
Abstract
Clinical manifestations of human coronavirus (HCoV)-related diseases are mostly related to the respiratory system, although secondary complications such as headache, anosmia, ageusia, and myalgia have been reported. HCoV infection and replication in chemosensory cells associated with ageusia and anosmia is poorly understood. Here, we characterized HCoV-OC43 and SARS-CoV-2 infection in two types of chemosensory cells, olfactory and taste cells, with their unique molecular and histological characteristics. We first assessed HCoV-OC43 infection in in vitro cultured human olfactory epithelial cells (hOECs) and fungiform taste papilla (HBO) cells. Interestingly, while both cell types were susceptible to HCoV-OC43 infection, viral replication rates were significantly reduced in HBO cells compared to hOECs. More interestingly, while culture media from hOECs was able to produce secondary infection in Vero cells, there was very limited secondary infection from HBO cells, suggesting that HBO cells may not be able to release infectious virus. On the other hand, unlike HCoV-OC43, SARS-CoV-2 showed comparable levels of viral infection rates in both hOECs and HBO cells. Furthermore, our RT-qPCR-based gene array studies revealed that several key genes involved in taste and olfactory functions were significantly altered by SARS-CoV-2 infection. These results may suggest a possible mechanism associated with chemosensory symptoms, such as anosmia and ageusia in patients infected with SARS-CoV-2.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
In late December 2019, a new outbreak of pneumonia, later associated to a new strain of coronavirus, started in Wuhan, China (Zhou et al. 2020). The virus was named SARS-CoV-2 from the International Committee on Taxonomy of Viruses (Coronaviridae Study Group of the International Committee on Taxonomy of Viruses 2020). SARS-CoV-2 outbreak spreads from China to other countries worldwide, first in Europe and Iran, then Americas and Asia (https://coronavirus.jhu.edu/data/animated-world-map). On March 11, 2020, the World Health Organization (WHO) has declared the outbreak a global pandemic (Cucinotta and Vanelli 2020). Coronaviruses (CoVs) are positive-sense-single-strand RNA viruses, enveloped, belonging to the Coronaviridae family, sub-family Orthocoronavirinae. CoVs can infect humans, causing intestinal and respiratory infections. Before SARS-CoV-2, there were six coronaviruses known to infect humans. The first human coronaviruses (HCoVs) were identified in 1965 and 1966, and they were named HCoV-OC43 and HCoV-229E (Tyrrell and Bynoe 1965; Hamre and Procknow 1966). They were shown to cause common cold and, in rare cases, infection of the respiratory tract (Bradburne et al. 1967). SARS-CoV-2 is the seventh coronavirus associated with humans (Zhou et al. 2020). SARS-CoV-2-associated acute respiratory disease has been defined as coronavirus disease 19 (COVID-19) (Coronaviridae Study Group of the International Committee on Taxonomy of Viruses 2020). Major symptoms of COVID-19 are high fever, dry cough, myalgia, and dyspnea, but patients may experience a broad spectrum of minor symptoms. The majority of patients with COVID-19 experience mild-to-moderate respiratory symptoms, although 5 to 10% can progress to severe disease, such as pneumonia and acute respiratory failure that can lead to persistent interstitial lung disease (Raghu and Wilson 2020; Konopka et al. 2021; Mehta et al. 2022). COVID-19 minor symptoms include disfunction in taste and smell, ranging from mild disfunction to complete loss, sputum production, diarrhea, headache, and lymphopenia (Tian et al. 2020; Huang et al. 2020; Ryu and Chun 2020; Carlos et al. 2020; Guan et al. 2020; Lai et al. 2020; Wang et al. 2020; Ren et al. 2020; Rothan and Byrareddy 2020; Xu et al. 2022). There is limited understanding of taste and smell disturbances in COVID-19 patients. Here, we investigated and compared human coronavirus HCoV-OC43 and SARS-CoV-2 infection kinetics in chemosensory taste and olfactory cells. Our results suggest that both taste and olfactory cells are susceptible to infection with SARS-CoV-2, and infection alters cellular gene expression associated with chemosensory functions.
