
Abstract
Brain-derived neurotrophic factor (BDNF) is an essential facilitator of neuronal plasticity. By counteracting the adverse effects of excessive stress-induced glucocorticoid signaling, BDNF has been implicated as a resilience factor to psychopathology caused by chronic stress. Insights into the effects of acute stress on peripheral BDNF levels in humans are inconclusive. The short-term interplay between BDNF and cortisol in response to acute psychosocial stress remains unexplored. Furthermore, it is unknown whether mental training that is effective at reducing cortisol reactivity can also influence BDNF during acute stress. In the current study, we investigated serum BDNF levels during an acute psychosocial stress paradigm, the Trier Social Stress Test (TSST), in 301 healthy participants (178 women, mean age = 40.65) recruited as part of the ReSource Project, a large-scale mental training study consisting of three distinct 3-month training modules. Using a cross-sectional study design, we first examined the relationship between BDNF and salivary cortisol in a control group with no mental training. Subsequent analyses focused on differences in BDNF stress levels between control and mental training groups. We show that serum BDNF is indeed stress-sensitive, characterized by a significant post-stress increase and subsequent decline to recovery. While respective increases in BDNF and cortisol were not associated, we found two indications for an antagonistic relationship. Higher BDNF peaks after stress were associated with steeper cortisol recovery. On the other hand, the magnitude of the cortisol stress response was linked to steeper BDNF recovery after stress. BDNF levels were not modulated by any of the mental training modules. Providing novel evidence for the dynamics of BDNF and cortisol during acute stress, our findings may further inform research on the physiological mechanisms involved in stress chronification and the associated health risks.
Similar content being viewed by others

Introduction
Brain-derived neurotrophic factor (BDNF) is abundantly expressed in the human brain, promotes neuronal growth and differentiation, and regulates adult synaptic plasticity [1, 2]. The expression of BDNF is highly regulated by neuronal activity [3]. BDNF has been implicated in a number of stress-related psychiatric diseases, most notably in mood disorders [2, 4]. Suggesting a direct contribution of the neurotrophin to pathogenesis, the neurotrophic hypothesis of depression [5] links chronic stress to a reduction in BDNF and consequent neuronal atrophy in depression-related brain areas (e.g., the hippocampus [6]). An upregulation of BDNF through pharmacotherapy [7] has been proposed to reverse atrophy, thereby counteracting the adverse effects of excessive stress-induced glucocorticoid release [8, 9]. Research further highlights the importance of the BDNF-Tropomyosin receptor kinase B (TrkB) signaling pathway in antidepressant action and restoration of synaptic plasticity [6, 10]. Chronically increased glucocorticoid signaling has been shown to downregulate BDNF expression [11], and the modulation of BDNF procession and secretion by glucocorticoids and its receptor system were recently suggested (reviewed in [12, 13]). In addition to opposite actions on synaptic plasticity, findings from animal models reveal the close relation of BDNF and glucocorticoids by demonstrating a role of BDNF in the regulation of the hypothalamic−pituitary−adrenal (HPA) axis and stress adaptation [14,15,16,17].
While the relation of chronic stress and BDNF is rather well established, far less is known about the effects of acute stress on BDNF levels. Overall, existing research suggests that BDNF is stress-sensitive. However, magnitude and direction of acute stress-induced BDNF changes differ substantially depending on the investigated species, site and type or duration of stress-induction [18]. As opposed to chronically elevated levels, transitory increases in glucocorticoids, for example, can increase BDNF levels [18]. Mechanistic links supporting the notion that these pathways work in conjunction were provided by studies showing that BDNF receptor TrkB signaling was mediated by glucocorticoids [19]. Importantly, BDNF signaling was shown to affect glucocorticoid-receptor function [20], and further, interactions of glucocorticoid receptor and TrkB in BDNF-stimulated neurotransmitter release were recently revealed [21].
Increases in peripheral BDNF levels measured both in serum and plasma are well documented for acute controllable stress in the form of physical exercise [22, 23]. However, evidence for an effect of psychosocial stress is scarce and inconclusive. The few studies investigating levels of serum BDNF in response to psychosocial stress used varying stress inductions in small and selective samples and have produced conflicting results: While one study found increased serum BDNF levels following psychosocial stress (Trier Social Stress Test; TSST [24]) [25], decreased serum BDNF levels were reported following a paradigm combining psychological and physical stressors [26].
In sum, while evidence for opposite actions of chronic stress and BDNF on synaptic integrity is accumulating, the effect of acute psychosocial stress on peripheral BDNF levels in humans remains poorly studied and ambiguous. Regarding the association between BDNF and glucocorticoids, research suggests a complex short-term interplay depending on dose and duration of glucocorticoid signaling [18, 27]. As higher acute stress responsivity and failure to recover from elevated HPA-axis activation are associated with long-term vulnerability to stress-related clinical conditions [28], and desynchronization of BDNF-TrkB and glucocorticoid-receptor signaling pathways contribute to stress maladaptation [29], exploring the interrelation of BDNF and cortisol under acute stress exposure in humans is a valid starting point to advance our understanding of stress-coping and related resilience or vulnerability.
The first aim of this study was therefore to provide evidence on whether acute psychosocial stress, induced with the TSST, influences peripheral BDNF levels. Further, we aimed to examine the relation between stress-induced changes in BDNF and cortisol.
Contemplative mental training interventions have become popular non-pharmacological treatment options aiming to promote mental and physical wellbeing in healthy and clinical populations [30,31,32,33]. The majority of established intervention protocols (e.g. the 8-week mindfulness-based stress reduction program; MBSR [34] or Mindfulness-Based Cognitive Therapy [35]) combine diverse mental practice types targeting functions such as attention, socio-affective, and cognitive skills [36]. Our second study aim was to explore whether these distinct mental practice types differentially influence stress-induced BDNF levels. Given the dynamic linkage of BDNF with stress [27] and cortisol [18], a reduction in cortisol reactivity may be one potential mechanism by which mental training alters BDNF release. In this context, and in the current participant sample, we showed substantially reduced cortisol reactivity to the TSST after the training of socio-affective and socio-cognitive mental capacities [37].
