





Abstract
Although it is often assumed that birds strongly prefer tailwinds for their migratory flights, we predict that a strategy of no wind selectivity (traveling independently of winds) may be more favorable than wind selectivity (traveling on tailwind occasions but stopping to rest under headwind occasions) for birds with low energy costs of travel relative to rest and for birds that cannot use stopover time for efficient fuel deposition. We test this prediction by analyzing the daily traveling or stopping as recorded by satellite tracking of five ospreys Pandion haliaetus, a species often using energy-saving thermal soaring, during their migration between northern Europe and Africa. Besides wind, precipitation is another weather factor included in the analyses because thermal soaring migrants are expected to stop and rest in rainy weather. In logistic regression analyses, taking into account the effects of latitude, behavior on previous day, season, date, and individual for discriminating between traveling and stopping days, we found a lack of influence of winds, suggesting that the ospreys travel or stop without regard to wind. This lack of wind selectivity under light and moderate winds is in agreement with our prediction. We expect a low degree of wind selectivity and thus regular flights under headwinds also among other types of birds that cannot use stopping time for efficient foraging and fuel deposition. We also found an unexpected lack of influence of precipitation, possibly because of relatively few instances with rainfall in combination with poor geographic precision for estimates of this weather variable.
Wind is of key importance for the migratory flights of birds, affecting both speed and cost of transport in a most profound way. Hence, it is not surprising that many studies have demonstrated that high migratory intensity often coincides with favorable tailwinds (e.g., reviews by Richardson, 1978, 1990), suggesting that wind is of primary importance for the departure decisions of migratory birds (Liechti and Bruderer, 1998). Still, there are evidence to suggest important differences in degree of wind selectivity between different species and categories of migrants, with some species showing a very distinct preference for tailwinds while others are only weakly selective of such winds, and regularly traveling also on occasions of opposed winds (Alerstam 1978) or show small effects of wind conditions on departure decisions (Rabøl, 1978; Schaub et al., 2004). However, only few studies have investigated the influence of weather on decisions of whether to travel or not at the individual level, and the results have been equivocal (Fransson, 1998; Åkesson and Hedenström, 2000).
The possible importance of flight cost as a determinant for birds' propensity to migrate under different wind conditions has been largely neglected in earlier evaluations of wind selectivity (Alerstam, 1979; Weber et al., 1998; Weber and Hedenström, 2000). We predict that wind selectivity will differ between birds depending on their flight performance, so that birds using energy-saving thermal soaring migration show a relatively low degree of wind selectivity as long as suitable thermal conditions are prevailing.
In this study, we investigate the daily traveling activity (traveling or stopping) of ospreys, as recorded by satellite tracking during their migration between northern Europe and Africa, in relation to winds and precipitation. This investigation of the birds' traveling decisions serves to test the prediction that raptors often using energy-saving thermal soaring show a low degree of wind selectivity and that they are therefore expected to regularly continue traveling also on days with opposed winds.
We include besides wind also precipitation in our analysis of migratory activity of ospreys. It seems to be general among migrating birds that they tend to stop traveling in rainy weather (e.g., reviews by Richardson, 1978, 1990). This is presumably because rain increases flight costs (to an unknown degree) and perhaps also because of orientation difficulties in rainy weather with much clouds and poor visibility. For thermal soaring migrants, rain has the additional detrimental effect of disrupting the development of thermals, and these soaring migrants would have to change to costly flapping flight when continuing to travel in rain. We therefore predict that thermal soaring migrants are equally or even more prone than other migrants to stop and rest in rainy weather. Given this prediction, it is important to include precipitation in our analysis in order to evaluate the birds' response to winds independently of the effect of rainy weather.