Materials and methods
Cell culture and reagents
hOECs, primary human olfactory epithelium cells, are provided by Dr. Chang Kyu Khan of the University Pennsylvania and described previously (Ozdener et al. 2020). HBO cells were derived from two male and two female volunteers and were developed in Dr. Ozdener’s Lab at Monell Chemical Senses Center (Ozdener and Rawson 2011; Ozdener et al. 2011, 2012). HBO cells were maintained in Iscove’s modified Dulbecco’s medium with 10% fetal bovine serum,1:5 ratio of MCDB 153 medium, 100 U/mL penicillin, 100 μg/mL streptomycin, and 0.5 μg/mL fungizone. hOECs were maintained in Iscove’s culture medium with 10% FBS and 100 U/mL penicillin/streptomycin. Vero cells were obtained from the American Type Cell Culture (ATCC) and maintained in Dulbecco’s Modified Eagle Medium, with 10% FBS and 100 U/mL penicillin/streptomycin. Cells were maintained at 37 °C in a humidified atmosphere with 5% CO2.
HCoV-OC43 and SARS-CoV-2 propagation and titration
HCoV-OC43 was obtained from ATCC (VR-1558™) and propagated in Vero cells for 6 days. Supernatants were then collected, clarified by centrifugation at 3000 rpm for 5 min, and freeze. RNA from supernatant was extracted using an RNA extraction kit (PureLink™ RNA Mini Kit, Invitrogen) following manufacturer’s specification, and the titer was calculated by RT-qPCR (RdRp). SARS-CoV-2 isolate was deposited by the Centers for Disease Control and Prevention and obtained through BEI Resources, NIAID, NIH: SARS-Related Coronavirus 2, Isolate USA-WA1/2020, NR-52281. It was propagated in Vero cells in BSL3 settings and titrated by RT-qPCRs (RdRp gene).
HCoV-OC43 and SARS-CoV-2 infection and replication
HBO, hOECs, and Vero cells were seeded in 60-mm dishes at concentration of 5 × 105 cells/dish. Twenty-four hours after seeding, cells were infected with HCoV-OC43 at different MOI (1, 10, 25, and 50 MOI) in 1 mL Opti-MEM® containing diluted virus. Five days after infection, culture media and cellular RNA were collected and processed for further analysis. To investigate kinetics of HCoV-OC43 infection, HBO, hOECs, and Vero cells were seeded in 6-well plate at concentration of 2.5 × 105 cells/well and infected with HCoV-OC43 at 25 MOI in 750 μL Opti-MEM® containing diluted virus. Culture media was collected daily up to 7 days, and cellular RNA was collected at 7 dpi. SARS-CoV-2 infection in HBO, hOECs, and Vero cells was performed in T75 flasks. Cells were infected with SARS-CoV-2 at MOI of 100. At 4 dpi, culture media and cellular RNA were collected. RNA from culture media and cells were purified using RNA extraction kit (PureLink™ RNA Mini Kit, Invitrogen) following manufacturer’s specification and analyzed by RT-qPCR for viral RNA copies.
Real-time RT-qPCR
HBO, hOEC, and VERO cells were infected either with HCoV-OC43 or SARS-CoV-2, and RNA from culture media or cell pellet was extracted using an RNA extraction kit (PureLink™ RNA Mini Kit, Invitrogen) following manufacturer’s specification. RT-qPCR assays were performed using Luna® Universal One-Step RT-qPCR Kit (New England Biolabs) in a LightCycler 96 instrument (Roche, Indianapolis, IN, USA). The reaction mixtures contained: 1 × Luna Universal One-Step Reaction Mix, 1 × Luna WarmStart® RT Enzyme Mix, 0.4 μM of forward primer, 0.4 μM of reverse primer, template RNA, and nuclease-free water to a final volume of 20 μL. The protocol was reverse transcription at 55 °C for 10 min, initial denaturation at 95 °C for 60 s, followed by 40 cycles of denaturation at 95 °C for 10 s, and extensions at 60 °C for 60 s, with single acquisition. The amplification steps were then followed by melting steps: initial denaturation at 95 °C for 15 s, followed by 60 s at 60 °C, and temperature increase with continuous readings for 15 s to reach 95 °C. The primers used are the following: HCoV-OC43 Forward: 5′-TACGAGTGTAGATGCCCGTC-3′ and HCoV-OC43 Reverse: 5′-CGCTCATAGCATTTCATCTC-3′, SARS-CoV-2 Forward: 5′-CACGCTGCTTCTGGTAATC-3′ and SARS-CoV-2 Reverse: 5′-CAGCATTACCATCCTGAGC-3′. pCR™2.1-HCoV-OC43-RdRp and pMAL-c5X-SARS-CoV-2-RdRp plasmids were used to create a standard curve for HCoV-OC43 and SARS-CoV-2, respectively, to calculate exact copies of virus in 1 mL culture media or 1 ug cellular RNA.
