
Abstract
Euglossini is an important tribe of Neotropical bees found primarily in wet forest environments, although their distribution extends to seasonal tropical forests and mountainous grassland habitats. However, little is known about the geography and diversity of euglossine bees in mountain regions, particularly in the Brazilian semi-arid Cerrado vegetation. We evaluated the distribution of euglossine bees along an elevational gradient in Serra do Cipó, Brazil, where the vegetation varies from savanna (Cerrado) at lower elevations to rupestrian grasslands in mountain summits, and an ecotone of cerrado/rupestrian grassland at mid elevations. We also tested whether bee species richness and abundance were influenced by environmental variables (vegetation and climate) that change with elevation. Bee abundance decreased with elevation, while species richness peaked at mid elevations. Species replacement along the elevational gradient led to differences in species composition between sites; but species diversity remained relatively constant, along the gradient. Abundance and diversity patterns were similar between the dry and rainy seasons, despite differences in species composition. We emphasize the importance of preserving corridors of mesic environments in the conservation of euglossine bees for maintaining ecosystem services in mountain savannas.
Similar content being viewed by others

1 Introduction
Bees are of great relevance worldwide because they provide important ecosystem services through the pollination of native and cultivated plant species (Michener 2007, Winfree and Kremen 2007, Garibaldi et al. 2013). With more than 20,000 species, bees are the most important group of animal pollinators on earth, both in natural and anthropized environments (Potts et al. 2016). Some groups of bees, such as the tribe Euglossini, have evolved tight relationships with certain groups of plants (i.e., orchids), where male bees pollinate orchid flowers as they collect floral volatiles produced as attractants (Dressler 1982). In this relationship, many orchid species exhibit high dependence on one or a few euglossine species, although the relationship is asymmetric and bee species may use floral resources from different orchid species (Ramírez et al. 2011) and from a number of plant families used as sources of nectar, pollen, and resins (Roubik and Hanson 2004, Ackerman and Roubik 2012). Despite the great importance of euglossine bees as exclusive pollinators of many plant species, little is known about patterns of diversity, distribution, and abundance of these bees in the Brazilian Cerrado.
One way to study the factors that determine bee abundance and distribution is to use mountain regions as natural labs, since mountains exhibit great geophysical and environmental variations at small spatial scales along the elevational gradient (Fernandes and Price 1992; Körner 2007; Fernandes et al. 2016). Most animal species have greater diversity in tropical latitudes (e.g., Gaston 2000); however, along elevational gradients, at least three patterns of species richness distribution have been described for entomological fauna (Ackerman 1983; Rahbek 1995). The first and most common pattern in tropical and temperate regions is the decline of insect species richness as elevation increases (e.g., ants: Araújo and Fernandes 2003; bees: Ackerman 1983, Hoiss et al. 2012; termites: Nunes et al. 2016; aculeata: Perillo et al. 2017). Second, an opposite pattern has been described for some insect groups, where species richness increases with elevation (e.g., herbivorous insects: Araújo and Fernandes 2003). A third pattern describes a distribution curve, where species richness peaks at intermediate elevations (e.g., beetles: Olson 1994; butterflies: Fleishman et al. 1998). However, few studies have documented bee abundance diversity patterns along elevational gradients (but see, Hoiss et al. 2012; Perillo et al. 2017).
Euglossine bees are most diverse at low latitudes and low to mid elevations, and they are typically forest organisms with greater abundance in the humid tropics, particularly in Central America and the Amazon forest (Dressler 1982; Nemésio 2008, 2016). Some large-bodied genera (e.g., Eufriesea and Eulaema) are common in mid to high elevations in Central America and the Andes, while euglossine bee communities in Costa Rica and Panama are most diverse at middle elevations (approx. 800 m, Roubik and Hanson 2004). Although euglossine bees have lower species richness and abundance in savanna vegetation, some species with greater tolerance to open areas are present in the Caatinga (Brazilian dry forest) and the Cerrado, where they play an important role in provisioning pollination services to native plants and crops (e.g., Silveira et al. 2002; Zanella 2000; Pokorny et al. 2014). In the Cerrado, euglossine bees are important pollinators of several plant families, such as Orchidaceae, Araceae, Euphorbiaceae, Gesneriaceae, and Solanaceae (Williams and Whitten 1983; Silveira et al. 2002). A peculiar characteristic of these bees, exclusive to males, is that they collect organic volatiles from orchids, while both males and females may collect nectar and pollen from other plant families (Williams 1982; Williams and Whitten 1983; Roubik and Hanson 2004). However, knowledge about the diversity, distribution, and ecology of euglossine bees in the open semi-arid habitats of Neotropical mountains is incipient (Nemésio and Silveira 2007; Faria and Silveira 2011; Moreira et al. 2017). One of the knowledge gaps in these ecosystems is the distribution of bee species along elevational gradients.
Another aspect of relevance in the study of bee abundance and diversity is seasonality. Euglossine bees may show strong seasonality, with greater abundance in the rainy season than in the dry season, when only species less sensitive to dry weather conditions persist (Rebêlo and Garófalo 1991). Euglossine bees are significantly affected by wind, temperature, plant diversity, availability of food resources, and access to nesting sites (Nemésio and Vasconcelos 2013; Antonini et al. 2016, 2017). At higher elevations in mountainous ecosystems, there is an increase in mean wind speed, a decrease in temperature, and a simplification of vegetation structure (e.g., Lawton et al. 1987), variables that may limit the distribution of euglossine bees in these environments, although large-bodied species are known to thrive in mid to high elevations in some regions (Roubik and Hanson 2004; McCravy et al. 2016).