The migration of ospreys has already been evaluated with respect to geographical pattern, speed, and orientation (e.g., Hake et al., 2001; Kjellén et al., 2001). Ospreys normally travel solitarily and regularly use both soaring and flapping flight (e.g., Kerlinger, 1989). A few satellite-tracked ospreys crossed large bodies of water where thermals cannot be exploited (Hake et al., 2001), but normally they use a high proportion of soaring flight on migration (Kjellén et al., 2001). Adults were found to compensate for lateral wind drift and gain considerable tailwind assistance (Thorup et al., 2003). In ospreys on autumn migration, Kjellén et al. (2001) found that on average 46 days were spent on migration (range 14–81 days), and of these 19 days were stopover days (range 0–44) at up to four sites for one individual.
After deriving predictions about wind selectivity in the next section, we proceed to analyze the traveling and stopover behavior of ospreys tracked by satellite in order to test whether they behave in agreement with predictions for migrants with low traveling costs.
PREDICTIONS
Consider a highly simplified situation where migrants having stored all fuel necessary for the journey, will travel on all days independently of winds (strategy 1: nonselectivity of winds) or will travel on days with tailwinds and stop to rest on headwind days (strategy 2: selectivity of winds). We assume that tailwind days, when the birds benefit from a wind effect w (w > 0; w is the wind effect relative to the birds' travel speed under calm conditions) and thus achieve a ground speed of (1 + w), occur with probability p (0 < p < 1). Headwind days, associated with a ground speed (1 − w), are then assumed to occur with probability (1 − p). The ratio of energy consumption during a day of traveling relative to that during a day of resting is assumed to be e (e > 1).
A low ratio of energy consumption during travel in relation to rest (i.e., low e) will be expected to promote strategy 1 (no wind selectivity). Gliding flight requires a comparatively low metabolic rate, and for birds using thermal soaring migration during 9 h of a traveling day, the ratio of energy consumption during a traveling day relative to a resting day may be as low as 1.75:1 (Alerstam, 2000). Such a low value of e is expected to be associated with a strategy of no wind selectivity, except under conditions of extremely large variability between tail- and headwinds and of a low probability of headwinds. For birds traveling by flapping flight, which requires a high metabolic rate, wind selectivity is expected to be more pronounced (cf. Discussion).
A large variability between tail- and headwinds (large w) will promote selectivity of winds. Birds are always expected to avoid traveling on occasions with headwind resistance w approaching 1, when they would make no headway in their preferred migratory direction. Hence, with w ≥ 1, selectivity of tailwinds (strategy 2) will be the only reasonable strategy.
An increased frequency (probability) of tailwinds will promote selectivity of tailwinds.
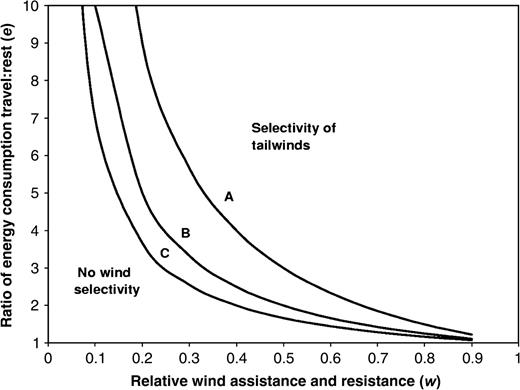
A plot showing under which conditions of relative rate of energy consumption during traveling (e) and relative wind assistance/resistance (w) a strategy of selectivity of tailwinds is more favorable than a strategy of no wind selectivity, and vice versa, under three different conditions of (A) infrequent tailwinds, (B) regular tailwinds, and (C) frequent tailwinds, respectively. The three curves show the functions of e on w as given by Equations 4–6 in the text.
ANALYSIS OF DAILY TRAVELING OR STOPPING OF MIGRATING OSPREYS
Methods
To examine the influence of weather on departure decisions and test our prediction that raptors which frequently use energy-saving thermal soaring flight show a low wind selectivity, we used data from satellite-tracked ospreys (Hake et al., 2001; Kjellén et al., 2001; Alerstam et al., 2006). We restricted the analysis to individuals that were tracked every day, omitting individuals with positions determined less often (usually every third day), because of the larger uncertainty in determining traveling and stopping days from data on birds tracked with longer time intervals. Five adult ospreys were tracked using solar-powered transmitters, which gives nearly daily positions on their migratory journeys, in several cases during both autumn and spring (Table 1 gives details for the individuals included). We used positions with a nominal accuracy within 1 km (65%; categories 3, 2, and 1 in the Argos system) or of unspecified accuracy (categories 0, A, and B in the Argos system, category Z not included; http://www.cls.fr/manuel/).