Plasmid constructs
Portion of HCoV-OC43-RdRp was TA cloned using primers from RT-qPCR. Primers were used in PCR to amplify portion of HCoV-OC43 RdRp protein (NCBI Ref: AY391777.1). Amplicon was then gel purified and ligated into pCR™2.1. Plasmid was labeled as pCR™2.1-HCoV-OC43-RdRp. SARS-CoV-2 RdRp coding sequence (NCBI Ref: NC_045512.2) was cloned into pMAL-c5X plasmid at NotI/BamHI restriction enzyme sites and labeled as pMAL-c5X-SARS-CoV-2-RdRp. The plasmid was created with the following primers: Forward: 5′-GCGGCCGCTCAGCTGAT-3′ and Reverse: 5′-GGATCCCTGTAAGACTGTA-3′.
Plaque-forming assay
Plaque-forming assay was performed following a protocol described previously (Baer and Kehn-Hall 2014). Briefly, Vero cells were seeded in 12-well-plate at concentration of 2.5 × 105 cells/well. Culture media from HBO, hOECs, and Vero cells infected with HCoV-OC43 at 25 MOI were used for infection of Vero cells. Serial dilutions of culture medium were prepared by diluting 100 μL of culture medium in 900 μL Opti-MEM® from 10−1 to 10−5. 350 μL of each viral dilution was added to confluent Vero cells and incubated at 37 °C overnight. The following day, viral dilutions were removed, and wells were washed with PBS and covered with a 1:1 mixture of 2X culture media and 0.6% agarose. At day 8 post-infections, cells were fixed with a solution of 10% formaldehyde for 90 min and then stained with crystal violet solution at 1% (CV) in 20% ethanol and dH2O. HCoV-OC43 titers were calculated by counting the plaque-forming units (PFU).
Immunocytochemistry of cultured human fungiform taste and cultured human olfactory cells
HBO cells were seeded on coverslip and then fixed with 4% PFA for 10 min at room temperature followed by blocking for 1 h at room temperature. After blocking, HBO cells were exposed to primary antibodies (Table 1). After overnight incubation of primary antibodies at 4 °C, HBO cells were washed three times with 1 × PBS for 10 min and incubated with secondary antibody (Table 1) diluted in blocking solution for 30 min at room temperature. Coverslips were washed with PBS 3 × 15 min, followed by Millipore water 3 × 20 min, and then mounted using VectaShield reagent with DAPI (Vector labs, H1200-10) and kept at 4 °C until visualization. Images were acquired with a Leica TCS SP2 spectral confocal microscope. The excitation wavelength used was 633 nm for Alexa Fluor 633, with emissions detected at appropriate wavelengths. Leica Scanware software was used to acquire confocal images by scanning unidirectionally at a 1024 × 1024 pixel format with 3-line and 2-frame averaging. A computer-controlled digital zoom was used to increase magnification to a maximum of 2.3 × with the 20 × and 40 × objective. Images were arranged and minimally adjusted for contrast and brightness using LCS software (Leica Microsystems) and Adobe Photoshop Elements 2.0 (Adobe Systems).