The mountains in the Espinhaço Range in eastern Brazil represent the second largest mountain range in South America and offer ideal conditions for testing hypotheses about environmental filters that influence species diversity (Fernandes and Price 1988; Fernandes et al. 2018). Studies on butterflies, dung beetles, galling insects, termites, and free-lining herbivores in these mountains have reported the three patterns of species richness previously described for elevational gradients (see review in Fernandes et al. 2016). In this study, we evaluated the species diversity and abundance of euglossine bees in the Espinhaço mountains of Brazil. The following hypotheses were tested: (i) the composition of the euglossine bee assemblages will be determined by seasonality; (ii) the assembly of euglossine bees varies along the elevational gradient, with abundance and richness of euglossine species decreasing with increasing elevation; and (iii) plant species richness and abundance as well as climatic variables are important factors that regulate the abundance and richness of euglossine bees along the elevation gradient.
2 Materials and methods
2.1 Study sites
The present study was conducted in the center-south portion of Serra do Espinhaço, in a locality known as Serra do Cipó, in the municipality of Santana do Riacho, Minas Gerais (MG), Brazil. The vegetation in this region is dominated by outcrops (quartzite) (for details, see Fernandes et al. 2016). Soils are sandy, shallow, and nutrient poor, and with high aluminum concentration and low water retention capacity (e.g., Schaefer et al. 2016; Silveira et al. 2016). The climate of the region is of the Cwb type, according to Köppen classification, mesothermic, with wet summers and dry winters, and with a mean annual precipitation of 1500 mm concentrated during wet season (November–April) and a mean annual temperature of 20 °C (Madeira and Fernandes 1999).
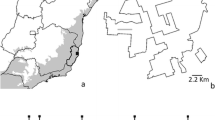
Fieldwork was conducted in seven areas at the sites of the Long-Term Ecological Research Program—Campos do Cipó (PELD-CRSC/CNPQ; 19° 16′–19° 21′ S, 43° 32′–43° 36′ W). These sites were located between elevations of 800 and 1400 m at intervals of approximately 100 m and separated by at least 2 km (Figure 1). The seven study areas comprise a physiognomic mosaic of vegetation, where soil and vegetation are quite heterogeneous. The savanna and grassland study sites vary from cerrado sensu stricto to rupestrian grasslands (for details, see Fernandes et al. 2016, Mota et al. 2018). The term rupestrian is related to the unique vegetation found in rocky outcrops in the Cerrado of Brazil.

Map of seven study sites along an elevational gradient in the Serra do Cipó (Minas Gerais, Brazil): a location of Espinhaço Mountain Range; b sampling areas at each elevation.
2.2 Sampling of bees
In order to evaluate if male euglossine abundance and richness respond to seasons and altitudinal gradient, male euglossine bees were sampled in the wet season (November–April) and the dry season (May–October) of 2017 and those from each one of the seven elevations were used.
At each collecting period, baits with fragrances (benzyl acetate, beta-ionone, cineole, methyl cinnamate, eugenol, methyl salicylate, and vanillin, after a pilot collection at the study site) were exposed for 48 h, following Bezerra and Martins (2001) for a total of 4704 trapping hours (seven traps in each one of the seven sampling plots—in the dry and wet seasons = 63 traps), and were placed 2 m apart from each other and fixed at about 1.5 m height at tree trunks (Antonini et al. 2016). Captured specimens were collected at 16:00 h of the second day, and frozen until they were prepared for taxonomic identification. All materials were identified using the keys of Rebelo and Moure (1995) and Nemésio (2009), and deposited in the entomological collection of the Laboratory of Evolutionary Ecology and Biodiversity at Universidade Federal de Minas Gerais.
2.3 Environmental variables
Climatic data from study sites along the elevational gradient were obtained from meteorological monitoring towers equipped with the Onset HOBO® U30 data logger. These stations were distributed at each study site, i.e., between 800 and 1400 m (Figure 1). The following climatic parameters were obtained: air temperature and humidity, wind speed, and solar radiation. For analyses, we used the mean of each variable over the study period. In order to analyze the local influence of vegetation on euglossine bees, we used data from plant community studies performed in the same areas where we conducted our sampling (see Mota et al. 2018). In this plant survey, 13 plots (10 × 10 m) were established along the elevational gradient and all woody plants with soil height diameter (DSH) ≥ 1 cm were sampled.
2.4 Data analysis
Data analysis was performed in the R environment (R Core Team 2017). In order to determine whether the assemblage of euglossine bees changes between the dry and rainy seasons, the Bray-Curtis dissimilarity matrix (previously calculated) was used to perform a non-metric multidimensional scaling (nMDS), followed by an analysis of similarity (ANOSIM). Subsequently, the proportional abundance of each species in the two seasons was plotted.
To test the hypothesis that euglossine bee assemblages vary with elevation and seasonality, generalized linear models (GLMs) were constructed, with abundance and richness designated as response variables, and elevation and “season of the year” as explanatory variables. The models used a negative binomial and quasi-Poisson distributions, respectively. Subsequently, the Chi-square test was used for negative binomial (O'Hara and Kotze 2010) and F-test for quasi-Poisson.
