





Abstract
The canopy-scope was recently introduced as a cheap, robust, portable and easy to use instrument for assessing canopy openness in forests. In this study, the method was tested in predominantly coniferous plantation forests, which were different in species and structure to the forests where the instrument was originally tested. The relationship between canopy openness (calculated from hemispherical photography) and canopy-scope score at individual points, despite being statistically significant, showed considerable scatter, especially at low values. Consequently, the canopy-scope is not recommended for obtaining estimates of canopy openness at a single point within plantation conifer stands. However, good results were obtained when values were averaged across a plot. Eight to 10 canopy-scope measurements in 0.25 ha were found to be sufficient to estimate canopy openness.
Introduction
Natural regeneration of seedlings beneath a forest canopy is playing an increasing role in British forestry, as alternative silvicultural systems to clearfell and replanting are becoming more widely used (Forestry Commission, 2004). A number of factors affect the success of natural regeneration: there must be a seed source nearby and a receptive seed bed (Nixon and Worrell, 1999), the microclimate and light regime must be suitable (Lieffers et al., 1999), and there must be freedom from vegetation competition (Cannell and Grace, 1993) and browsing (Gill, 1992). The amount of solar radiation transmitted through a canopy is critical: it directly influences seedling growth, the growth of competing vegetation, and the microclimate and moisture regime of a site. Transmitted radiation is closely linked to canopy openness (Whitmore et al., 1993). An assessment of canopy openness, in association with the other factors mentioned above, would assist foresters in assessing the regeneration potential of a site.
A variety of reasonably quick methods for estimating canopy openness exist, such as the moosehorn, spherical densiometer and the Clark and Clark index (for a description of these methods, see Jennings et al., 1999; Brown et al., 2000). Using hemispherical photography as the standard, Brown et al. (2000) compared a number of methods in a range of natural broadleaved forest types in Britain (canopy heights ranged from 18 to 30 m high) and in evergreen tropical rain forest (canopy 30–40 m high; see Brown et al., 2000) for a detailed description of these canopies. One method devised and tested was the canopy-scope (Brown et al., 2000), which is a sheet of transparent plastic engraved with a 5 × 5 grid of dots at 3-cm intervals. The user points the canopy-scope at the largest canopy gap and counts the number of dots that fall within the gap, with the canopy-scope held 20 cm from the eye. With the instrument used according to recommendations, the maximum score is 25. The method works on the assumption that the largest gap represents the gap size distribution of the canopy; the largest gap can therefore be used to represent the total canopy openness. Unlike other methods of estimating canopy openness (e.g. vertical sighting tube, spherical densiometer; see Jennings et al., 1999) the canopy-scope does not itself give an absolute measure of canopy openness; it must be used comparatively, or be calibrated against another method. Brown et al. (2000) found a strong relationship between canopy-scope score and canopy openness from hemispherical photographs, in stands where canopy openness was <30 per cent. The relationship was independent of forest type and canopy structure, and results showed low variability between operators.
The aim of this study was to investigate the relationship between canopy-scope score and canopy openness in a range of coniferous plantation canopy types different in species and structure from those reported in Brown et al. (2000).
Methods
Field sites
Measurements were taken in three existing experimental forest stands, using existing sample points. Each experiment was originally planned with input from a statistician, and in no case was it considered that the location of the sample points with respect to the overstorey trees would introduce statistical bias to the results. The first site consisted of two 1-ha plots in a 60-year-old European larch (Larix decidua Mill.) plantation at Aberfoyle (56° 11′ N, 4° 23′ W). One plot had recently been heavily thinned. Plot basal areas were 26.2 and 14.0 m2 ha−1, with stocking at 284 and 120 stems ha−1, and average tree height of 26 and 24 m. In each stand, measurements were taken at nine points systematically located in a 50 × 50 m central plot. The second site was a 1-ha plot in a 46-year-old Sitka spruce (Picea sitchensis (Bong.) Carr.) plantation at Kinlochard (56° 11′ N, 4° 29′ W). The stocking was 1370 stems ha−1, with a basal area of 59 m2 ha−1 and average tree height of 20 m. Measurements were taken at nine points in a 45 × 40 m inner plot. See Hale (2003) for the approximate layout of measurement points at Aberfoyle and Kinlochard. The stand was then thinned in five interventions to 15 m2 ha−1 (Hale, 2003) prior to clearfelling; measurements were repeated at each stand density, taken at the same nine points on each occasion. The third site was a 50-year-old underplanting trial in Wykeham (54° 17′ N, 0° 33′ W), where 12 conifer and broadleaved species had been planted between rows of 21-year-old Scots pine (Pinus sylvestris L.) in 1950. Two plots were used, with areas of 0.3 and 0.14 ha, stocking of 587 and 807 trees per hectare, and basal areas of 32 and 47 m2 ha−1, respectively. Average tree heights of the dominant species were 24.5 and 22.5 m, respectively. Measurements were taken at 14 existing vegetation sample plots located from 9 to 25 m apart (average 17 m) along transects crossing the plots in a zig-zag pattern. As a consequence of its history, the stands at Wykeham were more diverse in terms of diameter and height distributions than a typical plantation forest. All measurements in the larch and mixed stands were done with the trees in full leaf.
