





Abstract
Based on the known capacity of deoxynivalenol (DON) to target gut lymphoid tissue and IgA production, it was hypothesized that this mycotoxin interferes with the immune response to enteric reovirus infection. When mice were orally gavaged, first with 25 mg/kg bw DON, and then with reovirus serotype 1, strain Lang (T1/L) 2 or 12 h later, viral titers in the GI tract were 10-fold higher than control mice after 5 days. Virus was almost completely cleared in both treatment and control groups from intestinal tissue after 10 days. Real-time PCR indicated that, in infected control mice, reovirus λ2 core spike (L2 gene) RNA per g feces in infected mice that were pretreated with DON was significantly higher at 1, 3, and 5 days than in infected mice only. In reovirus-infected mice, DON at doses of 10 and 25 mg/kg bw but not 2 and 5 mg/kg bw increased fecal L2 RNA, whereas DON doses as low as 2 mg/kg potentiated L2 RNA levels in Peyer's patches (PP). Reovirus-specific IgA levels in feces of mice treated with DON were significantly elevated, as were specific IgA responses in lamina propria and PP fragment cultures. Similar effects were observed for serum IgA and IgG. DON suppressed IFN-γ responses in PP to reovirus at 3 and 5 days as compared to infected controls, while IL-2 mRNA concentrations were unaffected. Although reovirus alone did not induce Th2 cytokine mRNAs in PP, DON exposure significantly elevated IL-4, IL-6, and IL-10 mRNA expression at various times during the infection. ELISPOT revealed that mRNA expression data corresponded to suppression of IFN-γ- and enhancement of IL-4-producing cell responses in PP cultures from DON-treated mice. Taken together, these data suggest that DON transiently increased both severity of the reovirus infection and shedding in feces as well as elevated reovirus IgA responses. These effects corresponded to suppressed Th1 and enhanced Th2 cytokine expression.
Deoxynivalenol (DON or “vomitoxin”) is a trichothecene mycotoxin that commonly contaminates wheat, corn, and barley infected by the fungus Fusarium graminearum (Bai and Shaner, 2004). DON is resistant to milling, processing, and heating and, therefore, readily enters human food (Hazel and Patel, 2004; Jackson and Bullerman, 1999). This toxin has been associated worldwide with gastroenteritis as reflected by nausea, emesis, diarrhea, anorexia, and gastrointestinal (GI) hemorrhaging (Canady et al., 2001), making DON an important human health concern (Pestka and Smolinski, 2005). DON and other trichothecenes activate mitogen-activated protein kinases (MAPKs) in vitro and in vivo via a mechanism known as the ribotoxic stress response (Chung et al., 2003; Moon and Pestka, 2002, 2003; Zhou et al., 2003a,b). Leukocytes are extraordinarily sensitive to DON's effects and can respond to low toxin concentrations by upregulating immune- and inflammation-related genes and to high toxin concentrations by evoking apoptosis (Pestka et al., 2004).
The potential for trichothecene mycotoxins to alter human immune responses remains unknown. However, trichothecenes enhance or suppress systemic responses to experimental infections with Salmonella (Bottex et al., 1990; Kubena et al., 2001; Tai and Pestka, 1990; Ziprin and Elissalde, 1990), Listeria (Ziprin et al., 1987; Ziprin and McMurray, 1988), Staphylococcus (Cooray and Jonsson, 1990), and herpes simplex (Friend et al., 1983). Prolonged consumption of DON by mice induces elevated serum IgA and IgA immune complexes, kidney mesangial IgA deposition, and hematuria which are clinical signs of human IgA nephropathy (IgAN) (Pestka, 2003). DON-induced upregulation of IL-6 appears to be critical to DON-induced IgAN, based its robust induction in vivo (Zhou et al., 1997, 1998, 2003b), as well as studies employing IL-6 deficient mice (Pestka and Zhou, 2000), antibody neutralization (Yan et al., 1997), and ex vivo cultures (Yan et al., 1998). In addition, exposure to lipopolysaccharide potentiates DON-induced inflammation and cytotoxicity in the mucosal and systemic compartments (Islam et al., 2002; 2003; Islam and Pestka, 2003; Zhou et al., 1999, 2000). Although, DON's reported effects on the GI tract and IgA production suggest that this toxin might interfere with the immune response to enteric viruses, this possibility has not been systematically addressed.
Reovirus (respiratory enteric orphan virus) is a double-stranded RNA virus that has been isolated from the GI and respiratory tracts of both humans and animals (Nibert et al., 1996). Enteric reovirus infection produces a self-limited infection in which the virus is cleared from gastrointestinal tract within 7 to 14 days after onset (Barkon et al., 1996; Rubin et al., 1985). Mucosal immune responses to enteric reovirus infection have been well characterized with regards to induction of virus-specific intestinal IgA (London et al., 1987; Major and Cuff, 1996; Silvey et al., 2001) and serum IgG (Major and Cuff, 1996; Virgin et al., 1988), as well as cell-mediated immune responses (Cuff et al., 1993; London et al., 1987) and cytokine production (Fan et al., 1998; Mathers and Cuff, 2004) in gastrointestinal-associated lymphoid tissues (GALT) of mice. Notably, Cuff et al. (1998) successfully used enteric reovirus infection as a tool to assess the immunotoxic effects of cyclophosphamide on GI immune system, suggesting that this virus is an excellent laboratory model for investigating intestinal immune responses.
The objective of this research was to test the hypothesis that DON exposure modulates the immune response to enteric reovirus. The results indicated that DON impaired reovirus clearance but enhanced reovirus-specific immunoglobulin responses, which correlated with suppressed Th1 and enhanced Th2 cytokine responses, respectively.
MATERIALS AND METHODS
Mice.
Female B6C3F1 mice aged 7–10 weeks were obtained from Charles River. Mice were housed in laminar-flow microisolator cages with negative pressure and acclimated for 1 week prior to use in experiments at the MSU University Research Containment Facility in a BSL 2 room with humidity and temperature control. All animal studies followed NIH guidelines and were approved by the Michigan State University Committee on Animal Use and Care.
Virus.
Reovirus serotype 1, strain Lang (T1/L) was used for all experiments. The virus was grown in L929 fibroblast cells at 34°C in DMEM medium (Sigma) with 5% (v/v) fetal calf serum (Sigma), 100 U/ml of penicillin, 100 μg/ml of streptomycin (Sigma), and 0.25 μg/ml of amphotericin B (GIBCO). The third-passage plaque-purified virion (Fulong et al., 1988) was used for mouse infection and prepared by extraction with 1,1,2-trichloro-1,2,2-trifluoroethane (Sigma), followed by discontinuous CsCl gradient centrifugation as previously described (Silvey et al., 2001). Titers of the purified virus were determined by plaque assay (Cuff et al., 1990).
Experimental design and sample collection.