Species composition was evaluated through the Bray-Curtis dissimilarity metric (based on the log-transformed matrix of species abundance) using a distance-based linear model (DistLM) using also elevation and the interaction between elevation and seasons as the explanatory variables. This was followed by a distance-based redundancy analysis (dbRDA). To evaluate how the assembly changed along the gradient, the Bray-Curtis dissimilarity partitioning methodology was used to calculate the components of variation of the bee assemblages (Baselga 2017). This approach allowed us to evaluate how much the bee assemblage changes and how it is organized through the following components: (i) balanced variation of abundance, where the loss of individuals of one species is compensated by the occurrence of individuals of a different species; and (ii) abundance gradient, which accounts for the component of equal changes on the number of individuals of all species (Baselga 2017). For this analysis, the betapart package was used (Baselga 2017).
To test the influence of vegetation and climate on abundance and species richness, the dry and rainy seasons were evaluated separately, since the great differences in climate between the two seasons could generate noise in the models. Initially, the relationship between climatic and vegetation variables was tested separately by Spearman’s correlation and those with a high correlation (r > 0.70) were eliminated. The variables used for analyses of rainy season were as follows: amount of rainfall, temperature, air humidity, and plant species richness. For dry season analyses, we used the same variables except for temperature (high correlation with rainfall). Four GLMs were then constructed with the respective explanatory variables for each season and with abundance and richness of euglossine bees as response variables. These models used a quasi-Poisson distribution, except for abundance in the dry season, which used a negative binomial distribution.
3 Results
A total of 786 euglossine bee individuals were collected, distributed into three genera and 14 species (Figure 2). The genus Euglossa was the most abundant, with 472 individuals (11 species, 60.1%), followed by the genus Eulaema with 313 individuals (2 spp., 39.8%). A single individual of the genus Exaerete was collected, which represented 0.1% of the euglossine assembly in this study. The composition of the euglossine bee assemblage varied between dry and rainy seasons along the elevational gradient (R = 0.441, p = 0.001, Figure 3). Assemblages collected at different elevations were more similar to each other in the rainy season than in the dry season. This was evidenced by the lower dispersion of points for rainy season in the nMDS (Figure 4a). Additionally, the total number of euglossine bee species was different between the two seasons, with 14 species sampled in the rainy season and 10 species in the dry season.

Distribution of euglossine species along an elevational gradient in Serra do Cipó (MG, Brazil): The sizes of the circles represent the abundance of species at each elevation (log (N + 1)). White and full lines, gray and dotted lines, and circles represent the proportion within post-rainy and dry seasons, respectively.

Variation in the assemblages of euglossine bees sampled along an elevational gradient in the Serra do Cipó (MG, Brazil) in the dry (gray) and post-rainy seasons (white): a non-metric multidimensional scaling (nMDS), b proportion of the total of individuals of Euglossini species collected in the dry (gray) and post-rainy (white) seasons.

Partition of diversity based on Bray-Curtis dissimilarity of the euglossine bee assemblage in Serra do Cipó (MG, Brazil), considering each elevational pair (consecutive) and all points (total), for a overall community, b post-rainy season, and c dry season. The point on each bar indicates dissimilarity between each pair of bee assemblages (Bray-Curtis dissimilarity). The gray bars represent the proportion of the assemblage that is balanced in abundance, with the substitution of individuals of certain species by individuals of other species, and white bars represent the proportion of equal loss of individuals among all species (see Baselga 2017).
Elevation was negatively associated with euglossine bee abundance (X2 = 8.40; p = 0.004; Figure 4a). In contrast, elevation was not statistically related to bee species richness (F = 3.25, p = 0.102, Figure 4b) at Serra do Cipó. Species richness was similar between study sites at 800 to 1000 m elevation (8–10 species), which peaked at 1100 m (13 species) in the rainy season and had the lowest richness at the elevation in the dry season (Figure 4b). Despite the lack of statistical significance, richness showed a decreasing trend with increasing elevation to reach a minimum number of species at 1400 m (6 out of 14; Figure 4b). Regarding the differences between season, both abundance (X2 = 8.19, p = 0.004, Figure 4a) and richness (F = 8.53, p = 0.015, Figure 4b) showed higher values in the rainy season but there were no differences in the patterns between seasons neither for abundance (X2 = 1.00, p = 0.317, Figure 4a) nor for richness (F ≅ 0, p = 0.979, Figure 4b).
The species composition of the euglossine bee assemblage varied along the elevational gradient in Serra do Cipó (F = 3.53, p = 0.002), but there was no change in this variation due to the composition changes between seasons (F = 1.29, p = 0.279). Considering both seasons together, the diversity partition showed that the total euglossine assemblage along the gradient was 70% dissimilar (Figure 5), with two components explaining this dissimilarity in similar proportions: the gradual loss of individuals among the species (approximately 50%, Figure 5a), and the balanced variation in abundance, where loss of individuals in some euglossine species was compensated by the gain of individuals in other species (Figure 5a). Dissimilarity between consecutive elevational pairs from 800 to 1000 m reflected the bee abundance gradient (an equal loss of individuals by all species). From 1000 m up, the dissimilarity between bee assemblages was explained by an increase in species replacement (Figure 5a). Considering the season, we observed that these patterns in composition are driven by the rainy season, except for the proportion of balanced variation explaining the high shifts in composition between 1000 and 1100 m (Figure 5c). We did not find a significant contribution of climatic and vegetation variables to changes of species richness or abundance of euglossine bees along the elevational gradient in either season (Table I).

