
Abstract
Hypertension is considered to be the most important risk factor in the development of cardiovascular disease. An increasing body of evidence suggests that oxidative stress, which results in an excessive generation of reactive oxygen species (ROS), has a key role in the pathogenesis of hypertension. The modulation of the vasomotor system involves ROS as mediators of vasoconstriction induced by angiotensin II, endothelin-1 and urotensin-II, among others. The bioavailability of nitric oxide (NO), which is a major vasodilator, is highly dependent on the redox status. Under physiological conditions, low concentrations of intracellular ROS have an important role in the normal redox signaling maintaining vascular function and integrity. However, under pathophysiological conditions, increased levels of ROS contribute to vascular dysfunction and remodeling through oxidative damage. In human hypertension, an increase in the production of superoxide anions and hydrogen peroxide, a decrease in NO synthesis and a reduction in antioxidant bioavailability have been observed. In turn, antioxidants are reducing agents that can neutralize these oxidative and otherwise damaging biomolecules. The use of antioxidant vitamins, such as vitamins C and E, has gained considerable interest as protecting agents against vascular endothelial damage. Available data support the role of these vitamins as effective antioxidants that can counteract ROS effects. This review discusses the mechanisms involved in ROS generation, the role of oxidative stress in the pathogenesis of vascular damage in hypertension, and the possible therapeutic strategies that could prevent or treat this disorder.
Similar content being viewed by others

Introduction
Hypertension is considered the most important risk factor for the occurrence of cardiovascular disease.1 Oxidative stress has gained attention as one of the fundamental mechanisms responsible for the development of hypertension. Reactive oxygen species (ROS) have an important role in the homeostasis of the vascular wall; hence, they could be part of the mechanism that leads to hypertension.2, 3, 4 Thus, increased ROS production, reduced nitric oxide (NO) levels and reduced antioxidant bioavailability were demonstrated in experimental and human hypertension. Vascular superoxide is primarily derived from nicotinamide adenine dineucleotide phosphate (NADPH) oxidase when stimulated by hormones, such as angiotensin II (AT-II), endothelin-1 (ET-1) and urotensin II. In addition, increased ROS production may be generated by mechanical stimuli on the vascular wall, which increase with hypertension. ROS-induced vasoconstriction results from increased intracellular calcium concentration, thereby contributing to the pathogenesis of hypertension.2 Vasomotor tone is dependent on a delicate balance between vasoconstrictor and vasodilator forces that result from the interaction between the components of the vascular wall and the blood, all of which can be altered by oxidative stress. These findings have stimulated interest in antihypertensive therapies targeted toward decreasing ROS generation and/or increasing NO bioavailability. This review examines the available studies that highlight the role of oxidative stress in the underlying mechanism of high blood pressure and discusses those that consider the use of antioxidants in the prevention or treatment of this disorder.
Pathophysiology of hypertension
Endothelial dysfunction
Endothelial dysfunction has been implicated in the pathophysiology of different forms of cardiovascular disease, including hypertension. Endothelial dysfunction may be defined as impairment that is characterized by a shift in the actions of the endothelium toward reduced vasodilation, a proinflammatory state, and prothrombotic settings. These events lead to a state of vascular inflammation, which may be partially mediated by ROS formed by activated mononuclear cells.
Vascular oxidative stress and hypertension
Oxidative stress constitutes a unifying mechanism of injury in many types of disease processes, and it occurs when there is an imbalance between the generation of ROS and the antioxidant defense systems in the body. The ROS family comprises many molecules that have divergent effects on cellular function. Importantly, many of these actions are associated with pathological changes observed in cardiovascular disease. The effects of ROS are mediated through redox-sensitive regulation of multiple signaling molecules and second messengers.5, 6, 7 Several studies have demonstrated excessive amounts of ROS in essential hypertensive patients and various animal models of hypertension;8, 9, 10, 11, 12 these patients and experimental animals have a diminished antioxidant status,13 which contributes to the accumulating evidence that increased vascular oxidative stress could be involved in the pathogenesis of essential hypertension.2, 3, 14 Recently, a strong association between blood pressure and some oxidative stress-related parameters was demonstrated.15 Other studies showed that mouse models that involve genetic deficiencies in ROS-generating enzymes have lower blood pressure compared with their wild-type counterparts.16, 17 In addition, in cultured vascular smooth muscle cells (VSMCs) and isolated arteries from hypertensive rats and humans, ROS production is enhanced, redox-dependent signaling is amplified, and antioxidant bioactivity is reduced.18 The beneficial effects of classical antihypertensive agents, such as β-adrenergic blockers, angiotensin I converting enzyme (ACE) inhibitors, AT-II receptor antagonists, and calcium channel blockers, may be mediated, in part, by decreasing vascular oxidative stress.19, 20
Sources of ROS in the vascular wall. A variety of enzymatic and non-enzymatic sources of ROS exist in blood vessels. The best-characterized source of ROS is NADPH oxidase. In addition to NADPH oxidase, several other enzymes may contribute to ROS generation, including xanthine oxidase, NO synthase (NOS) and mitochondrial enzymes.
NADPH oxidase: NADPH oxidase is the primary biochemical source of ROS, particularly superoxide, in the vasculature. The kidney and vasculature are rich sources of NADPH oxidase-derived ROS, which have an important role in renal dysfunction and vascular damage under pathological conditions.12, 21 This system catalyses the reduction of molecular oxygen by NADPH, which functions as an electron donor, thus generating superoxide. NADPH oxidase is upregulated in hypertension by humoral and mechanical signals. AT-II is the most studied stimulus of the upregulation of NADPH oxidase, but ET-1 and urotensin II may also participate to the activation of NADPH oxidase, thereby resulting in increased ROS. It is likely that the most well-known function of NADPH oxidase-derived superoxide is the inactivation of NO in the reaction that forms peroxynitrite, which leads to impaired endothelium dependent vasodilation and the uncoupling of endothelial NO synthase (eNOS) to produce additional superoxide.16, 22 In the vasculature, the activation of NADPH oxidase has been strongly associated with hypertension.23
Uncoupled eNOS: The primary function of eNOS is NO production, which regulates vasodilation. Nevertheless, deficiency or oxidation of L-arginine and tetrahydrobiopterin (BH4), which are two essential cofactors for eNOS action, are associated with the uncoupling of the L-arginine-NO pathway that results in the decreased formation of NO and increased eNOS-mediated generation of superoxide. NADPH oxidase is the initial source of ROS. Superoxide combines with NO, which is synthesized by eNOS, to form peroxynitrite.24 In turn, peroxynitrite oxidizes and destabilizes eNOS to produce additional superoxide.22, 25 Superoxide also leads to BH4 oxidation (in fact, BH4 is highly sensitive to oxidation), which promotes eNOS uncoupling and further ROS production.
Xanthine oxidase: Xanthine oxidase is also an important source of ROS in the vascular endothelium.23, 26 It catalyzes the last two steps of purine metabolism. During this process, oxygen is reduced to superoxide. There is evidence that suggests the involvement of this enzyme in hypertension. Spontaneously hypertensive rats demonstrated elevated levels of endothelial xanthine oxidase and increased ROS production, which is associated with increased arteriolar tone.21 In addition to effects on the vasculature, xanthine oxidase may have a function in end-organ damage in hypertension.27
Mitochondria: Mitochondria are a major source and target of ROS. Some of the superoxide produced in the intermembrane space may be carried to the cytoplasm.28 Ubiquinol or coenzyme Q produces superoxide when partially reduced (semiquinone form) and an antioxidant when fully reduced.29 Complex I produces most of the superoxide that is generated by mammalian mitochondria in vitro. Complexes II and IV are not normally significant sites of ROS production. Mild uncoupling very effectively decreases the high superoxide production that occurs from complex I. A reduction in antioxidant enzymatic activity in patients with hypertension has been reported.30
Role of the vascular wall components
The endothelium senses mechanical and hormonal stimuli. In response, it releases agents that regulate vasomotor function. There is no doubt that the endothelium has a regulatory and protective role by generating vasorelaxing substances. Under some pathophysiological circumstances, endothelium-derived vasoconstricting factors, such as ET-1, AT-II, urotensin II, superoxide anions, vasoconstrictor prostaglandins and thromboxane A2, can be released and contribute to the paradoxical vasoconstrictor effects.
VSMCs function not only in the short-term regulation of blood vessel diameter and, therefore, blood pressure but also participate in long-term adaptation via structural remodeling. ROS mediate many of these pathophysiological processes.
The adventitia can contribute to hypertension by either reducing NO bioavailability or participating in vascular remodeling through ROS.
