
Abstract
Background:
Steroid sulphatase (STS) is one of the steroid-metabolising enzymes involved in desulphating inactive steroid sulphates and oestrogen sulphotransferase (EST) sulphates active oestrogen. The roles of both STS and EST have not been examined in oestrogen-dependent non-small-cell lung cancer (NSCLC).
Methods:
We evaluated the immunoreactivity of STS and EST in NSCLC cases using immunohistochemistry. The function of STS and EST was further demonstrated using NSCLC cell lines.
Results:
The immunoreactivity of STS and EST was detected in 49.5% and 27.8% of NSCLC cases, respectively. The immunoreactivity of STS was significantly higher in female adenocarcinoma cases. The STS-positive NSCLCs were also significantly correlated in an inversed manner with tumour size and cell proliferation and tended to be associated with better clinical outcome. However, the immunoreactivity of EST was significantly correlated with intracellular oestradiol concentration. Results of in vitro analysis demonstrated that oestrone sulphate (E1-S) induced and pregnenolone sulphate (Preg-S) inhibited the proliferation in STS-expressing cell lines. The inhibition by Preg-S was reversed by a specific progesterone receptor blocker. Simultaneous addition of E1-S and Preg-S significantly suppressed the proliferation.
Conclusion:
In NSCLC patients, STS is considered a good prognostic factor. Results of our present study also indicated the benefits of potential progesterone therapy for NSCLC patients.
Similar content being viewed by others


Main
Lung cancer is one of the most common types of malignancies in the world and its frequency has consistently increased in both men and women over the years in many parts of the world (Shibuya et al, 2002; Mathers and Loncar, 2006; Pharm et al, 2007). Lung cancer is histopathologically classified into small-cell and non-small-cell lung carcinoma (NSCLC) and the latter is further subclassified into the following three categories; adenocarcinoma, squamous cell carcinoma, and large cell carcinoma. Non-small-cell lung carcinoma accounts for ∼80% of all primary lung carcinomas and at least some of them have been recently proposed as one of oestrogen-dependent human malignancies. Results of previous studies demonstrated that female lifetime nonsmokers tended to develop NSCLC than male counterparts and some endocrine therapies for postmenopausal NSCLC patients resulted in survival benefits, although short-term improvement (Zang and Wynder, 1996). In addition, an increasing number of the reported studies demonstrated NSCLC as one of oestrogen-dependent tumours as in breast or endometrial malignancies (Schabath et al, 2004; Ganti et al, 2006; Niikawa et al, 2008). In postmenopausal breast cancer patients, intratumoural production of oestrogens occurs as a result of aromatisation of androgens into oestrogens, which is catalysed by the cytochrome P450 aromatase enzyme (Labrie, 1991; Sasano et al, 2008). Aromatase inhibitors such as anastrozole, letrozole, and exemestane have all been established as the first-line endocrine therapy for oestrogen receptor (ER)-positive postmenopausal breast carcinoma patients (Nabholtz et al, 2000; Mouridsen et al, 2001; Paridaens et al, 2008). In NSCLC tissues, Mah et al (2007) reported the presence of aromatase using immunohistochemistry and aromatase-positive older female patients had a greater survival than those who were aromatase negative. In addition, both ER blockers and aromatase inhibitors were reported to decrease the cell proliferation in NSCLC cells in both in vitro and in vivo studies (Kawai et al, 2005; Stabile et al, 2005; Weinberg et al, 2005; Dougherty et al, 2006; Koutras et al, 2009). Based upon these findings, some of the clinical trials employing oestrogen receptor blockers or aromatase inhibitors as endocrine therapeutic agents have currently been investigated in the patients with NSCLC. For instance, phase II clinical trials of combination therapy of epidermal growth factor receptor inhibitor erlotinib (Tarceva) with ER blocker fluvestrant (Faslodex) versus erlotinib alone in advanced NSCLC patients have been conducted (ClinicalTrials.gov Identifier: NCT00100854 and NCT00592007). In addition, a phase II randomised trial of fluvestrant and aromatase inhibitor anastrozole (Arimidex) as consolidation therapy in NSCLC patients who have received first-line platinum-based chemotherapy with or without monoclonal antibody for vascular endothelial growth factor Bevacizumab (Avastin) has been recently carried out (ClinicalTrials.gov Identifier: NCT00932152).
Steroid sulphatase (STS) has been recently focussed on as a novel therapeutic target of antioestrogen therapy for ER-positive breast cancer patients (Stanway et al, 2007; Foster et al, 2008; Geisler et al, 2011). The STS desulphates oestrogen (both oestradiol and oestrone) sulphates abundantly present in the circulating blood. Some investigators did demonstrate that oestrogens present in breast carcinoma cells were produced more frequently by STS pathway than by aromatase pathway (Tilson-Mallett et al, 1983; James et al, 1987). In addition, STS inhibitors have actually been demonstrated to be effective for breast cancer cell line MCF-7 (Selcer et al, 1997). It is also important, however, to note an involvement of oestrogen sulphotransferase (EST; SULT1E1), the other oestrogen-metabolising enzyme that biologically inactivates oestrogens by sulphating them in the activity of STS of particular tissues (Schabath et al, 2004). Both STS and EST were reported to be present in 74.3% and 51.4% of breast cancer patients, respectively (Suzuki et al, 2003). In the breast cancer microenvironment, both STS and EST have been considered to play important roles in the regulation of intratumoural oestrogen production. STS inhibitors have been developed as a novel endocrine therapy agent for ER-positive postmenopausal breast cancer patients based upon the results of these in vitro and in vivo studies above (Foster et al, 2008; Geisler et al, 2011). It is also known that STS catalyses the sulphonation of DHEA, cholesterol, and pregnenolone as well as oestrone. In addition, STS is also known to have eight alternatively spliced variants of its gene with corresponding tissue-specific transcripts (Dalla Valle et al, 2007; Nardi et al, 2009). However, to the best of our knowledge, the status of STS and EST expression has not been examined in NSCLC tissues, and thus the biological significances or characteristics of intratumoural STS and EST have remained unclear in lung cancer.
Therefore, in this study, we first examined the status of STS and EST in NSCLC tissues at both mRNA and protein levels using quantitative RT–PCR and immunohistochemistry and then evaluated the correlation between immunoreactivity and the clinicopathological factors of individual cases. In addition, oestrogen concentration in NSCLC tissues was also measured in order to study whether these intratumoural enzymes are functional or not. We further explored the mechanisms of regulation of STS gene expression in vitro in STS-transfected NSCLC cell line and endogenous STS-expressing cell line. Finally, we analysed the presence of splicing variants of STS mRNA, which may be involved in STS function in order to further study STS enzymatic activities in NSCLC.