DON, purified as described previously by Clifford et al. (2003), was dissolved in endotoxin-free water. Mice were treated once with 2 to 25 mg/kg bw DON po. The highest dose represents approximately one third the LD50 and was selected based on its demonstrated capacity to modulate gene expression in mice (Zhou et al., 1997). After 2 or 12 h, animals were infected by oral gavage with 3 × 107 plaque forming units (PFU) of reovirus in a total volume of 100 μl borate-buffered saline (pH 7.4) containing 2% (w/v) of gelatin (London et al., 1987). At specified time intervals, fecal pellets were collected and/or mice were bled from lateral saphenous vein (Hem et al., 1998). At experiment termination, mice were anesthetized with metaflurane, bled from retroorbital plexus, and immediately euthanized by cervical dislocation. Serum was separated from clotted blood samples and stored at 4°C for later analysis. Fecal pellets were collected at intervals after infection and weighed. The material was suspended at 0.1 g/ml in phosphate-buffered saline, held on ice for 2 h, and then sonicated for 15 s. Solutions were cleared by high speed centrifugation for 10 min at 4°C. Supernatants were used directly for specific antibody detection by ELISA and for total RNA purification. Intestines and spleens were procured from euthanized mice and processed as described below.
Lymphoid fragment cultures.
The lymphoid fragment culture system of Logan et al. (1991), as modified by Cuff et al. (1998), was used to study immunoglobulin secretion. Briefly, seven Peyer's patches (PP) per mouse were removed from intestine, pooled, and washed three times in HBSS with 5 μg/ml of gentamicin and twice with tissue culture medium (TCM) consisting of RPMI 1640 medium supplemented with 10% (v/v) FBS, 2 mM L-glutamine, 0.5 μM 2-mercaptoethanol, 5 μg/ml of gentamicin, 100 unit/ml penicillin, and 100 μg/ml streptomycin. Individual patches were cut in half and incubated collectively in 2 ml of TCM for 5 days at 37°C under 5% CO2 without additional stimulation. Lamina propria (LP) cultures were established, after removing PP, by splitting intestine longitudinally and cutting it into pieces 2–3 cm long. The fragments were washed at least three times with HBSS with 0.2% NaHCO3, 0.1M HEPES, and 5 μg/ml gentamicin. One half of the fragments were used for viral titer detection, and the other half were incubated twice for 30 min in 5 mM EDTA in HBSS to remove epithelial cells. Fragments were washed twice more and then incubated in 3 ml of TCM for 5 days at 37°C under 5% CO2. Culture supernatants were harvested and stored at 4°C for antibody detection by ELISA.
Virus titration.
For viral plaque counts of GI tract, intestinal segments were placed in 2 ml of sterile phosphate-buffered saline, freeze-thawed three times, and sonicated. Virus titers were measured by standard viral plaque assay (Major and Cuff, 1997). Briefly, tissues were homogenized in 3 ml saline containing 0.5% gelatin. Serial dilutions (100 μl) in TCM were incubated on monolayers of L-929 fibroblasts in 12-well tissue culture plates for 45 min at 34°C and thereafter overlaid with 3 ml of 1% agar in Medium 199 (Sigma) and cultured at 34°C. Plaques were counted after 7 days incubation.
Real-time PCR for reovirus.
For fecal pellet analyses, total RNA was extracted from 200 μl of supernatant of 10% (w/v) fecal suspension using TRIZOL (Invitrogen). PCR primers were selected from published sequence of λ2 core spike (L2 gene) of reovirus T1/L (Breun et al., 2001) as follows: forward, 5′ ctg acg tcg atc agg tcg ttg 3′ and reverse, 5′ gat gtg gca tgc atg cat gag 3′. The size of the expected amplicon was 97 bp. To denature dsRNA, 1 μg of the template was incubated for 5 min at 95°C with 250 pM of random primers (Promega) or L2 forward primer in total volume of 12 μl and then snap cooled on ice. Reverse transcriptase reaction was performed by adding 4 μl of 5× RT reaction buffer, 2 μl of 0.1M DTT, 1 μl of 10mM dNTP, and 1 μl of Superscript II reverse transcriptase (Invitrogen) at 42°C for 60 min. Purified reoviruses were added into 10% (w/w) of fecal pellet suspension from vehicle mice at concentrations of 0, 101, 102, 103, 104,105 106, 107 PFU/ml as standard sample for standard curve and sensitivity determination.
ELISA.
Serum, culture fluid, and supernatant of fecal suspension were assayed for virus-specific antibody by ELISA by a modification of the procedure of Major and Cuff (1997). Briefly, 96-well plates were coated overnight at 4°C with 50 μl /well of purified reovirus T1/L, diluted to 1 × 108 particles/ml in 0.1 M NaHCO3 (pH 9.6). Plates were washed four times with PBS with 0.1% Tween 20 (PBS-T) and blocked with 100 μl of blocking buffer (3% [w/v] bovine serum albumin [BSA] in PBS-T) for 2 h at room temperature. Serial dilutions (100 μl) of serum, culture fluids, or fecal supernatants were added to each well in duplicate and incubated at 4°C for 16 h. The plates were washed seven times with PBS-T, and 100 μl of goat anti-mouse IgG2A-, IgG1-, or IgA-HRP conjugates (1:3000) in blocking buffer was added to each well. Plates then were incubated at room temperature for 2 h. Following seven washes with PBS-T, 100 μl of K-Blue Max TMB substrate (Neogen) was added to each well, and plates were incubated at room temperature for 5 min. Reaction was terminated by adding 50 μl of 2 N H2SO4, and absorbance was read at 450 nm on ELISA reader. Absorbance 450 was used as endpoint for fecal and fragment cultures. For sera, titers were designated as the highest serum dilution that yielded absorbances of 0.2 or higher; the geometric mean antibody titer was then calculated.
Real-time PCR for cytokines.
For cytokine PCR, PPs were collected and immediately put into RNAlater reagent (Ambion) at 4°C. Total RNAs were isolated with TRIZOL (Invitrogen). Primer sequences for amplification of cytokines (Table 1) were designed according to the sequences of cytokine mRNA from Genebank and published data (Overbergh et al., 2003; Samuel and Knutson, 1983). RNA (1 μg) for amplification of cytokines was denatured by incubation at 70°C for 10 min with 250 pM Oligo (dT)12–15 primer (Invitrogen). Real-time PCR was performed as previously described (Kinser et al., 2004) on an ABI Prism 7700 Sequence Detector with ABI SYBR Green PCR core kit (Applied Biosystem). Reactions contained a total volume of 12.5 μl: 1 μM primer pair, 3 mM Mg+2, 0.8 mM dNTP mixture, 6 ng cDNA template, 0.075 μl Taq Gold DNA polymerase (ABI). Reactions were started by incubation at 95°C for 5 min and followed by 40 two-step thermal cycles of 15 s denaturation at 95°C, 60 s primer annealing and extension at 60°C. Real-time measurements were taken, and a threshold cycle (Ct) value for each sample was calculated. Three negative controls and known positive standard curve samples ranging from 101 to 108 copies/ml of target gene were used for each run. All samples were analyzed in duplicate.
