
Abstract
Maternal uniparental disomy for chromosome 14 (upd(14)mat) causes clinically discernible features such as pre- and/or postnatal growth failure, hypotonia, obesity, small hands, and early onset of puberty. The monoallelic expression patterns at the 14q32.2 imprinted region are tightly related to methylation status of the DLK1–MEG3 intergenic differential methylation region (DMR) and the MEG3-DMR that are severely hypermethylated after paternal transmission and grossly hypomethylated after maternal transmission. We examined this imprinted region in a 2 2/12-year-old Japanese patient who was born with a normal birth size (length, +0.2 SD; weight, −0.5 SD) and showed postnatal growth failure (height, −3.1 SD; weight, −3.4 SD), hypotonia, frontal bossing, micrognathia, and small hands. Methylation analysis, genotyping analysis, and deletion analysis were performed with blood samples of the patient and the parents, showing that the DMRs of this patient were grossly hypomethylated in the absence of upd(14)mat and deletion of the DMRs. The results indicate the occurrence of an epimutation (hypomethylation) affecting the normally methylated DMRs of paternal origin, and imply that epimutations should be examined in patients with upd(14)mat-like phenotype.
Similar content being viewed by others

Introduction
Maternal uniparental disomy for chromosome 14 (upd(14)mat) results in clinically discernible features such as pre- and postnatal growth failure, hypotonia, obesity, small hands, and early onset of puberty.1 Phenotypic development is consistent with chromosome 14q32.2 region harboring several paternally expressed genes (PEGs) such as DLK1 and RTL1 and maternally expressed genes (MEGs) such as MEG3 (alias GTL2), RTL1as (RTL1 antisense), and MEG8.2, 3 The parent-of-origin-specific monoallelic expression patterns are tightly related to methylation status of differential methylation regions (DMRs).4 For the 14q32.2 imprinted region, the previous studies have identified the intergenic DMR (IG-DMR) between DLK1 and MEG3 and the MEG3-DMR that are severely hypermethylated after paternal transmission and grossly hypomethylated after maternal transmission.5, 6, 7 In particular, the germline-derived IG-DMR plays a pivotal role in the imprinting regulation, because methylation pattern of the secondary MEG3-DMR is dependent on that of the IG-DMR.8
The upd(14)mat-like phenotype has also been exhibited by non-disomic patients. Temple et al9 described a patient with upd(14)mat-like phenotype and an epimutation (hypomethylation) of the normally methylated DMR of paternal origin. Kagami et al5 reported three patients with upd(14)mat-like phenotype and microdeletions affecting the 14q32.2 imprinted region including the DMRs of paternal origin. In this regard, the IG-DMR deletion from the paternally derived chromosome has no effect on the imprinting status, although that from the maternally derived chromosome results in a maternal to paternal epigenotypic alteration.5, 7 Thus, simple genotype–phenotype correlations can be applied for the three patients with the microdeletions, implying that loss of paternally derived DLK1 and RTL1 constitutes primary additive underlying factors for the development of upd(14)mat-like phenotype, although the perturbation of other imprinted genes could also have some effects.5
Here, we report an epimutation identified in a patient with upd(14)mat-like phenotype.
Patient and methods
Case report
This Japanese girl was born at 41 weeks of gestation after natural conception, with a history of mild oligohydramnios in the third trimester. At birth, her length was 50.0 cm (+0.2 SD), her weight 3.03 kg (−0.5 SD), and her head circumference 34.5 cm (+0.8 SD). The non-consanguineous parents were clinically normal, and the height was 161.0 cm (−1.7 SD) for the father and 154.5 cm (−0.7 SD) for the mother.
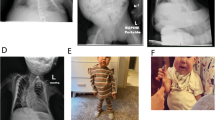
At 5 months of age, she was referred to us, because she was unable to control her head. Physical examination revealed generalized hypotonia without palsy and abnormal tendon reflex, and several somatic features such as frontal bossing, micrognathia, and small hands (Supplementary Figure 1). In addition, her length became below −2 SD of the mean from 10 months of age, while hypotonia was gradually ameliorated. She controlled her head at 7 months of age, sat without support at 11 month, and walked without support at 19 months. Repeatedly performed biochemical studies for hypotonia and growth failure were normal, as were skeletal roentgenograms and brain magnetic resonance imaging. The karyotype was 46XX in all the 30 lymphocytes examined. With a provisional diagnosis of Prader–Willi syndrome (PWS) that is primarily based on hypotonia and growth deficiency, fluorescence in situ hybridization (FISH) analysis for SNRPN and methylation analysis for the DMR at the SNRPN promoter region were performed,10 showing normal findings. In addition, hypomethylation of the H19-DMR and upd(7)mat, which can cause growth failure, were also excluded by previous methods.11, 12 On the last examination at 2 2/12 years of age, her height was 76.1 cm (−3.1 SD), her weight 7.9 kg (−3.4 SD), and her head circumference 44.9 cm (−1.9 SD). Her mental development appeared age appropriate.