Chemosensory gene arrays by quantitative reverse transcription-PCR (RT-PCR)
Gene expression services were provided by the UPENN Molecular Profiling Facility, including RNA isolation with the microRNeasy kit with DNase treatment (Qiagen) and quality control of the total RNA samples by Agilent Bioanalyzer and Nanodrop spectrophotometry. The expression was quantified in quadruplicate by RT-qPCR using TaqMan Assays (Applied Biosystems by Life Technologies, Foster City, CA, USA). The reverse transcription reaction was carried out with High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems) in 100 μL containing 2.0 μg RNA in 30.0 μL Nuclease Free Water, 4.0 μL of 25X (100 mM) dNTPs, 5 μL of Multiscribe Reverse Transcriptase (50 U/μL), 10.0 μL of 10X Reverse Transcription Buffer, 10.0 μL 10X Random Primer. For synthesis of cDNA, the reaction mixtures were incubated at 25 °C for 10 min, at 37 °C for 120 min, at 85 °C for 5 min and then held at 4 °C. Then, 4.5 μL of cDNA diluted 1:5 was amplified using 5.0 μL TaqMan 2 × Fast Universal PCR Master Mix with no AmpErase UNG (Applied Biosystems) and 0.5 μL of assay in a final volume of 10.0 μL. TaqMan probes for GAPDH (glyceraldehyde-3-phosphate dehydrogenase Hs02758991_g1), GNAL (Hs00181836_m1), OR51E2 (Hs00258239_s1), OMP (Hs01087269_s1), TAS2R46 (Hs00853124_s1), TAS2R14 (Hs00256800_s1), TAS2R20 (Hs00604340_s1), TAAR1 (Hs00373229_s1), OR8H1 (Hs01943789_s1), SCNN1A (Hs00168906_m1), SCNN1D (Hs00161595_m1), OR8K3 (Hs01943810_s1), PLCb2 (Hs01080541_m1), ACE2 (Hs01085333_m), TRPM5 (Hs05060590_s1), ADCY3 (Hs01086502_m1), OR5K1 (Hs07290899_s1), OR7D4 (Hs02339188_s1), TAS2R5 (Hs01549633_s1), OR5P3 (Hs01029920_s1), OR51E1 (Hs02339849_s1), OR2W1 (Hs01121978_s1), OR5A1 (Hs04980963_s1), T1R3 (Hs01387770_g1), and T2R38 (Hs00604294_s1) were ordered from Applied Biosystems (ThermoFisher). Quantitative PCR was run on a QuantStudio 12 K Flex Real-Time PCR system (Applied Biosystems), and the reaction mixtures were incubated at 95 °C for 10 min, followed by 40 cycles of 95 °C for 15 s and 60 °C for 1 min. The cycle threshold (Ct) values were calculated with QuantStudio Software version 1.2.4 (Applied Biosystems). Mean values of Ct triplets were calculated (single values differing ≥ 1 Ct were excluded), and ΔCt values were determined using mean Ct of GAPDH as reference (∆Ct = Ct target − Ct reference). Finally, 2−∆∆Ct values were calculated based on established methods (Livak and Schmittgen 2001).
Results
Characterization of cultured human taste and olfactory cells
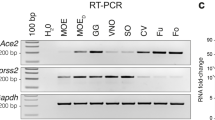
In mammals, taste papillae cells are morphologically and functionally distinct cells that detect a wide range of taste molecules. The taste papillae cells express several signaling proteins including the G-protein alpha-gustducin, phospholipase C-β2 (PLC-β2), which have been implicated in transduction of sweet, umami, and bitter tastes responses (Gilbertson et al. 2000; Pérez et al. 2002). Earlier, we demonstrated the expression of functional sweet (T1Rs) and bitter taste (T2Rs) receptors in a subset of taste type II cells (Zhao et al. 2003; Chandrashekar et al. 2006; Ozdener et al. 2011). Cultured human fungiform taste papilla (HBO) cells were examined for the presence of specific taste cell markers using standard immunocytochemical techniques. Staining of HBO cells with PLC-β2, gustducin, T1R2 (sweet receptor), and T2R38 (bitter receptor) antibodies indicated presence of these proteins (Fig. 1A). Approximately 25% of HBO cells expressed PLC-β2 and about 50% expressed gustducin, which are taste signaling molecules found in type II taste cells (Fig. 1A). We also observed immunoreactivity to T1R2 and T2R38 antibodies, each in about 5–8% of the cells. No reactivity was observed when primary antibodies were substituted with immunoglobulin generated from the same animal (data not shown). Human olfactory tissue is a specialized neuroepithelium in the middle and superior turbinate of the nasal cavity. Previously reports demonstrated the establishment of primary culture of human olfactory sensory neurons (OSNs) in vitro (Gomez et al. 2000b; Lane et al. 2002; Borgmann-Winter et al. 2009). A number of olfactory signal transduction and neuro-olfactory markers were identified in cultured human olfactory epithelial cells. The keys among these are olfactory marker protein, neuron specific tubulin, cyclic nucleotide gated channel subunit alpha 2 (CNGA2), and adenylate cyclase III (ACIII), an element of the olfactory transduction pathway (Rawson et al. 1997; Gomez et al. 2000a). We demonstrated the number of specific indicators of mature olfactory markers in vitro. The cultured human olfactory epithelial cells demonstrated the expression of adenylate cyclase 3 (AC3, ADCY3), key signal transduction molecule, and olfactory marker protein (OMP), a specific marker for mature olfactory receptor neurons. Neuron-specific tubulin (b-tubulin III) and CNGA2 molecules were analyzed by ICC staining, and their expressions were found in cultured human olfactory epithelial cells (Fig. 1B). More interestingly, expression of ACE2, the main receptor for SARS-CoV-2, was also analyzed in RNA samples obtained from both HBO cells and hOECs by RT-qPCR. As shown in Fig. 1C, relatively same levels of ACE2 mRNA were expressed by both HBO cells and hOECs.

