
Abstract
Endometriosis (EMS) is an estrogen-dependent gynecological disease with a low autophagy level of ectopic endometrial stromal cells (eESCs). Impaired NK cell cytotoxic activity is involved in the clearance obstruction of the ectopic endometrial tissue in the abdominopelvic cavity. Protopanaxadiol (PPD) and protopanaxatriol (PPT) are two metabolites of ginsenosides, which have profound biological functions, such as anti-cancer activities. However, the role and mechanism of ginsenosides and metabolites in endometriosis are completely unknown. Here, we found that the compounds PPD, PPT, ginsenoside-Rg3 (G-Rg3), ginsenoside-Rh2 (G-Rh2), and esculentoside A (EsA) led to significant decreases in the viability of eESCs, particularly PPD (IC50 = 30.64 µM). In vitro and in vivo experiments showed that PPD promoted the expression of progesterone receptor (PR) and downregulated the expression of estrogen receptor α (ERα) in eESCs. Treatment with PPD obviously induced the autophagy of eESCs and reversed the inhibitory effect of estrogen on eESC autophagy. In addition, eESCs pretreated with PPD enhanced the cytotoxic activity of NK cells in response to eESCs. PPD decreased the numbers and suppressed the growth of ectopic lesions in a mouse EMS model. These results suggest that PPD plays a role in anti-EMS activation, possibly by restricting estrogen-mediated autophagy regulation and enhancing the cytotoxicity of NK cells. This result provides a scientific basis for potential therapeutic strategies to treat EMS by PPD or further structural modification.
Similar content being viewed by others


Introduction
Endometriosis (EMS) is a common estrogen-dependent gynecological disease that is defined by the attachment of endometrial tissue at extrauterine ectopic sites where it forms invasive lesions1,2,3. It affects ~5–15% of all women of reproductive age and 20–50% of all infertile women4,5. There is strong evidence to support the important role of estrogen dependence, progesterone resistance, and defects in the immune system (e.g., impaired cytotoxic activity of NK cells) in EMS, leading to the enhanced benign tumor-like behaviors of refluxed endometrium (such as unrestrained proliferation and aggressive invasion)6,7,8,9.
Current medical therapies (e.g., progestins, androgens, gonadotropin-releasing hormone (GnRH) agonists, and aromatase inhibitors) focus primarily on reducing the systemic levels of estrogens, but these are of limited effectiveness, with frequent recurrence and considerable side effects1. Therefore, the most urgent need is to better understand the mechanisms underlying EMS, to enable the development of more effective treatments.
As a constitutive catabolic pathway, autophagy is the natural, regulated, destructive mechanism of the cell that mediates both non-specific and targeted sequestration of cellular organelles and other macromolecules, allowing the orderly degradation of cellular components in lysosomes and the recycling of bioenergetics metabolites10. Accumulating evidence has indicated that the level of autophagy in both the ectopic and eutopic endometrium of patients with EMS is decreased7,11,12,13,14,15. This aberrantly low autophagy is associated with the highly proliferative and lowly apoptotic capacities of ectopic endometrial stromal cells (eESCs)7,11,12,13,14. Our previous work has showed that estrogen promotes the survival of human ESCs via upregulating CXCL12/CXCR4-mediated autophagy inhibition7. The CXCR4-CXCL12 axis has also been shown to have both immune (e.g., lymphocyte chemotaxis, especially for NK cells) and non-immune functions (e.g., tissue repair, angiogenesis, cell invasion, and migration)16,17. In addition, our recent study has observed that the low autophagy of ESCs leads to the impaired cytotoxicity of NK cells in a co-culture system, which may be beneficial to the immune escape of ectopic ESCs8. Therefore, a new therapeutic strategy should be urgently required, that is, targeting autophagy and immunomodulation simultaneously.
As a traditional medicinal herb, ginseng is widely used in Asian countries and North America. Ginsenosides (e.g., ginsenoside-Rg3 (G-Rg3) and ginsenoside Rh2 (G-Rh2)) are the main components extracted from ginseng and have various pharmaceutical activities, such as antitumor, antioxidant, immunomodulatory, and anti-inflammatory activities18,19,20,21. As two metabolites of ginsenoside, protopanaxadiol (PPD) and protopanaxatriol (PPT) also exhibit activity against various cancer cells20,22,23. Various functional assays and molecular docking studies have provided evidence that ginsenosides can mediate their cellular activities by binding to the active sites of steroid receptors (e.g., estrogen receptor α (ERα))20. However, to date, no study has been published regarding the anti-EMS (a benign estrogen-dependent disease with tumor-like behaviors) activities of these ginsenosides and metabolites.
Therefore, the aim of this study was to investigate whether ginsenosides and metabolites have anti-EMS activity, and if so, whether this effect is dependent on the regulation of autophagy and ER in eESCs and the cytotoxicity of NK cells in vitro and in vivo.
Results
PPD has powerful anti-EMS activity in vitro
Esculentoside A (EsA), a triterpene saponin isolated from the root of the Chinese herb Phytolacca esculenta, is known for its anti-inflammatory and anti-oxidative effects24. It has been reported that EsA inhibits the growth of ectopic lesions in a rat EMS model, and its effect is stronger than that of the same dose of azole (a synthetic steroid that is used primarily in the treatment of EMS) (China Patent Number: CN200510110906.X). The chemical structures of PPD, PPT, G-Rg3, G-Rh2 and EsA are shown in Figure 1a. Owing to the anti-EMS activity and similar chemical structures among EsA, ginsenosides and metabolites, EsA was used as a positive control. First, we evaluated the effects of PPD, PPT, G-Rg3, G-Rh2, and EsA on eESCs and normal ESCs (nESCs) in vitro. As shown, treatment with PPD, PPT, G-Rh2, or EsA led to a significant decrease in eESC viability, especially at concentrations beyond 40 µM (Fig. 1b). Among these compounds, PPD has the most powerful inhibition on eESC viability, with an IC50 (half maximal inhibitory concentration) of 30.64 µM (Fig. 1c). However, none of these compounds (PPD, PPT, G-Rh2, or EsA) showed a significant effect on the viability of nESCs unless their concentrations were higher than 80 µM (Supplementary Figure 1). Further analysis showed that the expression levels of anti-apoptosis molecules B-cell lymphoma (Bcl)-xL and Bcl-2, and proliferation-related molecules Ki-67 and proliferating cell nuclear antigen (PCNA) in PPD-treated eESCs were obviously decreased compared with those of control eESC and EsA-treated eESCs (Fig. 1d, e). Conversely, the tumor metastasis suppressor CD8225 and pro-apoptotic molecules Bax and Bak were increased (Fig. 1d–f). These results indicated that a low level of PPD can markedly inhibits the viability of eESCs not nESCs in vitro.