Primer Sequences for Real-Time PCR Amplification
Gene | Genbank No. | Forward primer | Reverse primer | PCR product length (bp) |
|---|---|---|---|---|
| IL-4 | M25892 | 5′aca gga gaa ggg acg cca t3′ | 5′gaa gcc cta cag acg agc tca3′ | 95 |
| IL-10 | M37897 | 5′tgt gaa aat aag agc aag gca gtg3′ | 5′cat tca tgg cct tgt aga cac c3′ | 85 |
| IFN-γ | K00083 | 5′gag gaa ctg gca aaa gga tgg tga3′ | 5′tgt tgt tgc tga tgg cct gat tgt3′ | 97 |
| IL-2 | X01772 | 5′cct gag cag gat gga gaa tta ca3′ | 5′ tcc aga aca tgc cgc aga g3′ | 141 |
| IL-6 | X45452 | 5′gag gat acc act ccc aac aga cc3′ | 5′ aag tgc atc atc gtt gtt cat aca3′ | 141 |
| TGF-β | NM000660 | 5′ gga gga ggg acg agc tgg ttg aga 3′ | 5′ cga gac agc gca gtg cca aga agt | 97 |
Gene | Genbank No. | Forward primer | Reverse primer | PCR product length (bp) |
|---|---|---|---|---|
| IL-4 | M25892 | 5′aca gga gaa ggg acg cca t3′ | 5′gaa gcc cta cag acg agc tca3′ | 95 |
| IL-10 | M37897 | 5′tgt gaa aat aag agc aag gca gtg3′ | 5′cat tca tgg cct tgt aga cac c3′ | 85 |
| IFN-γ | K00083 | 5′gag gaa ctg gca aaa gga tgg tga3′ | 5′tgt tgt tgc tga tgg cct gat tgt3′ | 97 |
| IL-2 | X01772 | 5′cct gag cag gat gga gaa tta ca3′ | 5′ tcc aga aca tgc cgc aga g3′ | 141 |
| IL-6 | X45452 | 5′gag gat acc act ccc aac aga cc3′ | 5′ aag tgc atc atc gtt gtt cat aca3′ | 141 |
| TGF-β | NM000660 | 5′ gga gga ggg acg agc tgg ttg aga 3′ | 5′ cga gac agc gca gtg cca aga agt | 97 |
Primer Sequences for Real-Time PCR Amplification
Gene | Genbank No. | Forward primer | Reverse primer | PCR product length (bp) |
|---|---|---|---|---|
| IL-4 | M25892 | 5′aca gga gaa ggg acg cca t3′ | 5′gaa gcc cta cag acg agc tca3′ | 95 |
| IL-10 | M37897 | 5′tgt gaa aat aag agc aag gca gtg3′ | 5′cat tca tgg cct tgt aga cac c3′ | 85 |
| IFN-γ | K00083 | 5′gag gaa ctg gca aaa gga tgg tga3′ | 5′tgt tgt tgc tga tgg cct gat tgt3′ | 97 |
| IL-2 | X01772 | 5′cct gag cag gat gga gaa tta ca3′ | 5′ tcc aga aca tgc cgc aga g3′ | 141 |
| IL-6 | X45452 | 5′gag gat acc act ccc aac aga cc3′ | 5′ aag tgc atc atc gtt gtt cat aca3′ | 141 |
| TGF-β | NM000660 | 5′ gga gga ggg acg agc tgg ttg aga 3′ | 5′ cga gac agc gca gtg cca aga agt | 97 |
Gene | Genbank No. | Forward primer | Reverse primer | PCR product length (bp) |
|---|---|---|---|---|
| IL-4 | M25892 | 5′aca gga gaa ggg acg cca t3′ | 5′gaa gcc cta cag acg agc tca3′ | 95 |
| IL-10 | M37897 | 5′tgt gaa aat aag agc aag gca gtg3′ | 5′cat tca tgg cct tgt aga cac c3′ | 85 |
| IFN-γ | K00083 | 5′gag gaa ctg gca aaa gga tgg tga3′ | 5′tgt tgt tgc tga tgg cct gat tgt3′ | 97 |
| IL-2 | X01772 | 5′cct gag cag gat gga gaa tta ca3′ | 5′ tcc aga aca tgc cgc aga g3′ | 141 |
| IL-6 | X45452 | 5′gag gat acc act ccc aac aga cc3′ | 5′ aag tgc atc atc gtt gtt cat aca3′ | 141 |
| TGF-β | NM000660 | 5′ gga gga ggg acg agc tgg ttg aga 3′ | 5′ cga gac agc gca gtg cca aga agt | 97 |
ELISPOT for IFN-γ- and IL-4-producing cells.
Mice were treated with 25 mg/kg of DON for 2 h, then inoculated with 3 × 107 of purified reovirus. After 4 days, PP were removed, and lymphocytes dissociated by passage through a cell sieve. Single-cell suspensions were passed through nylon wool membrane to remove accessory B cells and enrich for T cells. Recovered cells were restimulated by infection of reovirus with one multiplicity of infection (M.O.I.) unit for 1 h in complete DMEM medium. Cytokine-producing cells were measured using R&D System ELISpot Kits (Minneapolis MN) according to manufacturer's instructions. Briefly, after blocking and washing plates, 5 × 105 infected PP cells were incubated in 100 μl at 37 °C for 36 h. Plates were washed, developed with peroxidase-labeled second antibody and substrate, and dried for 2 h at room temperature. Resultant spots were counted under stereoscope and reported as the mean number of spot forming cells (SFC) per 106 PP lymphocytes. Data from two independent experiments were pooled (n = 12) for statistical analyses.
Statistics.
Data were analyzed using Sigma Stat software (Jandel Scientific, San Rafael, CA). For comparisons of two groups of data, Student's t-test was performed. For comparison of multiple groups of data, a Kruskal-Wallis one-way ANOVA on ranks was used. Data sets were considered significantly different when p < 0.05.
RESULTS
DON Increases Reovirus Burden in Intestinal Tissue
The capacity of DON-treated mice to modulate reovirus intestinal infection was initially assessed. Mice were treated with 25 mg/kg bw DON and then infected with reovirus after 2 or 12 h. Viral load in the intestines increased to approximately 104 PFU/intestine in infected control mice after 5 days but was nearly completely cleared by 10 days (Fig. 1). In both groups of mice treated with DON, viral titers were 10-fold higher at day 5 than control mice, but by day 10 were not significantly different. These results suggested that the host response to reovirus was transiently suppressed by DON, thereby increasing the intestinal burden, but recovered after 10 days.
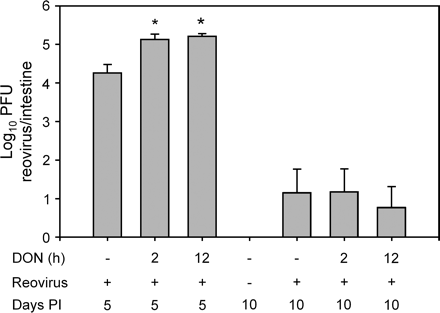
DON alters intestinal clearance of reovirus. Mice were treated with 0 or 25 mg/kg DON by oral gavage. After 2 or 12 h, mice were infected with 3 × 107 PFU of reovirus by oral gavage. Virus titers were determined by plaque assay 5 and 10 days after infection. Data are means ± SEM (n = 9). Bars with asterisk are significantly different from infected control at same timepoint by t-test (p > 0.05).