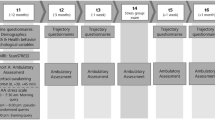
To target our research questions, 332 participants enrolled in the ReSource Project [38], a longitudinal training study involving three distinct 3-month training modules cultivating (a) Presence (present-moment focused attention and interoceptive awareness), (b) Affect (gratitude, compassion, prosocial motivation and dealing with difficult emotions) and (c) Perspective (metacognition and perspective-taking on self and others) in a within-subjects design (see Fig. 1). Based on the available literature, we hypothesized to find stress-induced increases in serum BDNF. In line with a growing body of research suggesting that the actions of glucocorticoids and BDNF are closely intertwined [12, 18], we expected to find an association of BDNF with cortisol. If the release of cortisol and BDNF following acute psychosocial stress was found to be associated, we further hypothesized mental training efficient in reducing cortisol reactivity [37, 39] to modulate BDNF reactivity. Based on our previous findings [37], the two social modules Affect and Perspective were expected to show the biggest effect on peripheral BDNF levels.

a Overview of the study design and participant numbers of the different groups in the cross-sectional stress-testing design. b Training modules and daily core exercises
Methods
Participants
All participants of the ReSource Project underwent a comprehensive mental health diagnostic interview with a trained clinical psychologist. The interview included a computer-assisted German version of the structured Clinical Interview for DSM-IV Axis-I disorders, the SCID-I DIA-X [40], and a personal interview for Axis-II disorders, the SCID-II [41, 42]. Volunteers were excluded if they fulfilled criteria for an Axis-I disorder within the past 2 years, or for schizophrenia, psychotic disorder, bipolar disorder, substance dependency or an Axis-II disorder at any time in their life. Further, medication intake influencing the HPA-axis resulted in exclusion from the study. For details on the multistep recruitment procedure, the complete list of inclusion/exclusion criteria and the sample description of the ReSource Project, see [38]. Three hundred and one participants (178 women, mean ± SD = 40.65 ± 9.3 years, 20–55 years) of the total 332 participants enrolled in the ReSource Project provided BDNF data and were consequently integrated in the present study. The missing 31 participants either dropped out of the study (n = 10; 6 women), were excluded from the study (n = 4; 3 women), were repeatedly unavailable for testing (n = 5; 3 women) or did not provide blood samples (n = 12; 8 women). Reasons for study dropout were time constraints (n = 5; 2 women) or discomfort with the experiments (n = 5; 4 women). Reasons for exclusion were medical (n = 3; 2 women) or prior mental training experience (n = 1 woman). Further sample characteristics and descriptive statistics are provided in Table S1 (Supplementary Materials). The ReSource Project was registered with the Protocol Registration System of ClinicalTrial.gov under the title “Plasticity of the Compassionate Brain” (Identifier NCT01833104). It was approved by the Research Ethics Boards of Leipzig University (ethic number: 376/12-ff) and Humboldt University Berlin (ethic numbers: 2013-20, 2013-29, 2014-10). Participants gave written informed consent, could withdraw from the study at any time and were financially compensated.
Experimental design
Each participant underwent stress testing once, at different time-points during the ReSource Project. This cross-sectional design (see Fig. 1) resulted in 124 participants (72 women) being tested without prior training (no-training-group, of which 78 participants were part of the retest control cohort and 46 were part of the training cohorts but tested at T0 prior to training onset). The remaining 177 participants were tested after completion of either one or two training modules: 44 at T1 following Presence, 45 at T1 following Affect, 44 at T2 following Presence and Affect, and 44 at T2 following Presence and Perspective training. Details on the matching of participants are available in the Supplementary Materials and Table S1.
Procedure
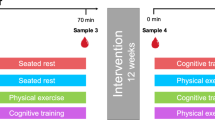
One testing session with a total duration of 130 min took place between 12 pm and 6 pm to account for potential diurnal variation in serum BDNF and glucocorticoid levels [43, 44]. A baseline blood sample for BDNF analysis was drawn 15 min after arrival (at −50 min relative to stressor onset). Following a resting period of 35 min, participants received TSST instructions (at −15 min) and, after a 15-min anticipation phase in which they additionally completed questionnaires (see Supplementary Materials), participants were exposed to the stress protocol (from 0 to 10 min). A post-stress blood sample was drawn immediately after termination of the stress test (at approx. 15 min), and a final sample after a recovery period shortly before the end of the testing session (at 60 min). A more detailed study protocol including specifics on the assessment of salivary cortisol is reported in [37]. In contrast to the previous study [37], which reported on a range of psychological and physiological stress markers, we here focused on the serum BDNF stress response and potential modulations after mental training. We calculated cortisol reactivity and recovery indices derived from the previous study (see below) to explore a potential relation to serum BDNF levels. Thus, differences in the respective study samples exist, as inclusion in the present study depended on the availability of blood serum samples.
Stress induction
Participants underwent the Trier Social Stress Test [24]. The TSST is the most frequently used psychosocial laboratory stress protocol to reliably elicit both a physiological and psychological stress response [45]. Following an anticipation phase (15 min in the current study), participants completed a mock job-interview (5 min) and a difficult mental arithmetic task (5 min) in the presence of a committee of alleged behavioral analysts. Key features for the efficacy of the TSST are socio-evaluative threat, unpredictability and uncontrollability [46].
Measures
Brain-derived neurotrophic factor (BDNF)
Blood (5.5 ml) was collected into serum vacutainers (Sarstedt), allowed to clot for 30−45 min, and subsequently centrifuged at 3500 rpm for 15 min. Serum was frozen at −80 °C until assay. BDNF concentrations in serum were determined with a quantitative sandwich enzyme immunoassay technique (R&D Systems, Inc., Minneapolis, MN, USA), using the recommended buffers, diluents and substrates. The optical density of the color reaction was read using a microtiter plate reader set at 450 nm. BDNF concentrations (in pg/ml) in each sample were calculated according to a standard curve. According to the manufacturer, the minimum detectable dose of total BDNF ranged from 0.372 to 1.35 pg/ml, with a mean value of 0.997 pg/ml. The intra- and interassay coefficients of variation of <7% were determined by duplicate analysis of >6% of randomly selected samples.