Methylation analysis
This study was approved by the Institutional Review Board Committees at Hokkaido University Hospital and National Center for Child Health and Development. After obtaining written informed consent, we examined the IG-DMR (CG4 and CG6) and the MEG3-DMR (Figure 1a), using bisulfite-treated leukocyte genomic DNA. For the IG-DMR, bisulfite sequencing was performed as reported previously,5 and the SNPs (rs12437020 for CG4 and rs10133627 for CG6) were also genotyped. For the MEG3-DMR, PCR amplification was performed with methylated and unmethylated allele-specific primers, as described previously.5, 6 A hitherto unreported upd(14)mat patient and the previously reported upd(14)pat patient5 were similarly analyzed with permission.

Summary of the molecular studies. (a) The regional physical map of the human chromosome 14q32.2 imprinted region. PEGs are shown in blue and MEGs in red; although it remains to be clarified whether DIO3 is a PEG, mouse Dio3 is known to be preferentially but not exclusively expressed from a paternally derived chromosome in embryos.13 WDR25 and BEGAIN appear biparentally expressed genes. The IG-DMR and the MEG3-DMR are depicted in green, and the FISH probes 1 and 2 covering the DMRs indicated in orange. The physical distance is ∼190 kb between BEGAIN and DLK1, ∼170 kb between DLK1 and MEG8, and ∼670 kb between MEG8 and DIO3. (b) Methylation patterns of the IG-DMR (CG4 and CG6) and the MEG3-DMR. Bisulfite sequencing has been performed for CG4 and CG6. Each line indicates a single clone and each circle denotes a CpG island; filled and open circles represent methylated and unmethylated cytosines, respectively. The SNP typing data for CG4 and CG6 are also shown. Methylated (M) and unmethylated (U) allele-specific PCR amplification has been performed for the MEG3-DMR. NC: negative control. (c) FISH analysis using FISH probe 1 (F1) for the IG-DMR and FISH probe 2 (F2) for the MEG3-DMR. The red signals (arrows) have been detected by the two FISH probes and the green signals (arrowheads) have been identified by an RP11-566I2 probe for 14q12 used as an internal control.
Genotyping analysis
We performed microsatellite analysis for 16 loci on chromosome 14 and SNP analysis for 39 loci around the DMRs (Supplementary Table 1). The primers used were as reported previously.5
Deletion analysis
Lymphocyte metaphase spreads were hybridized with a long and accurate (LA)-PCR product encompassing the IG-DMR and that spanning the MEG3-DMR (Figure 1a), together with an RP11-566I2 probe for 14q12 used as an internal control. Furthermore, the two LA-PCR products were also obtained from the patient and a control subject, and subjected to fragment size comparisons after restriction enzyme digestions, to detect a possible tiny deletion in the patient. The detailed methods for the deletion analysis have been reported previously.5
Results
Methylation analysis
The results are shown in Figure 1b. For the IG-DMR, CG4 and CG6 were grossly hypomethylated in the patient and the upd(14)mat patient, severely methylated in the upd(14)pat patient, and delineated in apparently mosaic patterns in the parents. In addition, the CG4 SNP typing indicated parental origin-dependent methylation patterns in the parents, and heterodisomy for the this region in the upd(14)mat patient. The CG6 SNP typing data were not informative. For the MEG3-DMR, PCR products were obtained with an unmethylated allele-specific primer pair alone in the patient and the upd(14)mat patient, with a methylated allele-specific primer pair alone in the upd(14)pat patient, and with both primer pairs in the parents.
Genotyping analysis
Microsatellite analysis demonstrated biparental origin of the two chromosome 14 homologs, and SNP analysis indicated lack of a segmental upd(14)mat around the DMRs (Supplementary Table 1).
