor primer and Super RT (HT Biotechnology Ltd, Cambridge, UK) following the manufacture’s instructions. The (dT)17-adaptor primer (GACTCGAGTCGACATCGATTTTTTTTTTTTTTTTT) was according to Frohman et al. (39). A portion of this cDNA was used to amplify the desired 3′-end sequence of mouse sFLT-1 using a gene-specific primer (GSP: GCCAGGAACATATACACAG) corresponding to bases 1941–1959 of mouse FLT-1 (40), and an adaptor primer (GACTCGAGTCGACATCGA), with a HYBAID Touchdown thermocycler in 20 μl of PCR cocktail comprising deoxynucleoside triphosphates (200 μm), 1× BioTaq polymerase buffer, Mg++ solution (1.5 mm), 1 U of BioTaq DNA polymerase (Bioline, London, UK). The reaction mixture was denatured at 95 C for 2 min, and then amplified for 35 cycles (95 C, 30 sec; 52 C, 1 min; 72 C, 40 sec), followed by a 3-min final extension at 72 C. After end repair, the PCR products were cloned in pCR-Script Amp SK (+) cloning vector (Stratagene, La Jolla, CA) and sequenced.
Probes
To generate probes for the specific detection of sFLT-1 or FLT-1 transcripts, or both, RT-PCR was performed to produce cDNA fragments using specific primers. A 105-bp long 3′-end cDNA fragment of mouse sFLT-1, which starts from the divergent site of FLT-1 and sFLT-1, was generated by RT-PCR using mRNA extracted from placenta of day 17 pregnant mice. The primers used were based on the unique 3′-end nucleotide sequence of mouse sFLT-1 obtained as described above. The 5′-primer was AGG TGA GCA CTG CGG CA (msflt-1), and the 3′-primer was ATG AGT CCT TTA ATG TTT GAC (msflt-2). A cDNA fragment corresponding to bases 3799–4011 of mouse FLT-1 nucleotide sequence (40), which showed little similarity to the other members of the receptor tyrosine kinase family, was also generated by RT-PCR as described above. The 5′-primer was (3799) TCA CCT GGA CTG AGA CCA AG (3819) and the 3′-primer was (3990) GTA CAA CAC CAC GGA GTT GTA (4011). Another cDNA fragment corresponding to bases 11–210 of mouse FLT-1 nucleotide sequence (40), which detects both sFLT-1 and FLT-1 transcripts, was also generated as described above. The 5′-primer was 11 CCG CGT CTT GCT CAC CAT G 29, and the 3′-primer was 210 ACC ATG AGT GGG CTG CCT C 192. cDNA fragments generated above were cloned in pCR-Script Amp SK(+) vector and sequenced.
In Situ Hybridization
In situ hybridization was carried out essentially as described by Clark et al. (31). Hybridization probes used were specific for detecting sFLT-1 or membrane-bound FLT-1 transcripts. This was confirmed by Northern blot analysis of total RNA from day 15 placenta (Fig. 2, A and B). To generate RNA probes for in situ hybridization, the constructs described above were linearized by digestion with the appropriate restriction endonuclease, and 33P-UTP (Amersham International PLC, Little Chalfont, U.K.)-labeled sense and antisense RNA probes were synthesized by in vitro transcription using T7- and T3-polymerases respectively (Ambion, Inc., Austin, TX).