Number of days with information available about traveling or stopping for the individual ospreys Pandion haliaetus tracked on autumn and spring migration using solar-powered transmitters
| ID | Seasons tracked | No. of days in autumn | No. of days in spring |
|---|---|---|---|
| F51 | A1997, S1998 | 52 | 25 |
| F53 | A1998 | 44 | |
| M51 | A1998, S1999, A1999, S2000, A2000, S2001 | 116 | 84 |
| M52 | A1998, S1999 | 34 | 13 |
| M54 | A1998, S1999, A1999 | 58 | 26 |
| Total | 304 | 148 |
| ID | Seasons tracked | No. of days in autumn | No. of days in spring |
|---|---|---|---|
| F51 | A1997, S1998 | 52 | 25 |
| F53 | A1998 | 44 | |
| M51 | A1998, S1999, A1999, S2000, A2000, S2001 | 116 | 84 |
| M52 | A1998, S1999 | 34 | 13 |
| M54 | A1998, S1999, A1999 | 58 | 26 |
| Total | 304 | 148 |
Time intervals with uncertain information about traveling or stopping have been excluded from analysis. A and S denote autumn and spring, respectively.
Number of days with information available about traveling or stopping for the individual ospreys Pandion haliaetus tracked on autumn and spring migration using solar-powered transmitters
| ID | Seasons tracked | No. of days in autumn | No. of days in spring |
|---|---|---|---|
| F51 | A1997, S1998 | 52 | 25 |
| F53 | A1998 | 44 | |
| M51 | A1998, S1999, A1999, S2000, A2000, S2001 | 116 | 84 |
| M52 | A1998, S1999 | 34 | 13 |
| M54 | A1998, S1999, A1999 | 58 | 26 |
| Total | 304 | 148 |
| ID | Seasons tracked | No. of days in autumn | No. of days in spring |
|---|---|---|---|
| F51 | A1997, S1998 | 52 | 25 |
| F53 | A1998 | 44 | |
| M51 | A1998, S1999, A1999, S2000, A2000, S2001 | 116 | 84 |
| M52 | A1998, S1999 | 34 | 13 |
| M54 | A1998, S1999, A1999 | 58 | 26 |
| Total | 304 | 148 |
Time intervals with uncertain information about traveling or stopping have been excluded from analysis. A and S denote autumn and spring, respectively.
We evaluated for each day whether a bird had been traveling or not, resulting in a total of 452 days of decisions (275 traveling days and 177 stopover days). Individuals were considered to be traveling if they moved more than 100 km during a day. In other cases, birds were considered to have made a stopover. In the vast majority of stopover days, the birds were stationary within distances of only a few kilometers. For the analyses of the influence of weather conditions on the birds' travel decisions, we used positions where the birds had spent the night, that is, at the location where the bird made the decision whether to travel or not.
For each of these positions, we obtained National Centers for Environmental Prediction Reanalysis weather data from early morning at 6Z (6 h GMT) on wind at 1000 hPa (corresponding to low altitude winds) and precipitation provided by the National Oceanic and Atmospheric Administration-Cooperative Institute for Research in Environmental Sciences Climate Diagnostics Center, Boulder, Colorado, USA, from their Web site at http://www.cdc.noaa.gov/. The data are given for a 2.5° latitude/longitude grid. For each position, perpendicular and tailwind components were calculated from the spatially interpolated (using linear interpolation) wind vector (wind direction and speed) in relation to the mean migration direction (cf. Thorup et al., 2003). Data on precipitation were given as average precipitation rate (kg m−2 s−1, corresponding to mm rain s−1) for a 6-h period starting at 6Z and were also interpolated spatially.