Deletion analysis
FISH probes 1 and 2 detected two signals in the patient (Figure 1c). The fragment size comparison after enzyme digestions showed no abnormal bands suggestive of a tiny deletion in the patient.
Discussion
This patient had hypomethylated DMRs in the absence of discernible maternal disomy affecting the DMRs or loss of the paternally derived DMRs. This implies the occurrence of an epimutation (hypomethylation) affecting the normally methylated DMRs of paternal origin. To our knowledge, such an epimutation (hypomethylation) has previously been identified only in a patient reported by Temple et al.9 Actually, the DMR examined in that patient appears to be a part of the MEG3-DMR rather than the IG-DMR on the basis of its position. It is likely, however, that the IG-DMR is also hypomethylated in that patient, because the MEG3-DMR can stay hypomethylated only in the presence of the hypomethylated IG-DMR.8
Clinical features of the two patients with epimutation are summarized in Table 1, together with those of upd(14)mat patients. Notably, clinical features are grossly similar in epimutation patients and upd(14)mat patients. Although our patient had no prenatal growth failure, lack of prenatal and/or postnatal growth failure has been described in several upd(14)mat patients,14, 15, 16 and this would be due to body growth being a multifactorial trait subject to multiple genetic and environmental factors.17 In this regard, it has been reported that clinical features are comparable between patients with paternal upd(14) and those with epimutations (hypermethylation) affecting the normally hypomethylated DMRs of maternal origin.5 Taken together, the methylation patterns of the DMRs appear to be closely related to the expression patterns of virtually all the imprinted genes on 14q32.2.
It is noteworthy that the patient was initially suspected as having PWS. Indeed, growth deficiency, hypotonia, and small hands are shared by upd(14)mat and PWS,18, 19 and upd(14)mat has occasionally been identified in patients referred for molecular examination of PWS.19, 20 Thus, upd(14)mat and epimutations should be considered in patients with PWS-like phenotype.18, 19
In summary, we observed an epimutation (hypomethylation) of the paternally derived DMRs in a patient with upd(14)mat-like phenotype. Further studies will identify epimutations in patients with upd(14)mat-like phenotype, thereby contributing to clarify the relevance of epimutations in human imprinted disorders.
References
Kotzot D : Maternal uniparental disomy 14 dissection of the phenotype with respect to rare autosomal recessively inherited traits, trisomy mosaicism, and genomic imprinting. Ann Genet 2004; 47: 251–260.
Cavaille J, Seitz H, Paulsen M, Ferguson-Smith AC, Bachellerie JP : Identification of tandemly-repeated C/D snoRNA genes at the imprinted human 14q32 domain reminiscent of those at the Prader-Willi/Angelman syndrome region. Hum Mol Genet 2002; 11: 1527–1538.
Charlier C, Segers K, Wagenaar D et al: Human-ovine comparative sequencing of a 250-kb imprinted domain encompassing the callipyge (clpg) locus and identification of six imprinted transcripts: DLK1, DAT, GTL2, PEG11, antiPEG11, and MEG8. Genome Res 2001; 11: 850–862.
Li E, Beard C, Jaenisch R : Role for DNA methylation in genomic imprinting. Nature 1993; 366: 362–365.
Kagami M, Sekita Y, Nishimura G et al: Deletions and epimutations affecting the human chromosome 14q32.2 imprinted region: implications for the phenotypic development in paternal and maternal uniparental disomy for chromosome 14. Nat Genet 2008; 40: 237–242.
Murphy SK, Wylie AA, Coveler KJ et al: Epigenetic detection of human chromosome 14 uniparental disomy. Hum Mutat 2003; 22: 92–97.
Lin SP, Youngson N, Takada S et al: Asymmetric regulation of imprinting on the maternal and paternal chromosomes at the Dlk1-Gtl2 imprinted cluster on mouse chromosome 12. Nat Genet 2003; 35: 97–102.
Takada S, Paulsen M, Tevendale M et al: Epigenetic analysis of the Dlk1-Gtl2 imprinted domain on mouse chromosome 12: implications for imprinting control from comparison with Igf2-H19. Hum Mol Genet 2002; 11: 77–86.
Temple IK, Shrubb V, Lever M, Bullman H, Mackay DJ : Isolated imprinting mutation of the DLK1/GTL2 locus associated with a clinical presentation of maternal uniparental disomy of chromosome 14. J Med Genet 2007; 44: 637–640.