a Chemical structures of PPD, PPT, G-Rg3, G-Rh2, and EsA. b After treatment with PPD, PPT, G-Rg3, G-Rh2 or EsA at different concentrations (0–160 µM) for 24 or 48 h, the viability of ectopic ESCs (eESCs, n = 6) was analyzed by the CCK-8 assay (one-way ANOVA). c IC50 of the compounds in eESCs. d, e After treatment with PPD or EsA (40 µM) for 48 h, the expression levels of Bcl-xL, Bcl-2, Bax, Ki-67, PCNA, and CD82 in ectopic ESCs (eESCs, n = 6) were analyzed by FCM (one-way ANOVA). f After treatment with PPD or EsA (40 µM) for 48 h, the expression levels of Bak in eESCs (n = 6) were analyzed by western blotting. Data are expressed as mean ± SEM. *P < 0.05, **P < 0.01, and ***P < 0.001 (compared to ctrl for 24 h); #P < 0.05, ##P < 0.01, and ##P < 0.001 (compared to ctrl for 48 h). NS no statistically difference
PPD suppresses estrogen-mediated autophagy inhibition of eESCs possibly by downregulating ERα and upregulating progesterone receptor α (PRα) in vitro
To investigate the potential effect of PPD on the autophagy of eESCs, quantitative real time polymerase chain reaction (qRT-PCR)-based analysis of 84 genes involved in autophagy (a human autophagy PCR array) was performed (Fig. 2a). As shown, 21 different genes (such as autophagy-related genes (ATG)2, ATG3, and ATG5, ESR1, SQSTEM1, and TGFB1) between control eESCs and PPD-treated eESCs (the differential fold >3 in eESCs) was found (Fig. 2b). The results of western blotting showed that PPD upregulated autophagy-related molecules microtubule-associated protein light chain 3B (LC3B)-II, Beclin-1, and downregulated p62 expression in eESCs (Fig. 2c), suggesting that PPD could enhance the autophagy of eESCs.

a Human Autophagy RT2 profiler™ PCR array was performed to evaluate the differential expression of autophagy-related genes in eESCs after treatment with or without PPD (40 µM) for 48 h. b Twenty-one different genes between control eESCs and PPD-treated eESCs (fold difference >3 in eESCs). c The eESCs (n = 6) were indicated with PPD (40 µM) or rapamycin (1 µM) for 48 h, and then western blotting was used to analyze the expression levels of LC3B, Beclin-1, p62, ERα, ERβ, PRα, and PRβ in eESCs. d, e After treatment with PPD (40 µM), 17β-estrogen (E2, 10−7M) or PPD plus E2 for 48 h, the expression levels of ERα, ERβ, PRα, PRβ, Beclin-1, and p62 in eESCs (eESCs, n = 6) were analyzed by western blotting. f After treatment with PPD (40 µM), ERα antagonist (MPP dihydrochloride, 2.7 nM), or PPD plus MPP dihydrochloride for 48 h, the expression levels of Beclin-1 and p62 in eESCs (n = 6) were analyzed by western blotting. ERαi MPP dihydrochloride, Rapa rapamycin
Meanwhile, treatment with PPD led to a significant decrease in ERα and increase in PRα in eESCs (Fig. 2c). PPD could restrict the stimulatory effect on ERα expression and inhibitory effect on autophagy induced by 17-β estradiol (E2) in eESCs (Fig. 2d, e). In addition, PPD alone, ERα antagonist MPP dihydrochloride alone or the combination of PPD and MPP dihydrochloride resulted in the elevated autophagy of eESCs (Fig. 2f). In addition, MPP dihydrochloride led to a low level of Bcl-2/Bcl-XL and a high level of Bax/Bak, similar to that with PPD, but there was no synergistic effect with the combined treatment of MPP dihydrochloride plus PPD (Supplementary Figure 2). These data suggest that PPD can downregulate ERα expression and upregulate PRα expression, and further induce a high level of autophagy and high ratio of pro-apoptosis molecules (Bax/Bak) to anti-apoptosis molecules (Bcl-2/Bcl-XL) in eESCs in vitro.
PPD activates NK cells in response to eESCs in a co-culture model
To explore the potential role of PPD in NK cells, NK cells were isolated from the peripheral blood of healthy women and were directly treated with PPD, or co-cultured with control eESCs or PPD-pretreated eESCs. Next, we found that both PPD treatment and co-culture with PPD-pretreated eESCs increased the expression of natural killer group 2A (NKG2A) and activating natural cytotoxicity receptors (NKp30 and NKp46)26,27 and cytokine interferon (IFN)-γ, and decreased IL-10 expression in NK cells (Fig. 3a, b).