NCEP Reanalysis variables are classified into classes, depending on the degree of influence of observations versus model. Wind data are strongly influenced by observations, but precipitation rate data are derived completely from the model and should be used with caution (Kalnay et al., 1996). Regional data sets on precipitation observations exist, for example, for Europe (Klein Tank et al., 2002). However, we could find no comparable large-scale data set for the total area covered by the ospreys. We compared the results using the two different precipitation data sets when analyzing European positions only. The Pearson's product-moment correlation coefficient between modeled precipitation rates and observed precipitation at nearest station was r = .35 (n = 295, p < .001). Because the two data sets on precipitation yielded very similar results, we present results using only NCEP Reanalysis data covering the whole migratory journey.
The following predictor variables were used for the analysis: (1) tailwind component, (2) sidewind component, (3) precipitation rate, and (4) latitude as numerical variables and (1) individual ID, (2) season, and (3) whether the bird had been migrating the previous day [migrating (t − 1)] as class variables. Including migrating (t − 1) resulted in a loss of 27 positions. The numerical variables were not strongly intercorrelated (none of the correlations being larger than .3, Pearson's product-moment r).
We modeled travel decisions by fitting logistic regression models on these predictor variables using PROC LOGISTIC in SAS 8.02 (SAS, 1990). The global model including intercept and all predictor variables fitted the data reasonably well (goodness-of-fit test: Pearson's χ2 = 423.45, p = .35, df = 413).
To assess the statistical evidence for the different variables, we use all-subsets model selection methods following Burnham and Anderson (2002) based on the Akaike information criterion (AIC) corrected for small sample size, AICc (Hurvich and Tsai, 1989). Inferences about the relative importance of each variable are based on the entire set of models (multimodel inference). Because model-averaged estimates based on logistic regression models can be heavily biased (Burnham and Anderson, 2002), we present only the sign of the estimates in logistic regressions.
We calculated AIC for all possible models (i.e., all combinations of including or excluding each variable). The models were then ranked according to AICc differences between the best model and model i (Δi). From these values, AICc weights (wi) were calculated to estimate the likelihood of each model. We then calculated predictor weights, w+(j), for each variable considered (a number between 0 and 1) as the sum of AIC weights for all models containing the variable. The variable with the largest predictor weight is then estimated to be the most important and the variable with the smallest weight the least important variable. A traditional hypothesis testing approach with stepwise selection yielded very similar results to those reported here.
RESULTS
Comparing the distribution of traveling and stopover days in relation to wind and precipitation (Figure 2) reveals that traveling on average was associated with relatively more tailwinds and less precipitation than stopping. However, the difference in weather conditions between traveling and stopover days was small. The mean tailwind component on traveling days (n = 275) is −0.44 m s−1 (SD = 3.8) and the precipitation rate 0.022 g m−2 s−1 (SD = 0.047) compared to −0.75 m s−1 (SD = 3.6) and 0.034 g m−2 s−1 (SD = 0.050), respectively, on stopover days (n = 177). The difference between traveling and stopover days is significant for precipitation (p = .02) but not significant for wind (p = .38). The precipitation rate on traveling days corresponds to 0.079 mm h−1 and on stopover days to 0.122 mm h−1.
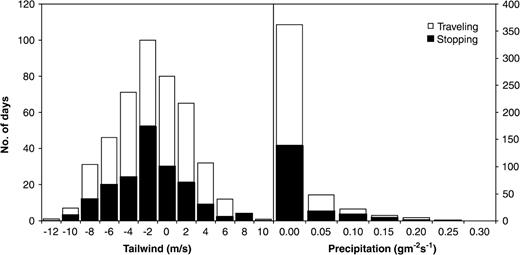
The number of days when the satellite tracked adult ospreys traveled (n = 275) or stopped (n = 177) in relation to tailwind and precipitation.