DON Increases Reovirus Shedding in Feces
Reovirus is shed in feces during intestinal infection, thus providing a convenient marker for monitoring effects of DON on host resistance to the virus. A real-time PCR method was devised for detection of reovirus λ2 core spike (L2 gene) RNA in fecal pellets. Agarose electrophoresis indicated that primers yielded a single 90 bp band corresponding to cDNA for this gene in fecal samples from control mice spiked with 800, 80, 8, 0.8 and 0.08 PFU/reaction, whereas no bands were generated in unspiked fecal samples or samples containing 0.008 PFU/reaction (Fig. 2A). A linear relationship existed between Ct values determined by real-time PCR and log viral titer (Fig. 2B), suggesting that this assay was sufficiently robust for measuring reovirus shedding in experimental mice. In a time-course study employing this method, 4 × 104 copies of L2 RNA/g feces were detectable at day 3 in infected control mice, but the gene was undetectable at 1, 5, and 7 days (Fig. 2C). In mice treated with DON for 2 h prior to infection, L2 RNA copies were significantly increased at 1, 3, and 5 days, with 3-day values exceeding infected controls by 6-fold. Similar results were found for mice treated with DON 12 h before reovirus infection (data not shown). These data indicate that, consistent with PFU measurements, markedly increased numbers of reovirus were shed into the intestine of DON-treated mice.
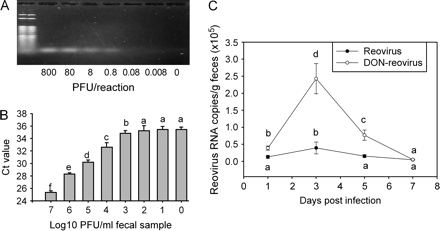
DON potentiates reovirus load in feces. For verification of specificity and sensitivity of real-time PCR for detection of reovirus L2 gene, purified reoviruses were added to supernatant of control mouse fecal pellet to PBS homogenate (1:10) at concentrations of 0 to 107 PFU/ml. Total RNAs were purified and L2 gene detected by real-time PCR. (A) PCR products after agarose gel electrophoresis and staining with ethidium bromide. (B) Relationship of virus concentration to threshold cycle (Ct) in real-time PCR. Data are means ± SEM (n = 6). Bars without same letter are significantly different (p < 0.05). (C) Mice were treated with 0 or 25 mg/kg DON by oral gavage for 2 h. Mice were infected with 3 × 107 PFU of reovirus po. Fecal pellets were collected at intervals and homogenized. Total RNAs were isolated and L2 gene detected by real-time PCR. Data are means ± SEM (n = 6) of viral L2 gene copies per gram fecal pellet. Points without same letter are significantly different (p < 0.05).
The dose-response effects of DON on L2 RNA in feces and PP of reovirus-infected mice were assessed at 4 days. DON at 10 and 25 mg/kg bw, but not 2 and 5 mg/kg bw, potentiated L2 RNA concentrations in fecal pellets (Fig. 3). However, L2 RNA levels in PP were significantly elevated in infected mice treated with DON at 2, 5, 10, and 25 mg/kg compared to infected control mice.
Dose-response effects of DON on reovirus L2 RNA in feces and Peyer's patches. Mice were treated with various doses of DON po and then, 2 h later, with 3 × 107 reovirus po. After 4 days, total fecal pellet and PP RNA were purified 4 days, and L-2 RNA was quantified by real-time PCR. Data are mean ± SEM (n = 6). Bars without same letter, differ significantly (p < 0.05).
DON Enhances Reovirus-Specific IgA Responses in Gut Mucosa
DON has previously been shown to alter regulation of the immunoglobulin response to intestinal antigens and induce polyclonal IgA synthesis in mice (Bondy and Pestka, 1991; Pestka et al., 1990a; Rasooly and Pestka, 1992; Rasooly et al., 1994). The capacity of DON to modulate the specific IgA response to reovirus in the gut mucosal compartment was therefore assessed by measuring fecal IgA with ELISA. Reovirus-specific IgA responses in infected controls were significantly elevated over uninfected controls at 8 and 10 days (Fig. 4). Reovirus-specific IgA levels in mice treated with DON 2 and 12 h prior to infection were slightly lower than infected or uninfected controls at 2 and 4 days but were markedly higher than infected controls at 8 and 10 days. Infection of control mice also increased reovirus-specific IgA in ex vivo LP fragment cultures at 5 and 10 days (Fig. 5). DON exposure of mice 2 h prior to reovirus infection significantly increased the IgA responses in LP cultures at 10 days and in PP cultures at 5 and 10 days. Thus, both in vivo and ex vivo data indicated that DON enhanced the mucosal IgA response to reovirus.
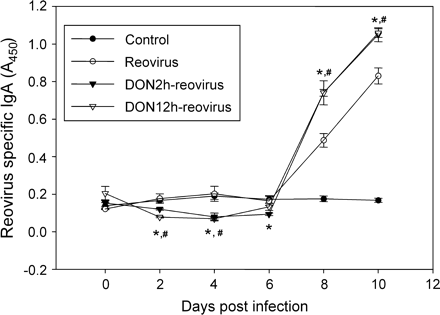
DON enhances reovirus-specific IgA response in feces. Mice were treated with 0 or 25 mg/kg of DON by oral gavage for 2 or 12 h and then orally infected with 3 ×107 PFU of reovirus. Fecal pellets were collected at 0, 2, 4, 6, 8, and 10 days after infection. Virus-specific IgA in fecal suspensions was determined by ELISA. Data are means ± SEM. Points marked with * and # indicate significant difference between control vehicle from groups given DON for 2 and 12 h, respectively (p < 0.05).
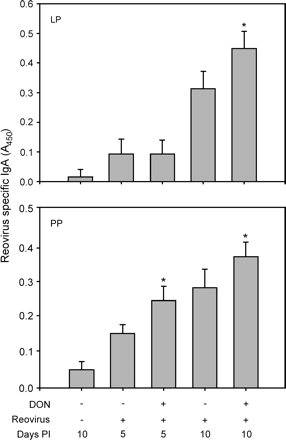
Ex vivo production of reovirus-specific IgA is potentiated in laminia propria (LP) and Peyer's patch (PP) cultures from DON-treated mice. Mice were treated with 0 or 25 mg/kg of DON by oral gavage for 2 or 12 h and then infected with 3 × 107 PFU of reovirus. LP and PP fragment cultures were incubated for 5 days without additional stimulation. Reovirus-specific IgA in cultured supernatants was determined by ELISA. Data are means ± SEM (n = 9). Bars marked with * are significantly different from vehicle control at corresponding time point by t-test.
DON Potentiates Reovirus-Specific IgA and IgG Responses in Systemic Compartment
Systemic immune responses to reovirus were also compared among experimental groups. Serum IgA titers in infected controls were markedly increased at 10 days but not at 5 days (Fig. 6). As found for fecal IgA, serum IgA titers in infected DON-treated mice were significantly higher than infected control mice. IgG2A is a major IgG isotype induced in serum by reovirus (Major and Cuff, 1996). All infected mice produced robust reovirus-specific IgG2A responses in serum after 10 days as illustrated by mean titers ranging from 3200 to 51,200 (Fig. 6). Significantly higher reovirus-specific IgG2A titers were found in sera of infected DON-treated mice at 10 days compared to infected controls.