Immunostaining of HBO cells and hOECs. A, B Both HBO cells and hOECs showed presence of HBO cell-specific (PLC-b2, T1R2, T2R38, and gustducin, A) and hOEC-specific biomarkers (Adcy3, OMP, CNGA2, and b-tubulin III, B). Images were acquired with a Leica TCS-SP2 confocal laser scanning microscope. Transmission images of corresponding fields are shown on the left. Nuclei of cells were stained as blue with DAPI. For controls, immunostaining with antibody specific immunoglobulin demonstrated the absence of nonspecific immunoreactivity (data not shown). Scale bars = 25 μm. C mRNA level of ACE2 in HBO cells and hOECs was processed by RT-qPCR. Data are shown as delta Ct (ΔCt) values of ACE2 gene versus GAPDH gene
HCoV-OC43 infection and replication in chemosensory cells
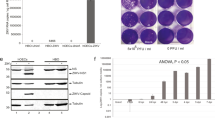
In order to characterize coronavirus infection and replication in chemosensory cells, Vero cells, human olfactory epithelial cells (hOECs), and human taste (HBO) cells were first infected with HCoV-OC43, a member of the Betacoronavirus family. Cells were first infected at different MOIs (1, 10, 25, and 50 MOI), and cellular RNA and culture media were collected and processed for RT-qPCR at 5 dpi. As expected, culture medium was tested positive for HCoV-OC43 RNA reaching a plateau with a viral load of ~ 1010 HCoV-OC43 RNA copies/mL culture medium (Fig. 2A). On the other hand, HBO culture medium viral loads were determined as ~ 2 × 105–3 × 105 HCoV-OC43 RNA copies/mL culture medium (Fig. 2B). Interestingly, hOECs showed a significantly higher viral loads with HCoV-OC43 infection (~ 106 to ~ 5 × 107 RNA copies/mL culture medium) compared to the HBO cells (Fig. 2C). Viral infection kinetic analysis of HCoV-OC43 infection in the chemosensory cells was also performed. Vero cells, HBO cells, and hOECs were infected with HCoV-OC43 at 25 MOI, and culture media were collected daily for 7 days. HCoV-OC43 RNA was extracted from culture media and analyzed for RT-qPCR. As shown in Fig. 2, panels D, E, and F, at day 1 post-infection, culture media from Vero cells, HBO, and hOECs were already tested positive, with a viral load of ~ 104 RNA copies/mL culture medium. Interestingly, Vero cells and hOECs viral load quickly reached a plateau in the 109 and 108 copies/mL culture media, respectively, at day 3, while HBO cells showed a more linear viral kinetics which reached to a plateau at day 6 post-infections with a viral load of ~ 2 × 106 RNA copies/mL culture medium. To further analyze viral replication, total RNA was extracted and processed for RT-qPCR at 7 dpi. In agreement with viral loads in culture media, Vero cells and hOECs showed higher viral RNA replication than HBO cells (Fig. 2G). Next, the infectivity of HCoV-OC43 viral particles released from infected Vero cells, HBO cells, and hOECs was determined by plaque-forming assays. Vero cells were plated in 6-well plate, and culture media from Vero cells, HBO cells, and hOECs (at 7 dpi) were used for infecting Vero cells for plaque-forming assays as described in the “Materials and methods” section. Vero cells were fixed and stained with crystal violet solution at 8 dpi, and the HCoV-OC43 titers were calculated. Interestingly, while culture media from Vero cells and hOECs were able to infect, replicate, and form plaques in Vero cells, HBO cells mainly failed to form any plaques (Fig. 2H). These results suggest that while olfactory cells are susceptible to productive and lytic infection with HCoV-OC43, taste cells are primary nonproductive and possibly infected abortively.