Cortisol
Saliva was sampled into Salivette collection devices (Sarstedt, Nümbrecht, Germany). Participants were instructed to place the collection swabs in their mouth and to refrain from chewing for 2 min. Salivettes were stored at −30 °C until assay (at the Department of Biological and Clinical Psychology, University of Trier, Germany). Cortisol levels (expressed in nmol/l) were determined in duplicate using a time-resolved fluorescence immunoassay [47] with intra-/interassay variabilities of <10/12%. As a measure of the magnitude of HPA-axis activation in response to stress (reactivity), a change score for cortisol increase was calculated (Δi, from baseline measurement to the average peak at 20 min after stressor onset). Representing stress recovery, a change score from the average peak to recovery at 55 min (Δr) was calculated. Stress recovery can take substantially longer than the increase to peak, is subject to comprehensive interindividual variability [48] and recognized as a major source of allostatic load [49]. Measuring cortisol levels in saliva using Salivettes is noninvasive and thus the method of choice in psycho-biological stress research [50, 51]. Saliva cortisol levels have been shown to be highly and reliably correlated to serum cortisol levels [52, 53].
ReSource training program
The ReSource Project investigated the effects of distinct mental training techniques. The training program was parceled into separate 3-month training modules (Presence, Affect, and Perspective), each cultivating distinct cognitive and socio-affective abilities [38]. Participants were divided in two 9-month training cohorts experiencing the modules in different orders, two retest control cohorts and one 3-month Affect training cohort. In detail, two training cohorts (TC1, TC2) started their training with the Presence Module. They then underwent Affect and Perspective Modules in different orders. To isolate the specific effects of the Presence Module, a third training cohort (TC3) underwent a 3-month Affect Module only (Fig. 1). Participants attended 3-day retreats before each training module. After the retreat, they met weekly for 2 h with their teachers in groups. On the remaining days, they practiced alone for approximately 30 min via cellphone-app or internet platform. The two core daily exercises varied depending on a given training module.
As illustrated in Fig. 1b, the processes targeted in the Presence Module are attention and interoceptive awareness, which are trained through the two meditation-based core exercises Breathing Meditation and Body Scan. The Affect Module targets the cultivation of social emotions such as compassion, loving kindness, and gratitude. It further aims to enhance prosocial motivation and dealing with difficult emotions. The two core exercises trained during the Affect Module are Loving-kindness Meditation and Affect Dyad [54]. In the Perspective Module participants trained metacognition and perspective-taking on self and others through the two core exercises Observing-thoughts Meditation and Perspective Dyad. Both dyadic exercises represent a form of contemplative dialogue or “loud meditation” and are realized in pairs of two. For details on the ReSource training modules, including specifics on training times and core exercises of each module, see [38].
Statistical analysis
Linear mixed effects models were fitted to accommodate the hierarchical data structure in which repeated measures were nested within subjects. In total, three multilevel models with random intercepts were calculated to test our research questions. The first two models were fitted only to the no-training group and tested for an effect of acute stress on repeated measures of BDNF (Model 1) as well as associations of stress-reactive BDNF with salivary cortisol (Model 2). Model 3 tested the effects of the specific mental training groups (no training, Presence, Affect, Presence/Affect, Presence/Perspective). All models included age and sex as covariates. Further information on the model-building process including specific model equations are provided in the Supplementary Materials. All analyses were performed using R version 3.5.1 [55] and package lme4 [56]. Significance was set to a level of p < 0.05. To account for skewedness, physiological data were ln-transformed before analysis.
Results
Participant numbers per group, as well as the available number of serum BDNF samples and cortisol reactivity/recovery measures are available in Table S2 (Supplementary Materials).
In an initial reliability check in the no-training group, the efficacy of the stress protocol in inducing marked physiological stress responses was confirmed. We observed cortisol increases of over 1.5 nmol/l, which is conventionally defined as a substantial cortisol response [57], in 75% of participants (also see [37]; minor deviations in results are due to slight divergence between the respective study samples).
Effects of acute stress on BDNF levels (no-training group)
As depicted in Fig. 2, BDNF levels significantly increased following acute stress (main effect of slope to peak; β30 = 0.001, t = 3.05, p = 0.003) and showed a significant decrease back to baseline afterwards (main effect of slope to recovery; β40 = −0.002, t = −4.85, p < 0.001). Higher age was associated with relatively increased peak BDNF levels (β02 = 0.006, t = 2.48, p = 0.014). Sex was unassociated with BDNF levels (p > 0.6). Table 1 displays the estimates of Model 1 parameters including covariates.

BDNF measures (pg/ml) collected at −50 min (baseline), +15 min (post-stress) and +60 min (recovery) relative to TSST onset at 0 min
Association of stress-reactive BDNF with physiological and psychological markers of stress (no-training group)
Model 2 explored the association between BDNF and cortisol activity during the TSST and revealed associations of BDNF with cortisol change scores (Table 2): Higher peak BDNF levels were associated with a steeper cortisol recovery slope (F174 = 12.31, p < 0.001), and a steeper BDNF decline (slope to recovery) was associated with higher cortisol increases after stress (F171 = 4.45, p = 0.036). BDNF reactivity (slope to peak) was not associated with cortisol increase. As in Model 1, age was associated with peak BDNF levels (p = 0.015) while sex was not (p = 0.13).
Effects of mental training on stress-reactive BDNF levels (all groups)
Model 3 tested for effects of specific training groups on BDNF levels during stress. As depicted in Table 3, the factor training group did not interact with BDNF peak levels (F504 = 0.73, p > 0.5), slope to peak (F570 = 0.78, p > 0.5) or slope to recovery (F569 = 1.58, p > 0.1). This implies that specific training groups (tested after single modules or combinations of modules) did not significantly differ from the no-training group regarding BDNF levels (peak, increase or recovery).
Discussion
The relation of chronic stress and basal levels of the neurotrophin BDNF has received growing attention [7, 58]. However, evidence for the effects of acute psychosocial stress on BDNF levels is rare, and the reciprocal interaction of BDNF with cortisol remains poorly understood. Furthermore, no study has investigated whether contemplative mental training, found to efficiently decrease stress-induced cortisol reactivity [37], also affects peripheral BDNF levels following an acute psychosocial stress paradigm.