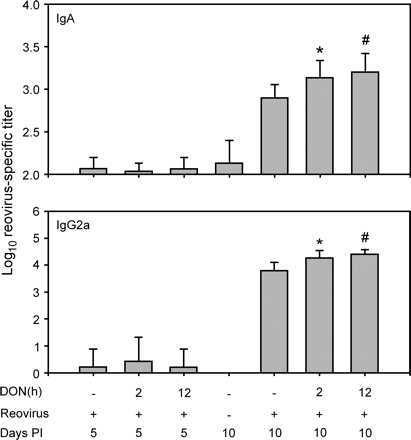
DON potentiates reovirus-specific IgA and IgG2A response in serum. Mice were treated with 0 or 25 mg/kg of DON by oral gavage for 2 to 12 h and then orally infected with 3 × 107 PFU of reovirus. Serum IgA and IgG2A titers were determined, 5 and 10 days after infection, by ELISA. Data are means ± SEM (n = 9). Bars with * and # indicate significant difference between vehicle control and groups given DON for 2 h and 12 h treatment, respectively (p < 0.05).
Ex vivo spleen fragment cultures also showed that DON-treated mice produced higher levels of reovirus-specific IgG2A then infected controls (Fig. 7). However, no differences between infected DON-treated and infected control mice responses were detectable in PP fragment cultures (data not shown). Specific IgG2A responses in LP cultures were negligible (data not shown). Reovirus also induced a weak IgG1 response after 10 days, and this was potentiated by DON (Fig. 8). Taken together, these results suggest that DON also enhanced the systemic IgA and IgG responses to reovirus.
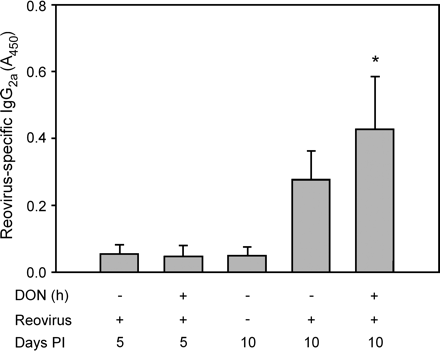
Ex vivo production of reovirus-specific IgG2A is potentiated in spleen cultures of DON-treated mice. Mice were treated with 0 or 25 mg/kg of DON by oral gavage for 2 or 12 h and then orally infected with 3 × 107 PFU of reovirus. Spleen fragment cultures were incubated for 5 days without additional stimulation. Reovirus-specific IgG2A of cultured supernatants were determined by ELISA. Data are means ± SEM (n = 9). Bars marked with * are significantly different from corresponding infected control given vehicle (p < 0.05).
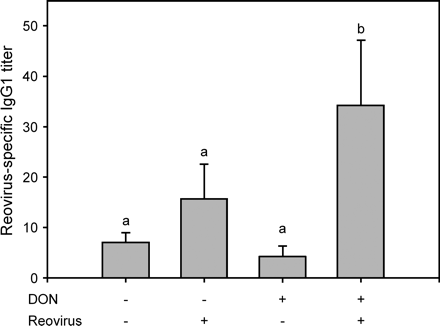
DON potentiates reovirus-specific IgG1 response in serum. Mice were treated with 0 or 25 mg/kg of DON by oral gavage for 2 h and then orally infected with 3 × 107 PFU of reovirus. Serum reovirus-specific IgG1 titers were determined by ELISA 10 days after infection. Data are means ± SEM (n = 5). Bars without same letter are significantly different (p < 0.05).
DON Suppresses Th1 Cytokine Response in PP
Since Th1 cytokines play a key role in regulating cell-mediated viral clearance, the effects of DON treatment on reovirus-induced IFN-γ and IL-2 mRNA expression were assessed. Reovirus infection of control mice resulted in increased IFN-γ mRNA expression at 3, 5, and 7 days compared to day 0 (Fig. 9), whereas effects on IL-2 mRNA (Fig. 9) were undetectable. Exposure to DON alone induced robust IFN-γ and IL-2 mRNA responses after 2 h (day 0). However, DON exposure suppressed subsequent IFN-γ responses to reovirus at 3 and 5 days as compared to infected controls. IL-2 mRNA concentrations in infected DON-treated mice returned to infected control levels at 1, 3, 5, and 7 days.
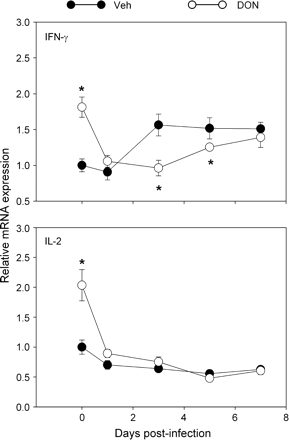
Kinetics of Th1 cytokine mRNA expression in Peyer's patches of DON-treated mice. Mice were treated with 0 or 25 mg/kg of DON by oral gavage for 2 h and then orally infected with 3 × 107 PFU of reovirus. After 0, 1, 3, 5, and 7 days, PP RNAs were isolated and relative expression of IFN-γ and IL-2 mRNA measured by real-time PCR. Data are means ± SEM (n = 6). Asterisk indicates significantly different from infected control (p < 0.05).
To confirm effects on IFN-γ secretion by T cells, PP cell suspensions were isolated at 4 days and subjected to ELISPOT analysis after passage through nylon wool and restimulation with reovirus (Table 2). These results revealed that DON exposure suppressed induction of total and CD4+ IFN-γ-producing cells from uninfected and reovirus-infected mice. These results are consistent with DON suppression of IFN-γ mRNA expression in PP of reovirus-infected mice.
ELISPOT Detection of IFN-γ and IL-4 Producing Cells in Reovirus-Challenged PP Cultures*
Spot forming cells per 106 | ||||||
|---|---|---|---|---|---|---|
| IFN-γ | ||||||
| Total | CD4+ | IL-4 | ||||
| DON | Reovirus | |||||
| − | − | 96 ± 5.0b | 40 ± 4.0b | 114 ± 5.0a | ||
| + | − | 79 ± 3.9a | 26 ± 5.6a | 149 ± 3.9c | ||
| − | + | 147 ± 6.4c | 70 ± 4.3c | 100 ± 6.4a | ||
| + | + | 107 ± 3.7b | 31 ± 6.7a | 134 ± 3.7b | ||
Spot forming cells per 106 | ||||||
|---|---|---|---|---|---|---|
| IFN-γ | ||||||
| Total | CD4+ | IL-4 | ||||
| DON | Reovirus | |||||
| − | − | 96 ± 5.0b | 40 ± 4.0b | 114 ± 5.0a | ||
| + | − | 79 ± 3.9a | 26 ± 5.6a | 149 ± 3.9c | ||
| − | + | 147 ± 6.4c | 70 ± 4.3c | 100 ± 6.4a | ||
| + | + | 107 ± 3.7b | 31 ± 6.7a | 134 ± 3.7b | ||
PP cultures (5 × 105/well) were prepared 4 days after DON and/or reovirus treatment of mice with 1 reovirus multiplicity of infection and ELISPOT performed. Data are mean ± SEM of two pooled experiments (n = 12).
Values without same letter in a column differ significantly (p < 0.05).