Variation in diversity parameters of the euglossine bee assemblage between the dry (gray dots and dashed line) and post-rainy (white dots and full line) seasons along an elevation gradient in Serra do Cipó (MG, Brazil): a species abundance and b species richness. Explanatory variables are represented by Ele (elevation), Sea (season), and Int. (interaction between the two variables).
4 Discussion
This study found a reduction in euglossine bee abundance with increasing elevation, changes in species community composition, and no statistical differences in species richness along an elevation gradient in Serra do Cipó (MG, Brazil). In relation to euglossine bee abundance, results corroborated the pattern observed for other bee assemblages along elevational gradients in other mountains of the world (e.g., Nemésio 2008; Hoiss et al. 2012; Perillo et al. 2017). This pattern is often related to the influence of environmental filters in mountain habitats, such as cold temperatures and strong winds that affect the flying capacity of bees (Kleinert-Giovannini 1982; Fernandes and Price 1988; Lomolino 2001). Although the climatic variables measured in this study were not related to bee abundance, vegetation structure was. At lower elevations, the Cerrado physiognomy is characterized by dense and stratified vegetation, which becomes shorter and structurally simpler at higher elevation habitats (Mota et al. 2018). Most euglossine bee species are associated with forest environments (Silveira et al. 2015) and they build their nests in pre-existing cavities in tree trunks or on the ground along ravines or hillsides (Silveira et al. 2002). Thus, lower elevation habitats may offer a greater number of nesting sites due to the greater coverage of woody plants (Mota et al. 2018), as well as soil protection against erosion. Taken together, these features may have favored greater populations of euglossine bees in the Cerrado environments in contrast to the more open habitats of rupestrian grasslands at higher elevations.
The richness of euglossine bee species in Serra do Cipó did not correlate with elevation or even with the vegetation and climatic variables measured. This lack of association with environmental variables apparently contradicts the findings from other studies (e.g., Hoiss et al. 2012; Antonini et al. 2016, 2017), but we offer potential explanations that might apply to these mountains. The rupestrian grassland is a highly complex habitat with gallery forests spreading out along the elevational gradient (see Fernandes and Price 1988, 1992; Coelho et al. 2016; Fernandes et al. 2016). As euglossine bees are mostly associated to forest habitats, where they have a greater availability of resources and nesting sites, bee species would have been attracted to the study sites primarily from neighboring forest habitats that are also found throughout the elevational gradient. It is well known that euglossine bees are capable of flying up to 23 km before returning to their nest on the same day (Janzen 1971). Wikelski et al. (2010), using aerial telemetry, showed that Exaerete frontalis individuals were found ca. 1.9 km in a 5-h flight. Although euglossine bees search for resources such as pollen, nectar, and volatile compounds in the flowers of rupestrian grasslands, many species that nidify in the forested habitats may use these habitats only as foraging sites (Azevedo et al. 2008). One important resource for male of Euglossine, volatile compounds from orchids, is well represented in the study area. According to Giulietti et al. (1987) in the campos rupestres of Serra do Cipó, there is 71 described species of orchids.
The observed changes in the composition of euglossine bee assemblages were due not only to the loss of individuals in some populations (abundance gradient), but also to the increase of individuals of other species (Baselga 2017), particularly above 1000 m. This may also influence the lack of a trend in the distribution of richness and diversity of euglossine species along the gradient, as also reported by Nemésio (2008). Overlap between species from lower and higher elevations has also been proposed as an explanation to the greater richness of euglossine bee species found at mid elevation sites in Costa Rica and Panama (Roubik and Ackerman 1987; Roubik and Hanson 2004). A potential explanation for this pattern is that, within a geographical domain, the random allocation of species with different distribution ranges produces a peak in species richness where the species overlap in the middle of the domain (Colwell and Lees 2000).
The most important climatic characteristic in the mountains of Serra do Espinhaço is the strong seasonality, which influences the phenology of its flora and fauna (Fernandes et al. 2016, 2018). Euglossine bees responded to seasonality all along the gradient, as indicated by the large difference in the composition of the bee assemblage between dry and rainy seasons. In the Brazilian Atlantic forest, it has been shown that some species of euglossine bees are highly seasonal, presenting an intimate relationship with the availability of floral resources (Rocha-Filho and Garófalo 2013). Likewise, studies in Panama have shown that some euglossine bee species have reduced activity or lower population sizes during certain times of the year (Ackerman 1983). These changes are thought to be related to spatial movements between different habitats associated with availability of food resources (Janzen 1971; Ackerman 1983). Throughout the elevational gradient, flowering of different plant species occurs all year round, although there is an increase in the number of flowering species between December and May (Belo et al. 2013; Rocha et al. 2016). This period corresponds to the rainy season, during which the highest similarity in composition of euglossine bee assemblages was recorded between different elevation sites (evidenced by the proximity of points in the NMDS, Figure 3a). Likewise, the rainy season corresponded to the period with the highest species richness, as shown also in other studies (Carvalho et al. 2006; Ramalho et al. 2009, Ackermann and Roubik 2012, McCravy et al. 2016).