Northern Blot Analysis
Total RNA (20 μg) was separated in a 1% (wt:vol) agarose/6% formaldehyde (wt:vol) gel prepared in 1× MEA buffer (20 mm 3-(N-morpholino)propane sulfonic acid, 5 mm NaAc, 1 mm EDTA, pH 7.O), transferred to nylon membrane (Amersham) by capillary blot, and fixed by UV cross-linking. The blot was then prehybridized in 50 ml of hybridization buffer comprising 50 mm Tris-HCl (pH 7.6), 0.1% (wt/vol) SDS, 10× Denhardts [0.2% (wt/vol) Ficoll, 0.2% (wt/vol) PVP, 0.2% (wt/vol) BSA], 0.1% sodium pyrophosphate (wt/vol), 6% polyethylene glycol (PEG 6000) (wt/vol), 6% NaCl (wt/vol), and 0.1 mg/ml salmon sperm DNA at 65 C for 2 h. Probes specific for sFLT-1 or FLT-1, or both transcripts, were prepared using cDNA fragments generated above and T7 QuickPrime Kit (Pharmacia Biotech) following the manufacturer’s instructions. Probe was labeled to a specific activity of 1–2 × 106 cpm/ng using [a32P] dCTP (Amersham). Before addition to the hybridization solution, the probe was denatured by boiling for 3 min. The incubation was continued at 65 C overnight. The blot was then washed in 1× SSC/0.1% (wt/vol) SDS at 65 C with three changes of washing buffer and wrapped in Saran wrap for autoradiography. The exposure was performed at −70 C overnight using double-coated x-ray film (Fuji Medical Systems, Stamford, CT) in conjunction with intensifying screens. The film was developed in an x-ograph automatic film developer.
RT-PCR
Extraction of total RNA from lung, liver, kidney, and uterus and preparation of cDNA were performed as already described. Similar amounts of RNA from each sample were used for RT and amplification. The specific sFLT-1 primers were msflt-1 and msflt-2 described above. The PCR program was as follows: 95 C for 1 min; 28 cycles (95 C, 30 sec; 54 C, 30 sec; 72 C, 30 sec); followed by a 3-min final extension at 72 C. RNA without RT was used as negative control.
Western Blot Analysis
Human sFLT-1 has been shown to bind heparin Sepharose (26). We therefore used this affinity matrix to partially purify the VEGF-binding activity in mouse placenta-conditioned medium. Conditioned medium from 20 mouse placentas (15 ml) was centrifuged and then loaded onto a HiTrap heparin 1-ml column (Pharmacia) equilibrated in PBS. The column was washed with 5 ml of PBS and 5 ml of 0.6 m NaCl/20 mm phosphate buffer (pH 7.4), and the activity was eluted with 1.2 m NaCl in the same buffer. The elute was then concentrated using Centricon-30 (Amicon, Inc., Beverly, MA) and washed in PBS. A portion of this was electrophoretically separated by 4–12% Bis-Tris NuPAGE (Novex, San Diego, CA) and transferred to nitrocellulose membrane (Amersham). The membrane was blocked with 5% BSA in PBS/0.1% Tween 20/0.1 m NaCl and probed with biotinylated anti-human FLT-1 antibody (0.15 μg/ml) (R&D Systems, Minneapolis, MN) in the same buffer containing 1% BSA. Immunoreactive bands were detected using streptavidin-horseradish peroxidase (Amersham) and Supersignal substrate (Pierce, Rockford, IL).
S-200 Gel Filtration Chromatography
Serum (50 μl) from nonpregnant and pregnant mice on days 11, 13, 15, and 17 of gestation (n = 3 for each stage) was incubated with 0.7 ng of 125I-labeled human VEGF (2300 Ci/mmol, Amersham), respectively, at room temperature overnight. Day 17 pregnant mouse serum in the presence of 100-fold excess of unlabeled recombinant human VEGF, or nonpregnant mouse serum in the presence of baculovirus-expressed recombinant human sFLT-1 (1 ng), was also incubated with the same amount of [125I]VEGF. Samples were analyzed by loading onto a 12.5-ml Sephacryl S-200 (Sigma Chemical Co., St. Louis, MO) gel column preequilibrated in PBS as described by Hill et al. (41). Forty fractions eluted in PBS, each of 460μ l, were collected, and the radioactivity of each fraction was counted using a γ-counter (Packard Instruments, Meriden, CT).
Acknowledgments
We thank Dr. Andrew Sharkey for helpful discussion of this work and Ms. Amanda Evans for her assistance with DNA sequencing.
D.S.C.J. was supported in part by Biotechnology and Biological Sciences Research Council (Fellowship PDF/22).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}