HCoV-OC43 infection in VERO cells, primary human taste (HBO) cells, and olfactory cells (hOECs). A–C VERO, HBO, and hOECs were infected with HCoV-OC43 at different MOI (1, 10, 25, and 50 MOI); RNA and media were collected. A HCoV-OC43-infected VERO cell culture media were processed for RT-qPCR. B HCoV-OC43-infected HBO cell culture media were processed for RT-qPCR. C HCoV-OC43-infected hOEC culture media were processed for RT-qPCR. D–H Vero cells, HBO cells, and hOECs were infected with HCoV-OC43 at 25 MOI; culture media were collected daily, and cellular RNA was extracted at 7 dpi. D HCoV-OC43-infected VERO cell culture media were processed for RT-qPCR. E OC43-infected HBO cell culture media were processed for RT-qPCR. F OC43-infected hOEC culture media were processed for RT-qPCR. G HCoV-OC43-infected Vero cell, HBO cell, and hOEC cellular RNAs were processed for RT-qPCR and shown as bar graph. H Quantification of number of plaques forming units/mL. Data are shown as mean EM of two independent replicates
SARS-CoV-2 infection and replication in chemosensory cells
Next, we also investigated SARS-CoV-2 infection in taste and olfactory cells. Vero, hOEC, and HBO cells were infected with SARS-CoV-2 as described in the “Materials and methods” section. Cellular RNA and culture media were collected and processed for RT-qPCR at 4 dpi. SARS-CoV-2 was capable of infecting both HBO cells and hOECs, although viral copies released from cells were about two logs lower than the Vero cells (Fig. 3A–C). Culture media from Vero cells were tested positive for SARS-CoV-2 RNA, with a viral count of 1.3 × 1010 RNA copies/mL culture medium, while media from HBO cells and hOECs were both tested positive with a viral count of ~ 2 × 108 RNA copies/mL culture medium. Moreover, cellular RNA was extracted from infected cells and processed also for RT-qPCR. As expected, SARS-CoV-2 RNA was in Vero cells with a viral count of ~ 2 × 1010 RNA copies/μg cellular RNA. Interestingly, in HBO cellular RNA, we found significantly more SARS-CoV-2 RNA copies than in hOECs, with a viral count of ~ 6 × 106 RNA copies/μg cellular RNA in HBO, and ~ 9 × 105 RNA copies/μg in hOEC cellular RNA. These results suggest that both HBO cells and hOECs are susceptible to SARS-CoV-2 infection. In addition, although viral loads were comparable in culture media, SARS-CoV-2 replication might be more efficient in HBO cells compared to the hOECs.

SARS-CoV-2 infection in primary human taste (HBO) cells, olfactory cells (hOECs), and Vero cells. HBO cells, hOECs, and Vero cells were infected with SARS-CoV-2, and cellular RNA along with culture media was collected. A SARS-CoV-2-infected hOEC culture media were processed for RT-qPCR. B SARS-CoV-2-infected HBO cell culture media were processed for RT-qPCR. C SARS-CoV-2-infected VERO cell culture media were processed for RT-qPCR. D SARS-CoV-2-infected hOEC cellular RNA was processed for RT-qPCR. E SARS-CoV-2-infected HBO cellular RNA was processed for RT-qPCR. F SARS-CoV-2-infected VERO cellular RNA was processed for RT-qPCR
SARS-CoV-2 modulates chemosensory gene expression in taste and olfactory cells
In order to determine the effect of SARS-CoV-2 infection on functional gene expression, total RNA was extracted from uninfected, and SARS-CoV-2-infected HBO cells and hOECs. Further, the expression of selected target genes in the uninfected vs. infected cells was measured by RT-qPCR (Fig. 4). In HBO cells, the PLC-β2 and TAS2R14 gene transcripts were found significantly upregulated, whereas SCNN1A, TAS220, and TAS38 mRNA expressions were found significantly reduced. Interestingly, ACE2, TRPM5, SCNN1D, TAS1R3, TAS2R46, and TAS2R5 receptor mRNA levels were found unchanged in infected HBO cells compared to uninfected HBO cells (Fig. 4A). The expression of mRNAs of ADCY3, GNAL, OMP, OR2W1, OR5A1 OR8H1, OR5P3, OR5K1, and OR8K3 were also found significantly upregulated in SARS-CoV-2-infected human-cultured olfactory epithelium cells compared to uninfected cells; however, the expressions of OR51E1, OR7D4, and TAAR1 mRNAs were found significantly downregulated, though ACE2, TRPM5, and OR51E2 mRNA amounts were not changed (Fig. 4B).