We found a total of 14 species of euglossine bees, a greater number than previously found in other Cerrado regions (e.g., Nemésio 2008: 9 species, Serra do Carraça, MG; Faria and Silveira 2011: 8 species, Brasilândia de Minas, MG; Viotti et al. 2013: 11 species, Serra do Espinhaço, Diamantina, MG; Nemésio 2016: 11 species, Panga Ecological Station, MG; Antonini et al. 2016: 12 species, State Park of Rio Preto, MG). We expect that the number of euglossine bees in the Espinhaço mountain range should be much larger simply due the larger area and spread of mountains over more than 10° latitude.
The species composition of euglossine bee assemblages changed at higher elevations, corroborating the pattern found for most groups of bees (Silveira and Cure 1993). At intermediate elevations, singletons were found, such as Euglossa violaceinfrons and Exaerete smaragdina. This second species is a rare cleptoparasite species recorded in the Cerrado biome (Nemésio 2016). The rare species of euglossine bees reported here were restricted to elevations where the rupestrian grassland vegetation was predominant. These data corroborate another study that focused on the distribution of bees in the Espinhaço rupestrian grasslands, which indicated that while population abundances are low, there is a high number of rare species in these environments (Azevedo et al. 2008).
This study documented elevational variation in species composition of euglossine assemblages with elevation, with the loss of individuals of some species and gain of individuals of other species at higher elevations. Given that global warming is one of the factors associated with the disappearance of bees (Kerr et al. 2015) and that mountain regions are the first to experience changes due to climate change, we suggest prioritizing the study of these organisms in mountain regions. The occurrence of severe climatic changes may result in a significant reduction in the populations of the most vulnerable species, or even lead to the extinction of those species restricted to higher elevational ranges (e.g., Eufriesea nigrohirta, endemic to the “campos ruprestres,” according to Nemesio 2005) whose populations are smaller and thus probably less stable (Raxworthy et al. 2008; Sundqvist et al. 2013). Long-term studies like those conducted by Roubik and Ackerman (1987) in Panama shed light onto the population stability of different Euglossine bees and demonstrate that some species maintain stable populations over time while others do not. Our study should serve as a baseline for conducting long-term sampling of bees to determine demographic patterns and vulnerability.
Bees are important pollinators in open habitats (Antonini et al. 2005) and agroecosystems (Giannini et al. 2017), and they can account for 70% of the pollination services in rupestrian grasslands (Guerra et al. 2016). Rupestrian grasslands occur within a matrix that contains patches of forest vegetation including gallery forests at lower elevations and forest beds at higher elevations (Coelho et al. 2016; Fernandes et al. 2016). These forest formations could represent an important source of pollinators for the adjacent rupestrian grasslands. Human modification of these habitats could also drive changes in bee communities. This is evident in the elevational gradient where non-forest vegetation formations prevail, and most plants depend on animals for pollination (Hoiss et al. 2012; Guerra et al. 2016). This study also indicates the importance of urgently increasing the knowledge about euglossine bees in tropical non-forest formations and their relevance in the reproductive success of endemic flora, as well as in the functioning of these endangered ecosystems.
Change history
17 July 2020
Following publication of the original article, the author group noticed an error related to the presentation of Figs. 4 and 5.
References
Ackerman J.D. (1983) Specificity and the mutual dependency of the orchid euglossine bee interaction. Biol. J. Linn. Soc. 20, 301–314
Antonini Y., Souza H.G., Jacobi C.M., Mury F.B. (2005) Diversidade e comportamento dos insetos visitantes florais de Stachytarpheta glabra Cham. (Verbenaceae), em uma área de campo ferruginoso, Ouro Preto, MG. Neotrop. Entomol. 34, 555–564
Antonini Y., Silveira R.A., Oliveira M.L., Martins C., Oliveira R. (2016) Orchid bee fauna responds to habitat complexity on a savanna area (Cerrado) in Brazil. Sociobiol. 63, 819–825
Antonini Y., Machado C.B., Galetti P.M. Jr., Oliveira M., Dirzo R., Fernandes G.W. (2017) Patterns of orchid bee species diversity and turnover among forested plateaus of central Amazonia. PLoS One 12, e0175884
Araújo L.M., Fernandes G.W. (2003) elevational patterns in a tropical ant assemblage and variation in species richness between habitats. Lundiana 4, 103–109