Ranking of the 127 candidate models shows five models with considerable support (Δi < 2) and 44 models with some support (Δi < 10). Model averaging (Table 2) shows very strong support for latitude and migrating (t − 1), with predictor weights of more than 0.99, and less support for season with a weight of 0.553. Sidewind has a predictor weight of 0.397, tailwind and precipitation have weights below 0.30, and ID has the lowest weight of 0.10. Furthermore, the sign of the effect for sidewind and precipitation is contrary to what would be expected (Table 2). The best-supported model includes the following three predictor variables (modeled as 0 = stopping and 1 = migrating): season (autumn estimate ± SE: −0.45 ± 0.23), latitude (−0.082 ± 0.018), and migrating (t − 1) (3.7 ± 0.4). Following a traditional hypothesis testing approach, the best-supported model has p < .001.
Relative importance of predictor variables included in the analysis of traveling decision in migrating ospreys Pandion haliaetus
| ID | Season (autumn) | Tailwind | Sidewind | Precipitation | Latitude | Migrating (t − 1) | |
|---|---|---|---|---|---|---|---|
| w+(j) | 0.098 | 0.553 | 0.262 | 0.397 | 0.278 | >0.999 | >0.999 |
| Sign of estimate (no. of models including variable) | (0) | −(3) | +(1) | +(2) | +(1) | −(5) | +(5) |
| ID | Season (autumn) | Tailwind | Sidewind | Precipitation | Latitude | Migrating (t − 1) | |
|---|---|---|---|---|---|---|---|
| w+(j) | 0.098 | 0.553 | 0.262 | 0.397 | 0.278 | >0.999 | >0.999 |
| Sign of estimate (no. of models including variable) | (0) | −(3) | +(1) | +(2) | +(1) | −(5) | +(5) |
Predictor weights, w+(j), based on model averaging over all possible models, are estimates of the relative importance of each variable. Signs of the effect of the predictor variables are based on the five logistic regression models with considerable support (Δi < 2). A positive sign means that high values of this variable are associated with a high probability of traveling relative to stopping (+ promotes traveling, − promotes stopping). In the best-supported models, the sign of individual variables was the same in all models including the variable.
Relative importance of predictor variables included in the analysis of traveling decision in migrating ospreys Pandion haliaetus
| ID | Season (autumn) | Tailwind | Sidewind | Precipitation | Latitude | Migrating (t − 1) | |
|---|---|---|---|---|---|---|---|
| w+(j) | 0.098 | 0.553 | 0.262 | 0.397 | 0.278 | >0.999 | >0.999 |
| Sign of estimate (no. of models including variable) | (0) | −(3) | +(1) | +(2) | +(1) | −(5) | +(5) |
| ID | Season (autumn) | Tailwind | Sidewind | Precipitation | Latitude | Migrating (t − 1) | |
|---|---|---|---|---|---|---|---|
| w+(j) | 0.098 | 0.553 | 0.262 | 0.397 | 0.278 | >0.999 | >0.999 |
| Sign of estimate (no. of models including variable) | (0) | −(3) | +(1) | +(2) | +(1) | −(5) | +(5) |
Predictor weights, w+(j), based on model averaging over all possible models, are estimates of the relative importance of each variable. Signs of the effect of the predictor variables are based on the five logistic regression models with considerable support (Δi < 2). A positive sign means that high values of this variable are associated with a high probability of traveling relative to stopping (+ promotes traveling, − promotes stopping). In the best-supported models, the sign of individual variables was the same in all models including the variable.
Thus, Table 2 demonstrates that traveling days were relatively more common than stopover days at southerly latitudes (Africa) compared to northerly latitudes (Europe) and that migration often took place by bouts of traveling days (several days of uninterrupted travel over longer distances), alternating with bouts of stopover days (remaining at the same site for several days).