Materials and methods
NSCLC tissues
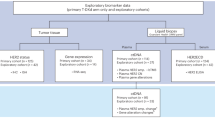
A total of 97 NSCLC cases were retrieved from the surgical pathology files at the Department of Pathology, Tohoku University Hospital (Sendai, Japan) and Ishinomaki Red Cross Hospital (Ishinomaki, Japan), respectively. These cases examined had all been fixed in 10% formalin and embedded in paraffin. The histologic types of these paraffin-embedded NSCLC cases were as follows: adenocarcinoma, 82 cases; squamous cell carcinoma, 15 cases. Among these cases, 59 cases from Tohoku University Hospital that had also been stored as frozen specimens were used for mRNA and tissue concentration studies because all corresponding frozen tissues in Ishinomaki Red Cross Hospital had been defrosted and denatured because of the blackout following an earthquake on 11 March 2011and subsequent tsunami. The histological types of these cases were as follows: adenocarcinoma, 44 cases; squamous cell carcinoma, 15 cases. Research protocols for this study were approved by the Ethics Committee at Tohoku University School of Medicine (#2008-444) and Ishinomaki Red Cross Hospital (#2008.6.30).
Immunohistochemistry
Immunohistochemistry was conducted by streptavidin-biotin method using a Histofine kit (Nichirei Co., Ltd, Tokyo, Japan). The lists of primary antibodies used in this study and their dilution for immunohistochemistry were as follows: anti-mouse monoclonal STS antibody (kindly provided by Kyowa Medex Co., Ltd, Tokyo, Japan), 0.37 mg ml−1; anti-rabbit polyclonal EST antibody (Medical & Biological Laboratories Co., Ltd, Nagoya, Japan), 1 : 1500; anti-mouse monoclonal aromatase antibody (contributed by Dr Evans DB, Novartis, Basel, Switzerland), 1 : 3000; and anti-mouse monoclonal Ki67 antibody (DAKO Cytomation, Carpinteria, CA, USA), 1 : 100. The slides were treated with a microwave (500 W for 15 min) and an autoclave (121 °C for 5 min) in citrate buffer (pH 6.0), respectively, for antigen retrieval for ER and Ki67 immunostaining. No treatment for antigen retrieval was performed in the staining of STS and aromatase. Immunoreactivities for STS, EST, and aromatase were detected in the cytoplasm of carcinoma cells, and cases that had >10% positive cells were considered as positive according to the results of previous published study (Suzuki et al, 2003; Abe et al, 2010). The Ki67 immunoreactivity was evaluated by counting more than 1000 carcinoma cells in hot spots and the percentage of immunoreactivity (labelling index (LI)) was subsequently obtained. Results of immunohistochemistry were independently evaluated by two of the authors (HK and KA).
Cell culture and reagents
The cell lines used in this study were as follows: human male lung adenocarcinoma, A549 and NCI-H23, which were obtained from American Type Culture Collection (ATCC, Manassas, VA, USA), LK87 (Cell Resource Center for Biomedical Research, Tohoku University, Sendai, Japan) and PC9 (Immuno-Biological Laboratories Co. (IBL), Ltd, Gunma, Japan); human female lung adenocarcinoma, NCI-H1975 (ATCC) and PC3 (Cell Resource Center for Biomedical Research), which is different from PC-3 prostate cancer cell line (ATCC, CRL-1435; Yamashita et al, 1996; Imai et al, 2004; Ishibashi et al, 2005b); human squamous cell lung carcinoma, LK2 (Cell Resource Center for Biomedical Research) and RERF-LC-AI (RIKEN Bio Resource Center, Tsukuba, Japan); and human breast carcinoma, T47D (ATCC). The cell lines obtained from ATCC and IBL were all obtained in 2010–2012 and authenticated by them. The other cell lines were previously used as reported by our group (Miki et al, 2010). All the cell lines examined were cultured in RPMI-1640 medium (Sigma-Aldrich Co., St Louis, MO, USA) containing 10% fetal bovine serum (FBS; Nichirei Co., Ltd) and maintained at 37 °C, 95% relative humidity, and 5% CO2. Mifepristone and pregnenolone sulphate were obtained from Sigma-Aldrich Co. Oestrone sulphate and ICI 182780 were purchased from Wako Pure Chemical Industries Ltd (Osaka, Japan).
Stable transfection
The stable transfection of a vector containing STS cDNA was performed using Lipofectamine LTX (Invitrogen, Gaithersburg, MD, USA) according to the manufacturer’s protocol in order to generate the cell lines with stable expression of STS from LK87 cells. The following vectors were kindly provided by ASKA Pharma Medical Co., Ltd (Kawasaki, Japan): pIRESneo2/hSTS, containing STS gene sequences; pIRESneo2, control vector. When LK87 cells were incubated in an ∼60% confluence and the mixture of the vectors and Lipofectamine LTX were added into LK87 cells. Following incubation for 24 h, the corresponding clones were selected in the medium containing G418 (Sigma-Aldrich Co.) for 2 weeks. The LK87 cells transfected with STS gene vector (indicated as LK87-STS) or control vector (LK87 Ctrl) were generated through these processes above.
Real-time quantitative RT–PCR
Total RNA was extracted from frozen tissues of 59 cases and NSCLC cell lines using Trizol reagent (Invitrogen). Frozen tissues were cut into pieces and homogenised with PT-2100 Polytron homogeniser (Kinematica, Inc, Luzem, Switzerland). Reverse-transcriptional reaction was performed using Quantitect Reverse Transcription kit (QIAGEN GmbH, Hilden, Germany) in order to synthesise cDNA from total RNA. Quantitative RT–PCR was carried out using the LightCycler System (Roche Diagnostics GmbH, Mannheim, Germany) and the FastStart DNA Master SYBR Green I with software version 3.5.3 (Roche Diagnostics GmbH).
Analysis of splicing variants of STS mRNA was evaluated using quantitative RT–PCR (Hot Star Taq Plus PCR; QIAGEN GmbH) and electrophoresis. Five types of splicing variants (i.e., 0a, 0b, 0c, 1a, and 1c) were analysed in this study. Tissues of placenta and thyroid and a breast cancer cell line MCF-7 were used as a positive control. The primers were synthesised by Nihon Gene Research Laboratories Inc. (Sendai, Japan). As an internal control, RPL13A mRNA was used. The expression rate of these genes was calculated and evaluated as the percentage compared with that of the internal control. The sequences of the primers used in this study are summarised in Supplementary Tables 1 and 2.
Liquid chromatography/electrospray tandem mass spectrometry
Tissue concentrations of oestrogen (oestrone and oestradiol) in 59 NSCLC tissues and progesterone (progesterone and pregnenolone) in 10 cases were measured using liquid chromatography-mass spectrometry/mass spectrometry (LC-MS/MS) analysis (ASKA Pharma Medical Co., Ltd). These concentrations were adjusted for tissue weight.