Canopy-scope measurements and hemispherical photography
A canopy-scope was made following the design of Brown et al. (2000). Canopy-scope readings were made by the same person in all plots. One reading with the canopy-scope was obtained at each of the measurement points described above, by holding it 20 cm from the eye, pointing it towards the largest gap, and counting the dots that appeared in the gap. In gaps which were an irregular shape, or very large, the canopy-scope was moved systematically to cover the gap in sections.
Hemispherical photographs were taken at each point using a Sigma 8-mm 180° lens (Sigma Corp. of America, New York, USA) mounted on a Nikon camera (FM2, Nikon Corporation, Tokyo, Japan) on a self-levelling mount, with Ilford HP5 400ASA black and white film (Ilford Imaging, Cheshire, UK). Negatives were scanned and analysed using Hemiview (Delta-T Devices, Cambridge, UK) to calculate canopy openness. The hemispherical photographs were reanalysed with all except the biggest gap blacked out, to investigate the relationship between the size of the largest gap and canopy openness.
Statistical analysis
Using pooled data from all sites, Spearman's rank correlation was calculated to enable direct comparison with the results presented in Brown et al. (2000). Pearson's product moment correlation and linear regression were also calculated to investigate the relationship between canopy-scope score and canopy openness in more detail. Where plot averages were analysed, an adjusted regression model was used (Ripley and Thompson, 1987), which accounts for the fact that both variables were averages and therefore subject to error, which violates a regression assumption that the x-axis (canopy-scope score) values were known exactly. All analyses were done using GENSTAT version 4.2 (VSN International Ltd, Oxford, UK).
Results
Figure 1 shows canopy-scope score against canopy openness for all measurement points. No measurements from this study are presented from stands with <5 per cent canopy openness, because previous work, using the same equipment and analysis methods as used here, has shown hemispherical photography to discriminate poorly between such low values (Hale, 2003). For comparison, the data from Brown et al. (2000) have been added (their figure 4c); they have expressed no lack of confidence in values obtained in dense canopies, so their results are reproduced in full.
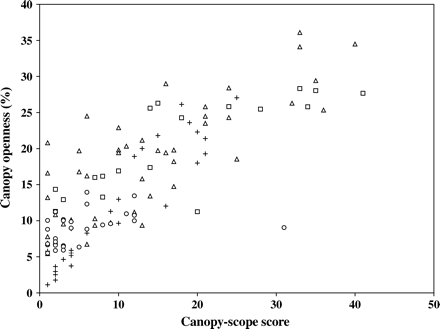
Total canopy openness (obtained from hemispherical photographs) plotted against canopy-scope score for all measurement points in the larch (squares), Sitka spruce (triangles) and mixed stands (circles). Data from Brown et al. (2000) are also shown (+).
Canopy-scope values higher than 25 were obtained in some gaps. Note that Brown et al. (2000) did not recommend using the canopy-scope where the largest gap was larger than 25 dots (corresponding to ∼30 per cent canopy openness). Canopy-scope score increases as canopy openness increases. Values from the Sitka spruce and larch stands cover a similar range; results from the mixed stand fall in the lower end of the range, and tend to show a lower canopy-openness for a given canopy-scope score.
Spearman's rank correlation was calculated for all data, and also excluded data with canopy-scope score higher than 25 (as recommended by Brown et al., 2000), to enable direct comparison against the results presented in Brown et al. (2000). All relationships were significant (P ≤ 0.001; Table 1). Using data from all sites gave a higher R2 than when sites with large gaps (canopy-scope score >25) were excluded (Table 1). Both correlation coefficients were lower than that obtained by Brown et al. (2000), indicating a poorer agreement between canopy-scope score and canopy openness.