The ASHG/ACMG Test and Technology Transfer Committee: Diagnostic testing for Prader–Willi and Angelman syndromes. Am J Hum Genet 1996; 58: 1085–1088.
Kagami M, Nagai T, Fukami M, Yamazawa K, Ogata T : Silver-Russell syndrome in a girl born after in vitro fertilization: partial hypermethylation at the differentially methylated region of PEG1/MEST. J Assist Reprod Genet 2007; 24: 131–136.
Yamazawa K, Kagami M, Ogawa M, Horikawa R, Ogata T : Placental hypoplasia in maternal uniparental disomy for chromosome 7. Am J Med Genet A 2008; 146: 514–516.
Tsai CE, Lin SP, Ito M, Takagi N, Takada S, Ferguson-Smith AC : Genomic imprinting contributes to thyroid hormone metabolism in the mouse embryo. Curr Biol 2002; 12: 1221–1226.
Rosa AL, Wu YQ, Kwabi-Addo B, Coveler KJ, Reid Sutton V, Shaffer LG : Allele-specific methylation of a functional CTCF binding site upstream of MEG3 in the human imprinted domain of 14q32. Chromosome Res 2005; 13: 809–818.
Aretz S, Raff R, Woelfle J et al: Maternal uniparental disomy 14 in a 15-year-old boy with normal karyotype and no evidence of precocious puberty. Am J Med Genet A 2005; 135: 336–338.
Takahashi I, Takahashi T, Utsunomiya M, Takada G, Koizumi A : Long-acting gonadotropin-releasing hormone analogue treatment for central precocious puberty in maternal uniparental disomy chromosome 14. Tohoku J Exp Med 2005; 207: 333–338.
Vogel F, Motulsky AG : Human Genetics: Problems and Approaches. Heidelberg: Springer-Verlag, 1986.
Gunay-Aygun M, Schwartz S, Heeger S, O'Riordan MA, Cassidy SB : The changing purpose of Prader–Willi syndrome clinical diagnostic criteria and proposed revised criteria. Pediatrics 2001; 108: E92.
Berends MJ, Hordijk R, Scheffer H, Oosterwijk JC, Halley DJ, Sorgedrager N : Two cases of maternal uniparental disomy 14 with a phenotype overlapping with the Prader–Willi phenotype. Am J Med Genet 1999; 84: 76–79.
Mitter D, Buiting K, von Eggeling F et al: Is there a higher incidence of maternal uniparental disomy 14 [upd(14)mat]? Detection of 10 new patients by methylation-specific PCR. Am J Med Genet A 2006; 140: 2039–2049.
Acknowledgements
This study was supported by grants for Child Health and Development (20C-2) and for Research on Children and Families (H18-005) from the Ministry of Health, Labor, and Welfare, and by Grants-in-Aid for Scientific Research (Priority Areas: 16086215) from the Ministry of Education, Culture, Sports, Science and Technology.
Author information
Authors and Affiliations
Corresponding author
Additional information
Disclosure
The authors have reported no conflicts of interest.
Supplementary Information accompanies the paper on European Journal of Human Genetics website (http://www.nature.com/ejhg)
Supplementary information
Rights and permissions
About this article
Cite this article
Hosoki, K., Ogata, T., Kagami, M. et al. Epimutation (hypomethylation) affecting the chromosome 14q32.2 imprinted region in a girl with upd(14)mat-like phenotype. Eur J Hum Genet 16, 1019–1023 (2008). https://doi.org/10.1038/ejhg.2008.90
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/ejhg.2008.90
Keywords
This article is cited by
-
Metabolic syndrome and underlying genetic determinants-A systematic review
Journal of Diabetes & Metabolic Disorders (2022)
-
Kagami–Ogata syndrome: a clinically recognizable upd(14)pat and related disorder affecting the chromosome 14q32.2 imprinted region
Journal of Human Genetics (2016)
-
Epimutations of the IG-DMR and the MEG3-DMR at the 14q32.2 imprinted region in two patients with Silver–Russell Syndrome-compatible phenotype
European Journal of Human Genetics (2015)
-
Novel deletions affecting the MEG3-DMR provide further evidence for a hierarchical regulation of imprinting in 14q32
European Journal of Human Genetics (2015)

{kind=link}