Regarding NSCLC cell lines, cells were cultivated in charcoal medium for 3 days and seeded into six-well plate at a density of 200 000 cells per well. Following 24 h of incubation, the cells were treated with 300 pg ml−1 of oestrone sulphate for 24 h (n=1) or 10 μ M of pregnenolone sulphate for 72 h (n=3) and the concentrations of the desulphated and synthesised products from them (oestrone from oestrone sulphate; pregnenolone and progesterone from pregnenolone sulphate) in the medium were evaluated by LC-MS/MS. The number of the cells was measured using cell counter (Sysmex CDA-500, Sysmex Corporation, Kobe, Japan) and the concentrations were adjusted for the cell numbers.
Proliferation assay
Proliferation assay was performed under both anchorage-dependent and -independent conditions. Cells were cultivated in charcoal medium for 3 days and seeded into 96-well plates under both normal adhesion condition (Asahi Glass Co., Inc, Tokyo, Japan) and low adhesion condition (EZ-BindShut Microplate; Asahi Glass Co., Inc.) at a density of 2000 (LK87-STS) and 5000 (NCI-H1975) cells per well, respectively. The cells were treated with steroids, its sulphates, or inhibitors for 24, 48, and 72 h in an anchorage-dependent condition or 1, 3, 5, and 7 days in an anchorage-independent condition and both assays were evaluated using WST-8 Cell Counting Kit-8 (Dojindo Laboratories, Kumamoto, Japan). The status of the cell number was evaluated as follows: (the average absorbance of treated cells/the average absorbance of control cells) × 100 (%).
Immunoblotting analysis
Total protein was extracted from the cell lines using Mammalian Protein Extraction Reagent (Pierce Biotechnology, Rockford, IL, USA) containing Halt Protease Inhibitor Cocktail (Pierce Biotechnology). After measuring the concentrations of the protein (Protein Assay Kit Wako: Wako Pure Chemical Industries, Ltd), total protein was individually subjected to SDS–PAGE (SuperSep Ace: Wako Pure Chemical Industries, Ltd). Proteins were then transferred onto Hybond P polyvinylidene difluoride membrane (GE Healthcare, Buckinghamshire, UK). The blots were then blocked in 5% non-fat dry skim milk powder (Wako Pure Chemical Industries Ltd) for 1 h at room temperature, and were then incubated with the primary antibodies overnight at 4 °C using Can Get Signal R Solution (Toyobo Biochemicals, Osaka, Japan). The dilutions of primary antibodies used in this study were summarised as follows: STS, 1 : 5000; EST, 1 : 500; ERα (Novocastra, Newcastle, UK), 1 : 500; ERβ (GeneTex, San Antonio, TX, USA), 1 : 200; PR (Chemicon, Temecula, CA, USA), 1 : 1000; β-actin (Sigma-Aldrich Co.), 1 : 1000. These antibody–protein complexes on the blots were detected using ECL-plus Western blotting detection reagents (GE Healthcare) following incubation with anti-mouse IgG horseradish peroxidase (GE Healthcare) for 60 min at room temperature. The corresponding protein bands were subsequently visualised and analysed with LAS-1000 cooled CCD-camera chemiluminescent image analyser and Multi Gauge v3.1 software (both from Fuji Photo Film Co. Ltd, Tokyo, Japan), respectively.
Statistical analysis
Statistical analyses were evaluated using Student’s t-test, Wilcoxon signed-rank test, Mann–Whitney U-test, χ2 test, and one-way ANOVA. Overall survival curve was generated according to the Kaplan–Meier method and statistical significance was calculated using the log-rank test. All statistical analyses were carried out using StatView 5.0 J software (SAS Institute Inc., Cary, NC, USA). Statistical significance was defined as P<0.05 in this study.
Results
Expression of STS and EST at mRNA and protein levels in NSCLC tissues
The immunoreactivity of STS and EST was detected in the cytoplasm of carcinoma cells in 48 (49.5%; Figure 1A) and 27 (27.8%; Figure 1B) of 97 NSCLC cases examined, respectively. The immunoreactivity of STS was not detected in morphologically normal lung tissues including bronchial epithelial cells and alveolar cells (data not shown), but EST immunoreactivity was weakly detected in bronchial epithelial cells (data not shown).

(A and B) Representative illustrations of immunohistochemistry of STS (A) and EST (B) in NSCLC tissues. Original magnification × 100. (C–F) Summary of overall survival analysis in 59 NSCLC cases or 44 lung adenocarcinoma cases according to the status of intratumoural STS (C and D) and EST (E and F). Positive cases are indicated by solid lines and negative cases by dashed lines. Survival curves were obtained using Kaplan–Meier method and P-values were evaluated using log-rank test.
The STS mRNA was widely detected in both tumour and nontumour sites examined in this study (Supplementary Figure 1A). However, EST mRNA was detected in tumour sites in only 6 out of 59 NSCLC cases examined (Supplementary Figure 1B). Both STS and EST mRNA expression levels demonstrated no statistical significant differences between those in tumour and normal sites (data not shown). Its immunoreactivity and mRNA expression levels were significantly correlated in EST (P=0.020), but not in STS (P=0.69; Supplementary Figure 2).
The immunoreactivity of STS was significantly more frequently detected in female and adenocarcinoma cases (P=0.0034 and 0.0009, respectively), and was inversely associated with tumour size and Ki67 LI of carcinoma cells (P=0.013 and 0.016, respectively; Table 1). The immunoreactivity of STS and EST tended to be coexpressed in NSCLC cases examined (P=0.060; Table 1). The status of EST immunoreactivity also tended to be associated with the cell proliferation of tumour cells (P=0.083), but no significant association was detected with any other clinicopathological parameters of the cases examined (Table 2). Aromatase expression was not correlated with both STS (P=0.99; Table 1) and EST (P=0.52; Table 2).
The status of STS immunoreactivity tended to be associated with better clinical outcome in 59 NSCLC cases (P=0.46, Figure 1C), especially in 44 lung adenocarcinoma cases (P=0.051, Figure 1D), but this tendency did not reach statistical significance. The EST-positive cases also tended to be associated with poorer overall survival (P=0.18, 59 cases; P=0.17, 44 lung adenocarcinoma cases; Figure 1E and F).
Correlation between the status of STS and EST and intratumoural oestrogen concentrations
In 59 NSCLC cases, the correlation between STS-immunopositive cases and intratumoural oestrogen (oestrone and oestradiol) concentration was not statistically significant (P=0.70 and 0.46, respectively; Figure 2A and C). However, intratumoural oestradiol but not oestrone concentration was significantly higher in EST-immunopositive cases (P=0.014 and 0.23, respectively; Figure 2B and D).

Correlation between the results of immunohistochemistry and oestrogen concentrations.Oestrone and estradiol concentrations in 59 NSCLC tissues were measured by LC-MS/MS and evaluated according to the status of intratumoural STS mRNA (oestrone, A; oestradiol, C) and EST mRNA expression (oestrone, B; oestradiol, D). Statistical significances were analysed by Mann–Whitney U-test and defined as P<0.05.