Spearman's rank correlation coefficient calculated for canopy openness and canopy-scope score
Spearmans's rank correlation | |||
|---|---|---|---|
| n | Correlation coefficient | ||
| Brown et al. (2000) | 28 | 0.86 | |
| This study (canopy-scope score ≤25) | 79 | 0.65 | |
| This study (all data) | 91 | 0.73 | |
Spearmans's rank correlation | |||
|---|---|---|---|
| n | Correlation coefficient | ||
| Brown et al. (2000) | 28 | 0.86 | |
| This study (canopy-scope score ≤25) | 79 | 0.65 | |
| This study (all data) | 91 | 0.73 | |
Results from Brown et al. (2000) are shown for comparison.
All correlations were significant (P ≤ 0.001).
Spearman's rank correlation coefficient calculated for canopy openness and canopy-scope score
Spearmans's rank correlation | |||
|---|---|---|---|
| n | Correlation coefficient | ||
| Brown et al. (2000) | 28 | 0.86 | |
| This study (canopy-scope score ≤25) | 79 | 0.65 | |
| This study (all data) | 91 | 0.73 | |
Spearmans's rank correlation | |||
|---|---|---|---|
| n | Correlation coefficient | ||
| Brown et al. (2000) | 28 | 0.86 | |
| This study (canopy-scope score ≤25) | 79 | 0.65 | |
| This study (all data) | 91 | 0.73 | |
Results from Brown et al. (2000) are shown for comparison.
All correlations were significant (P ≤ 0.001).
Linear regression was calculated to obtain a relationship linking canopy openness to canopy-scope score, both for each site and for the pooled data set (Table 2). The relationships between canopy-scope score and canopy openness were similar for the Sitka spruce and larch sites. For the mixed site, canopy openness increased more slowly with canopy-scope score, and the relationship was not significant (P ≤ 0.05). The best model for these data had a different slope and intercept for each species, rather than fitting a common slope to all species.
Regression coefficients and proportion of variation (R2) explained for comparison of canopy openness from hemispherical photographs and canopy-scope score
Linear regression | |||||||
|---|---|---|---|---|---|---|---|
| n | R2 | Intercept | Slope | ||||
| Brown et al. (2000) | 28 | 0.86 | 1.66 | 1.04 | |||
| This study (Sitka spruce) | 45 | 0.62 | 10.4 | 0.58 | |||
| This study (larch) | 18 | 0.62 | 13.1 | 0.42 | |||
| This study (mixed) | 28 | 0.09§ | 8.1 | 0.13 | |||
| This study (all data) | 91 | 0.56 | 9.87 | 0.55 | |||
| This study (plot averages) | 9 | 0.94† | 4.24‡ | 0.98‡ | |||
Linear regression | |||||||
|---|---|---|---|---|---|---|---|
| n | R2 | Intercept | Slope | ||||
| Brown et al. (2000) | 28 | 0.86 | 1.66 | 1.04 | |||
| This study (Sitka spruce) | 45 | 0.62 | 10.4 | 0.58 | |||
| This study (larch) | 18 | 0.62 | 13.1 | 0.42 | |||
| This study (mixed) | 28 | 0.09§ | 8.1 | 0.13 | |||
| This study (all data) | 91 | 0.56 | 9.87 | 0.55 | |||
| This study (plot averages) | 9 | 0.94† | 4.24‡ | 0.98‡ | |||
For comparison, regression was also calculated on data presented in Brown et al. (2000).
Simple regression;
adjusted regression; see text for explanation.
Not significant at P ≤ 0.05; all other regressions were significant (P ≤ 0.001).