Taking the effects of these two primary predictor variables, latitude and behavior on previous day, into account, there are still support for an additional effect of season (w+(j) > 0.5), with proportionally more traveling days than stopover days in spring compared to autumn. The remaining predictor variables have a low degree of importance in the statistical models, and they are probably not relevant for discriminating between traveling and stopover days. The models indicate that traveling days were associated with stronger crosswinds and more precipitation than stopover days. This is contrary to any biologically reasonable expectation about birds' decisions on whether to travel or not. Differences between the five individuals were of minor importance in the statistical models.
DISCUSSION
By considering that birds on migration in many cases cannot use stopover time for efficient fuel deposition, we arrive at the surprising prediction that a low degree of wind selectivity, with migration taking place not only in tailwinds but regularly also in headwinds, may be the most favorable strategy. This is particularly so for birds with low energy costs of travel relative to resting, like soaring migrants, and in situations of weak and moderate headwinds.
Comparing traveling and stopover days for ospreys during their migration between Sweden and Africa shows that there is a tendency toward slightly more tailwinds and less precipitation on traveling days (Figure 2). However, a more detailed analysis of the differences between traveling and stopover days using logistic regression and taking into account the possible effects of, besides wind and precipitation, latitude, behavior on previous day, season, and individual fails to demonstrate any influence of winds and precipitation. The shifting balance between traveling and stopover days depending on latitude, behavior on previous day, and autumn/spring season confirms the general patterns demonstrated in earlier studies of the migratory strategies of ospreys (Hake et al., 2001, Kjellén et al., 2001, Alerstam et al., 2006).
We conclude that the ospreys travel or stop without regard to wind conditions (at least in such moderate wind speeds as prevailed in this study). Hence, they show no wind selectivity, which is in agreement with our predictions that a bird species using thermal soaring flight on migration and thus low traveling cost should favor a strategy of no wind selectivity in moderate winds. Shamoun-Baranes et al. (2003) demonstrated for white storks (Ciconia ciconia), tracked by satellite during their thermal soaring migration between Europe and Sudan (NE Africa), that they traveled almost every day of their autumn migration period (which on average consisted of 25 traveling days and 1 stopover day). This gives further support to our prediction of little or no selectivity of winds for this category of migrants. Stopover days were more frequent during the storks' spring migration, but then in association with a period of foraging in the Middle East (Shamoun-Baranes et al., 2003).
Nocturnal migrants that can use the night either for traveling or resting (but not foraging) also represent an interesting case. From our schematic model, we may expect to find a lower degree of wind selectivity and thus more regular flights under headwinds (at least under weak and moderate headwinds) among this group of birds than among birds that have efficient foraging as an alternative option during their traveling time (cf. Schaub et al., 2004).
Our predictions about wind selectivity involve some important limitations:
In our schematic model, we assume that stopping on headwind occasions is associated with a cost of resting, with no or little foraging and fuel deposition taking place. This situation may be most likely among soaring migrants, which are particularly inclined to put on enough fuel stores for the whole migratory journey (Alerstam, 2000), among nocturnal migrants, which cannot use stopping nights for foraging, and among migrants traveling across habitats providing poor foraging conditions.
If stopping on headwind occasions can be used for efficient foraging and fuel deposition, selectivity of tailwinds for the migratory flights will be strongly promoted. In such cases, the birds' optimal departure decision will depend on both their fuel (energy) stores and the wind situation as evaluated by Weber et al. (1998) and Weber and Hedenström (2000). Generally, the predicted outcome from these evaluations is that birds most often will travel when favorable tailwind conditions prevail, although individual birds will not depart on all tailwind occasions (depending on their fuel load and deposition rate), and only under rather extreme conditions when tailwind occasions are rare are they expected to depart also under headwinds. Hence, a distinct preference for tailwinds during the migratory flights is expected for all migrants, irrespective of their rate of energy consumption during traveling, assuming that stopover time can always be used for efficient fuel deposition (Weber et al., 1998; Weber and Hedenström, 2000).
We have assumed that the most favorable strategy gives the lowest energy cost of migration (energy per distance covered). Considering time rather than energy, a bird showing no wind selectivity, traveling on all days including headwind days (with w < 1), will always complete migration in the shortest time. However, if the time for energy deposition is included in the total time required for migration, there will be little difference between the optimal strategies for energy and time minimization with respect to wind selectivity.