Role of vascular hormones and factors. NO: NO has an important role as a key paracrine regulator of vascular tone. Physiologically, NO inhibits leukocyte–endothelial cell adhesion, VSMC proliferation and migration, and platelet aggregation to maintain the health of the vascular endothelium. Therefore, it has many beneficial effects. The decrease in bioavailability of NO in the vasculature reduces vasodilatory capacity and contributes to hypertension. The enzyme that catalyzes the formation of NO from oxygen and arginine is NOS, which, in fact, is an entire family of enzymes. eNOS is the predominant NOS isoform in the vessel wall. Receptor-mediated agonist stimulation leads to rapid enzyme activation. Furthermore, shear stress and allosteric modulators are important modulators of eNOS activity.31 In addition to its vasorelaxing and antiproliferative roles, NO has an important role in antagonizing the effects of AT-II, endothelins and ROS. NO diffuses as a gas to the adjacent smooth muscle in which it interacts with different receptor molecules, such as the soluble guanylyl cyclase.
The normal production of NO has a crucial role in the maintenance of the physiological conditions within the cardiovascular system. L-arginine, which is a substrate for eNOS, seems to be a promising molecule in the preservation of NO formation. However, L-arginine failed to prevent blood pressure increases and left ventricle remodeling due to chronic treatment with methyl ester of N-nitro-L-arginine, which is an inhibitor of eNOS.32 The ACE inhibitor captopril completely prevented NO-deficient hypertension without improving NOS activity. NO also has an ACE down-regulating effect. Thiols protect NO from oxidation by scavenging ROS and by forming nitrosothiols; both of these effects prolong NO half-life and the duration of NO action.33, 34
Reduced NO levels can be attributed to elevated levels of ROS. Superoxide combines with NO to form peroxynitrite that oxidizes BH4 and destabilizes eNOS to produce more superoxide22, 24, 25 thus, further enhancing the development of oxidative stress. The balance between NO and AT-II in the vasomotor centers is important for the regulation of sympathetic tone.
Renin–Angiotensin system: The renin–angiotensin system has a key role in the development of cardiovascular disease. AT-II is a potent vasoactive peptide that can be formed in vascular beds that are rich in ACE. When AT-II production increases above normal levels, it induces vascular remodeling and endothelial dysfunction in association with increases in the levels of blood pressure. As a potent activator of NADPH oxidase, AT-II contributes to the production of ROS.35, 36 In rats and mice in which hypertension is induced by AT-II infusion, the expression of NADPH oxidase subunits, oxidase activity and the generation of ROS are all increased.37 AT-II not only increases NADPH oxidase activity but also upregulates superoxide dismutase activity, possibly to compensate the increased levels of ROS. In situations where this compensatory effect is efficient, ROS levels may appear normal even under prooxidant conditions. However, when ROS production becomes overwhelming, compensatory mechanisms are inadequate, and pathophysiological consequences occur.38
Captopril and enalapril prevented increases in blood pressure in young, spontaneously hypertensive rats by inhibiting ACE. Captopril, probably because of the antioxidant role of its thiol group, had a more effective hypotensive effect than enalapril.40 In contrast, NO not only antagonizes the effects of AT-II on vascular tone, cell growth and renal sodium excretion but also downregulates the synthesis of ACE and the expression of AT1 receptors. However, ACE inhibition upregulates eNOS expression. The ability of AT-II to induce endothelial dysfunction is also due to its ability to downregulate soluble guanylyl cyclase, thereby leading to impaired NO/cGMP signaling.
Acetylcholine: In vascular vessels, acetylcholine induces endothelium-dependent dilation via the production of endothelial factors, mainly NO, which then diffuses to the underlying VSMC, in which it induces VSMC relaxation. The decrease in NO bioavailability will lead to significantly reduced acetylcholine-mediated vasodilation.39, 40 The consequence of an overall increase in ROS is a reduction in the bioavailability of NO.
ET-1: Endothelins are potent vasoconstrictor isopeptides that are produced in different vascular tissues, including the vascular endothelium. ET-1 is the main endothelin generated by the endothelium and is the most important endothelin in the cardiovascular system. When ET-1 is administered in high concentrations, it behaves as a potent vasoconstrictor that is capable of exerting an array of physiological effects, including the potential to alter arterial pressure. ET-1 mediates its effects through two receptors, ETA and ETB. The ETA receptor mediates contractions via the activation of NADPH oxidase, xanthine oxidase, lipoxygenase, uncoupled NOS and mitochondrial respiratory chain enzymes. The ETB receptor induces relaxation in endothelial cells.41 Many factors that normally stimulate ET-1 synthesis, (for example, thrombin and AT-II) also cause the release of vasodilators, such as prostacyclin (PGI2) and/or NO, which oppose the vasoconstricting function of ET-1. It has been reported that essential hypertension is characterized by increased ET-1-mediated vasoconstrictor tone, which is an effect that seems to be dependent on decreased endothelial ETB-mediated NO production that is attributable to the impaired NO bioavailability.
Urotensin-II: Urotensin-II is a potent vasoactive peptide42 and indeed is the most potent identified vasoconstrictor. It functions through the activation of NADPH oxidase. The role of urotensin-II in disease is not well understood. The constrictor response to urotensin-II appears to be variable and highly dependent on the vascular bed examined. Vasoconstriction is not its only effect because UT receptors have been found in other organs.43, 44 Urotensin-II has also been shown to function as a potent vasodilator in some isolated vessels.45
Norepinephrine: VSMC are primarily innervated by the sympathetic nervous system through adrenergic receptors. Three types of adrenoceptors are present within VSMC namely: α1, α2 and β2. Norepinephrine stimulates VSMC proliferation. In addition, over-expression of inducible NOS increases blood pressure via central activation of the sympathetic nervous system, which is mediated by an increase in oxidative stress.5
Prostaglandins: PGI2, which is another endothelium-dependent vasodilator, relaxes the VSMC. PGI2 is released in higher amounts in response to the binding of ligands, such as thrombin, arachidonic acid, histamine or serotonin. The enzymes prostaglandin H2 synthase uses arachidonic acid as a substrate to produce prostaglandin H2. Prostaglandin H2 is converted to vasoactive molecules, such as PGI2. The enzyme isoform, prostaglandin H2 synthase-2, may mediate vascular dysfunction under oxidative stress conditions. Thus, peroxynitrite inhibits the enzymatic activity of PGI2 synthase, thereby impairing PGI2-mediated vasodilation.
Homocysteine: This molecule may have an important role in the pathogenesis of essential hypertension.3 Elevated homocysteinemia diminishes the vasodilation by NO, increases oxidative stress, stimulates the proliferation of VSMC and alters the elastic properties of the vascular wall. Thus, homocysteine contributes to the elevation of blood pressure.46 Additionally, elevated homocysteine levels could lead to oxidant injury of the endothelium.3 The correction of elevated homocysteinemia by the administration of the vitamins B12, B6 and folic acid could be a useful adjuvant therapy for hypertension.3, 47 However, further controlled randomized trials are necessary to establish the efficacy of these therapeutic agents.
A hypothesis for the role of vascular oxidative stress in hypertension is depicted in Figure 1.

Schematic summary of the role of vascular oxidative stress in the pathogenesis of hypertension. ACE, angiotensin converting enzyme; AT-II, angiotensin II; AT1, type 1 angiotensin II receptor; ET-1, endothelin 1; ETA, type A endothelin receptor; UT-II, urotensin II; UTR, urotensin-II receptor; ACh, acetylcholine; NO, nitric oxide; eNOS, nitric oxide synthase; PGI2, prostacyclin; ROS, reactive oxygen species.