ELISPOT Detection of IFN-γ and IL-4 Producing Cells in Reovirus-Challenged PP Cultures*
Spot forming cells per 106 | ||||||
|---|---|---|---|---|---|---|
| IFN-γ | ||||||
| Total | CD4+ | IL-4 | ||||
| DON | Reovirus | |||||
| − | − | 96 ± 5.0b | 40 ± 4.0b | 114 ± 5.0a | ||
| + | − | 79 ± 3.9a | 26 ± 5.6a | 149 ± 3.9c | ||
| − | + | 147 ± 6.4c | 70 ± 4.3c | 100 ± 6.4a | ||
| + | + | 107 ± 3.7b | 31 ± 6.7a | 134 ± 3.7b | ||
Spot forming cells per 106 | ||||||
|---|---|---|---|---|---|---|
| IFN-γ | ||||||
| Total | CD4+ | IL-4 | ||||
| DON | Reovirus | |||||
| − | − | 96 ± 5.0b | 40 ± 4.0b | 114 ± 5.0a | ||
| + | − | 79 ± 3.9a | 26 ± 5.6a | 149 ± 3.9c | ||
| − | + | 147 ± 6.4c | 70 ± 4.3c | 100 ± 6.4a | ||
| + | + | 107 ± 3.7b | 31 ± 6.7a | 134 ± 3.7b | ||
PP cultures (5 × 105/well) were prepared 4 days after DON and/or reovirus treatment of mice with 1 reovirus multiplicity of infection and ELISPOT performed. Data are mean ± SEM of two pooled experiments (n = 12).
Values without same letter in a column differ significantly (p < 0.05).
DON Enhances Th2 Cytokine Response to Reovirus in PP
Th2 cytokines have been associated with Ig responses. Although reovirus did not induce Th2 cytokine mRNAs in PPs, DON exposure for 2 h significantly elevated IL-6 and IL-10 and suppressed IL-4 after 2 h (day 0) (Fig. 10). Interestingly, IL-6 mRNA expression in infected DON-treated mice decreased sharply to the same level as infected controls after 1 day and stayed at that level throughout the remainder of the experiment. IL-4 mRNA expression was also reduced in infected DON-treated mice compared to infected controls at 1 day but was increased at 3, 5, and 7 days post-infection. DON also elevated IL-10 mRNA expression in reovirus-infected mice at 1 day, but this decreased to control values at 3 days.
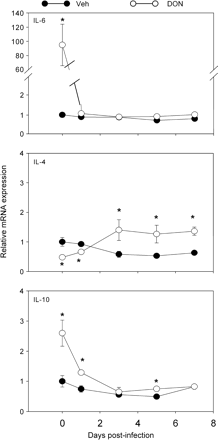
Kinetics of Th2 cytokine mRNA expression in Peyer's patches of DON-treated mice. Mice were treated as described in Fig. 9 legend, and relative expression of IL-6, IL-10, and IL-4 mRNA was determined by real-time PCR. Data are means ± SEM (n = 6). Asterisk indicates significantly different from infected control (p < 0.05).
Given that the effect of DON on IL-4 mRNA was most robust, ELISPOT assay was conducted after 4 days on PP lymphocytes. DON exposure suppressed reovirus-induced IL-4-producing cells in cells isolated from uninfected and reovirus-infected mice (Table 2). The results suggested that DON treatment enhanced Th2 cytokine expression prior to and after reovirus infection.
DON Suppresses TGF-β mRNA Expression
TGF-β is known to be a switch factor in the IgA response. DON exposure significantly elevated TGF-β mRNA in PP within 2 h (Fig. 11). However, at 3, 5, and 7 days, TGF-β mRNA expression was less in mice treated with reovirus and DON than with reovirus alone. These data suggest that elevated IgA switching resulting from increased TGF-β might not explain aforementioned increases in IgA production.
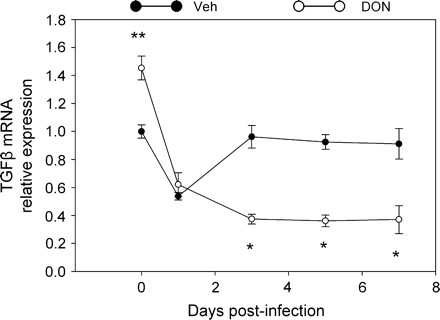
Kinetics of TGF-B mRNA expression in Peyer's patches of DON-treated mice. Mice were treated as described in Fig. 9 legend, and relative expression of TGF-B mRNA was determined by real-time PCR. Data are means ± SEM (n = 6). Asterisk indicates significantly different from infected control (p < 0.05).
DISCUSSION
The capacity of many natural toxins and environmental toxicants to modulate immunity has potentially serious implications relative to mounting appropriate responses to infectious agents as well as generating untoward inflammatory or autoimmune responses (Germolec, 2004). The trichothecenes have a unique capacity to up- and down-regulate immune function by disrupting intracellular signaling within leukocytes (Pestka et al., 2004). The results presented herein demonstrate that a single DON exposure transiently suppressed the host response to reovirus as reflected by increased severity of GI-infection and fecal shedding of the virus. The sublethal DON dose (25 mg/kg) used for most experiments here has been used in previous studies evaluating the effects of this toxin on immune function (Zhou et al., 1997, 1998). Since this dose is incapable of inducing apoptosis in thymus, PP, spleen or bone marrow (Zhou et al., 1999, 2000), altered reovirus clearance is not explainable by leukocyte death in the mucosal or systemic immune compartments. Rather, suppression appeared to correspond with suppressed IFN-γ and increased IL-4 mRNA expression in PP. Ultimately, DON-treated mice were still capable of resolving the infection, and this corresponded with enhanced mucosal IgA as well as systemic IgA, IgG2A, and IgG1 titers.
The critical immunologic determinants for resistance to reovirus and other enteric viruses remain unclear. Resistance is multifactorial and includes elements of innate and adaptive immunity. It is likely that decreased IFN-γ responses in DON-exposed mice contributed to impaired cellular or innate immune responses in early resistance to infection. The observation that DON inhibited IFN-γ expression early during the infection is consistent with diminished clearance of reovirus infection and a suppressed Th1 response. Suppression of IFN-γ production has been previously shown to be mediated by IL-4 (Szabo et al., 2003), IL-6 (Diehl and Rincon, 2002), and IL-10 (Conti et al., 2003). Thus, upregulation of these Th2 cytokines by DON might have mediated IFN-γ suppression. IFN-γ plays a key role in antiviral immunity by multiple mechanisms including suppression of viral replication, macrophage activation, inducible nitric oxide synthase expression, and stimulation of specific cytotoxic immunity via cell-surface-bound antigen associated MHC proteins (Boehm et al., 1997; Shtrichman and Samuel, 2001). Suppression of IFN-γ expression might have impaired one or more of these mechanisms, thus prolonging reovirus replication and survival. Future efforts should focus on clarifying how DON suppression of IFN-γ modulates reovirus-induced changes of macrophage, NK, dendritic, or T-cell function as well as epithelial cell physiology, pIgR levels, or immune tolerance.
Ultimately, the IFN-γ response recovered (day 8) and likely facilitated removal of the virus in conjunction with elevated reovirus-specific IgA. It should be noted that IgG2A response is a major isotype of IgG induced by reovirus (Major and Cuff, 1996), and this is considered to be Th1 dependent, with IFN-γ acting as a switch factor. The capacity to mount a robust IgG2A response by day 10 is consistent with recovery of IFN-γ production and a Th1 immune response.