a, b NK cells isolated from PBMCs were treated with PPD or were co-cultured with control eESCs or PPD-pretreated eESCs (n = 6) for 48 h, and then the expression levels of IFN-γ, IL-10, NKG2A, NKp30, and NKp46 in NK cells were analyzed by FCM. Ctrl: NK cells alone; PPD: NK cells treated with PPD; ESC: NK cells co-cultured with control eESCs; ESC-PPD: NK cells co-cultured with PPD-pretreated eESCs (one-way ANOVA). c, d After co-culture with control eESCs (ESC-NK) or PPD-pretreated eESCs (ESC-PPD-NK) for 48 h, the NK cells were collected and further co-cultured with fresh eESCs (n = 6) for another 48 h. The expression levels of Bcl-2, Bcl-xL, Ki-67, and CD82 in eESCs were analyzed by FCM. (Student’s t-test). Data are expressed as mean ± SEM. *P < 0.05, **P < 0.01, ***P < 0.001, and ****P < 0.0001
To further investigate the role of these NK cells in the viability of eESCs, we collected NK cells after co-culture with control eESCs (ESC-NK) or PPD-pretreated eESCs (ESC-PPD-NK), and then co-cultured with fresh eESCs (Fig. 3c). As shown, compared with ESC-NK co-cultured eESCs, the expression of Bcl-2, Bcl-xL, and Ki-67 was markedly decreased, and the expression of CD82 was increased in ESC-PPD-NK co-cultured eESCs (Fig. 3d). These data suggest that PPD can induce the activation directly and indirectly by acting on eESCs, and PPD-pretreated eESC-cultured NK cells may further restrict the growth and invasion in vitro.
Low doses of PPD do not influence ERα or PRα expression, proliferation, or the autophagy of the uterine endometrium in a mouse EMS model
To evaluate the potential therapeutic value of PPD in EMS, we analyzed the role of PPD in eutopic endometrium. According to the procedure of Figure 4a, the mouse EMS model was constructed by allotransplantation, and was intraperitoneally injected with low doses of PPD. As shown, PPD did not change the expression of ERα, PRα, Ki-67, Beclin-1, and LC3B in mouse uterine endometrium (Fig. 4b, c), suggesting that intraperitoneal injection of low-dose PPD does not regulate ERα and PRα expression, proliferation, and autophagy of uterine endometrium in a mouse EMS model.

a–c The EMS model of BALB/C mice was constructed by allotransplantation and was intraperitoneally injected with low doses of PPD (15 mg/kg, 100 µl) on day 4 and day 10. On day 14, the expression levels of ERα, ERβ, PR, Ki-67, Beclin-1, and LC3B in mouse uterine tissues were detected by immunofluorescence (IF, original magnification: ×200) (b) and immunohistochemistry (IHC) (c)
PPD reverses the inhibitory effects of estrogen on the autophagy of mouse ectopic lesions and the activation of NK cells
As shown, all compounds (PPD, PPT, G-Rg3, G-Rh2, and EsA) at a high dose could significantly reduce the number and weight of ectopic lesions in a mouse EMS model, especially PPD (Fig. 5a). A significant inhibitory effect on the weight of mouse ectopic lesions could be achieved with the administration of low doses of PPD (Fig. 5b).

a The number and weight of ectopic lesions in a mouse EMS model after intraperitoneal injection of the compounds (PPD, PPT, G-Rg3, G-Rh2, or EsA) at a high dose (45 mg/kg, 100 µl) (one-way ANOVA). b The number and weight of ectopic lesions in mouse EMS model after intraperitoneal injection of PPD at a high dose (PPD H, 45 mg/kg, 100 µl), medium dose (PPD M, 30 mg/kg, 100 µl) or low-dose (PPD L, 15 mg/kg, 100 µl) (one-way ANOVA). c–f The mouse EMS model was intraperitoneally injected with low doses of PPD (15 mg/kg, 100 µl), E2 (150 µg/kg,100 µl; intramuscular injection) and or PPD (15 mg/kg, 100 µl; intraperitoneal injection), or 3-MA (50 mg/kg, 100 µl; intraperitoneal injection) on day 4 and day 10. On day 14, the expression levels of ERα, ERβ, PR, Ki-67, Beclin-1, and LC3B in mouse ectopic lesions were detected by IF (original magnification: ×200) (c) and IHC (d–f)
Subsequently, we found that PPD led to the decrease in ERα and the increase in PRα in ectopic lesions (Fig. 5c), echoing the results in vitro. E2 significantly promoted the proliferation of ectopic lesions, and this effect could be reversed by PPD (Fig. 5d). E2 suppressed the expression of Beclin-1 and LC3B of ectopic lesions as well as 3-methyladenine (3-MA, an autophagy inhibitor), and PPD also completely abrogated it (Fig. 5e, f).
In addition, treatment with E2 downregulated Granzyme B and NKG2D in CD45+CD3e-DX5+NK cells from mouse peritoneal fluids (Fig. 6a, b). Although the intraperitoneal injection of PPD had no similarly effects on NK cells, the combination of PPD and E2 induced the activation of NK cells compared with E2 alone (Fig. 6a, b).

a, b The mouse EMS model was intraperitoneally injected with low doses of PPD, E2 and/or PPD, or 3-MA on day 4 and day 10. On day 14, the expression levels of Granzyme B, NKG2D, IFN-γ, and perforin in CD45+CD3e-DX5+NK cells from mouse peritoneal fluid were analyzed by FCM (one-way ANOVA). Data are expressed as mean ± SEM. *P < 0.05, **P < 0.01, ***P < 0.001, and ****P < 0.0001
PPD suppresses the estrogen-mediated growth of mouse ectopic lesions
Finally, we analyzed the effect of PPD and E2 on the number and weight of ectopic lesions in a mouse EMS model. As shown, E2 increased the number and weight of mouse ectopic lesions (Fig. 7a, b). In contrast, PPD not only decreased the number and weight of mouse ectopic lesions, but also restricted the stimulatory effect of E2 on the growth of mouse ectopic lesions (Fig. 7a, b). These data suggest that PPD suppresses the growth of mouse ectopic lesions induced by estrogen and plays a role in anti-mouse EMS activity.