This review has discussed the importance of ROS in the vasculature and its relation to hypertension; however, it is important to also emphasize the evidence that hypertensive stimuli, such as high salt and AT-II, promote the production of ROS not only in the vasculature but also in the kidney and the central nervous system. In addition, each of these sites contributes either to hypertension or to the adverse sequelae of this disease.47
Role of oxidative stress in the kidney
Evidence suggests a key role for ROS in the pathophysiological processes of several renal diseases; these diseases are considered causes and consequences of hypertension. Regarding glomerular alterations, ROS mediate lipoprotein glomerulopathy and other inflammatory glomerular lesions.48 A recent study demonstrated that NADPH oxidase activation and the production of ROS through lipid raft clustering is an important molecular mechanism that triggers homocysteine-mediated oxidative injury of podocytes. This injury may represent an early event that initiates glomerulosclerosis during hyperhomocysteinemia.49 One of the underlying mechanisms of ROS-mediated tubulointerstitial injury is the exposure of tubular cells to LDL, which may result in tubulointerstitial damage due to NADPH oxidase-mediated ROS production.50 AT-II has a pivotal role not only in the progression of tubulointerstitial injury but also in obstructive nephropathy.51, 52 It activates NADPH oxidase and, subsequently, generates superoxide that leads to hypertrophy of the renal tubular cells.53
There is evidence which suggests that a high-fat diet induces renal inflammation and aggravation of blood pressure via ROS in spontaneously hypertensive rats.54 Additionally, metabolic syndrome is a risk factor for chronic kidney disease (CKD) that is at least in part independent of diabetes and hypertension and probably mediated by ROS. Moreover, the onset and maintenance of renal damage may worsen metabolic syndrome features, such as hypertension, leading to potential vicious cycles.55
There are several oxidative stress-mediated mechanisms involved in endothelial dysfunction in CKD.56 ROS are elevated in CKD and are related to endothelium-dependent vascular reactivity and systolic blood pressure.57 High ROS and increased levels of the endogenous asymmetric dimethylarginine were reported to be novel risk factors for endothelial dysfunction.58 Moreover, high levels of asymmetric dimethylarginine were reported in CKD and were associated with increased intima-media thickness and cardiovascular events.59 In renovascular hypertension, oxidative stress in the ischemic kidney has a major role in the maintenance of hypertension in two kidney-one clip rats.60
Role of oxidative stress in the central nervous system
In addition to the kidney and the vasculature itself, the sympathetic nervous system, which is regulated in the central nervous system, is involved in the pathogenesis of hypertension.61 Recent studies strongly suggest that central sympathetic outflow is increased in hypertension.62 There is also evidence that increased ROS generation in the brainstem contributes to the neural mechanisms of hypertension in hypertensive rats.63
The rostral ventrolateral medulla is the major vasomotor center and is essential for the maintenance of basal vasomotor tone.64, 65 Some findings strongly indicate that ROS in the rostral ventrolateral medulla is increased in stroke-prone spontaneously hypertensive rats and thereby contributes to the neural mechanisms of hypertension through activation of the sympathetic nervous system.64 The paraventricular nucleus of the hypothalamus is most likely also involved in the ROS-mediated neural mechanism of hypertension.60, 66 There is evidence that other regions of the brain are likewise involved in ROS-mediated hypertension. These investigations suggest that increased intracellular superoxide production in the subfornical organ is critical to the development of AT-II-induced hypertension.67
Antioxidants in hypertension
This section discusses the antihypertensive role of endogenous and exogenous antioxidants that have demonstrated an ability to alter the function of blood vessels and participate in the main redox reactions involved in the pathophysiology of hypertension.
Endogenous antioxidants
The main endogenous antioxidant enzyme systems involve superoxide dismutase, catalase and glutathione peroxidase (GSH-Px). GSH-Px reduces both hydrogen peroxide and organic hydroperoxides into water and alcohols, respectively.68, 69 Catalase catalyzes the breakdown of hydrogen peroxide to water and molecular oxygen. Superoxide dismutase is a major superoxide scavenger in humans and converts the superoxide anion to hydrogen peroxide, which is a substrate for catalase and GSH-Px.70 The decrease in enzymatic antioxidant activity in hypertensive humans and in animal models of hypertension has been widely reported, which suggests that reduced antioxidant defenses are involved in this disease.71, 72, 73, 74, 75, 76
Another endogenous antioxidant is glutathione, which displays direct antioxidant activity by providing the hydrogen at the sulfur atom to transform a free radical into a stable molecule.77 An impairment of the glutathione system in hypertension has been demonstrated.78, 79 Recent studies have demonstrated the importance of the thioredoxin system in the maintenance of a reduced redox state and in the pathophysiology of hypertension.80 It has been reported that impaired thioredoxin expression in spontaneously hypertensive rats81 and in endothelial cells negatively regulates ROS production in VSMC through thioredoxin upregulation.82 Thus, thioredoxin could be an important therapeutic target for the prevention and treatment of oxidative stress and hypertension.83, 84, 85
Recent studies have shown that the antioxidant effects of mildly elevated serum bilirubin levels may protect against diseases that are associated with oxidative stress86 through mechanisms that decrease oxidative stress and increase NO levels.87, 88 A mild increase within the physiological range of serum bilirubin concentration is negatively correlated with hypertension.89 Finally, uric acid has long been recognized for its antioxidant properties and is the most abundant scavenger of free radicals in humans. High concentrations of uric acid are associated with increased serum antioxidant capacity and reduced oxidative stress.90, 91 However, there is evidence of a potential association between the plasma concentration of uric acid, blood pressure and cardiovascular risk.92, 93, 94, 95, 96 This situation can be explained by the hyperactivity of xanthine oxidase found in hypertension.21, 27
Exogenous antioxidants
Vitamin C. Vitamin C is a potent water-soluble antioxidant. In the vascular wall, it functions as an enzyme modulator that upregulates eNOS and downregulates NADPH oxidase.97 Most studies have demonstrated an inverse relationship between plasma ascorbate levels and blood pressure in both normotensive and hypertensive populations.15 Treatment with antioxidants has been shown to improve vascular function and reduce blood pressure in animal models98, 99 and in human hypertension.100, 101 Vitamin C may have favorable effects on vascular dilation, possibly through its antioxidant effects on NO.102, 103, 104
Nevertheless, several small and short-term clinical trials on the effect of vitamin C supplements on blood pressure have yielded inconsistent findings.105, 106, 107, 108, 109, 110, 111 The lack of antihypertensive efficacy observed in studies using supplementation with vitamin C alone could be due to the decreased bioavailability of NO under conditions of oxidative stress. It was shown that these effects are mediated in part by the ability of vitamin C to protect BH4 from oxidation and thereby increase the enzymatic activity of eNOS.112 In addition, this uncertain clinical beneficial effect of vitamin C as an antihypertensive agent could be due to the lack of consideration of its pharmacokinetic properties. It was experimentally determined that the antihypertensive effect of vitamin C is expected to occur at a concentration of 10 mM,103 which is an unattainable plasma level in humans through oral administration; that concentration is required for vitamin C to compete efficiently with the reaction of NO with superoxide. The renal ascorbic acid threshold occurs at daily vitamin C doses between 60 and 100 mg. The plasma is completely saturated at doses of 400 mg daily and higher, producing a steady-state plasma concentration of approximately 80 μM.113 Thus, the antihypertensive effect may only be active in plasma following high doses of vitamin C infusion.
Vitamin E. This major lipid-soluble antioxidant has received considerable attention for its antioxidant potential. Epidemiological data support an association between high dietary vitamin E intake and a reduced incidence of cardiovascular disease.56 Increasing evidence indicates that vitamin E can function as a biological modifier independently of its antioxidant activity. Available experimental evidence shows that vitamin E is capable of dose-dependently regulating mitochondrial generation of superoxide and hydrogen peroxide.
However, intervention trials have not been convincing, and several studies have demonstrated no beneficial effect of vitamin E on cardiovascular disease outcomes.114, 115, 116, 117 Moreover, a study using supplementation with vitamin E, either as α-tocopherol or mixed tocopherols, showed a significant increase in blood pressure, pulse pressure and heart rate in individuals with type 2 diabetes.118 It should be noted that sufficiently high concentrations in the vascular microenvironment that would enable vitamin E to interfere effectively with all components of oxidative stress is unlikely to be achieved.119
Association of Vitamins C and E. Ascorbic acid may reduce the α-tocopheroxyl radical and may be required for the beneficial vascular effects of α-tocopherol.120 In fact, the effect of α-tocopherol seems to be dependent on tissue saturation with vitamin C, and both vitamins may function synergistically to provide optimal conditions for endothelial NO formation.121 Thus, the association of vitamins C and E is expected to have an antihypertensive effect, probably because this combined therapy provides a reinforcement of their individual properties through a complementary effect.122
Despite the biological effects of both vitamin C and E, long-term clinical trials have failed to consistently support their antihypertensive effects in high-risk cardiovascular patients. Some short-term trials have shown that supplemental antioxidant vitamin intake lowers blood pressure,106, 123, 124, 125 but the majority of long-term clinical trials did not demonstrate any antihypertensive effects of antioxidant vitamins. However, most of these studies lacked rigorous exclusion criteria in the selection of subjects to avoid the influence of confounders.126 It should be noted that most of the subjects in cohorts of large trials had irreversible cardiovascular disease. A study performed with newly diagnosed hypertension, without end-organ damage, showed an association between oxidative-stress-related parameters and blood pressure thus, suggesting a role for oxidative stress in the pathogenesis of essential hypertension.127, 128 The available data suggest that there might be a beneficial antihypertensive effect of vitamins C and E if they are administered during the phase of endothelial dysfunction, which precedes established vascular damage. In contrast, an antihypertensive effect in patients with significant cardiovascular disease should not be expected because the chronic damaging effects of oxidative stress may be irreversible. Thus, it seems that ROS have a more important role in the induction rather than in the maintenance of hypertension.