Regression coefficients and proportion of variation (R2) explained for comparison of canopy openness from hemispherical photographs and canopy-scope score
Linear regression | |||||||
|---|---|---|---|---|---|---|---|
| n | R2 | Intercept | Slope | ||||
| Brown et al. (2000) | 28 | 0.86 | 1.66 | 1.04 | |||
| This study (Sitka spruce) | 45 | 0.62 | 10.4 | 0.58 | |||
| This study (larch) | 18 | 0.62 | 13.1 | 0.42 | |||
| This study (mixed) | 28 | 0.09§ | 8.1 | 0.13 | |||
| This study (all data) | 91 | 0.56 | 9.87 | 0.55 | |||
| This study (plot averages) | 9 | 0.94† | 4.24‡ | 0.98‡ | |||
Linear regression | |||||||
|---|---|---|---|---|---|---|---|
| n | R2 | Intercept | Slope | ||||
| Brown et al. (2000) | 28 | 0.86 | 1.66 | 1.04 | |||
| This study (Sitka spruce) | 45 | 0.62 | 10.4 | 0.58 | |||
| This study (larch) | 18 | 0.62 | 13.1 | 0.42 | |||
| This study (mixed) | 28 | 0.09§ | 8.1 | 0.13 | |||
| This study (all data) | 91 | 0.56 | 9.87 | 0.55 | |||
| This study (plot averages) | 9 | 0.94† | 4.24‡ | 0.98‡ | |||
For comparison, regression was also calculated on data presented in Brown et al. (2000).
Simple regression;
adjusted regression; see text for explanation.
Not significant at P ≤ 0.05; all other regressions were significant (P ≤ 0.001).
For the pooled data set, the regression equation explained 56 per cent of the variance in the data, compared with 86 per cent for the data of Brown et al. (2000). Although statistically significant (P ≤ 0.001), the regression relationship is of limited practical use for estimating canopy openness from a single canopy-scope measurement when the scatter in the data is considered. For example, for a canopy-scope score of 6, canopy openness varied between 6 and 24 per cent in the Sitka spruce stands, and between 8 and 14 per cent in the mixed stands.
Figure 2 shows the data from Figure 1, presented here as the average canopy openness plotted against the average canopy-scope score for each plot. This shows a strong relationship between average canopy openness and average canopy-scope score, and a simple regression explained 94 per cent of the variation (although note that statistically this must be interpreted with care; see Methods) (Table 2). The adjusted regression model, which allowed for both variables being averages, exhibited no lack of fit, the intercept was not significantly different from zero, and the slope was similar to that obtained by Brown et al. (2000) (Table 2). The agreement between average canopy-scope score and average canopy openness holds even for plots where many of the individual points fell above the recommended maximum of 25 for the canopy-scope score. The high R2 and the fit of all data points close to the line illustrate the fact that plots of all species are well described by a single relationship.
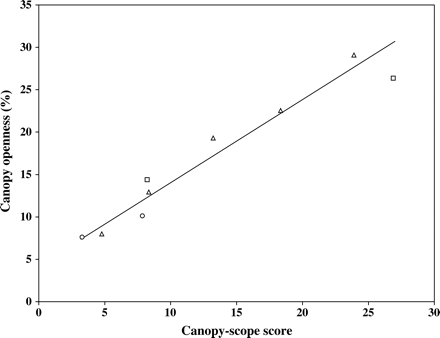
Average canopy openness plotted against average canopy-scope score for the larch (squares), Sitka spruce (triangles) and mixed stands (circles). The regression line is shown; see Table 2 for regression coefficients.
Figure 3 shows the canopy openness of the largest gap (calculated by blacking out the remainder of the hemispherical photograph) plotted against the total canopy openness for each photograph (as plotted in Figure 1). For comparison, the equivalent points from Brown et al. (2000) are also shown (their figure 5a). Regression equations fitted to these data sets show a lower slope for this study than for Brown et al. (2000) (Table 3), indicating that, although the two data sets appear to agree well at canopy openness values below 20 per cent, above this they diverge, with the Brown et al. (2000) data having a higher proportion of total canopy openness contributed by the largest gap.
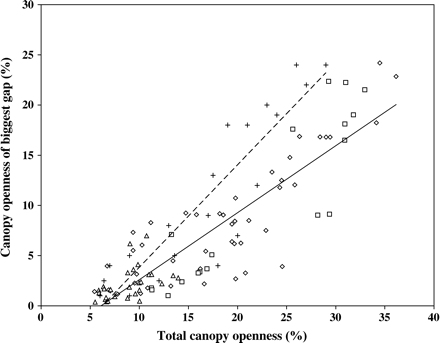
Canopy openness of the largest gap plotted against total canopy openness, for the larch (squares), Sitka spruce (triangles) and mixed (circles) stands (solid regression line). Data from Brown et al. (2000) are also shown (+; dashed regression line). See Table 3 for regression coefficients.