In a large sample of healthy participants, we show an increase in serum BDNF following acute psychosocial stress induction. While increases in serum BDNF and salivary cortisol were unassociated, there was evidence for a negative relation of BDNF with cortisol: higher overall levels of BDNF were linked to a steeper cortisol recovery slope, and a steeper BDNF recovery slope (i.e., a faster return of stress-reactive BDNF to baseline levels) was linked to higher cortisol stress reactivity. These results demonstrate a dynamic interaction of BDNF and cortisol following a single psychosocial stress exposure in humans, and mirror previous accounts on their long-term antagonism. Stress-reactive BDNF levels were uninfluenced by distinct mental training practices. This null result is in correspondence with the finding of uncorrelated reactivity of BDNF and cortisol in suggesting that—although both measures show reciprocal dynamics during acute stress—independent mechanisms of action drive their respective initial release.
Consistent with our findings, increased serum BDNF levels after acute psychosocial stress have been found in a small sample of 14 alcohol dependent and 10 healthy male adults [25]. Likewise, acute physical exercise is a reliable trigger of serum BDNF release [22]. However, a recent study found a post-stress decrease in serum BDNF levels in a mixed-sex sample of 68 young adults [26]. Besides differences in sample size and characteristics, we attribute these conflicting results to methodical discrepancies between studies, particularly regarding the utilized stress-inductions. While both our and Meng’s study [25] employed a classical psychosocial stress task, the TSST [24], Sharma et al. [26] employed a combination of psychosocial and physical stressor. Although the paradigm used in the latter study was of considerable duration (30 min), it may have lacked the necessary strength to elicit a substantial stress response, which is conventionally affirmed by a cortisol surge of at least 1.5 nmol/l from baseline [57]. Overall, given our considerably larger sample size and successful stress induction, the current study offers strong evidence for the sensitivity of serum BDNF levels to acute psychosocial stress induction in healthy adults.
We found associations of BDNF and cortisol levels during stress. Contrary to our hypothesis, however, BDNF reactivity was unrelated to cortisol reactivity. Instead, higher overall BDNF levels (measured in terms of the BDNF peak at 20 min after stressor onset) were associated with a steeper cortisol recovery slope. Also, a steeper BDNF decline (i.e., return of stress-reactive BDNF levels to baseline) was associated with higher cortisol stress reactivity. These associations are in line with accounts suggesting BDNF and glucocorticoids to act in conjunction during stress [12, 14, 18]. Specifically, inverse associations between the two markers can be hypothesized based on findings in which sustained or chronic stress decreased BDNF levels [59]. We suggest that individuals displaying a steeper cortisol recovery after acute stress experience are generally less prone to suffer from the effects of excessive long-term cortisol signaling, including the potential suppression of BDNF levels. The ability to adequately terminate an acute stress response and to swiftly recover cortisol to resting levels is crucial in preventing sustained activation of the stress-system and thus highly adaptive in reducing vulnerability to stress-related disease [60]. Interestingly, it was not the increase in BDNF, but rather the BDNF peak—which, in our model, is indicative of overall BDNF levels—that was linked to a steeper cortisol stress recovery. Thus, our finding may reflect the proposed long-term antagonism of BDNF and chronic stress: individuals displaying a “healthy” stress response (i.e., a steep recovery following stress-induced cortisol release) show higher levels of BDNF.
Alternatively, the found association could denote an influence of BDNF on the reduction of HPA-axis activation after stress. The pivotal role of the hippocampus in regulating HPA-axis activation, especially in terminating the stress response, is well established [61, 62]. Animal models have further specified a role of hippocampal BDNF in the proper regulation of acute stress responses by showing that adult-born hippocampal neurons were required for normal stress regulation [15], and that hippocampal BDNF expression fostered resilience to chronic stress [16]. Since peripheral BDNF levels seem to approximate levels in the brain ([63], but also see below for a more detailed discussion of this issue), the present finding of higher serum BDNF relating to steeper cortisol recovery may indicate synaptic integrity and proper functioning of stress-regulatory brain areas [64]. Research suggests that a deficient cortisol recovery is associated with higher chronic stress [65], accumulation of allostatic load [49, 66], and associated health risks such as depression [67]. Our finding may hint at the potential value of acute stress-reactive serum BDNF levels as a diagnostic risk marker for stress-chronification.
The second association found between BDNF and cortisol again confirms an inverse relation of the two markers: a higher stress-induced cortisol increase was associated with a steeper post-stress BDNF decline. We interpret this association as compliant with the previously found inverse dynamic of both measures and the long-term antagonistic relationship of BDNF and cortisol. While we are aware that the potentially involved molecular mechanisms of the acute crosstalk between BDNF and cortisol are incompletely understood [12, 18], it could be speculated that the association of higher stress-induced cortisol release with steeper post-stress BDNF decline may indicate a direct regulatory influence of circulating cortisol on BDNF recovery levels. Most robustly, glucocorticoids have been found to downregulate BDNF gene expression [58]. Suri and Vaidya [13] further suggest that glucocorticoids may exert regulatory effects on BDNF at several sites and different levels of BDNF pathways including synthesis and secretion. Findings show that BDNF protein levels rapidly respond to stress [27, 68] and significant changes in both BDNF and cortisol are detectable in the periphery within 20 min after intervention onset [22, 69]. In line with these accounts, it may be conceivable that we observed a direct downregulation of BDNF through elevated post-stress cortisol levels.
So far, the only studies indirectly linking BDNF and cortisol in an acute stress-setting in humans were investigations of the BDNF Val66Met gene polymorphism [70,71,72,73]. In detail, different genotypes—implicating differences in activity-dependent BDNF trafficking and secretion [74]—were shown to modulate cortisol responses to psychosocial stressors. In the present study, we extend this evidence by showing an association of peripheral levels of both BDNF and cortisol during acute stress. Interestingly, our exploratory analyses (Supplementary Materials) reveal that serum BDNF levels were unassociated with alpha-amylase, heart rate (HR), high frequency heart rate variability (HF-HRV) or subjective-psychological stress. Regarding alpha-amylase, which we used as proxy of sympathetic adrenomedullary (SAM) reactivity, this extends prior research showing that the BDNF Val66Met polymorphism did not interact with alpha-amylase release during the TSST [73]. Overall, HPA-axis reactivity measured in terms of cortisol release seems to be most closely connected to BDNF secretory pathways.