Expression and function of STS and EST in NSCLC cell lines
Steroid sulphatase was detected in all NSCLC cell lines examined in this study at mRNA levels (Figure 3A). In most of the cell lines examined, EST mRNA was expressed at markedly low levels, but was detected in LK87, PC3, and NCI-H1975 (Figure 3B). The PR mRNA expression in LK87 and NCI-H1975 was higher than other NSCLC cell lines examined in this study and not detected in PC3 (Figure 3C). Both STS and EST were detected in the above three cell lines at both mRNA and protein levels (Figure 3A, B). Among these cell lines examined, LK87 and NCI-H1975 expressed both ERβ and PR but not PC3, as demonstrated at mRNA levels (Figure 3D), although at relatively low levels compared with those of breast cancer cell line T47D. The ERα protein was not detected in these NSCLC cell lines (Figure 3D). Relatively high level of STS expression at protein levels was detected in LK87-STS cells compared with LK87 Ctrl (Figure 3E).

The status of STS, EST, and hormone receptors in NSCLC cell lines. Summary of the basal expression of STS (A), EST (B), and PR (C) at mRNA levels in carcinoma cell lines derived from male adenocarcinoma (black box), female adenocarcinoma (grey box), and squamous cell lung carcinoma (white box). (D) Summary of the basal levels of STS, EST, ERα, ERβ, and PR at protein levels in LK87, PC3, and NCI-H1975. Each protein band was quantified compared with β-actin as loading control and relative quantification of each band is indicated. The breast carcinoma cell line T47D was used as a positive control for ERα, ERβ, and PR. (E) Summary of STS expression at protein levels in LK87 transfected with a control vector (LK87 Ctrl) and the vector containing STS gene (LK87-STS).
We measured the oestrone concentrations in culture medium following an administration of oestrone sulphates in LK87 Ctrl, LK87-STS, PC3, and NCI-H1975, and its concentrations were evaluated in all the cell lines examined (Supplementary Figure 3). The highest activity of STS was in LK87-STS among these four cell lines examined, with LK87 Ctrl higher than NCI-H1975 (Supplementary Figure 3).
Effects of the sulphated sex steroids on NSCLC cell lines
We measured the concentrations of both pregnenolone and progesterone in cultured medium following the addition of the sulphated form of pregnenolone in LK87 Ctrl and LK87-STS. Concentrations of both these steroids were significantly increased in LK87-STS compared with LK87 Ctrl (Figure 4A and B). Results of proliferation assay following an addition of oestrone sulphate in LK87-STS indicated that oestrone sulphates significantly induced the cell proliferation in a concentration-dependent manner (Figure 4C). Pregnenolone sulphates significantly inhibited the cell proliferation and mifepristone reversed the inhibition (Figure 4D). When the sulphated forms of both oestrone and pregnenolone were simultaneously administered at the final concentration of 1, 10, and 100 nM, the significant decrement of the cell number was noted in all these conditions in LK87-STS cells (Figure 4E).

Characteristics of LK87-STS cells. (A and B) Pregnenolone and progesterone concentrations in LK87 Ctrl and LK87-STS cells treated with pregnenolone sulphate. The concentrations of pregnenolone and progesterone contained in cultured medium following administration of 10 μ M pregnenolone sulphate for 72 h were measured using LC-MS/MS (n=3) in order to evaluate STS activity. Results indicated that the concentrations of both pregnenolone (A) and progesterone (B) were significantly higher in LK87-STS cells than in LK87 Ctrl. (C–E) Summary of proliferation assay using LK87-STS cells. The effects of (C) oestrone sulphates (0, 10 pM, 100 pM, and 1 nM) at day 3 (n=6) and of (D) pregnenolone sulphates (Preg-S, 10 μ M) and mifepristone (1.0 μ M) at day 3 (n=6) of cell proliferation. (E) The effects of simultaneous administration of oestrone sulphate (E1-S; 1, 10, and 100 nM) and pregnenolone sulphate (Preg-S; 1, 10, and 100 nM) at day 5 in an anchorage-independent condition is shown (n=6). P-values were evaluated using Student’s t-test. *P<0.05, **P<0.01, ***P<0.001.
In both PC3 and NCI-H1975 cells, concentrations of pregnenolone and progesterone in culture medium following an addition of pregnenolone sulphates were detected at a relatively low level compared with LK87-STS (Figure 5A and B). However, the cell proliferation in NCI-H1975 was increased by an addition of oestrone sulphate and also decreased by that of pregnenolone sulphate (Figure 5C and D). In addition, the inhibition by pregnenolone sulphate above was reversed by mifepristone (Figure 5D). Simultaneous addition of the sulphated forms of oestrone and pregnenolone significantly inhibited the growth of NCI-H1975 in the same condition as in LK87-STS (Figure 5E).

Characteristics of NCI-H1975 cells. (A and B) Pregnenolone and progesterone concentrations in PC3 and NCI-H1975 cells treated with pregnenolone sulphate (n=3). The concentrations of pregnenolone and progesterone contained in cultured medium following administration of 10 μ M pregnenolone sulphate for 72 h was measured using LC-MS/MS in order to evaluate STS activity in these cells. The concentrations of both pregnenolone (A) and progesterone (B) were detected in these two cell lines examined. (C–E) Summary of proliferation assay using NCI-H1975 cells. The effects of (C) oestrone sulphates (0, 100 pM, 1 nM, and 10 nM) at day 3 (n=6) and of (D) pregnenolone sulphates (Preg-S, 10 μ M) and mifepristone (1.0 μ M) at day 3 (n=6) of cell proliferation. (E) The effects of simultaneous administration of oestrone sulphate (E1-S; 1, 10, and 100 nM) and pregnenolone sulphate (Preg-S; 1, 10, and 100 nM) at day 5 in an anchorage-independent condition is demonstrated. P-values were evaluated using Student’s t-test. *P<0.05, **P<0.01, ***P<0.001.
Progesterone concentrations in lung adenocarcinoma tissues
We further analysed the concentration of progesterone and evaluated the correlation of tissue progesterone concentration with STS/EST expression in 10 lung adenocarcinoma cases. The higher concentration of tissue progesterone was detected in STS-positive cases and lower in EST-positive patients, but the correlation did not reach statistical significances (P=0.73 and 0.14, respectively; Supplementary Figure 4). In addition, PR was expressed in only 1 out of the 10 cases examined and STS mRNA was not detected in this particular case.
Analysis of splicing variants of STS in NSCLC
We analysed the status of the splicing variants of STS mRNA (i.e., exons 0a, 0b, 0c, 1a, and 1b) in both NSCLC tissues and cell lines. Exons 0a, 0b, and 1c were detected in both tumour and adjacent nonpathological lung tissues of NSCLC tissues, but exons 0c and 1a of STS mRNA were detected only in tumour tissues (Supplementary Table 3 and Supplementary Figure 5). Exon 1a STS mRNA was detected in 2 of the 10 NSCLC tissues but not in the 2 NSCLC cell lines.