Relative quantification of taste and olfactory receptors mRNA expression measured by real-time [quantitative] PCR (RT-PCR) in SARS-CoV-2 infected vs. uninfected HBO cells (A) and hOECs (B). Normalization factors were calculated as the geometric mean of the expression levels of the GAPDH reference gene. Values are expressed as group mean; fold change 2−∆∆Ct vs. GAPDH gene, stars indicate a significant difference (p < 0.05) between infected (n = 3) and uninfected (n = 3) independent samples. SARS-CoV-2 infection of taste and olfactory cells resulted to changes of expression level of candidate receptors
Discussion
Although SARS-CoV-2 selectively infects cells (< 1%) in the human body, it causes many clinical and pathological symptoms and damages in a number of organs. Therefore, understanding the cellular pathophysiology of SARS-CoV-2 infection is critically necessary. Chemosensory-deficient symptoms are one of the most common symptoms but again one of the least understood symptoms of COVID-19 patients (Zazhytska et al. 2022). Although many recent studies have reported impairment in taste and smell in COVID-19 patients (Ralli et al. 2020; Hopkins et al. 2020; Menni et al. 2020; Giacomelli et al. 2020; Parma et al. 2020), limited evidence has been reported on the molecular mechanisms underlying these impairments. Many hypotheses were made for possible reasons behind these chemosensory dysfunctions seen in affected individuals with limited supportive data. Hypotheses of anosmia involve loss of olfactory sensory neurons and damage in the olfactory sustentacular cells, most likely due to the inflammatory response to the infection (Han et al. 2020). Our results suggest that human olfactory cells are susceptible to the productive infection with both hCOV-OC43 and SARS-CoV-2 viruses. Similar to anosmia, mechanisms involved in ageusia have not been clarified yet. Main hypothesis also involves the destruction of gustatory receptor cells by cytotoxic inflammatory cytokines and chemokines during the disease (Srinivasan 2021). Interestingly, coronavirus infections in taste cells have not been deeply investigated, although angiotensin-converting enzyme 2 (ACE2) has been found in type II taste cells (Doyle et al. 2021). ACE2 is the primary SARS-CoV-2 receptor (Hoffmann et al. 2020b), and it has been reported to be expressed in different type of cells, where SARS-CoV-2 infection has been demonstrated (Monteil et al. 2020; Hoffmann et al. 2020a; Lamers et al. 2020). We also report that ACE2 mRNA is expressed in both taste and olfactory cells in our model of chemosensory cells, HBO, and hOECs, respectively. Furthermore, we demonstrate that taste cells are also susceptible to infection with both hCOV-OC43 and SARS-CoV-2. Interestingly, hCOV-OC43 infection rates were significantly low in HBO cells and virions released from these cells were not capable of infecting Vero cells. Viral RNA loads in culture media suggest that HBO cells are able to release virus but most likely these virions are defective in infecting new cells. Actual mechanisms of defective hCOV-OC43 replication in taste cells and its cellular and molecular determinants remain to be determined. On the other hand, SARS-CoV-2 infection in taste cells resulted in comparable viral loads in culture media but significantly higher replication rates in these cells than olfactory cells, suggesting a possible mechanism for observed ageusia in infected individuals. ACE2 mRNA levels were either unchanged or upregulated in our cellular models. Our results are in line with previous publications, where increase in ACE2 was demonstrated in endothelial cells and in lungs from COVID-19 infected patients (Garvin et al. 2020; Ackermann et al. 2020; Gerard et al. 2021).
During SARS-CoV-2 infection, temporary (short-term) and persistent (long-term) altered olfactory functions were observed but its etiology is still unknown for both conditions. It has been reported earlier that many viruses including coronaviruses may cause transient loss/changes in olfactory function due to mainly inflammatory responses. Injury to peripheral and central nervous system including olfactory cells is caused from massive activation of cytokines (Doty 2022). Typically, recovery of olfactory function is shorter in SARS-CoV-2 infection compared to non-SARS-CoV-2 post-viral infection which could be due to inflammation in the olfactory epithelium or damage to the olfactory receptor neurons. Persistent loss of olfactory function is caused by the presence of SARS-CoV-2 transcripts in olfactory neuronal and surrounding cells. SARS-CoV-2 infection of non-neuronal cell types leads to olfactory dysfunction in COVID-19 patients (Brann et al. 2020). We, here, report significant downregulation of olfactory receptor (OR) genes and the key genes of the OR signaling pathway.