Based on the assumption of related stress-reactivity in cortisol and BDNF and previous findings showing specific reductions in cortisol reactivity (but not recovery) after the ReSource modules Affect and Perspective [37], we expected to find according modulations of BDNF following mental training. Surprisingly, the training groups did not differ in post-stress BDNF release or recovery. To the best of our knowledge, no other study has so far addressed the question of whether acute stress-induced BDNF release is modulated by different types of contemplative mental practices. In fact, only one study using contemplative mental training interventions has examined training effects on basal BDNF levels. In this study [75], healthy volunteers took part in a 3-month retreat. Next to mindfulness-based mental training with a focus on body and breath, participants practiced yoga and underwent a strict dietary change. While basal serum BDNF levels were found to increase throughout the retreat, effects cannot be attributed to the mental training per se, since specifically physical exercise has been shown to robustly modulate the availability of BDNF in blood [23]. Regarding acute stress-reactive BDNF levels, we conclude that serum BDNF changes are not sensitive to mental training interventions focusing on present-moment attention, socio-affective or socio-cognitive capacities. Although our findings show a reciprocal link of peripheral BDNF and cortisol levels, the fact that respective post-stress increases were not associated suggests independent dynamics of respective release mechanisms. While it is an inherent claim of many mental training protocols to foster stress resilience by providing emotional, motivational or cognitive capacities relevant in (psychological) stress-coping, BDNF release during acute stress seems less susceptible to changes in these psychological skills. The absence of an association of BDNF and subjective-psychological stress experience (see Table S3, Supplementary Materials) further supports this notion.
Our study is subject to some limitations. A more frequent assessment of serum BDNF would have been beneficial to acquire a fine-grained notion of the stress-induced dynamics of BDNF and its relation to cortisol. Although we most likely accurately captured the BDNF peak with our post-stress measurement [25, 76], more detailed insights into the post-stress recovery phase of BDNF would have been informative. As Vega et al. [77] suggest that increased serum BDNF levels can return to resting levels within 15 min after intervention, interindividual differences in the circadian rhythm of peripheral BDNF [30] might have added additional variance to our recovery BDNF measurement. Lastly, it remains speculative to which extent the observed changes in serum BDNF also apply to central BDNF. Several studies show large variation of BDNF in different brain sites [27]. Others suggest that at least hippocampal BDNF translated to peripheral BDNF in different species [78, 79], and that human serum BDNF levels correlated with other markers of cortical integrity [80]. For obvious reasons, human studies on the in vivo relation of central and peripheral BDNF levels are rare. In accordance with the assumption that peripheral BDNF reflects central levels of the neurotrophin, however, Pillai et al. [63] showed moderate correlations of BDNF protein levels in cerebrospinal fluid and plasma. Regarding the relation of salivary and central levels of cortisol, several findings suggest a meaningful association: salivary cortisol levels relate to total and free serum cortisol levels [50, 53] and further, high correlations of peripheral (plasma) and cerebrospinal fluid cortisol concentrations were reported [81]. Cortisol readily enters the brain mainly via passive diffusion [82] where central feedback loops are essential for regulation of HPA-axis activity [62]. However, actions of cortisol are further determined by receptor-mediated sensitivity for cortisol signaling [50], which was not assessed in the present study but may be particularly relevant in terms of vulnerability or resilience to stress-induced disease [28, 29]. Future studies, preferentially in vulnerable populations, should determine the predictive value of acute, stress-reactive BDNF-cortisol associations in relation to pathogenesis.
In summary, we demonstrate in a large healthy adult population that serum BDNF levels are stress-sensitive. We also provide novel evidence on the calibration of peripheral BDNF and cortisol during acute psychosocial stress. Our findings indicate an acute inverse relationship, consistent with the proposed long-term antagonism of BDNF and chronic stress (and concomitant cortisol release). Future research addressing the involved molecular mechanisms is critically needed, particularly to elucidate potential acute regulatory influences of BDNF and cortisol onto each other. Yet, our results suggest that measuring peripheral BDNF levels during acute stress may be informative in assessing HPA-axis functioning. While increasing evidence on the interplay of BDNF and cortisol pathways suggests reciprocal influences on several levels and differing timescales [29, 83], the present findings clarify their acute relation on the peripheral level. The current results may thus contribute to our understanding of how stress responses, mediated by both BDNF and cortisol, can turn from adaptive to pathologic.
Funding and disclosure
TS as the principal investigator received funding for the ReSource Project from the European Research Council (ERC) under the European Community’s Seventh Framework Programme (FP7/2007–2013) ERC grant agreement number 205557 and the Max Planck Society. The authors declare no competing interests.
Data availability
The datasets generated and/or analyzed for the current study are not publicly available due to ongoing analyses in the context of the large-scale ReSource Project. The data are available upon request for replication purposes.
References
Huang EJ, Reichardt LF. Neurotrophins: roles in neuronal development and function. Annu Rev Neurosci. 2001;24:677–736.
Lu B, Nagappan G, Lu Y. BDNF and synaptic plasticity, cognitive function, and dysfunction. In: Lewin GR, Carter BD, editors. Neurotrophic factors. Berlin, Heidelberg: Springer Berlin Heidelberg; 2014. p. 223–50.
Thoenen H. The changing scene of neurotrophic factors. Trends Neurosci. 1991;14:165–70.
Autry AE, Monteggia LM. Brain-derived neurotrophic factor and neuropsychiatric disorders. Pharmacol Rev. 2012;64:238–58.
Duman RS, Monteggia LM. A neurotrophic model for stress-related mood disorders. Biol Psychiatry. 2006;59:1116–27.
Castrén E, Rantamäki T. The role of BDNF and its receptors in depression and antidepressant drug action: reactivation of developmental plasticity. Dev Neurobiol. 2010;70:289–97.
Duman RS, Aghajanian GK, Sanacora G, Krystal JH. Synaptic plasticity and depression: new insights from stress and rapid-acting antidepressants. Nat Med. 2016;22:238–49.