Discussion
In breast cancer tissues, STS immunoreactivity was correlated with tumour size, and was significantly associated with an increased risk of recurrence and an adverse clinical outcome (Suzuki et al, 2003). However, STS-positive NSCLC patients had a smaller tumour size and lower tumour cell proliferation as evaluated by Ki-67 index. In addition, a better overall survival of the patients tended to be associated with the presence of intratumoural STS expression in NSCLC, especially in lung adenocarcinoma patients. Steroid sulphatase is known as a poor prognostic marker for breast cancer patients and its targeted agents have been currently developed (Stanway et al, 2007; Foster et al, 2008; Geisler et al, 2011). Steroid sulphatase in breast cancer desulphates oestrone sulphate and produces biologically active oestrogen, oestradiol, resulting in higher cell proliferation by binding to ERα and eventually in adverse clinical outcome (Suzuki et al, 2003). Intratumoural expression of STS has also been reported in several types of tumours such as endometrial carcinoma (Utsunomiya et al, 2004), thymoma (Ishibashi et al, 2005a), and colon carcinoma (Sato et al, 2009). All these above reports indicated that intratumoural oestrogen, oestradiol, and/or oestrone are elevated through STS enzymatic activity and are related to tumour progression. In NSCLC tissues, providing that intratumoural oestrogen was increased by STS as demonstrated in other tumours above, it is reasonably postulated that intratumoural STS could be positively correlated with tumour cell proliferation status. However, in our present study, an inverse correlation was detected between STS immunoreactivity and the cell proliferation markers. In addition, the status of intratumoural STS immunoreactivity was not significantly correlated with intratumoural oestrone and oestradiol concentrations in NSCLC tissues examined. Therefore, based upon the above findings, we hypothesised that the possible biological roles of intratumoural STS in NSCLC, which were clearly different from those in other malignancies, may be due to the intratumoural status of EST, which has the opposite enzymatic activity to STS, or the other steroids desulphated by STS, and proceeded with further analysis in order to explore this hypothesis.
As demonstrated in human breast cancer patients, EST also plays pivotal roles in intratumoural oestrogen metabolism/production (Suzuki et al, 2003). In our present study, EST immunoreactivity was detected in approximately one-third of all NSCLC cases examined and tended to be correlated with higher cellular proliferation of tumour cells. In addition, oestradiol concentration was significantly higher in EST-positive cases examined. Oestrogen sulphotransferase in breast carcinoma cells was reported to sulphate oestrogens, which subsequently results in decrement of biologically active oestrogens in tumour tissue microenvironment, resulting in lower proliferation of tumour cells (Suzuki et al, 2003). However, the correlations between the status of intratumoural EST and oestradiol concentrations in NSCLC cells were rather opposite to those in breast cancer tissues in our study. We previously demonstrated the significant positive correlation between aromatase mRNA expression and intratumoural oestradiol concentration in NSCLC (Niikawa et al, 2008), suggesting that aromatase may increase oestradiol concentrations and higher proliferation of tumour cells in NSCLC carcinoma cells, which is more important than intratumoural EST in this type of cancer. In addition, it is also true that only 8 out of 59 cases examined were immunohistochemically positive for EST, and that EST mRNA could not be detected in the great majority of the cases. It awaits further investigations for clarification of the significance of intratumoural EST in human NSCLC.
Steroid sulphatase catalyses the sulphonation of DHEA, cholesterol, and pregnenolone as well as oestrone (Figure 6). In human leukocyte, an affinity of STS for pregnenolone sulphate was relatively higher than the sulphated forms of DHEA and oestrone (Hirato et al, 1991). Pregnenolone is well known to be synthesised from cholesterol by P450scc and StAR, and to be subsequently converted into DHEA by CYP17 or progesterone by 3β-hydroxysteroid dehydrogenase (3β-HSD). We previously reported that a significantly high concentration of progesterone was detected in NSCLC cases expressing progesterone-producing enzymes such as StAR, P450scc, and 3β-HSD, and the treatment of cholesterol increased progesterone concentration in NSCLC cell lines (Ishibashi et al, 2005b). In addition, PR-positive NSCLC cases were significantly associated with a better overall survival and progesterone did inhibit cell proliferation in PR-expressing NSCLC cell lines (Ishibashi et al, 2005b). Therefore, we hypothesised that progesterone may be synthesised from pregnenolone sulphates by STS and 3β-HSD in NSCLC tissues and result in an overall better clinical outcome of the patients.

Simple diagram of the metabolism of oestrogen and progesterone. ER, oestrogen receptor; EST, oestrogen sulphotransferase; HSD, hydroxysteroid dehydrogenase; PR, progesterone receptor; STS, steroid sulphatase.
In our present study, we first evaluated STS and EST expression at both mRNA and protein levels in all the cell lines evaluated in order to select appropriate NSCLC cell lines. The STS expression was widely detected in NSCLC cell lines examined in this study but EST was below the level of detection limits. In addition, the expression level of STS was higher in PC3 and NCI-H1975 derived from female adenocarcinoma than those from male adenocarcinoma and squamous cell carcinoma. The cell lines LK87, PC3, and NCI-H1975 all expressed both STS and EST, but the amounts of ERα, ERβ, and PR expression in PC3 were much smaller than in the other cell lines. Therefore, based upon these results, we selected LK87-STS to evaluate the activity of stably expressing STS by vector transfection, and NCI-H1975 to analyse the function of endogenous STS. The STS activity of parental LK87 or the cell without transfection was higher than that of NCI-H1975 when oestrone concentration was measured after addition of its sulphate (Supplementary Figure 3).