Regression coefficients and proportion of variation (R2) explained for comparison of canopy openness of largest gap and total canopy openness
n | R2 | Intercept | Slope | |
|---|---|---|---|---|
| Brown et al. (2000) | 20 | 0.82 | −6.30 | 1.02 |
| This study (all data) | 91 | 0.79 | −4.06 | 0.67 |
n | R2 | Intercept | Slope | |
|---|---|---|---|---|
| Brown et al. (2000) | 20 | 0.82 | −6.30 | 1.02 |
| This study (all data) | 91 | 0.79 | −4.06 | 0.67 |
For comparison, regression was also calculated on data presented in Brown et al. (2000). Both regressions were significant (P ≤ 0.001).
Regression coefficients and proportion of variation (R2) explained for comparison of canopy openness of largest gap and total canopy openness
n | R2 | Intercept | Slope | |
|---|---|---|---|---|
| Brown et al. (2000) | 20 | 0.82 | −6.30 | 1.02 |
| This study (all data) | 91 | 0.79 | −4.06 | 0.67 |
n | R2 | Intercept | Slope | |
|---|---|---|---|---|
| Brown et al. (2000) | 20 | 0.82 | −6.30 | 1.02 |
| This study (all data) | 91 | 0.79 | −4.06 | 0.67 |
For comparison, regression was also calculated on data presented in Brown et al. (2000). Both regressions were significant (P ≤ 0.001).
Discussion and conclusions
The relatively poor agreement between canopy-scope score and canopy openness obtained in this study suggests that in contrast to the results of Brown et al. (2000), the canopy-scope does not give a good indication of the canopy openness at an individual point in the forest types investigated. The lack of a significant relationship between canopy-scope score and canopy openness in the mixed plots was influenced by the relatively low range of values measured at this site, and by the outlying point which has a notably high canopy-scope for the canopy openness. At this sample point the canopy was dominated by a single large gap, with few openings elsewhere. The largest gap generally contributes a smaller proportion of the total canopy openness in this study than in Brown et al. (2000). This may be because plantation forests support a more regular canopy structure than natural forests, with gaps more evenly distributed throughout the hemisphere when viewed from a point on the ground. Alternatively, it may be due to a difference in the canopy structure of broadleaved and coniferous woodlands. The stands that had closest agreement with the data of Brown et al. (2000) were the mixed stands, which have a structure atypical of plantation forests, with a broad range of tree heights and diameters. To test species and stand structure differences more fully, measurements should be made in natural conifer and plantation broadleaf woodlands.
Despite the limitation discussed above on the use of the canopy-scope on a point-by-point basis in plantation coniferous forests, the average plot results indicate that if a number of measurements are taken across a plot, then the average canopy-scope score could be used to estimate canopy openness. The results from this study found that 8–10 measurements in a plot size of 0.25 ha were sufficient. This is practical given the portability and ease of use of the canopy-scope. Application of the sampling strategy to larger areas was not assessed in this study, but could be done by systematically increasing the number of measurements taken in a stand of, for example, 2 ha, and noting at what point the average value for the stand stopped changing significantly. It is likely that the sampling intensity of a large area, particularly of fairly uniform woodland, would be less intensive than that used in this study. Sampling could also be linked to the monitoring strategy for continuous cover forestry proposed by Kerr et al. (2002), with a canopy-scope measurement being taken at each sample point. The crude method of moving the canopy-scope within a large gap to give scores higher than 25 was not thought to introduce errors in this study. To test this rigorously, it would be desirable to compare the 25-dot canopy-scope with a larger version with more dots that could cover larger gaps.
Considering the differences in the canopy types studied here and by Brown et al. (2000), the broad agreement between the data sets supports the conclusion that the canopy-scope can be used in a wide range of forest types as a tool for assessing canopy openness, and is a satisfactory compromise between practicality and rigour.
We thank Cameron Maxwell, Colin Edwards and Owen Davies for field measurements, Tom Connolly for statistical analysis and Gary Kerr for constructive comments on an earlier draft.
References
Brown, N., Jennings, S., Wheeler, P. and Nabe-Nielson, J.
Cannell, M.G.R. and Grace, J.
Forestry Commission
Gill, R.M.A.
Hale, S.E.
Jennings, S.B., Brown, N.D. and Sheil, D.
Kerr, G., Mason, W.L., Boswell, R. and Pommerening, A.
Lieffers, V.J., Messier, C., Stadt, K.J., Gendron, F. and Comeau, P.G.
Nixon, C.J. and Worrell, R.
Ripley, B.D. and Thompson, M.

{kind=link}
{kind=link}
{kind=link}