Martinowich K, Manji H, Lu B. New insights into BDNF function in depression and anxiety. Nat Neurosci. 2007;10:1089–93.
McEwen BS. Protective and damaging effects of stress mediators: central role of the brain. Dialogues Clin Neurosci. 2006;8:367.
Björkholm C, Monteggia LM. BDNF—a key transducer of antidepressant effects. Neuropharmacology. 2016;102:72–9.
Duman RS, Heninger GR, Nestler EJ. A molecular and cellular theory of depression. Arch Gen Psychiatry. 1997;54:597–606.
Numakawa T, Odaka H, Adachi N. Actions of brain-derived neurotrophic factor and glucocorticoid stress in neurogenesis. Int J Mol Sci. 2017;18:2312.
Suri D, Vaidya V. Glucocorticoid regulation of brain-derived neurotrophic factor: relevance to hippocampal structural and functional plasticity. Neuroscience. 2013;239:196–213.
Jeanneteau FD, Lambert WM, Ismaili N, Bath KG, Lee FS, Garabedian MJ, Chao MV. BDNF and glucocorticoids regulate corticotrophin-releasing hormone (CRH) homeostasis in the hypothalamus. Proc Natl Acad Sci USA. 2012;109:1305–10.
Snyder JS, Soumier A, Brewer M, Pickel J, Cameron HA. Adult hippocampal neurogenesis buffers stress responses and depressive behaviour. Nature. 2011;476:458–61.
Taliaz D, Loya A, Gersner R, Haramati S, Chen A, Zangen A. Resilience to chronic stress is mediated by hippocampal brain-derived neurotrophic factor. J Neurosci. 2011;31:4475–83.
Tapia-Arancibia L, Rage F, Givalois L, Arancibia S. Physiology of BDNF: focus on hypothalamic function. Front Neuroendocrinol. 2004;25:77–107.
Jeanneteau FD, Chao MV. Are BDNF and glucocorticoid activities calibrated?. Neuroscience. 2013;239:173–95.
Jeanneteau FD, Garabedian MJ, Chao MV. Activation of Trk neurotrophin receptors by glucocorticoids provides a neuroprotective effect. Proc Natl Acad Sci USA. 2008;105:4862–7.
Lambert WM, Xu CF, Neubert TA, Chao MV, Garabedian MJ. & Jeanneteau FD. Brain-derived neurotrophic factor signaling rewrites the glucocorticoid transcriptome via glucocorticoid receptor phosphorylation. Mol Cell Biol. 2013;33:3700–14.
Numakawa T, Kumamaru E, Adachi N, Yagasaki Y, Izumi A, & Kunugi H. Glucocorticoid receptor interaction with TrkB promotes BDNF-triggered PLC-γ signaling for glutamate release via a glutamate transporter. Proc Natl Acad Sci USA. 2009;106:647–52.
Rojas Vega S, Strüder HK, Wahrmann BV, Schmidt A, Bloch W, & Hollmann W. Acute BDNF and cortisol response to low intensity exercise and following ramp incremental exercise to exhaustion in humans. Brain Res. 2006;1121:59–65.
Szuhany KL, Bugatti M, Otto MW. A meta-analytic review of the effects of exercise on brain-derived neurotrophic factor. J Psychiatr Res. 2015;60:56–64.
Kirschbaum C, Pirke KM, Hellhammer DH. The Trier Social Stress Test—a tool for investigating psychobiological stress responses in a laboratory setting. Neuropsychobiology. 1993;28:76–81.
Meng D, Wu T, Rao U, North CS, Xiao H, Javors MA, & Adinoff B. Serum NPY and BNDF response to a behavioral stressor in alcohol-dependent and healthy control participants. Psychopharmacology. 2011;218:59.
Sharma S, Graham R, Rohde R, & Ceballos N.A. Stress-induced change in serum BDNF is related to quantitative family history of alcohol use disorder and age at first alcohol use. Pharmacol Biochem Behav. 2017;153:12–17.
Gray J, Milner T, McEwen B. Dynamic plasticity: the role of glucocorticoids, brain-derived neurotrophic factor and other trophic factors. Neuroscience. 2013;239:214–27.
Holsboer F, Ising M. Stress hormone regulation: biological role and translation into therapy. Annu Rev Psychol. 2009;61:81–109.
Jeanneteau F, Borie A, Chao MV, Garabedian MJ. Bridging the gap between BDNF and glucocorticoid effects on brain networks. Neuroendocrinology. 2018. https://doi.org/10.1159/000496392. PMID: 30572337.
Cain SW, Chang AM, Vlasac I, Tare A, Anderson C, Czeisler CA, & Saxena R. Circadian rhythms in plasma brain-derived neurotrophic factor differ in men and women. J Biol Rhythms. 2017;32:75–82.
Galante J, Galante I, Bekkers MJ, & Gallacher J. Effect of kindness-based meditation on health and well-being: a systematic review and meta-analysis. Journal of consulting and clinical psychology. 2014;82:1101.
Grossman P, Niemann L, Schmidt S, & Walach H. Mindfulness-based stress reduction and health benefits: a meta-analysis. Journal of psychosomatic research. 2004;57:35–43.
Khoury B, Sharma M, Rush SE, & Fournier C. Mindfulness-based stress reduction for healthy individuals: a meta-analysis. Journal of psychosomatic research. 2015;78:519–28.
Kabat-Zinn J. Wherever you go, there you are: Mindfulness meditation in everyday life. New York: Hyperion; 1994.
Segal ZV, Williams JMG, Teasdale JD. Mindfulness-based cognitive therapy for depression. New York, NY, US: Guilford Press; 2012.
Dahl CJ, Lutz A, Davidson RJ. Reconstructing and deconstructing the self: cognitive mechanisms in meditation practice. Trends Cogn Sci. 2015;19:515–23.
Engert V, Kok BE, Papassotiriou I, Chrousos GP, & Singer T. Specific reduction in cortisol stress reactivity after social but not attention-based mental training. Sci Adv. 2017;3:e1700495.
Singer T, Kok BE, Bornemann B, Zurborg S, Bolz M, & Bochow C. The ReSource Project: background, design, samples, and measurements. Leipzig: Max Planck Institute for Human Cognitive and Brain Sciences; 2016.