We then demonstrated that both progesterone and pregnenolone concentrations were detected when pregnenolone sulphates were administered to both LK87-STS and NCI-H1975, suggesting that pregnenolone sulphates were desulphated by STS and then immediately converted into progesterone by 3β-HSD, which was also demonstrated to be associated with intratumoural progesterone concentration in NSCLC tissues in our previous study (Ishibashi et al, 2005b). Therefore, progesterone converted from pregnenolone sulphates through STS and 3β-HSD was considered to promote the antiproliferative actions mediated by PR in both cells, and this suppressive effects was indeed recovered by an administration of mifepristone, a PR blocker, in our present study. The serum level of oestrone sulphate is usually approximately the level of 3.8 nM (premenopausal women at early follicular phase), 6.3 nM (premenopausal women at late follicular phase), 1.1 nM (postmenopausal women), and 2.1 nM (adult men) (Adly et al, 2006; Greenlee et al, 2007). However, the serum concentrations of pregnenolone sulphate are usually the same between male and female subjects. Its concentration reaches a peak at ∼30 years of age and subsequently decreases throughout the ages up to ∼100 nM after 60 years (Tagawa et al, 2000; Havlíková et al, 2002). Therefore, we then added the sulphated forms of oestrone and pregnenolone simultaneously administered at ∼10 nM as their standard concentrations in order to evaluate which sulphated forms play more important roles in NSCLC. Oestrone sulphates did increase the cell proliferation but significant reduction of the cell number was detected even in the condition containing a higher concentration of oestrone sulphates than that of pregnenolone sulphates. The progesterone treatment also decreased STS expression at mRNA levels in LK87 and NCI-H1975 but oestradiol did not (data not shown), suggesting that the sensitivity to progesterone may be higher than that to oestrogen in NSCLC cells. This may be also caused by an oestrogen-induced PR expression, because PR gene is well known to have oestrogen-response element and to be controlled by oestrogen (Thi et al, 1975; Nardulli et al, 1988; Wei et al, 1988). Oestrone sulphate cannot bind to ER at all and therefore fails to stimulate any oestrogen-induced biological responses. Oestrone sulphates were reported to be desulphated and converted to oestradiol (Vignon et al, 1980; MacIndoe et al, 1988) that subsequently induced PR expression at mRNA levels in human breast carcinoma cell line MCF-7 (Pasqualini et al, 1986). Therefore, the proliferation may be induced by oestrogen but progesterone further inhibited the growth in NSCLC through oestrogen-induced PR in carcinoma cells, which may account for a better clinical outcome in STS-positive NSCLC cases examined in our study, but it awaits further investigations, such as studying the larger number of the patients for PR and STS expression, for clarification.
The presence of a variety of splicing variants of STS mRNA, which have the transcription start points in untranslated region, has been evaluated mostly in breast carcinoma cells (Dalla Valle et al, 2007; Nardi et al, 2009). Eight types of splicing variants of STS mRNA (i.e., 0a–c and 1a–e) have so far been reported in the literature and their expression patterns have also been reported to influence STS activity and expression (Dalla Valle et al, 2007; Nardi et al, 2009). Among these splicing variants of STS mRNA, exon 1d was expressed in a rather restricted number of tissues and at low expression levels, but exon 1b was expressed throughout the body (Dalla Valle et al, 2007). Therefore, in our present study, we evaluated five exons, 0a–c, 1a, and 1c, in NSCLC tissues and cell lines. We demonstrated that 0c and 1a STS mRNA were detected only in tumour but not in nonpathological areas of NSCLC tissues examined. Results of our present study also revealed that all five patterns of STS exons were detected in lung cancer tissues examined. Three splicing variants of STS mRNA, 0c, 1a, and 1c, were below the detection levels in nontumour tissues of lung cancer tissues in our present study. Nardi et al (2009) demonstrated the expression of 1a STS mRNA in one case of lung cancer, and its corresponding nontumour tissues and higher expression level of exon 1a was detected in tumour tissues. These results as well as those of our present study suggested that the switching of STS promoter usage may be related to malignant transformation in human lung. In addition, exon 1a may also play an important role in STS enzymatic activity in lung cancer as reported in placenta and breast cancer (Dalla Valle et al, 2007; Nardi et al, 2009).
The results of epidemiological studies demonstrated that the combined treatment of oestrogen and progesterone increased both the incidence and mortality of female postmenopausal lung cancer patients (Chlebowski et al, 2009). On the other hand, the above effects were not detected in the groups of postmenopausal women taking only oestrogen (Chlebowski et al, 2010). However, it is also important to note that in both reported studies, the status of ERβ and PR in the lung carcinoma cells was not studied. Therefore, it is reasonably postulated that the presence of these receptors could have influenced the different clinical outcome of the patients, especially those receiving both oestrogen and progesterone. In addition, the microenvironment of cancer including the carcinoma–stromal interactions is considered to play pivotal roles in the progression of human malignancies, but the potential roles of progesterone in tumour microenvironment have not been necessarily well characterised. Therefore, an involvement of both oestrogen and progesterone receptors may be required to stimulate the lung cancer growth, but it awaits further investigations for clarification.
There have been more controversies as to the actions of progesterone compared with those of oestrogen in lung cancer. For instance, Check et al (2010) reported that PR blocker mifepristone demonstrated antitumour activities in mouse models of lung cancer. However, in our present study, mifepristone restored the inhibition of cell proliferation as a result of pregnenolone sulphate administration. In addition, Stabile et al (2011) also reported that human lung cancer patients with relatively high ERβ/low PR status were associated with poorer prognosis, which is not necessarily consistent with the results of our present study. One of the reasons for this discrepancy is the possible influence of tumour microenvironment such as stromal cells and other extracellular factors on the results because the results of the above-mentioned studies had all been obtained from these in vivo models. We previously demonstrated that cytokines derived from the stromal cells of human lung cancer tissues induced aromatase expression in lung carcinoma cells (Miki et al, 2010). Therefore, it is reasonably postulated that cytokines or other factors derived from carcinoma–stromal interactions may influence the overall balance and/or effects of oestrogen and/or progesterone in human lung cancer. Therefore, further studies are required to clarify the roles of STS and EST with relation to microenvironment of human lung cancer.
In breast cancer, STS is a poorer prognostic factor and its targeted drugs have been currently developed as a novel antioestrogen therapy (Geisler et al, 2011). Many studies have indicated that lung cancer is an oestrogen-dependent cancer as well as breast cancer (Schabath et al, 2004; Ganti et al, 2006; Niikawa et al, 2008). We also demonstrated that intratumoural progesterone was considered to be increased through the intratumoural STS activity and contributes to suppression of cancer growth. However, studies regarding the possible roles of nuclear receptor signalling in the lung and its diseases have not been well studied compared with those in breast cancer. Therefore, further studies including those of intratumoural progesterone signalling may provide new insights into the potential roles of STS inhibitors.