A most interesting analogous case to our wind selectivity reasoning for birds is the energetics of migration by selective tidal stream transport in fish (Metcalfe et al., 1990). Metcalfe et al. (1990) calculated for plaice (Pleuronectes platessa) the relative favorability of selective tidal stream transport (with the fish coming up into midwater and swimming when the tidal stream is flowing in the migratory direction and waiting stationary on the bottom when the stream is in the opposite direction) versus continuous swimming (swimming in midwater throughout the periods of alternating tidal stream directions) in relation to tidal stream speeds and size of the fish.
The lack of influence of precipitation is surprising and contrary to our prediction. One possible explanation is that the large-scale precipitation measure used in our analyses reflects the local rainfall situation rather poorly. Furthermore, there were relatively few days with precipitation in the data set (cf. Figure 2), which, in combination with poor geographic precision for the rainfall estimate, may contribute to make this weather variable of little use for discriminating between traveling and stopping days. Ospreys have occasionally been observed on spring migration traveling in rainy weather (K Thorup, personal observation) or at night (Alerstam et al., 2006) when there are no thermals. The ospreys' response to rainfall during migration remains to be investigated more closely through field observations, and it would call for an interesting explanation if they were found to travel regularly also in rainy weather.
Satellite tracking was supported by the Crafoord Foundation and the Swedish Natural Science Research Council. We thank Judy Shamoun-Baranes and an anonymous reviewer for helpful suggestions for improving the manuscript.
References
Åkesson S, Hedenström A,
Alerstam T,
Alerstam T, Hake M, Kjellén N, 2006. Temporal and spatial patterns of repeated migratory journeys by ospreys. Anim Behav 71:555–566.
Burnham KP, Anderson DR,
Fransson T,
Hake M, Kjellén N, Alerstam T,
Hurvich CM, Tsai CL,
Kalnay E, Kanamitsu M, Kistler R, Collins W, Deaven D, Gandin L, Iredell M, Saha S, White G, Woollen J, Zhu Y, Chelliah M, Ebisuzaki W, Higgins W, Janowiak J, Mo KC, Ropelewski C, Wang J, Leetmaa A, Reynolds R, Jenne R, Joseph D,
Kjellén N, Hake M, Alerstam T,
Klein Tank AMG, Wijngaard JB, Können GP, Böhm R, Demarée G, Gocheva A, Mileta M, Pashiardis S, Hejkrlik L, Kern-Hansen C, Heino R, Bessemoulin P, Müller-Westermeier G, Tzanakou M, Szalai S, Pálsdóttir T, Fitzgerald D, Rubin S, Capaldo M, Maugeri M, Leitass A, Bukantis A, Aberfeld R, Van Engelen AFV, Forland E, Mietus M, Coelho F, Mares C, Razuvaev V, Nieplova E, Cegnar T, Antonio Lópes J, Dahlström B, Moberg A, Kirchhofer W, Ceylan A, Pachaliuk O, Alexander LV, Petrovic P,
Liechti F, Bruderer B,
Metcalfe JD, Arnold GP, Webb PW,
Richardson WJ,
Richardson WJ,
Schaub M, Liechti F, Jenni L,
Shamoun-Baranes J, Baharad A, Alpert P, Berthold P, Yom-Tov Y, Dvir Y, Leshem Y,
Thorup K, Alerstam T, Hake M, Kjellén N,
Weber TP, Alerstam T, Hedenström A,
Author notes
aZoological Museum, University of Copenhagen, Universitetsparken 15, DK-2100 Copenhagen, Denmark
bDepartment of Animal Ecology, Lund University, Ecology Building, SE-22362 Lund, Sweden
cWildlife Damage Centre, Grimsö Wildlife Research Station, Swedish University of Agricultural Sciences, SE-73091 Riddarhyttan, Sweden

{kind=link}
{kind=link}