Fan Y, Tang YY, Posner MI. Cortisol level modulated by integrative meditation in a dose‐dependent fashion. Stress Health. 2014;30:65–70.
Wittchen HU, Pfister H. DIA-X-Interviews: Manual für Screening-Verfahren und Interview. Frankfurt: Swets & Zeitlinger; 1997.
First MB, Gibbon M, Spitzer RL, Williams JBW, Benjamin LS. Structured clinical interview for DSM-IV Axis II personality disorders, (SCID-II). Washington, DC: American Psychiatric Press, Inc; 1997.
Wittchen HU, Zaudig M, Fydrich T. SKID—Strukturiertes Klinisches Interview für DSM-IV. Achse I und Achse II. Göttingen: Hogrefe; 1997.
Bus BAA, Molendijk ML, Penninx BJWH, Buitelaar JK, Kenis G, Prickaerts J, & Voshaar RO. Determinants of serum brain-derived neurotrophic factor. Psychoneuroendocrinology. 2011;36:228–39.
Piccinni A, Marazziti D, Del Debbio A, Bianchi C, Roncaglia I, Mannari C, & Dell'Osso L. Diurnal variation of plasma brain‐derived neurotrophic factor (BDNF) in humans: an analysis of sex differences. Chronobiol Int. 2008;25:819–26.
Allen AP, Kennedy PJ, Cryan JF, Dinan TG, & Clarke G. Biological and psychological markers of stress in humans: focus on the Trier Social Stress Test. Neurosci Biobehav Rev. 2014;38:94–124.
Dickerson SS, Kemeny ME. Acute stressors and cortisol responses: a theoretical integration and synthesis of laboratory research. Psychol Bull. 2004;130:355–91.
Dressendorfer RA, Dressendörfer RA, Kirschbaum C, Rohde W, Stahl F, & Strasburger CJ. Synthesis of a cortisol-biotin conjugate and evaluation as a tracer in an immunoassay for salivary cortisol measurement. J Steroid Biochem Mol Biol. 1992;43:683–92.
Kudielka BM, Wüst S. Human models in acute and chronic stress: assessing determinants of individual hypothalamus–pituitary–adrenal axis activity and reactivity. Stress. 2010;13:1–14.
McEwen BS, Seeman T. Protective and damaging effects of mediators of stress: elaborating and testing the concepts of allostasis and allostatic load. Ann NY Acad Sci. 1999;896:30–47.
Hellhammer DH, Wüst S, Kudielka BM. Salivary cortisol as a biomarker in stress research. Psychoneuroendocrinology. 2009;34:163–71.
Kirschbaum C, Hellhammer DH. Salivary cortisol in psychoneuroendocrine research: recent developments and applications. Psychoneuroendocrinology. 1994;19:313–33.
Lippi G, Dipalo M, Buonocore R, Gnocchi C, Aloe R, & Delsignore R. Analytical evaluation of free testosterone and cortisol immunoassays in saliva as a reliable alternative to serum in sports medicine. J Clin Lab Anal. 2016;30:732–5.
Poll E-M, Kreitschmann-Andermahr I, Langejuergen Y, Stanzel S, Gilsbach JM, Gressner A, & Yagmur E. Saliva collection method affects predictability of serum cortisol. Clin Chim Acta. 2007;382:15–19.
Kok BE, Singer T. Effects of contemplative dyads on engagement and perceived social connectedness over 9 months of mental training: a randomized clinical trial. JAMA Psychiatry. 2017;74:126–34.
R Core Team. R: a language and environment for statistical computing. R Foundation for Statistical Computing. Vienna, Austria. 2018. https://www.R-project.org.
Bates D, Maechler M, Bolker B, Walker S. lme4: Linear mixed-effects models using Eigen and S4. R package version 1.1–7. 2014 http://CRAN.R-project.org/package=lme4.
Miller R, Plessow F, Kirschbaum C, & Stalder T. Classification criteria for distinguishing cortisol responders from nonresponders to psychosocial stress: evaluation of salivary cortisol pulse detection in panel designs. Psychosom Med. 2013;75:832–40.
Egeland M, Zunszain PA, Pariante CM. Molecular mechanisms in the regulation of adult neurogenesis during stress. Nat Rev Neurosci. 2015;16:189.
Gold P. The organization of the stress system and its dysregulation in depressive illness. Mol psychiatry. 2015;20:32–47.
De Kloet ER, Joëls M, Holsboer F. Stress and the brain: from adaptation to disease. Nat Rev Neurosci. 2005;6:463.
Herman JP, Figueiredo H, Mueller NK, Ulrich-Lai Y, Ostrander MM, Choi DC, & Cullinan WE. Central mechanisms of stress integration: hierarchical circuitry controlling hypothalamo–pituitary–adrenocortical responsiveness. Front Neuroendocrinol. 2003;24:151–80.
Ulrich-Lai YM, Herman JP. Neural regulation of endocrine and autonomic stress responses. Nat Rev Neurosci. 2009;10:397.
Pillai A, Kale A, Joshi S, Naphade N, Raju MSVK, Nasrallah H, & Mahadik SP. Decreased BDNF levels in CSF of drug-naive first-episode psychotic subjects: correlation with plasma BDNF and psychopathology. Int J Neuropsychopharmacol. 2010;13:535–9.
McEwen BS, Gray JD, Nasca C. Recognizing resilience: Learning from the effects of stress on the brain. Neurobiol Stress. 2015;1:1–11.
Matthews KA, Gump BB, Owens JF. Chronic stress influences cardiovascular and neuroendocrine responses during acute stress and recovery, especially in men. Health Psychol. 2001;20:403.
Sapolsky RM, Romero LM, Munck AU. How do glucocorticoids influence stress responses? Integrating permissive, suppressive, stimulatory, and preparative actions. Endocr Rev. 2000;21:55–89.
Burke HM, Davis MC, Otte C, & Mohr DC. Depression and cortisol responses to psychological stress: a meta-analysis. Psychoneuroendocrinology. 2005;30:846–56.
Marmigère F, Givalois L, Rage F, Arancibia S, & Tapia‐Arancibia L. Rapid induction of BDNF expression in the hippocampus during immobilization stress challenge in adult rats. Hippocampus. 2003;13:646–55.