a, b Number and weight of ectopic lesions from the mouse EMS model, which was treated as described in Figure 6. Data are expressed as mean ± SEM. *P < 0.05, **P < 0.01, and ***P < 0.001
Discussion
Ginsenosides within the dammarane-type consist mainly of three types classified according to their genuine aglycone moieties: PPD, PPT, and ocotillol. The potential health effects of ginsenosides include anti-inflammatory, antistress, anticarcinogenic, immunomodulatory, antiallergic, antiatherosclerotic, and antihypertensive effects as well as antidiabetic effects and regulatory effects on blood pressure and metabolism28,29,30. Here, we found that PPD, PPT, G-Rg3, and G-Rh2, especially PPD, could inhibit viability and growth as well as EsA both in vitro and in vivo. Additionally, low doses of PPD did not influence the growth of nESCs or the eutopic endometrium. These results suggest that PPD may be a safe and effective strategy for the treatment of EMS.
Autophagy has been linked to various pathophysiological processes, including cell death, development, tumorigenesis, and immunity31,32. We had found that autophagy in eESCs was significantly decreased, and estrogen suppressed the autophagy of ESCs by upregulating CXCL12 and CXCR4 expression. Here, the analysis of autophagy-related genes in this study showed that PPD enhanced the transcription of several autophagy-related genes in eESCs, such as ATGs and MAP1L3CB. Of note, the ESR1 and TGFB mRNA levels were decreased. Under hormonal control, TGF-β is abundantly and differentially expressed in the endometrium and is involved in the promotion of ESC survival, cell adhesion and invasion, angiogenesis33. Additionally, TGF-β contributes to the suppression of the immune system in the environment of ectopic lesions such as the induction of IL-10+T help (Th)17 and regulatory T (Treg) cell differentiation and the impairment of NK cells cytotoxicity9,33,34,35. Therefore, the anti-EMS effects of PPD may also be dependent on the regulation of TGFB.
Estrogen and progestogen modulate proliferation, apoptosis, and autophagy in the human endometrium and in endometriotic cells and tissues, further contributing to the origin and development of EMS7,36. In the current study, we also observed that E2 treatment led to increased Ki-67 expression and a low autophagy level in ectopic lesions and elevated numbers and weights of ectopic lesions in a mouse EMS model. In addition, studies using ER knockout transgenic mice and treatment with ER subtype-selective ligands have indicated the crucial roles of ERs in EMS37,38,39,40. Progesterone resistance was observed in EMS. The molecular basis of progesterone resistance in EMS may be related to an overall reduction in the levels of PR37. The progesterone responses are mediated primarily via binding to and activation of the nuclear receptors (PR-A and PR-B)38,41. We found that PPD not only decreased ERα expression but also upregulated PRα expression in eESCs in vitro and in vivo. Our previous works have reported that progesterone signaling can suppress the inhibitory effect of estrogen on ESC autophagy7. Therefore, the stimulation of PR likely plays an important role in anti-EMS by PPD.
Here, we observed that PPD upregulated Bax and Bak, and downregulated Bcl-2 and Bcl-xL in ectopic ESCs possibly by downregulating ERα. In addition to ER signaling, other signaling pathways such as PI3K/AKT, ROS and NF-κB participate in the regulation process of ginsenosides in cell apoptosis and death42,43,44,45. Under the regulation of estrogen, these signaling pathways are also involved in regulating ESC’s apoptosis in EMS36,46,47,48. Therefore, PPD may also regulate these apoptosis-related proteins via these signaling pathways (e.g., PI3K/AKT, ROS, and NF-κB) in EMS, a finding that require further research.
Ginsenosides are structurally described as triterpenoid saponins that contain a steroidal backbone. Although most naturally occurring ginsenosides have bulky sugar side chains that pose a huge steric hindrance for these molecules to bind to steroid receptors, various functional assays and molecular docking studies have provided evidence to show that ginsenosides can mediate their cellular activities by binding to the active sites of steroid receptors and downregulating the expression of estrogen receptors20,22,49. On the other hand, other studies suggest that ginsenosides may modulate their cellular actions via pathways independent of steroid receptors50,51,52. However, the regulatory mechanism of ginsenosides for hormone receptors remain unclear and warrants further research.
The impaired cytotoxic activity of NK cells is associated with several physiological and pathological processes, including EMS14,27,53. The effector functions of NK cells include the cytotoxicity and secretion of cytokines (e.g., IFN-γ, TNF-α). The most potent activated receptors of NK cells are the antibody-dependent cell-mediated cytotoxicity (ADCC)-mediating molecule CD16 and NKG2D26,27. Moreover, NK cells recognize their ligands in tumor or virus-infected cells and mediate natural cytotoxicity through a set of activating natural cytotoxicity receptors (e.g., NKp30, and NKp46)26,27. In the current study, PPD promoted the cytotoxic activity of NK cells, such as via the upregulation of granzyme B and perforin, especially under the stimulation of estrogen. Of note, we did not observe the effect of PPD alone on the activation NK cells in the mouse EMS model. This result may have resulted from the low concentration and or complex microenvironment in EMS, but the mechanisms deserve further investigation.
EMS is also an inflammatory disorder. The action of estrogen not only is essential for endometriotic tissue growth but also contributes to ongoing inflammation, neovascularization, and associated pain54. Ginsenosides are known to have anti-cancer properties based on their anti-inflammatory activation, and their low toxicities render them excellent candidates for cancer therapy20. For example, ginseng total saponins markedly reduce the production of pro-inflammatory cytokines (e.g., TNF-α, interleukin (IL)-1β, and IL-6) in lipopolysaccharide (LPS)-stimulated rat astrocyte and microglia cultures in vitro. These inflammatory cytokines were also increased in the eESCs, ectopic lesions and peritoneal fluids from patients with EMS40,55. Therefore, the anti-EMS property may be partly dependent on the anti-inflammatory effects, which requires further research.