Allopurinol. Xanthine oxidase is an important source of ROS in the vascular endothelium.24 It catalyzes the last two steps of purine metabolism and produces uric acid. Xanthine oxidase activity is associated with an increasing arteriolar tone and, therefore, hypertension.129, 130 Xanthine oxidase inhibitors, such as allopurinol, have shown marked improvements in endothelial function in various cohorts that were at risk for cardiovascular events. Treatment with allopurinol results in the reduction in blood pressure in adolescents,92 spontaneously hypertensive rats93 and patients with CKD.131 Nevertheless, most of the evidence gathered to date was derived from smaller mechanistic studies, and the few large randomized controlled trials have not shown a significant mortality benefit of using these agents.132
Selenium. Selenium is an essential trace element and an integral part of many proteins with catalytic and structural functions. It exerts an antioxidant function mainly in the form of selenocysteine residues, which are an integral constituent of ROS-detoxifying selenoenzymes, such as GSH-Px, thioredoxin reductases and selenoprotein P.133 Maintenance of full GSH-Px and thioredoxin reductase activity by adequate dietary intake of selenium has been proposed to be useful in the prevention of several cardiovascular disorders.112 In addition, selenium is capable of preventing the binding of nuclear factor kappa B to its nuclear response elements in DNA.134 Nuclear factor kappa B has a key role in inflammation and the production of ROS. The inhibition of nuclear factor kappa B is presumed to be the result of the binding of the selenium to the essential thiols of this transcription factor.135
The antioxidant properties of selenium have been documented in several trials.134, 136, 137, 138, 139, 140, 141 At low doses, selenium can provide significant protection of the human coronary artery endothelium against oxidative stress-mediated damage.133 In an animal model, dietary supplementation with selenium was associated with low levels of cardiac oxidative damage and increased antioxidant expression, as well as a reduction in disease severity and mortality in spontaneously hypertensive rats.142 A reduced selenium concentration in hypertensive pregnancies has been associated with a decrease in GSH-Px activity.143 Thus, it is reasonable to conclude that a deficiency in selenium might be an underestimated risk factor for the development of high blood pressure.144
N -acetylcysteine. The antioxidant N-acetylcysteine, which is a sulfhydryl group donor, improves renal dysfunction and decreases arterial pressure and renal injury in salt-sensitive hypertension.145 The inhibition of oxidative stress in hypertensive states probably contributes to the therapeutic effects of N-acetylcysteine, which are likely mediated via a NO-dependent mechanism.146 This protective mechanism is exerted by the prevention of BH4 oxidation by the increased superoxide.147 In addition, this molecule can protect against oxidative damage by inhibiting lipid peroxidation and scavenging ROS.148, 149
Polyphenols. Polyphenols are the most abundant antioxidants in the human diet. They can function as ROS scavengers, iron chelators and enzyme modulators150, 151 and may possibly enhance the production of NO.152, 153 In humans, circulating NO concentration increases after the consumption of polyphenols.154 However, some studies have shown increased blood pressure that is mediated by the association of polyphenols with vitamin C.155
Diet. There is sufficient evidence to suggest that dietary approaches may help to prevent and control high blood pressure. There are dietary approaches to the prevention and management of hypertension, that is, the moderate use of sodium and alcohol, as well as the increased intake of potassium, plant fibers, calcium and foods such as salmon, nuts and wine.156 In a Mediterranean population with an elevated fat consumption, a high fruit and vegetable intake is inversely associated with blood pressure levels.157 Short-term studies indicated that specialized diets may prevent or ameliorate mild hypertension; most notable are the Dietary Approaches to Stop Hypertension (DASH) diet, which is high in fruits, vegetables and low-fat dairy products.158 It has been reported that a low sodium DASH diet is effective in reducing blood pressure in hypertensive patients, particularly in those receiving antihypertensive medications.159 In addition, the DASH diet showed significant beneficial effects on cardiovascular risk.160, 161, 162 In overweight or obese persons with above-normal blood pressure, the addition of exercise and weight loss to the DASH diet resulted in even larger blood pressure reductions, greater improvements in vascular and autonomic function and reduced left ventricular mass.163, 164
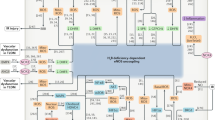
A summary of the exogenous antioxidant mechanisms that likely account for their anti-hypertensive effects is shown in Table 1 and Figure 2.

Schematic summary of the exogenous antioxidant mechanisms likely accounting for anti-hypertensive effects. NO, nitric oxide; eNOS, nitric oxide synthase; ROS, reactive oxygen species; BH4, tetrahydrobiopterin; oxBH4, oxidized tetrahydrobiopterin; NAC, N-acetylcysteine; VSMC, vascular smooth muscle cells.
Conclusions and perspectives
There is considerable evidence supporting the view that oxidative stress is involved in the pathophysiology of hypertension. ROS are mediators of the major physiological vasoconstrictors that increase intracellular calcium concentration. In addition, superoxide reduces the bioavailability of NO and enhances superoxide production via uncoupled eNOS; this further enhances oxidative stress, which is a major factor in hypertension.
Antioxidant therapy can curtail the development of hypertension in animal models, but remains controversial in humans. Possible confounding factors in patients include co-existing pathologies and treatments, lack of selection of treatments according to ROS levels, among others. However, the dietary intake of antioxidants and polyphenols could have an effect in the primary prevention or reduction of hypertension. Although the role of antioxidant therapy remains an open question, these interventions are expected to be more efficient in the prevention rather than in the reduction of established hypertension.
References
Yusuf S, Hawken S, Ounpuu S, Dans T, Avezum A, Lanas F, McQueen M, Budaj A, Pais P, Varigos J, Lisheng L . Effect of potentially modifiable risk factors associated with myocardial infarction in 52 countries (the INTERHEART Study): case control study. Lancet 2004; 364: 937–952.
Paravicini TM, Touyz RM . Redox signalling in hypertension. Cardiovasc Res 2006; 71: 247–258.
Rodrigo R, Passalacqua W, Araya J, Orellana M, Rivera G . Implications of oxidative stress and homocysteine in the pathophysiology of essential hypertension. J Cardiovasc Pharmacol 2003; 42: 453–461.
Lassègue B, Griendling K . Reactive oxygen species in hypertension, an update. AJH 2004; 17: 852–860.
Kimura S, Zhang GX, Nishiyama A, Shokoji T, Yao L, Fan YY, Rahman M, Abe Y . Mitochondria-derived reactive oxygen species and vascular MAP kinases: comparison of angiotensin II and diazoxide. Hypertension 2005; 45: 438–444.
Hool LC, Corry B . Redox control of calcium channels: from mechanisms to therapeutic opportunities. Antioxid Redox Signal 2007; 9: 409–435.
Yoshioka J, Schreiter ER, Lee RT . Role of thioredoxin in cell growth through interactions with signaling molecules. Antioxid Redox Signal 2006; 8: 2143–2145.
Lacy F, Kailasam MT, O’Connor DT, Schmid-Schonbein GW, Parmer RJ . Plasma hydrogen peroxide production in human essential hypertension: role of heredity, gender, and ethnicity. Hypertension 2000; 36: 878–884.
Stojiljkovic MP, Lopes HF, Zhang D, Morrow JD, Goodfriend TL, Egan BM . Increasing plasma fatty acids elevates F2-isoprostanes in humans: implications for the cardiovascular risk factor cluster. J Hypertens 2002; 201: 1215–1221.
Redon J, Oliva MR, Tormos C, Giner V, Chaves J, Iradi A, Saez GT . Antioxidant activities and oxidative stress byproducts in human hypertension. Hypertension 2003; 41: 1096–1101.
Tanito M, Nakamura H, Kwon YW, Teratani A, Masutani H, Shioji K, Kishimoto C, Ohira A, Horie R, Yodoi J . Enhanced oxidative stress and impaired thioredoxin expression in spontaneously hypertensive rats. Antioxid Redox Signal 2004; 6: 89–97.
Touyz RM . Reactive oxygen species, vascular oxidative stress, and redox signaling in hypertension: what is the clinical significance? Hypertension 2004; 44: 248–252.
Briones AM, Touyz RM . Oxidative stress and hypertension: current concepts. Curr Hypertens Rep 2010; 12: 135–142.
Bengtsson SH, Gulluyan LM, Dusting GJ, Drummond GR . Novel isoforms of NADPH oxidase in vascular physiology and pathophysiology. Clin Exp Pharmacol Physiol 2003; 30: 849–854.
Rodrigo R, Prat H, Passalacqua W, Araya J, Guichard C, Bächler JP . Relationship between oxidative stress and essential hypertension. Hypertens Res 2007; 30: 1159–1167.
Landmesser U, Dikalov S, Price SR, McCann L, Fukai T, Holland SM, Mitch WE, Harrison DG . Oxidation of tetrahydrobiopterin leads to uncoupling of endothelial cell nitric oxide synthase in hypertension. J Clin Invest 2003; 111: 1201–1209.
Gavazzi G, Banfi B, Deffert C, Fiette L, Schappi M, Herrmann F, Krause KH . Decreased blood pressure in NOX1-deficient mice. FEBS Lett 2006; 580: 497–504.