Change history
16 April 2013
This paper was modified 12 months after initial publication to switch to Creative Commons licence terms, as noted at publication
References
Abe K, Miki Y, Ono K, Mori M, Kakinuma H, Kou Y, Kudo N, Koguchi M, Niikawa H, Suzuki S, Evans DB, Sugawara S, Suzuki T, Sasano H (2010) Highly concordant coexpression of aromatase and estrogen receptor beta in non-small cell lung cancer. Hum Pathol 41: 190–198
Adly L, Hill D, Sherman ME, Sturgeon SR, Fears T, Mies C, Ziegler RG, Hoover RN, Schairer C (2006) Serum concentrations of estrogens, sex hormone-binding globulin, and androgens and risk of breast cancer in postmenopausal women. Int J Cancer 119: 2402–2407
Check JH, Dix E, Wilson C, Check D (2010) Progesterone receptor antagonist therapy has therapeutic potential even in cancer restricted to males as evidenced from murine testicular and prostate cancer studies. Anticancer Res 30: 4921–4923
Chlebowski RT, Schwartz AG, Wakelee H, Anderson GL, Stefanick ML, Manson JE, Rodabough RJ, Chien JW, Wactawski-Wende J, Gass M, Kotchen JM, Johnson KC, O’Sullivan MJ, Ockene JK, Chen C, Hubbell FA Women’s Health Initiative Investigators (2009) Estrogen plus progestin and lung cancer in postmenopausal women (Women’s Health Initiative trial): a post-hoc analysis of a randomised controlled trial. Lancet 374: 1243–1251
Chlebowski RT, Anderson GL, Manson JE, Schwartz AG, Wakelee H, Gass M, Rodabough RJ, Johnson KC, Wactawski-Wende J, Kotchen JM, Ockene JK, O’Sullivan MJ, Hubbell FA, Chien JW, Chen C, Stefanick ML (2010) Lung cancer among postmenopausal women treated with estrogen alone in the women’s health initiative randomized trial. J Natl Cancer Inst 102: 1413–1421
Dalla Valle L, Toffolo V, Nardi A, Fiore C, Armanini D, Belvedere P, Colombo L (2007) The expression of the human steroid sulfatase-encoding gene is driven by alternative first exons. J Steroid Biochem Mol Biol 107: 22–29
Dougherty SM, Mazhawidza W, Bohn AR, Robinson KA, Blankenship KA, Huff MO, McGregor WG, Klinge CM (2006) Gender difference in the activity but not expression of estrogen receptors alpha and beta in human lung adenocarcinoma cells. Endocr Relat Cancer 13: 113–134
Foster PA, Reed MJ, Purohit A (2008) Recent developments of steroid sulfatase inhibitors as anti-cancer agents. Anticancer Agents Med Chem 8: 732–738
Ganti AK, Sahmoun AE, Panwalkar AW, Tendulkar KK, Potti A (2006) Hormone replacement therapy is associated with decreased survival in women with lung cancer. J Clin Oncol 24: 59–63
Geisler J, Sasano H, Chen S, Purohit A (2011) Steroid sulfatase inhibitors: promising new tools for breast cancer therapy? J Steroid Biochem Mol Biol 125: 39–45
Greenlee H, Atkinson C, Stanczyk FZ, Lampe JW (2007) A pilot and feasibility study on the effects of naturopathic botanical and dietary interventions on sex steroid hormone metabolism in premenopausal women. Cancer Epidemiol Biomarkers Prev 16: 1601–1609
Havlíková H, Hill M, Hampl R, Stárga L (2002) Sex- and age-related changes in epitestosterone in relation to pregnenolone sulfate and testosterone in normal subjects. J Clin Endocrinol Metab 87: 2225–2231
Hirato K, Suzuki T, Hondo T, Saito H, Yanaihara T (1991) Steroid sulfatase activities in human leukocytes: biochemical and clinical aspects. Endocrinol Jpn 38: 597–602
Imai M, Nakamura T, Akiyama T, Horii A (2004) Infrequent somatic mutations of the ICAT gene in various human cancers with frequent 1p-LOH and/or abnormal nuclear accumulation of b-catenin. Oncol Rep 12: 1099–1103
Ishibashi H, Suzuki T, Suzuki S, Moriya T, Kaneko C, Nakata T, Sunamori M, Handa M, Kondo T, Sasano H (2005a) Estrogen inhibits cell proliferation through in situ production in human thymoma. Clin Cancer Res 11: 6495–6504
Ishibashi H, Suzuki T, Suzuki S, Niikawa H, Lu L, Miki Y, Moriya T, Hayashi S, Handa M, Kondo T, Sasano H (2005b) Progesterone receptor in non-small cell lung cancer - a potent prognostic factor and possible target for endocrine therapy. Cancer Res 65: 6450–6458
James VHT, McNeill JM, Lai LC, Newton CJ, Ghilchik MW, Reed MJ (1987) Aromatase activity in normal breast and breast tumour tissue: in vivo and in vitro studies. Steroids 50: 269–279
Kawai H, Ishii A, Washiya K, Konno T, Kon H, Yamaya C, Ono I, Minamiya Y, Ogawa J (2005) Estrogen receptor alpha and beta are prognostic factors in non-small cell lung cancer. Clin Cancer Res 11: 5084–5089
Koutras A, Giannopoulou E, Kritikou I, Antonacopoulou A, Evans TR, Papavassiliou AG, Kalofonos H (2009) Antiproliferative effect of exemestane in lung cancer cells. Mol Cancer 8: 109
Labrie F (1991) Intracrinology. Mol Cell Endocrinol 78: C113–C118
MacIndoe JH, Woods G, Jeffries L, Hinkhouse M (1988) The hydrolysis of oestrone sulfate and dehydroepiandrosterone sulfate by MCF-7 human breast cancer cells. Endocrinology 123: 1281–1287
Mah V, Seligson DB, Li A, Márquez DC, Wistuba II, Elshimali Y, Fishbein MC, Chia D, Pietras RJ, Goodglick L (2007) Aromatase expression predicts survival in women with early-stage non-small cell lung cancer. Cancer Res 67: 10484–10490
Mathers CD, Loncar D (2006) Projections of global mortality and burden of disease from 2002 to 2030. PLoS Med 3: e442
Miki Y, Suzuki T, Abe K, Suzuki S, Niikawa H, Iida S, Hata S, Akahira J, Mori K, Evans DB, Kondo T, Yamada-Okabe H, Sasano H (2010) Intratumoural localization of aromatase and interaction between stromal and parenchymal cells in the non-small cell lung carcinoma microenvironment. Cancer Res 70: 6659–6669
Mouridsen H, Gershanovich M, Sun Y, Pérez-Carrión R, Monnier A, Apffelstaedt J, Smith R, Sleeboom HP, Jänicke F, Pluzanska A, Dank M, Becquart D, Bapsy PP, Salminen E, Snyder R, Lassus M, Verbeek JA, Staffler B, Chaudri-Ross HA, Dugan M (2001) Superior efficacy of letrozole versus tamoxifen as first-line therapy for postmenopausal women with advanced breast cancer: results of a phase III study of the International Letrozole Breast Cancer Group. J Clin Oncol 19: 2596–2606
Nabholtz JM, Buzdar A, Pollak M, Harwin W, Burton G, Mangalik A, Steinberg M, Webster A, von Euler M for the Arimidex Study Group (2000) Anastrozole is superior to tamoxifen as first-line therapy for advanced breast cancer in postmenopausal women: results of a North American multicenter randomized trial. J Clin Oncol 18: 3758–3767