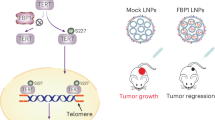
Collectively, our findings, schematized in Figure 8, reveal that PPD can downregulate the expression of ERα and upregulate the expression of PRα. The inhibition of estrogen/ERα signaling mediated by PPD further induces the autophagy of eESCs, activates the cytotoxicity of NK cells in response to eESCs, suppresses the growth of ectopic lesions, enhances the immune surveillance of ectopic lesions, and finally inhibits the development of EMS. Therefore, PPD should have potential therapeutic value for treating diseases such as EMS. Differences in the saponin structure markedly influence bioactivity and bioavailability. Therefore, further structural modification of PPD is needed to identify better anti-EMS drugs.

PPD can downregulate the expression of ERα and upregulate the expression of PRα. The inhibition of estrogen/ERα signaling mediated by PPD further induces the autophagy of eESCs and activates the cytotoxicity of NK cells (upregulation of granzyme B, NKG2D, IFN-γ, and perforin) in response to eESCs, and suppresses the growth of ectopic lesions and enhances the immune surveillance to ectopic lesions finally inhibiting the development of EMS
Materials and methods
Patients and sample collection
The study protocol was approved by the Human Research Ethics Committee of Obstetrics and Gynecology Hospital, Fudan University, and written informed consent was obtained from all patients. All of the endometriotic tissues were obtained by laparoscopy from 43 patients with EMS at the Obstetrics and Gynecology Hospital of Fudan University. Normal endometrium was obtained through hysterectomy from patients with leiomyoma (six cases) as healthy control samples. None of the included patient experienced complications related to pelvic inflammatory disease and no patient took any medication or received hormonal therapy within six months prior to surgery. All the samples were obtained in the proliferation phase of the cell cycle, as confirmed histologically according to established criteria.
Isolation and culture of ESCs
We isolated human nESCs from the endometrium of healthy control subjects, and eESCs from ectopic lesions of women with EMS according to a previously described method25. It supplied >98% vimentin+CK7- ESCs, as confirmed by flow cytometry (FCM) analysis.
Purification of NK cells
Peripheral blood mononuclear cells (PBMCs) were isolated from healthy fertile women. Human NK cells were isolated from PBMCs using magnetic beads (Miltenyi Biotec, Bergisch Gladbach, Germany) for in vitro experiments. These NK cells were directly treated with PPD (40 µM, Sigma-Aldrich Co. LLC., USA); or were co-cultured with control eESCs or PPD-pretreated eESCs for 48 h, and then these NK cells were collected to analyze the expression of IFN-γ, IL-10, NKG2A, NKp30, and NKp40 by FCM; or were further co-cultured with fresh eESCs for another 48 h, and then the expression levels of Bcl-2, Bcl-xL, Ki-67, and CD82 in eESCs were analyzed by FCM.
The cell-counting kit-8 (CCK-8) assay
The eESCs were treated with PPD, PPT, G-Rg3, G-Rh2, or EsA (0–160 µM, Sigma-Aldrich Co. LLC., USA) for 24 or 48 h, with 0.1% DMSO as a blank control. Next, these cells were collected, and the viability was detected by the CCK-8 assay (Dojindo, Japan). According to the manufacturer’s protocol, the CCK-8 reagent was added to each well and cells were incubated at 37°C for 1–4 h. The absorbance (optical density) at 450 nm was measured and used to represent the cell viability. Each experiment was performed in six parallel wells and repeated three times.
FCM
After treatment with PPD (40 µM) or EsA (40 µM), or co-culture with control eESCs (ESC-NK) or PPD-pretreated eESCs (ESC-PPD-NK), these eESCs were collected and the expression levels of Bcl-2, Bcl-xL, Bax, Ki-67, PCNA, and or CD82 (all from Biolegend, USA) were analyzed by FCM according to the manufacturer’s instructions. In addition, the expression levels of IFN-γ, IL-10, NKG2A, NKp30, and NKp40 (all from Biolgend) in CD56+NK cells were analyzed by FCM. Isotypic IgG antibodies were used as controls. The samples were analyzed using a FACS-Calibur flow cytometer (Becton Dickinson, USA) and Cellquest software (Becton Dickinson). Statistical analysis was conducted using isotype-matched controls as references.
Human Autophagy RT2 profiler™ PCR array
The eESCs were indicated with PPD (40 µM) for 48 h, with 0.1% DMSO as a blank control. Next, human Autophagy RT2 profiler™ PCR array (Catalog No. PAHS-084ZR, 96-well format) was utilized to analyze the transcriptional levels of these autophagy-related genes in eESCs as described previously7,56. The plates were processed using an Applied Biosystems 7500 fast RT-PCR system (Applied Biosystems, USA), and the data were interpreted with SABiosciences’ web-based PCR array analysis tool.