Touyz RM, Schiffrin EL . Increased generation of superoxide by angiotensin II in smooth muscle cells from resistance arteries of hypertensive patients: role of phospholipase D-dependent NAD(P)H oxidase-sensitive pathways. J Hypertens 2001; 19: 1245–1254.
Ghiadoni L, Magagna A, Versari D, Kardasz I, Huang Y, Taddei S, Salvetti A . Different effect of antihypertensive drugs on conduit artery endothelial function. Hypertension 2003; 41: 1281–1286.
Yoshida J, Yamamoto K, Mano T, Sakata Y, Nishikawa N, Nishio M, Ohtani T, Miwa T, Hori M, Masuyama T . AT1 receptor blocker added to ACE inhibitor provides benefits at advanced stage of hypertensive diastolic heart failure. Hypertension 2004; 43: 686–691.
Feairheller DL, Brown MD, Park JY, Brinkley TE, Basu S, Hagberg JM, Ferrell RE, Fenty-Stewart NM . Exercise training, NADPH oxidase p22phox gene polymorphisms, and hypertension. Med Sci Sports Exerc 2009; 41: 1421–1428.
Zou MH, Cohen RA, Ullrich V . Peroxynitrite and vascular endothelial dysfunction in diabetes mellitus. Endothelium 2004; 11: 89–97.
Lassègue B, Clempus RE . Vascular NAD(P)H oxidases: specific features, expression, and regulation. Am J Physiol Regul Integr Comp Physiol 2003; 285: 277–297.
Kuzkaya N, Weissmann N, Harrison DG, Dikalov S . Interactions of peroxynitrite, tetrahydrobiopterin, ascorbic acid, and thiols: implications for uncoupling endothelial nitric-oxide synthase. J Biol Chem 2003; 278: 22546–22554.
Laursen JB, Somers M, Kurz S, McCann L, Warnholtz A, Freeman BA, Tarpey M, Fukai T, Harrison DG . Endothelial regulation of vasomotion in apoE-deficient mice: implications for interactions between peroxynitrite and tetrahydrobiopterin. Circulation 2001; 103: 1282–1288.
Viel EC, Benkirane K, Javeshghani D, Touyz RM, Schiffrin EL . Xanthine oxidase and mitochondria contribute to vascular superoxide anion generation in DOCA-salt hypertensive rats. Am J Physiol Heart Circ Physiol 2008; 295: 281–288.
Laakso JT, Teräväinen TL, Martelin E, Vaskonen T, Lapatto R . Renal xanthine oxidoreductase activity during development of hypertension in spontaneously hypertensive rats. J Hypertens 2004; 22: 1333–1340.
Han D, Antunes F, Canali R, Rettori D, Cadenas E . Voltage-dependent anion channels control the release of the superoxide anion from mitochondria to cytosol. J Biol Chem 2003; 278: 5557–5563.
Eto Y, Kang D, Hasegawa E, Takeshige K, Minakami S . Succinate-dependent lipid peroxidation and its prevention by reduced ubiquinone in beef heart submitochondrial particles. Arch Biochem Biophys 1992; 295: 101–106.
Zhou L, Xiang W, Potts J, Floyd M, Sharan C, Yang H, Ross J, Nyanda AM, Guo Z . Reduction in extracellular superoxide dismutase activity in African-American patients with hypertension. Free Radic Biol Med 2006; 41: 1384–1391.
Michel JB, Feron O, Sase K, Prabhakar P, Michel T . Caveolin versus calmodulin. Counterbalancing allosteric modulators of endothelial nitric oxide synthase. J Biol Chem 1997; 272: 25907–25912.
Simko F, Luptak I, Matuskova J, Krajcirovicova K, Sumbalova Z, Kucharska J, Gvozdjakova A, Simko J, Babal P, Pechanova O, Bernatova I . L-arginine fails to protect against myocardial remodelling in L-NAME-induced hypertension. Eur J Clin Invest 2005; 35: 362–368.
Zhang Y, Hogg N . S-Nitrosothiols: cellular formation and transport. Free Radic Biol Med 2005; 38: 831–838.
Sládková M, Kojsová S, Jendeková L, Pechánová O . Chronic and acute effects of different antihypertensive drugs on femoral artery relaxation of L-NAME hypertensive rats. Physiol Res 2007; 56: 85–91.
Touyz RM . Reactive oxygen species and angiotensin II signaling in vascular cells - implications in cardiovascular disease. Braz J Med Biol Res 2004; 37: 1263–1273.
Hitomi H, Kiyomoto H, Nishiyama A . Angiotensin II and oxidative stress. Curr Opin Cardiol 2007; 22: 311–315.
Landmesser U, Cai H, Dikalov S, McCann L, Hwang J, Jo H, Holland SM, Harrison DG . Role of p47(phox) in vascular oxidative stress and hypertension caused by angiotensin II. Hypertension 2002; 40: 511–515.
Taniyama Y, Griendling K . Reactive oxygen species in the vasculature: molecular and cellular mechanisms. Hypertension 2003; 42: 1075–1081.
Pechánová O . Contribution of captopril thiol group to the prevention of spontaneous hypertension. Physiol Res 2007; 56: 41–48.
Bitar MS, Wahid S, Mustafa S, Al-Saleh E, Dhaunsi GS, Al-Mulla F . Nitric oxide dynamics and endothelial dysfunction in type II model of genetic diabetes. Eur J Pharmacol 2005; 511: 53–64.
Gomez-Alamillo C, Juncos LA, Cases A, Haas JA, Romero JC . Interactions between vasoconstrictors and vasodilators in regulating hemodynamics of distinct vascular beds. Hypertension 2003; 42: 831–836.
Djordjevic T, BelAiba RS, Bonello S, Pfeilschifter J, Hess J, Görlach A . Human urotensin II is a novel activator of NADPH oxidase in human pulmonary artery smooth muscle cells. Arterioscler Thromb Vasc Biol 2005; 25: 519–525.
Matsushita M, Shichiri M, Imai T, Iwashina M, Tanaka H, Takasu N, Hirata Y . Co-expression of urotensin II and its receptor (GPR14) in human cardiovascular and renal tissues. J Hypertens 2001; 19: 2185–2190.
Jegou S, Cartier D, Dubessy C, Gonzalez BJ, Chatenet D, Tostivint H, Scalbert E, LePrince J, Vaudry H, Lihrmann I . Localization of the urotensin II receptor in the rat central nervous system. J Comp Neurol 2006; 495: 21–36.
Stirrat A, Gallagher M, Douglas SA, Ohlstein EH, Berry C, Kirk A, Richardson M, MacLean MR . Potent vasodilator responses to human urotensin-II in human pulmonary and abdominal resistance arteries. Am J Physiol Heart Circ Physiol 2001; 280: 925–928.
Rodrigo R, Passalacqua W, Araya J, Orellana M, Rivera G . Homocysteine and essential hypertension. J Clin Pharmacol 2003; 43: 1299–1306.
Harrison DG, Gongora MC . Oxidative stress and hypertension. Med Clin North Am 2009; 93: 621–635.
Rodrigo R, Rivera G . Renal damage mediated by oxidative stress: a hypothesis of protective effects of red wine. Free Radic Biol Med 2002; 33: 409–422.
Zhang C, Hu JJ, Xia M, Boini KM, C Brimson, PL Li . Redox signaling via lipid raft clustering in homocysteine-induced injury of podocytes. Biochimica et Biophysica Acta 2010; 1803: 482–491.
Piccoli C, Quarato G, D’Aprile A, Montemurno E, Scrima R, Ripoli M, Gomaraschi M, Cirillo P, Boffoli D, Calabresi L, Gesualdo L, Capitanio N . Native LDL-induced oxidative stress in human proximal 1 tubular cells: multiple players involved. J Cell Mol Med 2009 (in press).
Klahr S . Urinary tract obstruction. Semin Nephrol 2001; 21: 133–145.
Grande MT, Pérez-Barriocanal F, López-Novoa JM . Role of inflammation in túbulo-interstitial damage associated to obstructive nephropathy. J Inflamm (Lond) 2010; 22: 7–19.
Sachse A, Wolf G . Angiotensin II-induced reactive oxygen species and the kidney. J Am Soc Nephrol 2007; 18: 2439–2446.
Chung S, Park CW, Shin SJ, Lim JH, Chung HW, Youn DY, Kim HW, Kim BS, Lee JH, Kim GH, Chang YS . Tempol or candesartan prevents high-fat diet-induced hypertension and renal damage in spontaneously hypertensive rats. Nephrol Dial Transplant 2010; 25: 389–399.
Guarnieri G, Zanetti M, Vinci P, Cattin MR, Pirulli A, Barazzoni R . Metabolic syndrome and chronic kidney disease. J Ren Nutr 2010; 20: 19–23.
Malyszko J . Mechanism of endothelial dysfunction in chronic kidney disease. Clin Chim Acta 2010; 411: 1412–1420.