Although class-switching to IgA-committed B cells is thought to involve TGF-β (Nakamura et al., 1996; van Ginkel et al., 1999), our results suggest that the increased reovirus-specific IgA response could not be explained by elevated TGF-β. It is, further, not clear whether Th1 or Th2 cells are beneficial for optimal secretory IgA production. However, Th2 cytokines play a role in terminal differentiation of B cells that are already committed to IgA, whereas Th1-type cytokine IFN-γ is implicated in the induction of the polymeric Ig receptor (pIgR) needed for transport of secretory IgA (van Ginkel et al., 2000). The Th2 cell produces more IL-4, thereby expanding Th2 cells, which support the associated immune response involving secretion of IL-4, IL-5, IL-6, IL-9, IL-10, IL-13. IL-4 supports IgG1 subclass and IgE production, but IgA is also produced during a Th2-dominated response (Boyaka et al., 2001; Yamamoto et al., 1996). Thus, increased IL-4 might act at the level of class switching or by driving differentiation of IgA-committed B cells, along with IL-6, to plasma cells.
Enhanced mucosal and systemic IgA responses to reovirus are congruent with previous findings that chronic DON exposure potentiates total serum and fecal IgA (Dong et al., 1991; Forsell et al., 1986; Greene et al., 1994) as well as ex vivo PP and splenic IgA production (Bondy and Pestka, 1991; Dong et al., 1991; Forsell et al., 1986; Greene et al., 1994). Chronic DON exposure also enhances IgA responses to intestinal and self antigens (Pestka et al., 1990b; Rasooly and Pestka, 1992, 1994; Rasooly et al., 1994). These earlier findings, consistent with DON-induced potentiation of reovirus-specific IgA, occurred in feces, serum, and fragment cultures during late-stage infection observed here. Previously, we have observed that DON is an effective inducer of IL-6 (Zhou et al., 1998, 1999) and that elevated IL-6 is crucial to increased IgA responses observed in mice chronically exposed to DON (Pestka and Zhou, 2000). Thus, it was not surprising to observe that acute DON-exposed mice had markedly increased IL-6 mRNA expression in PP. Further evidence of a skewed Th2 response were DON potentiation of serum IgG1 as well as the marked elevations of IL-4 and IL-10 mRNA in PPs of DON-exposed mice. All three of these cytokines can promote proliferation and terminal differentiation of Ig-secreting cells and downregulate Th1 responses (Conti et al., 2003; Diehl and Rincon, 2002; Mathers and Cuff, 2004; Yamamoto et al., 1996). Taken together, these data are consistent in DON enhancement of the Th2 response to reovirus.
The mechanism by which DON rapidly enhances the cytokine responses has been characterized previously. In T cells and macrophages, DON can increase cytokine gene expression by enhancing both activation of key transcription factors (Chung et al., 2003; Li et al., 2000; Ouyang et al., 1996; Wong et al., 2002; Zhou et al., 2003a) and cytokine mRNA stability (Chung et al., 2003; Li et al., 1997; Wong et al., 2002). Both enhanced transactivition and mRNA stabilization of immune and inflammatory genes by DON have been linked to activation of p38 and ERK 1/2 MAPKs via the ribotoxic stress response (Chung et al., 2003; Moon and Pestka, 2002; Moon et al., 2003). Two signaling transducers upstream of the MAPKs that appeared to mediate DON's action are double-stranded RNA-activated protein kinase R (Zhou et al., 2003b) and the Src-family tyrosine kinases (Zhou and Pestka, in press). The potential for interactive effects of reovirus with these transduction pathways is a potential target for further study.
Assessment of potential xenobiotic modulation of mucosal immune function has lagged behind well-established methods for measuring systemic immunotoxicity (Bondy and Pestka, 2005; Kawabata et al., 1995). The results presented herein and previously (Cuff et al., 1998) suggest reovirus model to be a robust approach for monitoring dysregulation of host resistance as well as Th1 and Th2 responses by immunotoxic chemicals. Real-time PCR is a rapid, sensitive method for detection of viral genomic RNA and DNA and can be used to monitor viral infections (Idesawa et al., 2004; Pang et al., 2004; Van Rijn et al., 2004). The demonstration that virus contamination in feces could be monitored using reovirus L2 gene real-time PCR and that it correlated with PFU assays in the intestine indicates that this is a facile approach for noninvasive monitoring of the course of reovirus infection in experimental mice.
These results have relevance to human safety assessment. The dose-response experiment suggested that as little as 2 mg/kg bw of DON potentiated reovirus L2 RNA levels in PP, whereas 10 mg/kg bw was required for significant potentiation of this RNA in fecal contents. Differences between the two endpoints might relate to RT PCR sensitivity within these two matrices. In addition, the mRNA might exist as two species in these matrices. In feces, it is likely the PCR is detecting viral RNA, whereas RNA in PP might include viral RNA as well as mRNA undergoing translation. Nevertheless, the data suggest that a relatively low dose of DON was sufficient to increase the viral burden in this model. The 2 mg/kg bw dose is equivalent to 13 ppm of DON in mouse diet, and such concentrations have been found in contaminated human food samples (Abouzied et al., 1991). These findings are particularly important in view of the 100-fold safety factor that is used establishing tolerable daily intake of natural toxins (Pestka and Smolinski, 2005). Future studies to relate to how chronic exposure to DON at low concentrations might affect response to reovirus challenge.
Taken together, the results presented herein suggest DON transiently suppresses the host response to enteric reovirus by interfering with the Th1-driven IFN-γ response, thereby increasing severity of the infection and virus shedding in feces. Corresponding increases in DON-mediated Th2 cytokines, notably IL-4 and IL-6, might concurrently drive elevated reovirus-specific IgA and IgG responses that were observed at experiment termination. From the perspective of host resistance, these observations are important for two reasons. First, elevated viral load in intestinal tissue is likely to increase inflammation and discomfort to the host during the infection process. Second, elevated fecal shedding will enhance the spread of this virus among individuals. From the perspective of immunotoxicity, these data are important because they suggest that, in addition to induction of apoptosis, trichothecenes might also suppress host response by selectively modulating cytokine responses to an infecting pathogen. In the future, it will be desirable to further explore mechanisms for Th1/Th2 modulation in this and other viral models following acute and chronic DON exposure.
This study was supported by a Strategic Research Grant from the Michigan State University Foundation and by Public Health Service Grants AI034544 (CC), ES03553 (JP), and DK058833 (JP) from the National Institutes for Health. We thank Dr. Norb Kaminski and Dr. Jack Harkema for advice on experiment design and Drs. Hui-Ren Zhou, Zahidul Islam, and Qunshan Jia for technical assistance. Conflict of interest: none declared.