Azevedo A.A., Silveira F.A., Aguiar C.M.L., Pereira V.S. (2008) Fauna de abelhas (Hymenoptera, Apoidea) nos campos rupestres da Cadeia do Espinhaço (Minas Gerais e Bahia, Brasil): riqueza de espécies, padrões de distribuição e ameaças para conservação. Megadiversidade 4, 126–157
Baselga A. (2017) Partitioning abundance-based multiple-site dissimilarity into components: balanced variation in abundance and abundance gradients. Methods Ecol. Evol. 8, 799–808
Belo R.M., Negreiros D., Fernandes G.W., Silveira F.A.O., Ranieri B.D., Morelato P.C. (2013) Fenologia reprodutiva e vegetativa de arbustos endêmicos de campo rupestre na Serra do Cipó, Sudeste do Brasil. Rodriguésia 64, 817–828
Bezerra C.P., Martins C.F. (2001) Diversidade de Euglossinae (Hymenoptera, Apidae) em dois fragmentos de Mata Atlântica localizados na região urbana de João Pessoa, Paraíba, Brasil. Rev. Bras. Zool. 18, 823–835
Carvalho C.C.D., Rêgo M.M.C., Mendes F.N. (2006) Dinâmica de populações de Euglossina (Hymenoptera, Apidae) em mata ciliar, Urbano Santos, Maranhão, Brasil. Iheringia Sér. Zool. 96, 249–256
Coelho M.S., Fernandes G.W., Pacheco P., Diniz V., Meireles A. et al (2016) Archipelago of montane forests surrounded by rupestrian grasslands: new insights and perspectives. In Fernandes G.W. (Ed.), Ecology and Conservation of Mountain-top Grasslands in Brazil (pp. 129–153). Springer International, Switzerland
Colwell R.K., Lees D.C. (2000) The mid-domain effect: geometric constraints on the geography of species richness. Trends Ecol. Evol. 15, 70–76
Dressler R.L. (1982) Biology of the orchid bees. Annu. Rev. Ecol. Syst. 13, 373–394
Faria L.R.R., Silveira F.A. (2011) The orchid bee fauna (Hymenoptera, Apidae) of a core area of the Cerrado, Brazil: the role of riparian forests as corridors for forest-associated bees. Biota Neotrop. 11, 87–94
Fernandes G.W., Price P.W. (1988) Biogeographical gradients in galling species richness. Oecologia. 76, 161–167
Fernandes G.W., Price P.W. (1992) The adaptive significance of insect gall distribution: survivorship of species in xeric and mesic habitats. Oecologia 90, 14–20
Fernandes G.W., Almeida H.A., Nunes C.A., Xavier J.H.A., Cobb N.S. (2016) Cerrado to rupestrian grasslands: patterns of species distribution and the forces shaping them along an altitudinal gradiente. In Fernandes G.W. (Ed.), Ecology and conservation of mountaintop Grasslands in Brazil (pp. 345–377). Springer International Publishing, Switzerland
Fernandes G.W., Pedroni F., Sancheza M., Scariot A., Aguiar L.M.S. et al (2018) Cerrado: em busca de soluções sustentáveis. Vertente, Rio de Janeiro
Fleishman E., Austin G.T., Weiss A.D. (1998) An empirical test of Rapoport’s rule: elevational gradients in montane buttery communities. Ecology 79, 2482–2493
Garibaldi L.A., Steffan-Dewenter I., Winfree R., Aizen M.A., Bommarco R., Cunningham S.A., Kremen C., Carvalheiro L.G., Harder L.D., Afik O., Bartomeus I., Benjamin F., Boreux V., Cariveau D., Chacoff N.P., Dudenhöffer J.H., Freitas B.M., Ghazoul J., Greenleaf S., Hipólito J., Holzschuh A., Howlett B., Isaacs R., Javorek S.K., Kennedy C.M., Krewenka K.M., Krishnan S., Mandelik Y., Mayfield M.M., Motzke I., Munyuli T., Nault B.A., Otieno M., Petersen J., Pisanty G., Potts S.G., Rader R., Ricketts T.H., Rundlöf M., Seymour C.L., Schüepp C., Szentgyörgyi H., Taki H., Tscharntke T., Vergara C.H., Viana B.F., Wanger T.C., Westphal C., Williams N., Klein A.M. (2013) Wild Pollinators Enhance Fruit Set of Crops Regardless of Honey Bee Abundance. Science 339, 1608–1611
Gaston K.J. (2000) Global patterns in biodiversity. Nature. 405, 220–227
Giannini T.C., Costa W.F., Cordeiro G.D., Imperatriz-Fonseca V.L., Saraiva A.M., Biesmeijer J. et al (2017) Projected climate change threatens pollinators and crop production in Brazil. PLoS One 12, e0182274
Giulietti A.M., Menezes N.L., Pirani J.R., Meguro M., Wanderley M.G.L. (1987) Flora da Serra do Cipó, Minas Gerais: caracterização e lista das espécies. Boletim de Botânica da Universidade de São Paulo 9, 1–151
Guerra T.J., Carstensen D.W., Morellato L.P.C., Silveira F.A., Costa F.V. (2016) Mutualistic interactions among free-living species in Rupestrian Grasslands. In Fernandes G.W. (Ed.), Ecology and Conservation of Mountaintop Grasslands in Brazil (pp. 291–314). Springer International Publishing, Switzerland
Hoiss B., Krauss J., Potts S.G., Roberts S., Steffan-Dewenter I. (2012) Altitude acts as an environmental filter on phylogenetic composition, traits and diversity in bee communities. Proc. R. Soc. B Biol. Sci. 279, 4447–4456
Janzen D.H. (1971) Euglossine bees as long-distance pollinators of tropical plants. Science. 171, 203–205
Kerr J.T., Pindar A., Galpern P., Packer L., Potts S.G., Roberts S.M., Rasmont P., Schweiger O., Colla S.R., Richardson L.L., Wagner D.L., Gall L.F., Sikes D.S., Pantoja A. (2015) Climate change impacts on bumblebees converge across continents. Science 349, 177–180