Costa-Hong V, Bortolotto LA, Jorgetti V, Consolim-Colombo F, Krieger EM, Lima JJ . Oxidative stress and endothelial dysfunction in chronic kidney disease. Arq Bras Cardiol 2009; 92: 381–386.
Zoccali C, Bode-Böger S, Mallamaci F, Benedetto F, Tripepi G, Malatino L, Cataliotti A, Bellanuova I, Fermo I, Frölich J, Böger R . Plasma concentration of asymmetrical dimethylarginine and mortality in patients with end-stage renal disease: aprospective study. Lancet 2001; 358: 2113–2117.
Nanayakkara PW, Teerlink T, Stehouwer CD, Allajar D, Spijkerman A, Schalkwijk C, ter Wee PM, van Guldener C . Plasma asymmetric dimethylarginine (ADMA) concentration is independently associated with carotid intima-media thickness and plasma soluble vascular cell adhesión molecule-1 (sVCAM-1) concentration in patients with mild-to-moderate renal failure. Kidney Int 2005; 68: 2230–2236.
Campos RR, Oliveira-Sales EB, Nish EM, Boim MA, Dolnikoff MS, Bergamaschi CT . The role of oxidative stress in renovascular hypertension Special Series: Stress and Hypertension. Clin Exp Pharmacol Physiol 2010 (in press).
Grassi G . Assessment of sympathetic cardiovascular drive in human hypertension: achievements and perspectives. Hypertension 2009; 54: 690–697.
Guyenet PG . The sympathetic control of blood pressure. Nat Rev Neurosci 2006; 7: 335–346.
Kishi T, Hirooka Y, Kimura Y, Ito K, Shimokawa H, Takeshita A . Increased reactive oxygen species in rostral ventrolateral medulla contribute to neural mechanisms of hypertension in stroke-prone spontaneously hypertensive rats. Circulation 2004; 109: 2357–2362.
Hirooka Y, Sagara Y, Kishi T, Sunagawa K . Oxidative stress and central cardiovascular regulation. Pathogenesis of hypertension and therapeutic aspects. Circ J 2010; 74: 827–835.
Sved AF, Ito S, Sved JC . Brainstem mechanisms of hypertension: role of the rostral ventrolateral medulla. Curr Hypertens Rep 2003; 5: 262–268.
Oliveira-Sales EB, Nishi EE, Carillo BA, Boim MA, Dolnikoff MS, Bergamaschi CT, Campos RR . Oxidative stress in the sympathetic promotor neurons contributes to sympathetic activation in renovascular hypertension. Am J Hypertens 2009; 22: 484–492.
Zimmerman MC, Lazartigues E, Sharma RV, Davisson RL . Hypertension caused by angiotensin II infusion involves increased superoxide production in the central nervous system. Circ Res 2004; 95: 210–216.
Andreoli SP . Reactive oxygen molecules, oxidant injury and renal disease. Pediatr Nephrol 1991; 5: 733–742.
Meister A, Anderson ME . Glutathione. Annu Rev Biochem 1983; 52: 711–760.
Rubanyi GM, Vanhoutte PM . Superoxide anions and hyperoxia inactivate endothelium-derived relaxing factor. Am J Physiol 1986; 250: 822–827.
Lee SK, Arunkumar S, Sirajudeen KN, Singh HJ . Glutathione system in young spontaneously hypertensive rats. J Physiol Biochem 2010; 66: 321–327.
de Cavanagh EM, Ferder LF, Ferder MD, Stella IY, Toblli JE, Inserra F . Vascular structure and oxidative stress in salt-loaded spontaneously hypertensive rats: effects of losartan and atenolol. Am J Hypertens 2010; 23: 1318–1325.
Zhou L, Xiang W, Potts J, Floyd M, Sharan C, Yang H, Ross J, Nyanda AM, Guo Z . Reduction in extracellular superoxide dismutase activity in African-American patients with hypertension. Free Radic Biol Med 2006; 41: 1384–1391.
Kashyap MK, Yadav V, Sherawat BS, Jain S, Kumari S, Khullar M, Sharma PC, Nath R . Different antioxidants status, total antioxidant power and free radicals in essential hypertension. Mol Cell Biochem 2005; 277: 89–99.
Kedziora-Kornatowska K, Czuczejko J, Pawluk H, Kornatowski T, Motyl J, Szadujkis-Szadurski L, Szewczyk-Golec K, Kedziora J . The markers of oxidative stress and activity of the antioxidant system in the blood of elderly patients with essential arterial hypertension. Cell Mol Biol Lett 2004; 9: 635–641.
Zhan CD, Sindhu RK, Pang J, Ehdaie A, Vaziri ND . Superoxide dismutase, catalase and glutathione peroxidase in the spontaneously hypertensive rat kidney: effect of antioxidant-rich diet. J Hypertens 2004; 22: 2025–2033.
Wernerman J, Hammarqvist F . Modulation of endogenous glutathione availability. Curr Opin Clin Nutr Metab Care 1999; 2: 487–492.
Da Silva AP, Marinho C, Gonçalves MC, Monteiro C, Laires MJ, Falcão LM, Nogueira JB, Bich M . Decreased erythrocyte activity of methemoglobin and glutathione reductases may explain age-related high blood pressure. Rev Port Cardiol 2010; 29: 403–412.
Joles JA, Wesseling S, Braam B . Renal glutathione S-transferase mu type 1 expression is already reduced in new-born spontaneously hypertensive rats. J Hypertens 2010; 28: 633–634.
Ebrahimian T, Touyz RM . Thioredoxin in vascular biology: role in hypertension. Antioxid Redox Signal 2008; 10: 1127–1136.
Tanito M, Nakamura H, Kwon YW, Teratani A, Masutani H, Shioji K, Kishimoto C, Ohira A, Horie R, Yodoi J . Enhanced oxidative stress and impaired thioredoxin expression in spontaneously hypertensive rats. Antioxid Redox Signal 2004; 6: 89–97.
Xu S, He Y, Vokurkova M, Touyz RM . Endothelial cells negatively modulate reactive oxygen species generation in vascular smooth muscle cells: role of thioredoxin. Hypertension 2009; 54: 427–433.
Widder JD, Fraccarollo D, Galuppo P, Hansen JM, Jones DP, Ertl G, Bauersachs J . Attenuation of angiotensin II-induced vascular dysfunction and hypertension by overexpression of Thioredoxin 2. Hypertension 2009; 54: 338–344.
Maulik N, Das DK . Emerging potential of thioredoxin and thioredoxin interacting proteins in various disease conditions. Biochim Biophys Acta 2008; 1780: 1368–1382.
World CJ, Yamawaki H, Berk BC . Thioredoxin in the cardiovascular system. J Mol Med 2006; 84: 997–1003.
Vítek L, Ostrow JD . Bilirubin chemistry and metabolism; harmful and protective aspects. Curr Pharm Des 2009; 15: 2869–2883.
Vera T, Stec DE . Moderate hyperbilirubinemia improves renal hemodynamics in angiotensin ii dependent hypertension. Am J Physiol Regul Integr Comp Physiol 2010; 299: R1044–R10499.
LeBlanc RM, Navar LG, Botros FT . Bilirubin exerts renoprotective effects in angiotensin II-hypertension. Am J Med Sci 2010; 340: 144–146.
Chin HJ, Song YR, Kim HS, Park M, Yoon HJ, Na KY, Kim Y, Chae DW, Kim S . The bilirubin level is negatively correlated with the incidence of hypertension in normotensive Korean population. J Korean Med Sci 2009; 24: 50–56.
Waring WS, Webb DJ, Maxwell SR . Systemic uric acid administration increases serum antioxidant capacity in healthy volunteers. J Cardiovasc Pharmacol 2001; 38: 365–371.
Waring WS, Convery A, Mishra V, Shenkin A, Webb DJ, Maxwell SR . Uric acid reduces exercise-induced oxidative stress in healthy adults. Clin Sci (Lond) 2003; 105: 425–430.
Feig DI, Soletsky B, Johnson RJ . Effect of allopurinol on blood pressure of adolescents with newly diagnosed essential hypertension: a randomized trial. JAMA 2008; 300: 924–932.
Mazzali M, Hughes J, Kim Y, Jefferson JA, Kang DK, Gordon KL, Lan HY, Kivlighn S, Johnson RJ . Elevated uric acid increases blood pressure in the rat by a novel crystal-independent mechanism. Hypertension 2001; 38: 1101–1106.
Mazzali M, Kanbay M, Segal MS, Shafiu M, Jalal D, Feig DI, Johnson RJ . Uric acid and hypertension: cause or effect?. Curr Rheumatol Rep 2010; 12: 108–117.
Baker JF, Schumacher HR . Update on gout and hyperuricemia. Int J Clin Pract 2010; 64: 371–377.