Nardi A, Pomari E, Zambon D, Belvedere P, Colombo L, Dalla Valle L (2009) Transcriptional control of human steroid sulfatase. J Steroid Biochem Mol Biol 115: 68–74
Nardulli AM, Greene GL, O’Malley BW, Katzenellenbogen BS (1988) Regulation of progesterone receptor message ribonucleic acid and protein levels in MCF-7 cells by oestradiol: analysis of estrogen’s effect on progesterone receptor synthesis and degradation. Endocrinology 122: 935–944
Niikawa H, Suzuki T, Miki Y, Suzuki S, Nagasaki S, Akahira J, Honma S, Evans DB, Hayashi S, Kondo T, Sasano H (2008) Intratumoural estrogens and estrogen receptors in human non-small cell lung carcinoma. Clin Cancer Res 14: 4417–4426
Paridaens RJ, Dirix LY, Beex LV, Nooij M, Cameron DA, Cufer T, Piccart MJ, Bogaerts J, Therasse P (2008) Phase III study comparing exemestane with tamoxifen as first-line hormonal treatment of metastatic breast cancer in postmenopausal women: the European organization for research and treatment of cancer breast cancer cooperative group. J Clin Oncol 26: 4883–4890
Pasqualini JR, Gelly C, Lecerf F (1986) Estrogen sulfates: biological and ultra-structural responses and metabolism in MCF-7 human breast cancer cells. Breast Cancer Res 8: 233–240
Pharm TM, Fujino Y, Matsuda S, Yoshimura T (2007) Premature mortality due to cancer in Japan, 1995 and 2005. Int J Cancer 127: 190–194
Sasano H, Suzuki T, Miki Y, Moriya T (2008) Intracrinology of estrogens and androgens in breast carcinoma. J Steroid Biochem Mol Biol 108: 181–185
Sato R, Suzuki T, Katayose Y, Miura K, Shiiba K, Tateno H, Miki Y, Akahira J, Kamogawa Y, Nagasaki S, Yamamoto K, Ii T, Egawa S, Evans DB, Unno M, Sasano H (2009) Steroid sulfatase and estrogen sulfotransferase in colon carcinoma: regulators of intratumoural estrogen concentrations and potent prognostic factors. Cancer Res 69: 914–922
Schabath MB, Wu X, Vassilopoulou-Sellin R, Vaporciyan AA, Spitz MR (2004) Hormone replacement therapy and lung cancer risk: a case-control analysis. Clin Cancer Res 10: 113–123
Selcer KW, Hedge PV, Li PK (1997) Inhibition of oestrone sulfatase and proliferation of human breast cancer cells by nonsteroidal (p-O-sulfamoyl)-N-alkanoyl tyramines. Cancer Res 57: 702–707
Shibuya K, Mathers CD, Boschi-Pinto C, Lopez AD, Murray CJ (2002) Global and regional estimates of cancer mortality and incidence by site: II. Results for the global burden of disease 2000. BMC Cancer 2: 37
Stabile LP, Lyker JS, Gubish CT, Zhang W, Grandis JR, Siegfried JM (2005) Combined targeting of the estrogen receptor and the epidermal growth factor receptor in non-small cell lung cancer shows enhanced antiproliferative effects. Cancer Res 65: 1459–1470
Stabile LP, Dacic S, Land SR, Lenzner DE, Dhir R, Acquafondata M, Landreneau RJ, Grandis JR, Siegfried JM (2011) Combined analysis of estrogen receptor beta-1 and progesterone receptor expression identifies lung cancer patients with poor outcome. Clin Cancer Res 17: 154–164
Stanway SJ, Delavault P, Purohit A, Woo LW, Thurieau C, Potter BV, Reed MJ (2007) Steroid sulfatase: a new target for the endocrine therapy of breast cancer. Oncologist 12: 370–374
Suzuki T, Nakata T, Miki Y, Kaneko C, Moriya T, Ishida T, Akinaga S, Hirakawa H, Kimura M, Sasano H (2003) Estrogen sulfotransferase and steroid sulfatase in human breast carcinoma. Cancer Res 63: 2762–2770
Tagawa N, Tamanaka J, Fujinami A, Kobayashi Y, Takano T, Fukata S, Kuma K, Tada H, Amino N (2000) Serum dehydroepiandrosteroine, dehydroepiandrosteroine sulfate, and pregnenolone sulfate concentrations in patients with hyperthyroidism and hypothyroidism. Clin Chem 46: 523–528
Thi MT, Baulieu EE, Milgrom E (1975) Comparison of the characteristics and of the hormonal control of endometrial and myometrial progesterone receptor. J Endocrinol 66: 349–356
Tilson-Mallett N, Santner SJ, Feil PD, Santen RJ (1983) Biological significance of aromatase activity in human breast tumours. J Clin Endocrinol Metab 57: 1125–1128
Utsunomiya H, Ito K, Suzuki T, Kitamura T, Kaneko C, Nakata T, Niikura H, Okamura K, Yaegashi N, Sasano H (2004) Steroid sulfatase and estrogen sulfotransferase in human endometrial carcinoma. Clin Cancer Res 10: 5850–5856
Vignon F, Terqui M, Westley B, Derocq D, Rochefort H (1980) Effects of plasma estrogen sulfates in mammary cancer cells. Endocrinology 106: 1079–1086
Wei LL, Krett NL, Francis MD, Gordon DF, Wood WM, O’Malley BW (1988) Multiple human progesterone receptor message ribonucleic acids and their autoregulation by progestin agonists and antagonists in breast cancer cells. Mol Endocrinol 2: 62–72
Weinberg OK, Marquez-Garban DC, Fishbein MC, Goodglick L, Garban HJ, Dubinett SM, Pietras RJ (2005) Aromatase inhibitors in human lung cancer therapy. Cancer Res 65: 11287–11291
Yamashita J, Tashiro K, Yoneda S, Kawahara K, Shirakusa T (1996) Local increase in polymorphonuclear leukocyte elastase is associated with tumor invasiveness in non-small cell lung cancer. Chest 109: 1328–1334
Zang EA, Wynder EL (1996) Differences in lung cancer risk between men and women: examination of the evidence. J Natl Cancer Inst 88: 183–192
Acknowledgements
We appreciate Erina Iwabuchi, Miki Mori, and Katsuhiko Ono (Department of Pathology, Tohoku University Graduate School of Medicine) for their skillful technical support.
Author information
Authors and Affiliations
Corresponding author
Additional information
This work is published under the standard license to publish agreement. After 12 months the work will become freely available and the license terms will switch to a Creative Commons Attribution-NonCommercial-Share Alike 3.0 Unported License.
Supplementary Information accompanies this paper on British Journal of Cancer website
Supplementary information
Rights and permissions
From twelve months after its original publication, this work is licensed under the Creative Commons Attribution-NonCommercial-Share Alike 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-sa/3.0/
About this article
Cite this article
Iida, S., Kakinuma, H., Miki, Y. et al. Steroid sulphatase and oestrogen sulphotransferase in human non-small-cell lung carcinoma. Br J Cancer 108, 1415–1424 (2013). https://doi.org/10.1038/bjc.2013.84
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/bjc.2013.84
Keywords
This article is cited by
-
Exploring estrogenic activity in lung cancer
Molecular Biology Reports (2017)