Kleinert-Giovannini A. (1982) The influence of climatic factors on flight activity of Plebeia emerina Friese (Hymenoptera, Apidae, Meliponinae) in winter. Rev. Brasil. Ent. 26, 1–13
Körner C. (2007) The use of ‘altitude’ in ecological research. Trends Ecol. Evol. 22, 569–574
Lawton J.H., Macoarvin M., Heads P.A. (1987) Effects of altitude on the abundance and species richness of in insect’s herbivores. Ecology. 56, 147–160
Lomolino M.V. (2001) Elevation gradients of species-density: historical andprospective views. Glob. Ecol. Biogeogr. 10, 3–13
Madeira J.A., Fernandes G.W. (1999) Reproductive phenology of sympatric species of Chamaecrista (Leguminosae) in Serra do Cipó, Brazil. J. Trop. Ecol. 15, 463–479
McCravy K.W., Van Dyke J., Creedy T.J., Roubik D.W. (2016) Orchid bees (Hymenoptera: Apidae: Euglossini) of Cusuco National Park, state of Cortés, Honduras. Fla. Entomol. 99, 765–769
Michener C.D. (2007) The bees of the world, 2nd edn. Johns Hopkins Press, Baltimore
Moreira E.F., Santos R.L.S., Silveira M.S., Boscolo D., Neves E.L., Viana B.F. (2017) Influence of landscape structure on Euglossini composition in open vegetation environments. Biota. Neotrop. 17, e20160294
Mota G.S., Luz G.R., Mota N.M., Coutinho E.S., Veloso M.D.M., Fernandes G.W., Nunes Y.R.F. (2018) Changes in species composition, vegetation structure, and life forms along an altitudinal gradient of rupestrian grasslands in south-eastern Brazil. Flora. 238, 32–42
Nemésio A. (2005) Description of the male Eufriesea nigrohirta (Friese, 1899) (Hymenoptera: Apidae) with comments on the holotype, species biology and distribution. Lundiana 6, 41–45
Nemésio A. (2008) Orchid bee community (Hymenoptera, Apidae) at an altitudinal gradient in a large forest fragment in southeastern Brazil. Rev. Bras. Zooc. 10, 251–258
Nemésio A. (2009) Orchid bees (Hymenoptera: Apidae) of the Brazilian Atlantic Forest. Zootaxa. 2041, 1–242
Nemésio A. (2016) Orchid bees (Hymenoptera, Apidae) from the Brazilian savanna-like ‘Cerrado’: how to adequately survey under low population densities? North-Western. J. Zool. 12, 230–238
Nemésio A., Silveira F.A. (2007) Diversity and distribution of orchid bees (Hymenoptera: Apidae) with a revised checklist of species. Neotrop. Entomol. 36, 874–888
Nemésio A., Vasconcelos H.L. (2013) Beta diversity of orchid bees in a tropical biodiversity hotspot. Biodivers. Conserv. 22, 1647–1661
Nunes C.A., Quintino A.V., Constantino R., Negreiros D., Reis R. Jr., Fernandes G.W. (2016) Patterns of taxonomic and functional diversity of termites along a tropical elevational. Biotropica. 49, 186–194
O'Hara R.B., Kotze D.J. (2010) Do Not Log-Transform Count Data. Methods in Ecology and Evolution. Br. Ecol. Soc. 1, 118–122. https://doi.org/10.1111/j.2041-210X.2010.00021.x
Olson D.M. (1994) The distribution of leaf litter invertebrates along a Neotropical altitudinal gradient. J. Trop. Ecol. 10, 129–150
Perillo L.N., Neves F.S., Antonini Y., Martins R.P. (2017) Compositional changes in bee and wasp communities along Neotropical mountain altitudinal gradient. PLoS One 12, e0182054
Pokorny T., Loose D., Dyker G., Quezada-Euán J.J.G., Eltz T. (2014) Dispersal ability of male orchid bees and direct evidence for long-range flights. Apidologie. 46, 224–237
Potts S.G., Imperatriz-Fonseca V., Ngo H.T., Aizen M.A., Biesmeijer J.C., Breeze T.D., Dicks L.V., Garibaldi L.A., Hill R., Settele J., Vanbergen A.J. (2016) Safeguarding pollinators and their values to human well-being. Nature. 540, 220–229
Rahbek C. (1995) The elevational gradient of species richness: a uniform pattern? Ecography. 18, 200–205
Ramalho A.V., Gaglianone M.C., Oliveira M.L.D. (2009) Assembleias de abelhas Euglossina (Hymenoptera, Apidae) em fragmentos de Mata Atlântica no sudeste do Brasil. Rev. Bras. Entomol. 53, 95–101
Ramírez S.R., Eltz T., Fujiwara M.K., Gerlach G., Goldman-Huertas B., Tsutsui N.D., Pierce N.E. (2011) Asynchronous diversification in a specialized plant-pollinator mutualism. Science 333, 1742–1746
Raxworthy C.J., Pearson R.G., Rabibisoa N., Rakotondrazafy A.M., Ramanamanjato J.B. et al (2008) Extinction vulnerability of tropical montane endemism from warming and upslope displacement: a preliminary appraisal for the highest massif in Madagascar. Glob. Chang. Biol. 14, 1703–1720
Rebêlo J.M.M., Garófalo C.A. (1991) Diversidade e sazonalidade de machos de Euglossini (Hymenoptera, Apidae) e preferências por iscas-odores em um fragmento de floresta no sudeste do Brasil. Rev. Bras. Biol. 51, 787–799
Rebelo J.M.M., Moure J.S. (1995) The species of Euglossa Latreille from the northeast of São Paulo, Brazil (Apidae, Euglossinae). Rev. Bras. Zool. 12, 445–466