Nagahama K, Inoue T, Iseki K, Touma T, Kinjo K, Ohya Y, Takishita S . Hyperuricemia as a predictor of hypertension in a screened cohort in Okinawa, Japan. Hypertens Res 2004; 27: 835–841.
Ulker S, McKeown PP, Bayraktutan U . Vitamins reverse endothelial dysfunction through regulation of eNOS and NAD(P)H oxidase activities. Hypertension 2003; 41: 534–539.
Nishikawa Y, Tatsumi K, Matsuura T, Yamamoto A, Nadamoto T, Urabe K . Effects of vitamin C on high blood pressure induced by salt in spontaneously hypertensive rats. J Nutr Sci Vitaminol (Tokyo) 2003; 49: 301–309.
Reckelhoff JF, Kanji V, Racusen LC, Schmidt AM, Yan SD, Marrow J, Roberts LJ, Salahudeen AK . Vitamin E ameliorates enhanced renal lipid peroxidation and accumulation of F2-isoprostanes in aging kidneys. Am J Physiol 1998; 274: 767–774.
Chen X, Touyz RM, Park JB, Schiffrin EL . Antioxidant effects of vitamins C and E are associated with altered activation of vascular NADPH oxidase and superoxide dismutase in stroke-prone SHR. Hypertension 2001; 38: 606–611.
Atarashi K, Ishiyama A, Takagi M, Minami M, Kimura K, Goto A, Omata M . Vitamin E ameliorates the renal injury of Dahl Salt-sensitive rats. Am J Hypertens 1997; 10: 116–119.
Vita JA, Frei B, Holbrook M, Gokce N, Leaf C, Keaney Jr JF . L-2-Oxothiazolidine-4-carboxylic acid reverses endothelial dysfunction in patients with coronary artery disease. J Clin Invest 1998; 101: 1408–1414.
Jackson TS, Xu A, Vita JA, Keaney Jr JF . Ascorbate prevents the interaction of superoxide and nitric oxide only at very high physiological concentrations. Circ Res 1998; 83: 916–922.
Duffy SJ, Gokce N, Holbrook M, Hunter LM, Biegelsen ES, Huang A, Keaney Jr JF, Vita JA . Effect of ascorbic acid treatment on conduit vessel endothelial dysfunction in patients with hypertension. Am J Physiol Heart Circ Physiol 2001; 280: 528–534.
Duffy SJ, Gokce N, Holbrook M, Huang A, Frei B, Keaney Jr JF, Vita JA . Treatment of hypertension with ascorbic acid. Lancet 1999; 354: 2048–2049.
Fotherby MD, Williams JC, Forster LA, Craner P, Ferns GA . Effect of vitamin C on ambulatory blood pressure and plasma lipids in older persons. J Hypertens 2000; 18: 411–415.
Block G, Mangels AR, Norkus EP, Patterson BH, Levander OA, Taylor PR . Ascorbic acid status and subsequent diastolic and systolic blood pressure. Hypertension 2001; 37: 261–267.
Ghosh SK, Ekpo EB, Shah IU, Girling AJ, Jenkins C, Sinclair AJ . A double-blind, placebo-controlled parallel trial of vitamin C treatment in elderly patients with hypertension. Gerontology 1994; 40: 268–272.
Duffy SJ, Gokee N, Holbrook M, Huang A, Frei B, Keany JFJ . Treatment of hypertension with ascorbic acid. Lancet 2004; 354: 2048–2049.
Galley HF, Thornton J, Howdle PD, Walber BE, Webster NR . Combination oral antioxidant supplementation reduces blood pressure. Clin Sci 1997; 92: 361–365.
Mullan BA, Young IS, Fee H, McCance DR . Ascorbic acid reduces blood pressure and arterial stiffness in type 2 diabetes. Hypertension 2002; 40: 797–803.
Steinbrenner H, Sies H . Protection against reactive oxygen species by selenoproteins. Biochim Biophys Acta 2009; 1790: 1478–1485.
Padayatty SJ, Katz A, Wang Y, Eck P, Kwon O, Lee JH, Chen S, Corpe C, Dutta A, Dutta SK, Levine M . Vitamin C as an antioxidant: evaluation of its role in disease prevention. J Am Coll Nutr 2003; 22: 18–35.
Rapola JM, Virtamo J, Ripatti S, Huttunen JK, Albanes D, Taylor PR, Heinonen OP . Randomised trial of a-tocopherol and b-carotene supplements on incidence of major coronary events in men with previous myocardial infarction. Lancet 1997; 349: 1715–1720.
GISSI-Prevenzione Investigators. Dietary supplementation with n-3 polyunsaturated fatty acids and vitamin E after myocardial infarction: results of the GISSI-Prevenzione Trial. Lancet 1999; 354: 447–455.
Lonn E, Bosch J, Yusuf S, Sheridan P, Pogue J, Arnold JM, Ross C, Arnold A, Sleight P, Probstfield J, Dagenais GR, HOPE and HOPE-TOO Trial Investigators. Effects of long-term vitamin E supplementation on cardiovascular events and cancer: a randomized controlled trial. JAMA 2005; 293: 1338–1347.
Lee IM, Cook NR, Gaziano JM, Gordon D, Ridker PM, Manson JE, Hennekens CH, Buring JE . Vitamin E in the primary prevention of cardiovascular disease and cancer: the Women's Health Study: a randomized controlled trial. JAMA 2005; 294: 56–65.
Ward NC, Wu JH, Clarke MW, Puddey IB, Burke V, Croft KD, Hodgson JM . The effect of vitamin E on blood pressure in individuals with type 2 diabetes: a randomized, double-blind, placebo-controlled trial. J Hypertens 2007; 25: 227–234.
Münzel T, Keaney Jr JF . Are ACE inhibitors a ‘magic bullet’ against oxidative stress? Circulation 2001; 104: 1571–1574.
Heller R, Werner-Felmayer G, Werner ER . Antioxidants and endothelial nitric oxide synthesis. Eur J Clin Pharmacol 2006; 62: 21–28.
Heller R, Werner-Felmayer G, Werner ER . Alpha-tocopherol and endothelial nitric oxide synthesis. Ann N Y Acad Sci 2004; 1031: 74–85.
Bilodeau JF, Hubel CA . Current concepts in the use of antioxidants for the treatment of preeclampsia. J Obstet Gynaecol Can 2003; 25: 742–750.
Mullan B, Young IS, Fee H, McCance DR . Ascorbic acid reduces blood pressure and arterial stiffness in type 2 diabetes. Hypertension 2002; 40: 804–809.
Galley HF, Thornton J, Howdle PD, Walker BE, Webster NR . Combination oral antioxidant supplementation reduces blood pressure. Clin Sci 1997; 92: 361–365.
Plantinga Y, Ghiadoni L, Magagna A, Giannarelli C, Franzoni F, Taddei S, Salvetti A . Suplementation with vitamins C and E improves arterial stiffness and endothelial function in essential hypertensive patients. Am J Hypertens 2007; 20: 392–397.
Rodrigo R, Guichard C, Charles R . Clinical pharmacology and therapeutic use of antioxidant vitamins. Fundam Clin Pharmacol 2007; 21: 111–127.
Rodrigo R, Prat H, Passalacqua W, Araya J, Bächler JP . Decrease in oxidative stress through supplementation of vitamins C and E is associated with a reduction in blood pressure in patients with essential hypertension. Clin Sci (Lond) 2008; 114: 625–634.
Rodrigo R, Vinay J, Castillo R, Cereceda M, Asenjo R, Zamorano J, Araya J, Castillo-Koch R, Espinoza J, Larraín E . Use of vitamins C and E as a prophylactic therapy to prevent postoperative atrial fibrillation. Int J Cardiol 2010; 138: 221–228.
Suzuki H, DeLano FA, Parks DA, Jamshidi N, Granger DN, Ishii H, Suematsu M, Zweifach BW, Schmid-Schonbein GW . Xanthine oxidase activity associated with arterial blood pressure in spontaneously hypertensive rats. Proc Natl Acad Sci USA 1998; 95: 4754–4759.
DeLano FA, Parks DA, Ruedi JM, Babior BM, Schmid-Schönbein GW . Microvascular display of xanthine oxidase and NADPH oxidase in the spontaneously hypertensive rat. Microcirculation 2006; 13: 551–566.
Goicoechea M, Vinuesa SG, Verdalles U, Ruiz-Caro C, Ampuero J, Rincón A, Arroyo D, Lu ño J . Effect of allopurinol in chronic kidney disease progression and cardiovascular risk. Clin J Am Soc Nephrol 2010; 5: 1388–1393.
George J, Struthers A . The role of urate and xanthine oxidase in vascular oxidative stress: future directions. Ther Clin Risk Manag 2009; 5: 799–803.