Tang SW, Chu E, Hui T, Helmeste D, & Law C. Influence of exercise on serum brain-derived neurotrophic factor concentrations in healthy human subjects. Neurosci Lett. 2008;431:62–5.
Armbruster D, Müller-Alcazar A, Strobel A, Lesch KP, Kirschbaum C, & Brocke B. BDNF val 66 met genotype shows distinct associations with the acoustic startle reflex and the cortisol stress response in young adults and children. Psychoneuroendocrinology. 2016;66:39–46.
Jiang R, Babyak MA, Brummett BH, Siegler IC, Kuhn CM, & Williams RB. Brain-derived neurotrophic factor (BDNF) Val66Met polymorphism interacts with gender to influence cortisol responses to mental stress. Psychoneuroendocrinology. 2017;79:13–9.
Shalev I, Lerer E, Israel S, Uzefovsky F, Gritsenko I, Mankuta D, & Kaitz M. BDNF Val66Met polymorphism is associated with HPA axis reactivity to psychological stress characterized by genotype and gender interactions. Psychoneuroendocrinology. 2009;34:382–8.
Tsuru J, Tanaka Y, Ishitobi Y, Maruyama Y, Inoue A, Kawano A, & Masuda K. Association of BDNF Val66Met polymorphism with HPA and SAM axis reactivity to psychological and physical stress. Neuropsychiatr Dis Treat. 2014;10:2123.
Egan MF, Kojima M, Callicott JH, Goldberg TE, Kolachana BS, Bertolino A, & Lu B. The BDNF val66met polymorphism affects activity-dependent secretion of BDNF and human memory and hippocampal function. Cell. 2003;112:257–69.
Cahn BR, Goodman MS, Peterson CT, Maturi R, & Mills PJ. Yoga, meditation and mind-body health: increased BDNF, cortisol awakening response, and altered inflammatory marker expression after a 3-month yoga and meditation retreat. Front Hum Neurosci. 2017;11:315.
Gold SM, Schulz KH, Hartmann S, Mladek M, Lang UE, Hellweg R, & Heesen C. Basal serum levels and reactivity of nerve growth factor and brain-derived neurotrophic factor to standardized acute exercise in multiple sclerosis and controls. J Neuroimmunol. 2003;138:99–105.
Vega SR, Strüder HK, Wahrmann BV, Schmidt A, Bloch W, & Hollmann W. Acute BDNF and cortisol response to low intensity exercise and following ramp incremental exercise to exhaustion in humans. Brain Res. 2006;1121:59–65.
Klein AB, Williamson R, Santini MA, Clemmensen C, Ettrup A, Rios M, & Aznar S. Blood BDNF concentrations reflect brain-tissue BDNF levels across species. Int J Neuropsychopharmacol. 2011;14:347–53.
Sartorius A, Hellweg R, Litzke J, Vogt M, Dormann C, Vollmayr B, & Gass P. Correlations and discrepancies between serum and brain tissue levels of neurotrophins after electroconvulsive treatment in rats. Pharmacopsychiatry. 2009;42:270–6.
Lang UE, Hellweg R, Seifert F, Schubert F, & Gallinat J. Correlation between serum brain-derived neurotrophic factor level and an in vivo marker of cortical integrity. Biol psychiatry. 2007;62:530–5.
Kancheva R, Hill M, Novak Z, Chrastina J, Kancheva L, & Starka L. Neuroactive steroids in periphery and cerebrospinal fluid. Neuroscience. 2011;191:22–7.
Mason BL, Pariante CM, Jamel S, & Thomas SA. Central nervous system (CNS) delivery of glucocorticoids is fine-tuned by saturable transporters at the blood-CNS barriers and nonbarrier regions. Endocrinology. 2010;151:5294–305.
Arango-Lievano M, Lambert WM, Bath KG, Garabedian MJ, Chao MV, & Jeanneteau, F. Neurotrophic-priming of glucocorticoid receptor signaling is essential for neuronal plasticity to stress and antidepressant treatment. Proc Natl Acad Sci USA. 2015;112:15737.
Acknowledgements
We are thankful to the members of the Social Neuroscience Department involved in the ReSource Project over many years, in particular to Astrid Ackermann, Christina Bochow, Matthias Bolz, and Sandra Zurborg for managing the large-scale longitudinal study, to Elisabeth Murzik, Nadine Otto, Sylvia Tydecks, and Kerstin Träger for help with recruiting and data archiving, to Henrik Grunert for technical assistance, and to Hannes Niederhausen and Torsten Kästner for data management. Thank you also to the research assistants and students, especially Anna Koester, whose help with data collection was indispensable.
Author information
Authors and Affiliations
Contributions
TS initiated and developed the ReSource Project and secured all funding with the exception of the BDNF analysis. TS and VE designed the experiment. Data collection and statistical analyses were performed by RL and VE. GPC funded and IP, FA and EM performed the BDNF assay. RL drafted and all authors contributed to writing the manuscript and approved its final version for submission.
Corresponding author
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
About this article

Cite this article
Linz, R., Puhlmann, L.M.C., Apostolakou, F. et al. Acute psychosocial stress increases serum BDNF levels: an antagonistic relation to cortisol but no group differences after mental training. Neuropsychopharmacol. 44, 1797–1804 (2019). https://doi.org/10.1038/s41386-019-0391-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41386-019-0391-y
This article is cited by
-
Interplay between stress, sleep, and BDNF in a high-risk sample of young adults
Scientific Reports (2023)
-
Essential amino acid supplements ingestion has a positive effect on executive function after moderate-intensity aerobic exercise
Scientific Reports (2023)
-
Intricate role of sleep deprivation in modulating depression: focusing on BDNF, VEGF, serotonin, cortisol, and TNF-α
Metabolic Brain Disease (2023)
-
Clustering suicidal phenotypes and genetic associations with brain-derived neurotrophic factor in patients with substance use disorders
Translational Psychiatry (2021)
-
Circulating neurotrophins and hemostatic risk factors of atherothrombotic cardiovascular disease at baseline and during sympathetic challenge: the SABPA study
Scientific Reports (2021)