Protein extraction and western blotting
The eESCs were treated with PPD (40 µM), rapamycin (1 µM, sigma), E2 (10-7M, Sigma), E2 plus PPD, ERα antagonist (MPP dihydrochloride; 2.7 nM; Tocris Bioscience, USA), or PPD plus MPP dihydrochloride for 48 h, and then cells were washed in phosphate buffered saline (PBS), detached with a cell scraper and centrifuged for 20 min at 12,000 r.p.m. at 4 °C. The pellet was resuspended in high efficiency cell tissue rapid lysis buffer (RIPA; Beyotime, Shanghai, China) containing 1% phenylmethanesulfonylfluoride (PMSF; Beyotime) proteinase and 1% phosphatase inhibitors (Roche, USA). Cell lysates were boiled for 10 min at 95 °C and then were stored at −80 °C. Protein concentrations were quantified using the BCA protein assay kit (Beyotime). Total proteins (20 µg) were electrophoresed in SDS-PAGE gels (EpiZyme scientific) using a Miniprotein III system (Bio-Rad, USA) and were transferred to PVDF membranes (Millipore, USA) at 2 h, followed by overnight incubation with primary antibody against Bax, Bak, Beclin-1, p62, LC3B, ERα, PR, or GAPDH (1:1000; Cell Signaling Technology, USA), and ERβ (1:200; Santa Cruz Biotechnology, Santa Cruz, CA, USA) at 4 °C. Then PVDF membranes were washed three times with PBST solution and were incubated at room temperature for 1 h in peroxidase-conjugated goat anti-rabbit IgG secondary antibodies (1:5000; Bioworld Technology, Co. Ltd. USA). Thereafter the membrane was washed three times and processed for chemiluminescence using the Immobilon Western Chemiluminescent HRP Substrate Kit (Millipore).
Mouse EMS model
A group of adult female BALB/C mice was purchased from the Laboratory Animal Facility of Fudan University and was used for this study. They were maintained for 2 weeks at the animal facility before use. The Animal Care and Use Committee of Shanghai First Maternity and Infant Hospital, Tongji University School of Medicine approved all the animal protocols.
We constructed an intraperitoneal EMS model. On Day 0, the uterus of female BALB/C mice (Donor mice) was minced, and then the tissue debris was intraperitoneally injected into female BALB/C mice (for recipient mice, the ratio of the uterus to intraperitoneal injection of mice was 1:2). On day 4 and day 10, the EMS mice were intraperitoneally injected with the compounds (PPD, PPT, G-Rg3, G-Rh2, or EsA) at a high dose (45 mg/kg, 100 µl), medium dose (30 mg/kg, 100 µl) or low-dose (15 mg/kg, 100 µl). In addition, some EMS mice were treated with E2 (150 µg/kg,100 µl; intramuscular injection), and/or PPD (15 mg/kg, 100 µl; intraperitoneally injection), or 3-MA (50 mg/kg, 100 µl; intraperitoneally injection), 0.1% DMSO (100 µl) as a control. On Day 14, the EMS-like lesions and peritoneal fluids were collected and detected. The number and weight of EMS-like lesions were counted. Immunohistochemistry (IHC) and immunofluorescence (IF) were used to analyze the expression of ERα, ERβ, PR, Ki-67, Beclin-1 and LC3B in the EMS-like lesions. Additionally, the expression levels of IFN-γ, NKG2D, granzyme B and perforin (all from Biolegend) in CD45+CD3e-DX5+NK cells in peritoneal fluid were analyzed by FCM.
IHC and IF
For IHC, paraffin sections (5 µm) of the EMS-like lesions and uterine endometrium were dehydrated in graded ethanol and then were incubated with hydrogen peroxide and 1% bovine serum albumin/TBS to block endogenous peroxidase. The samples were then incubated with rabbit anti-human Ki-67 (1:500; Abcam, USA), Beclin-1 (1:200; Abcam) and LC3B (1:500; Abcam) or rabbit IgG isotype (Abcam) overnight at 4 °C in a humid chamber. After washing three times with TBS, the sections were overlaid with peroxidase-conjugated goat anti-rabbit IgG, and the reaction was developed with 3,3-diaminobenzidine (DAB) and counterstained with hematoxylin.
For IF, according to a previous procedure7, EMS-like lesions and uterine endometrium tissues were incubated with anti-goat ERα, anti-goat ERβ, anti-goat PR antibody (1:100, Abcam) in PBS at 4 °C overnight. The slides were then incubated with Alexa Fluor 488-conjugated donkey anti-goat secondary antibody (1:500; Abcam). The nuclei were then stained with 4′,6-diamidino-2-phenylindole (DAPI; Beyotime, China). Images were captured with a confocal microscope (Leica, Germany).
Statistics
All values are shown as the mean ± SEM. The data were analyzed with GraphPad Prism version 5 by t-test for two groups or one-way analysis of variance using Tukey’s post hoc test for multiple groups. Differences were considered statistically significant at P < 0.05.
References
Bulun, S. E. Endometriosis. N. Engl. J. Med. 360, 268–279 (2009).
Giudice, L. C. & Kao, L. C. Endometriosis. Lancet 364, 1789–1799 (2004).
Han, S. J. et al. A new isoform of steroid receptor coactivator-1 is crucial for pathogenic progression of endometriosis. Nat. Med 18, 1102–1111 (2012).
Eskenazi, B. & Warner, M. L. Epidemiology of endometriosis. Obstet. Gynecol. Clin. North Am. 24, 235–258 (1997).
Pritts, E. A. & Taylor, R. N. An evidence-based evaluation of endometriosis-associated infertility. Endocrinol. Metab. Clin. North Am. 32, 653–667 (2003).
Pollacco, J., Sacco, K., Portelli, M., Schembri-Wismayer, P. & Calleja-Agius, J. Molecular links between endometriosis and cancer. Gynecol. Endocrinol. 28, 577–581 (2012).
Mei, J. et al. Estrogen promotes the survival of human secretory phase endometrial stromal cells via CXCL12/CXCR4 up-regulation-mediated autophagy inhibition. Hum. Reprod. 30, 1677–1689 (2015).
Yu, J. J. et al. IL15 promotes growth and invasion of endometrial stromal cells and inhibits killing activity of NK cells in endometriosis. Reproduction 152, 151–160 (2016).
Yang, H. L. et al. The crosstalk between endometrial stromal cells and macrophages impairs cytotoxicity of NK cells in endometriosis by secreting IL-10 and TGF-β. Reproduction 154, 815–825 (2017).
He, C. & Klionsky, D. J. Regulation mechanisms and signaling pathways of autophagy. Annu. Rev. Genet. 43, 67–93 (2009).