Miller S, Walker SW, Arthur JR, Nicol F, Pickard K, Lewin MH, Howie AF, Beckett GJ . Selenite protects human endothelial cells from oxidative damage and induces thioredoxin reductase. Clin Sci (Lond) 2001; 100: 543–550.
Faure P, Ramon O, Favier A, Halimi S . Selenium supplementation decreases nuclear factor-kappa B activity in peripheral blood mononuclear cells from type 2 diabetic patients. Eur J Clin Invest 2004; 34: 475–481.
Kim IY, Stadtman TC . Inhibition of NF-kappaB DNA binding and nitric oxide induction in human T cells and lung adenocarcinoma cells by selenite treatment. Proc Natl Acad Sci 1997; 94: 12904–12907.
Campbell L, Howie F, Arthur JR, Nicol F, Beckett G . Selenium and sulforaphane modify the expression of selenoenzymes in the human endothelial cell line EAhy926 and protect cells from oxidative damage. Nutrition 2007; 23: 138–144.
Takizawa M, Komori K, Tampo Y, Yonaha M . Paraquat-induced oxidative stress and dysfunction of cellular redox systems including antioxidative defense enzymes glutathione peroxidase and thioredoxin reductase. Toxicol In Vitro 2007; 21: 355–363.
Faure P . Protective effects of antioxidant micronutrients (vitamin E, zinc and selenium) in type 2 diabetes mellitus. Clin Chem Lab Med 2003; 41: 995–998.
Brigelius-Flohé R, Banning A, Schnurr K . Selenium-dependent enzymes in endothelial cell function. Antioxid Redox Signal 2003; 5: 205–215.
Ito Y, Fujita T . Trace elements and blood pressure regulation. Nippon Rinsho 1996; 54: 106–110.
Zhou X, Ji WJ, Zhu Y, He B, Li H, Huang TG, Li YM . Enhancement of endogenous defenses against ROS by supra-nutritional level of selenium is more safe and effective than antioxidant supplementation in reducing hypertensive target organ damage. Med Hypotheses 2007; 68: 952–956.
Lymbury RS, Marino MJ, Perkins AV . Effect of dietary selenium on the progression of heart failure in the ageing spontaneousl hypertensive rat. Mol Nutr Food Res 2010; 54: 1436–1444.
Mistry HD, Wilson V, Ramsay MM, Symonds ME, Broughton Pipkin F . Reduced selenium concentrations and glutathione peroxidase activity in preeclamptic pregnancies. Hypertension 2008; 52: 881–888.
Nawrot TS, Staessen JA, Roels HA, Den Hond E, Thijs L, Fagard RH, Dominiczak AF, Struijker-Boudier HA . Blood pressure and blood selenium: a cross-sectional and longitudinal population study. Eur Heart J 2007; 28: 628–633.
Tian N, Rose RA, Jordan S, Dwyer TM, Hughson MD, Manning Jr RD . N-Acetylcysteine improves renal dysfunction, ameliorates kidney damage and decreases blood pressure in salt-sensitive hypertension. J Hypertens 2006; 24: 2263–2270.
Pechánová O, Zicha J, Kojsová S, Dobesová Z, Jendeková L, Kunes J . Effect of chronic N-acetylcysteine treatment on the development of spontaneous hypertension. Clin Sci (Lond) 2006; 110: 235–242.
Zembowicz A, Hatchett RJ, Radziszewski W, Gryglewski RJ . Inhibition of endothelial nitric oxide synthase by ebselen. Prevention by thiols suggests the inactivation by ebselen of a critical thiol essential for the catalytic activity of nitric oxide synthase. J Pharmacol Exp Ther 1993; 267: 1112–1118.
De la Fuente M, Victor VM . Ascorbic acid and N-acetylcysteine improve in vitro the function of lymphocytes from mice with endotoxin-induced oxidative stress. Free Radic Res 2001; 35: 73–84.
Penugonda S, Mare S, Goldstein G, Banks WA, Ercal N . Effects of N-acetylcysteine amide (NACA), a novel thiol antioxidant against glutamate-induced cytotoxicity in neuronal cell line PC12. Brain Res 2005; 1056: 132–138.
Rodrigo R, Bosco C . Oxidative stress and protective effects of polyphenols: comparative studies in human and rodent kidney. A review. Comp Biochem Physiol C Toxicol Pharmacol 2006; 142: 317–327.
Pietta P, Simonetti P, Gardana C, Brusamolino A, Morazzoni P, Bombardelli E . Relationship between rate and extent of catechin absorption and plasma antioxidant status. Biochem Mol Biol Int 1998; 46: 895–903.
Duarte J, Andriambeloson E, Diebolt M, Andriantsitohaina R . Wine polyphenols stimulate superoxide anion production to promote calcium signaling and endothelialdependent vasodilatation. Physiol Res 2004; 53: 595–602.
Zenebe W, Pechánová O, Andriantsitohaina R . Red wine polyphenols induce vasorelaxation by increased nitric oxide bioactivity. Physiol Res 2003; 52: 425–432.
Pechánová O, Rezzani R, Babál P, Bernátová I, Andriantsitohaina R . Beneficial effects of provinols: cardiovascular system and kidney. Physiol Res 2006; 55: 17–30.
Ward NC, Hodgson JM, Croft KD, Burke V, Beilin LJ, Puddey IB . The combination of vitamin C and grape-seed polyphenols increases blood pressure: a randomized, double-blind, placebo-controlled trial. J Hypertens 2005; 23: 427–434.
Appel LJ, Moore TJ, Obarzanek E, Vollmer WM, Svetkey LP, Sacks FM, Bray GA, Vogt TM, Cutler JA, Windhauser MM, Lin PH, Karanja N . A clinical trial of the effects of dietary patterns on blood pressure. DASH Collaborative Research Group. N Engl J Med 1997; 336: 1117–1124.
Alonso A, de la Fuente C, Martín-Arnau AM, de Irala J, Martínez JA, Martínez-González MA . Fruit and vegetable consumption is inversely associated with blood pressure in a Mediterranean population with a high vegetable-fat intake: the Seguimiento Universidad de Navarra (SUN) Study. Br J Nutr 2004; 92: 311–319.
Savica V, Bellinghieri G, Kopple JD . The effect of nutrition on blood pressure. Annu Rev Nutr 2010; 30: 365–401.
Nowson CA, Wattanapenpaiboon N, Pachett A . Low-sodium dietary approaches to stop hypertension-type diet including lean red meat lowers blood pressure in postmenopausal women. Nutr Res 2009; 29: 8–18.
Azadbakht L, Fard NR, Karimi M, Baghaei MH, Surkan PJ, Rahimi M, Esmaillzadeh A, Willett WC . Effects of the dietary approaches to stop hypertension (DASH) eating plan on cardiovascular risks among type 2 diabetic patients: a randomized cross-over clinical trial. Diabetes Care 2010 (in press).
Chen ST, Maruthur NM, Appel LJ . The effect of dietary patterns on estimated coronary heart disease risk: results from the Dietary Approaches to Stop Hypertension (DASH) trial. Circ Cardiovasc Qual Outcomes 2010; 3: 484–489.
Levitan EB, Wolk A, Mittleman MA . Relation of consistency with the dietary approaches to stop hypertension diet and incidence of heart failure in men aged 45–79 years. Am J Cardiol 2009; 104: 1416–1420.
Smith PJ, Blumenthal JA, Babyak MA, Craighead L, Welsh-Bohmer KA, Browndyke JN, Strauman TA, Sherwood A . Effects of the dietary approaches to stop hypertension diet, exercise, and caloric restriction on neurocognition in overweight adults with high blood pressure. Hypertension 2010; 55: 1331–1338.
Blumenthal JA, Babyak MA, Hinderliter A, Watkins LL, Craighead L, Lin PH, Caccia C, Johnson J, Waugh R, Sherwood A . Effects of the DASH diet alone and in combination with exercise and weight loss on blood pressure and cardiovascular biomarkers in men and women with high blood pressure: the ENCORE study. Arch Intern Med 2010; 170: 126–135.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Rodrigo, R., González, J. & Paoletto, F. The role of oxidative stress in the pathophysiology of hypertension. Hypertens Res 34, 431–440 (2011). https://doi.org/10.1038/hr.2010.264
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/hr.2010.264
Keywords
This article is cited by
-
Onion quercetin inhibits vascular endothelial cell dysfunction and prevents hypertension
European Food Research and Technology (2024)
-
Arterial stiffness and hypertension
Clinical Hypertension (2023)
-
Association between raised blood pressure and elevated serum liver enzymes among active-duty Royal Thai Army personnel in Thailand
BMC Cardiovascular Disorders (2023)
-
Antihypertensive, antioxidant, and renal protective impact of integrated GJD with captopril in spontaneously hypertensive rats
Scientific Reports (2023)
-
Association between the accumulation of elevated serum γ-glutamyltransferase level and risk of atrial fibrillation: a nationwide cohort study
Scientific Reports (2023)