Rocha N.M.W.B., Carstensen D.W., Fernandes G.W., Le Stradic S., Buisson E., Morellato L.P.C. (2016) Phenology Patterns across a Rupestrian Grassland Altitudinal Gradient. In Fernandes G.W. (Ed.), Ecology and Conservation of Mountaintop Grasslands in Brazil (pp. 275–289). Springer International Publishing, Switzerland
Rocha-Filho L.C., Garófalo C.A. (2013) Phenological patterns and preferences for aromatic compositions by male Euglossini Bees (Hymenoptera, Apidae) in two coastal ecosystems of the Brazilian Atlantic Forest. Neotrop. Entomol. 43, 9–20
Roubik D.W. (2012) Can extinction risk help explain plant-pollinator specificity among euglossine bee pollinated plants? Oikos 121, 1821–1827
Roubik D.W., Ackerman J.D. (1987) Long-term ecology of euglossine orchid-bees (Apidae: Euglossini) in Panama. Oecologia 73, 321. https://doi.org/10.1007/BF00385247
Roubik D.W., Hanson P.E. (2004) Orchid bees of tropical America: Biology and field guide. Editorial INBIO, San José
Schaefer C.E., Cândido H.G., Corrêa G.R., Nunes J.A., Arruda D.M. (2016) Soils associated with rupestrian grasslands. In Fernandes G.W. (Ed.), Ecology and Conservation of mountain-top grasslands in Brazil (pp. 55–69). Springer International, Switzerland
Silveira F.A., Cure J.R. (1993) High altitude bee fauna of Southeastern Brazil: Implications for biogeographic patterns. (Hymenoptera: Apoidea). Stud. Neotrop. Fauna Environ. 28, 47–55
Silveira F.A., Melo G.A.R., Almeida E.A.B. (2002) Abelhas brasileiras -sistemática e identificação. Edição dos Autores, Belo Horizonte
Silveira G.C., Freitas R.F., Tosta T.H.A., Rabelo L.S., Gaglianone M.C., Augusto S.C. (2015) The orchid bee fauna in the Brazilian savanna: do forest formations contribute to higher species diversity? Apidologie. 46, 197–208
Silveira F.A.O., Negreiros D., Barbosa N.P.U., Buisson E., Carmo F.F. et al (2016) Ecology and evolution of plant diversity in the endangered campo rupestre: a neglected conservation priority. Plant Soil 403, 129–152
Sundqvist M.K., Sanders N.J., Wardle D.A. (2013) Community and ecosystem responses to elevational gradients: processes, mechanisms, and insights for global change. Annu. Ver. Ecol. Evol. Syst. 44, 261–280
Viotti M.A., Moura F.R., Lourenço A.P. (2013) Species diversity and temporal variation of the orchid-bee fauna (Hymenoptera, Apidae) in a conservation gradient of a rocky field area in the Espinhaço Range, state of Minas Gerais, southeastern Brazil. Neotrop. Entomol. 42, 565–575
Wikelski M., Moxley J., Eaton-Mordas A., López-Uribe M.M., Holland R. et al (2010) Large-Range Movements of Neotropical Orchid Bees Observed via Radio Telemetry. PLoS One 5, e10738
Williams N.H. (1982) The biology of orchids and Euglossine bees. In Arditti J. (Ed.), Orchid Biology: reviews and perspectives (pp. 120–171). Cornell University Press, Ithaca
Williams N.H., Whitten W.M. (1983) Orchid floral fragrances and males Euglossini bees: Methods and advances in the last sesquidecade. Biol. Bull. 164, 355–395
Winfree R., Griswold I.T, Kremen C. (2007) Effect of Human Disturbance on Bee Communities in a Forested Ecosystem. Conserv. Biol. 21, 213–223
Zanella F.C.V. (2000) The bees of the Caatinga (Hymenoptera, Apoidea, Apiformes): a species list and comparative notes regarding their distribution. Apidologie 31, 579–592
Acknowledgments
We thank Graziella França Monteiro for field assistance and Rodrigo Assunção Silveira for help with orchid bees’ identification. We also thank two anonymous reviewers for their comments on earlier versions of this manuscript.
Funding
CNPq provided scholarship to GWF and YA and research grant (PELD-CRSC). This research was also supported by CAPES (scholarship to FMS), PAPIIT grants to SMR and MQ (UNAM). Programa Iberoamericano de Ciencia y Tecnología para el Desarrollo RED CYTED-SEPODI (417RT0527).
Author information
Authors and Affiliations
Contributions
FMS, WB, YA, and GWF: design and interpretation of the data; FMS performed experiments and analysis; all authors wrote the paper and participated in the revisions of it. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Manuscript Editor: Alexandra Klein
Structure et composition de la communauté d'abeilles euglossines le long d'un gradient d'altitude des prairies du Campo rupestre.
abeilles à orchidée / Campo rupestre / Cerrado brésilien / écologie de montagne / savane néotropicale.
Struktur und Zusammensetzung der Euglossinen-Bienengemeinschaft entlang eines Höhengradienten in einer Grasland-Vegetation des Campo rupestre.
Prachtbienen / Campo rupestre / brasilianische Cerrado / Gebirgsökologie / neotropische Savanne.
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
dos Santos, F.M., Beiroz, W., Antonini, Y. et al. Structure and composition of the euglossine bee community along an elevational gradient of rupestrian grassland vegetation. Apidologie 51, 675–687 (2020). https://doi.org/10.1007/s13592-020-00752-7
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13592-020-00752-7