Choi, J., Jo, M., Lee, E., Oh, Y. K. & Choi, D. The role of autophagy in human endometrium. Biol. Reprod. 86, 70 (2012).
Choi, J., Jo, M., Lee, E., Kim, H. J. & Choi, D. Differential induction of autophagy by mTOR is associated with abnormal apoptosis in ovarian endometriotic cysts. Mol. Hum. Reprod. 20, 309–317 (2014).
Choi, S., Shin, H., Song, H. & Lim, H. J. Suppression of autophagic activation in the mouse uterus by estrogen and progesterone. J. Endocrinol. 221, 39–50 (2014).
Yang, H. L. et al. Autophagy in endometriosis. Am. J. Transl. Res. 9, 4707–4725 (2017).
Ruiz, A. et al. Effect of hydroxychloroquine and characterization of autophagy in a mouse model of endometriosis. Cell Death Dis. 7, e2059 (2016).
Wu, X. et al. Human first-trimester trophoblast cells recruit CD56brightCD16- NK cells into decidua by way of expressing and secreting of CXCL12/stromal cell-derived factor 1. J. Immunol. 175, 61–68 (2005).
Meng, W., Xue, S. & Chen, Y. The role of CXCL12 in tumor microenvironment. Gene 641, 105–110 (2018).
Christensen, L. P. Ginsenosides chemistry, biosynthesis, analysis, and potential health effects. Adv. Food Nutr. Res. 55, 1–99 (2009).
Zhang, Q., Kang, X. & Zhao, W. Antiangiogenic effect of low-dose cyclophosphamide combined with ginsenoside Rg3 on Lewis lung carcinoma. Biochem. Biophys. Res. Commun. 342, 824–828 (2006).
Wong, A. S., Che, C. M. & Leung, K. W. Recent advances in ginseng as cancer therapeutics: a functional and mechanistic overview. Nat. Prod. Rep. 32, 256–272 (2015).
Li, B. et al. Ginsenoside Rh2 induces apoptosis and paraptosis-like cell death in colorectal cancer cells through activation of p53. Cancer Lett. 301, 185–192 (2011).
Gu, C. J. et al. Protopanaxadiol and metformin synergistically inhibit estrogen-mediated proliferation and anti-autophagy effects in endometrial cancer cells. Am. J. Transl. Res. 9, 4071–4082 (2017).
Xu, F. Y. et al. The antitumor activity study of ginsenosides and metabolites in lung cancer cell. Am. J. Transl. Res. 8, 1708–1718 (2016).
Wu, F., Yi, Y., Sun, P. & Zhang, D. Synthesis, in vitro inhibitory activity towards COX-2 and haemolytic activity of derivatives of esculentoside A. Bioorg. Med. Chem. Lett. 17, 6430–6433 (2017).
Li, M. Q. et al. CD82 gene suppression in endometrial stromal cells leads to increase of the cell invasiveness in the endometriotic milieu. J. Mol. Endocrinol. 47, 195–208 (2011).
Cooper, M. A. et al. Human natural killer cells: a unique innate immunoregulatory role for the CD56 (bright) subset. Blood 97, 3146–3151 (2001).
Lanier, L. L. NK cell recognition. Annu. Rev. Immunol. 23, 225–227 (2005).
Chang, Y. S., Seo, E. K., Gyllenhaal, C. & Block, K. I. Panax ginseng: a role in cancer therapy? Integr. Cancer Ther. 2, 13–33 (2003).
Attele, A. S., Wu, J. A. & Yuan, C. S. Ginseng pharmacology: multiple constituents and multiple actions. Biochem. Pharmacol. 58, 1685–1693 (1999).
Kitts, D. D. & Hu, C. Efficacy and safety of ginseng. Public Health Nutr. 3, 473–485 (2000).
Levine, B., Mizushima, N. & Virgin, H. W. Autophagy in immunity and inflammation. Nature 469, 323–335 (2011).
Auberger, P. & Puissant, A. Autophagy, a key mechanism of oncogenesis and resistance in leukemia. Blood 129, 547–552 (2017).
Omwandho, C. O., Konrad, L., Halis, G., Oehmke, F. & Tinneberg, H. R. Role of TGF-betas in normal human endometrium and endometriosis. Hum. Reprod. 25, 101–109 (2010).
Chang, K. K. et al. IL-27 triggers IL-10 production in Th17 cells via a c-Maf/RORγt/Blimp-1 signal to promote the progression of endometriosis. Cell Death Dis. 8, e2666 (2017).
Li, M. Q. et al. CD4+Foxp3+regulatory T cell differentiation mediated by endometrial stromal cell-derived TECK promotes the growth and invasion of endometriotic lesions. Cell Death Dis. 5, e1436 (2014).
Reis, F. M., Petraglia, F. & Taylor, R. N. Endometriosis: hormone regulation and clinical consequences of chemotaxis and apoptosis. Hum. Reprod. 19, 406–418 (2013).
Bulun, S. E. et al. Estrogen receptor-beta, estrogen receptor-alpha, and progesterone resistance in endometriosis. Semin Reprod. Med. 28, 36–43 (2010).
Kulak, J. Jr., Fischer, C., Komm, B. & Taylor, H. S. Treatment with bazedoxifene, a selective estrogen receptor modulator, causes regression of endometriosis in a mouse model. Endocrinology 152, 3226–3232 (2011).
Harris, H. A., Bruner-Tran, K. L., Zhang, X., Osteen, K. G. & Lyttle, C. R. A selective estrogen receptor-beta agonist causes lesion regression in an experimentally induced model of endometriosis. Hum. Reprod. 20, 936–941 (2005).
Zhao, Y. et al. Dual suppression of estrogenic and inflammatory activities for targeting of endometriosis. Sci. Transl. Med. 7, 271ra9 (2015).