References
Abouzied, M. M., Azcona, J. I., Braselton, W. E., and Pestka, J. J. (
Bai, G., and Shaner, G. (
Barkon, M. L., Haller, B. L., and Virgin, H. W. I. V. (
Boehm, U., Klamp, T., Groot, M., and Howard, J. C. (
Bondy, G. S., and Pestka, J. J. (
Bondy, G. S., and Pestka, J. J. (
Bottex, C., Martin, A., and Fontanges, R. (
Boyaka, P. N., Marinaro, M., Jackson, R. J., van Ginkel, F. W., Cormet-Boyaka, E., Kirk, K. L., Kensil, C. R., and McGhee, J. R. (
Breun, L. A., Broering, T. J., McCutcheon, A. M., Harrison, S. J., Luongo, C. L., and Nibert, M. L. (
Canady, R. A., Coker, R. D., Rgan, S. K., Krska, R., Kuiper-Goodman, T., Olsen, M., Pestka, J. J., Resnik, S., and Schlatter, J. (
Chung, Y. J., Zhou, H. R., and Pestka, J. J. (
Clifford, L. J., Jia, Q., and Pestka, J. J. (
Conti, P., Kempuraj, D., Kandere, K., Di, G. M., Barbacane, R. C., Castellani, M. L., Felaco, M., Boucher, W., Letourneau, R., and Theoharides, T. C. (
Cooray, R., and Jonsson, P. (
Cuff, C. F., Cebra, C. K., Rubin, D. H., and Cebra, J. J. (
Cuff, C. F., Fulton, J. R., Barnett, J. B., and Boyce, C. S. (
Cuff, C. F., Lavi, E., Cebra, C. K., Cebra, J. J., and Rubin, D. H. (
Diehl, S., and Rincon, M. (
Dong, W., Sell, J. E., and Pestka, J. J. (
Fan, J. Y., Boyce, C. S., and Cuff, C. F. (
Forsell, J. H., Witt, M. F., Tai, J. H., Jensen, R., and Pestka, J. J. (
Friend, S. C., Schiefer, H. B., and Babiuk, L. A. (
Fulong, D. B., Nibert, M. L., and Fields, B. N. (
Germolec, D. R. (
Greene, D. M., Azcona-Olivera, J. I., and Pestka, J. J. (
Hazel, C. M., and Patel, S. (
Hem, A., Smith, A. J., and Solberg, P. (
Idesawa, M., Sugano, N., Ikeda, K., Oshikawa, M., Takane, M., Seki, K., and Ito, K. (
Islam, Z., and Pestka, J. J. (
Islam, Z., King, L. E., Fraker, P. J., and Pestka, J. J. (
Islam, Z., Moon, Y. S., Zhou, H. R., King, L. E., Fraker, P. J., and Pestka, J. J. (
Jackson, L. S., and Bullerman, L. B. (
Kawabata, T. T., Burleson, G. R., Ernst, P. B., and Ullrich, S. E. (
Kinser, S., Jia, Q., Li, M., Laughter, A., Cornwell, P. D., Corton, J. C., and Pestka, J. J. (
Kubena, L. F., Bailey, R. H., Byrd, J. A., Young, C. R., Corrier, D. E., Stanker, L. H., and Rottinghaust, G. E. (
Li, S., Ouyang, Y. L., Dong, W., and Pestka, J. J. (
Li, S., Ouyang, Y., Yang, G. H., and Pestka, J. J. (
Logan, A. C., Chow, K. P., George, A., Weinstein, P. D., and Cebra, J. J. (
London, S. D., Rubin, D. H., and Cebra, J. J. (
Major, A. S., and Cuff, C. F. (
Major, A. S., and Cuff, C. F. (
Mathers, A. R., and Cuff, C. F. (
Moon, Y., and Pestka, J. J. (
Moon, Y., and Pestka, J. J. (
Moon, Y., Uzarski, R., and Pestka, J. J. (
Nakamura, M., Kondo, S., Sugai, M., Nazarea, M., Imamura, S., and Honjo, T. (
Nibert, M. L., Schiff, L. A., and Fields, B. N. (
Ouyang, Y. L., Li, S., and Pestka, J. J. (
Overbergh, L., Giulietti, A., Valckx, D., Decallonne, B., Bouillon, R., and Mathieu, C. (
Pang, X. L., Lee, B., Boroumand, N., Leblanc, B., Preiksaitis, K., and Yu, C. C. Ip. (
Pestka, J. J. (
Pestka, J. J., Dong, W., Warner, R. L., Rasooly, L., and Bondy, G. S. (
Pestka, J. J., Moorman, M. A., and Warner, R. L. (
Pestka, J. J., and Smolinski, A. T. (
Pestka, J. J., and Zhou, H. R. (
Pestka, J. J., Zhou, H. R., Moon, Y., and Chung, Y. J. (
Rasooly, L., Abouzied, M. M., Brooks, K. H., and Pestka, J. J. (
Rasooly, L., and Pestka, J. J. (
Rasooly, L., and Pestka, J. J. (
Rubin, D. H., Kornstein, M. J., Eaton, M. A., and Anderson, A. O. (
Samuel, C. E., and Knutson, G. S. (
Shtrichman, R., and Samuel, C. E. (
Silvey, K. J., Hutchings, A. B., Vajdy, M. Petzke, M. M., and Neutra, M. R. (
Szabo, S. J., Sullivan, B. M., Peng, S. L., and Glimcher, L. H. (
Tai, J. H., and Pestka, J. J. (
van Ginkel, F. W., Nguyen, H. H., and McGhee, J. R. (
van Ginkel, F. W., Wahl, S. M., Kearney, J. F., Kweon, M. N., Fujihashi, K., Burrows, P. D., Kiyono, H., and McGhee, J. R. (
Van Rijn, P. A., Wellenberg, G. J., Hakze-van der Honing, R., Jacobs, L., Moonen, P. L., and Feitsma, H. (
Virgin, H. W., Bassel-Buby, R., Fields, B. N., and Tyler, K. L. (
Wong, S. S., Zhou, H. R., and Pestka, J. J. (
Yamamoto, M., Vancott, J. L., Okahashi, N., Marinaro, M., Kiyono, H., Fujihashi, K., Jackson, R. J., Chatfield, S. N., Bluethmann, H., and McGhee, J. R. (
Yan, D., Zhou, H. R., Brooks, K. H., and Pestka, J. J. (
Yan, D., Zhou, H. R., Brooks, K. H., and Pestka, J. J. (
Zhou, H. R., Harkema, J. R., Hotchkiss, J. A., Yan, D., Roth, R. A., and Pestka, J. J. (
Zhou, H. R., Harkema, J. R., Yan, D., and Pestka, J. J. (
Zhou, H. R., Islam, Z., and Pestka, J. J. (
Zhou, H. R., Islam, Z., and Pestka, J. J. (
Zhou, H. R., Lau, A. S., and Pestka, J. J. (
Zhou, H. R., Yan, D., and Pestka, J. J. (
Zhou, H. R., Yan, D., and Pestka, J. J. (
Ziprin, R. L., and Elissalde, M. H. (
Ziprin, R. L., Holt, P. S., and Mortensen, R. F. (
Author notes
*Department of Microbiology and Molecular Genetics, †Department of Food Science and Human Nutrition, ‡Center for Integrative Toxicology, Michigan State University, East Lansing, Michigan 48824; and §Department of Microbiology and Immunology, Robert C. Byrd Health Sciences Center, West Virginia University, Morgantown, West Virginia 26506

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Comments