Al-Sabbagh, M., Lam, E. W. & Brosens, J. J. Mechanisms of endometrial progesterone resistance. Mol. Cell. Endocrinol. 358, 208–215 (2012).
Wu, Q. et al. Ginsenoside Rh4 induces apoptosis and autophagic cell death through activation of the ROS/JNK/p53 pathway in colorectal cancer cells. Biochem. Pharmacol. 148, 64–74 (2018).
Cui, J. et al. Ginsenoside Rg2 protects PC12 cells against β-amyloid25-35-induced apoptosis via the phosphoinositide 3-kinase/Akt pathway. Chem. Biol. Interact. 275, 152–161 (2017).
Yuan, Z., Jiang, H., Zhu, X., Liu, X. & Li, J. Ginsenoside Rg3 promotes cytotoxicity of Paclitaxel through inhibiting NF-κB signaling and regulating Bax/Bcl-2 expression on triple-negative breast cancer. Biomed. Pharmacother. 89, 227–232 (2017).
Huang, G. et al. Notoginsenoside R1 attenuates glucose-induced podocyte injury via the inhibition of apoptosis and the activation of autophagy through the PI3K/Akt/mTOR signaling pathway. Int. J. Mol. Med 39, 559–568 (2017).
Li, M. Q. et al. CXCL8 enhances proliferation and growth and reduces apoptosis in endometrial stromal cells in an autocrine manner via a CXCR1-triggered PTEN/AKT signal pathway. Hum. Reprod. 27, 2107–2116 (2012).
Park, S., Lim, W., Bazer, F. W. & Song, G. Apigenin induces ROS-dependent apoptosis and ER stress in human endometriosis cells. J. Cell. Physiol. 233, 3055–3065 (2018).
Banu, S. K., Lee, J., Speights, V. O. Jr., Starzinski-Powitz, A. & Arosh, J. A. Selective inhibition of prostaglandin E2 receptors EP2 and EP4 induces apoptosis of human endometriotic cells through suppression of ERK1/2, AKT, NFkappaB, and beta-catenin pathways and activation of intrinsic apoptotic mechanisms. Mol. Endocrinol. 23, 1291–1305 (2009).
Park, J. et al. Effects of ginseng on two main sex steroid hormone receptors: estrogen and androgen receptors. J. Ginseng Res. 41, 215–221 (2017).
Leung, K. W. et al. Signaling pathway of ginsenoside-Rg1 leading to nitric oxide production in endothelial cells. FEBS Lett. 580, 3211–3216 (2006).
Song, I. H. & Buttgereit, F. Non-genomic glucocorticoid effects to provide the basis for new drug developments. Mol. Cell. Endocrinol. 246, 142–146 (2006).
Hafezi-Moghadam, A. et al. Acute cardiovascular protective effects of corticosteroids are mediated by non-transcriptional activation of endothelial nitric oxide synthase. Nat. Med. 8, 473–479 (2002).
Osuga, Y. et al. Lymphocytes in endometriosis. Am. J. Reprod. Immunol. 65, 1–10 (2011).
Bulun, S. E. et al. Role of estrogen receptor-β in endometriosis. Semin. Reprod. Med. 30, 39–45 (2012).
Burney, R. O. & Giudice, L. C. Pathogenesis and pathophysiology of endometriosis. Fertil. Steril. 98, 511–519 (2012).
Deshpande, N. P. et al. Campylobacter concisus pathotypes induce distinct global responses in intestinal epithelial cells. Sci. Rep. 6, 34288 (2016).
Acknowledgements
We thank Dr. Yi-Qin Wang in the Department of Pathology, Hospital of Obstetrics and Gynecology, Fudan University for helping with histological analysis. This study was supported by the Major Research Program of National Natural Science Foundation of China (NSFC) (nos. 91542108, 81471513, 31671200, 81601354, and 81671460), the Shanghai Rising-Star Program 16QA1400800, the Development Fund of Shanghai Talents (201557), the Oriented Project of Science and Technology Innovation from Key Lab. of Reproduction Regulation of NPFPC (CX2017-2), the National Basic Research Program of China (2015CB943300), the National Key Research and Development Program of China (2017YFC1001401), the Program for Shanghai leaders, the Program of Shanghai Outstanding Academic Leader (15XD1500900), the National Science Foundation of Jiangsu Province (no. BK20160128), and Fundamental Research Funds for the Central Universities (no. 021414380180).
Author information
Authors and Affiliations
Contributions
B.Z. and W.J.Z. conducted all experiments and prepared the figures and the manuscript. C.J.G., K.W., H.L.Y., and J.J.Y. assisted with animal trials and IHC assay. J.M. and X.F.H. helped to edit the manuscript. J.S.S. helped to design the study and edit the manuscript. F.Y.X. and D.J.L. helped to edit the manuscript. M.Q.L. and L.P.J. designed the study, supervised the project, and edited the manuscript. All the authors were involved in writing the manuscript.
Corresponding authors
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Edited by B. Zhivotovsky
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Zhang, B., Zhou, WJ., Gu, CJ. et al. The ginsenoside PPD exerts anti-endometriosis effects by suppressing estrogen receptor-mediated inhibition of endometrial stromal cell autophagy and NK cell cytotoxicity. Cell Death Dis 9, 574 (2018). https://doi.org/10.1038/s41419-018-0581-2
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41419-018-0581-2
This article is cited by
-
ERG mediates the inhibition of NK cell cytotoxicity through the HLX/STAT4/Perforin signaling pathway, thereby promoting the progression of myocardial infarction
Journal of Physiology and Biochemistry (2024)
-
CCL20/CCR6 axis mediates macrophages to promote proliferation and migration of ESCs by blocking autophagic flux in endometriosis
Stem Cell Research & Therapy (2022)
-
Anti-inflammatory cytokines in endometriosis
Cellular and Molecular Life Sciences (2019)