Unlike olfactory deficiency, changes in taste sensory has drawn less interest in the public and in the scientific community. Many studies reported viruses can infect olfactory cells but infection of the taste cells has been mostly overlooked. Previous studies demonstrated that rabies virus infects taste bud cells in dogs and as well as in human (Li et al. 1994; Shiwa et al. 2018). However, mechanism of this infection was not reported. Possible explanation could be that the virus may reach the taste buds by retrograde transport from the infected brain. Thus, an infection of taste buds which do not contain neurons may not be threatening for the infected individual. Similarly, an explanation of the SARS-CoV-2-induced ageusia (taste loss) could be that taste nerves might be damaged following central nervous infection by SARS-CoV-2 (Matschke et al. 2020). Another possible explanation of taste impairment could be due to cytokine storms caused by SARS-CoV-2 in taste cells as well as in surrounding tongue epithelial cells. It is highly possible that cytokine storm could make taste cells be permissive to SARS-CoV-2. ACE2 has been shown to be overexpressed in the presence of IFNs (Ziegler et al. 2020). Thus, a distant production of IFN from infected keratinocytes could lead to ACE2 expression in taste bud cells which could in turn be infected by SARS-CoV-2 (Meunier et al. 2021).
Data availability
The data that support the findings of this study are available from the authors upon reasonable request.
References
Ackermann M, Verleden SE, Kuehnel M et al (2020) Pulmonary vascular endothelialitis, thrombosis, and angiogenesis in Covid-19. N Engl J Med 383:120–128. https://doi.org/10.1056/NEJMoa2015432
Baer A, Kehn-Hall K (2014) Viral concentration determination through plaque assays: using traditional and novel overlay systems. JoVE 52065. https://doi.org/10.3791/52065
Borgmann-Winter KE, Rawson NE, Wang H-Y et al (2009) Human olfactory epithelial cells generated in vitro express diverse neuronal characteristics. Neuroscience 158:642–653. https://doi.org/10.1016/j.neuroscience.2008.09.059
Bradburne AF, Bynoe ML, Tyrrell DA (1967) Effects of a “new” human respiratory virus in volunteers. Br Med J 3:767–769. https://doi.org/10.1136/bmj.3.5568.767
Brann DH, Tsukahara T, Weinreb C et al (2020) Non-neuronal expression of SARS-CoV-2 entry genes in the olfactory system suggests mechanisms underlying COVID-19-associated anosmia. Sci Adv 6:eabc5801. https://doi.org/10.1126/sciadv.abc5801
Carlos WG, Dela Cruz CS, Cao B et al (2020) Novel wuhan (2019-nCoV) coronavirus. Am J Respir Crit Care Med 201:P7–P8. https://doi.org/10.1164/rccm.2014P7
Chandrashekar J, Hoon MA, Ryba NJP, Zuker CS (2006) The receptors and cells for mammalian taste. Nature 444:288–294. https://doi.org/10.1038/nature05401
Coronaviridae Study Group of the International Committee on Taxonomy of Viruses (2020) The species severe acute respiratory syndrome-related coronavirus: classifying 2019-nCoV and naming it SARS-CoV-2. Nat Microbiol 5:536–544. https://doi.org/10.1038/s41564-020-0695-z
Cucinotta D, Vanelli M (2020) WHO declares COVID-19 a pandemic. Acta Bio Medica Atenei Parmensis 91:157–160. https://doi.org/10.23750/abm.v91i1.9397
Doty RL (2022) Olfactory dysfunction in COVID-19: pathology and long-term implications for brain health. Trends Mol Med 28:781–794. https://doi.org/10.1016/j.molmed.2022.06.005
Doyle ME, Appleton A, Liu Q-R et al (2021) Human type II taste cells express angiotensin-converting enzyme 2 and are infected by severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2). Am J Pathol 191:1511–1519. https://doi.org/10.1016/j.ajpath.2021.05.010
Garvin MR, Alvarez C, Miller JI et al (2020) A mechanistic model and therapeutic interventions for COVID-19 involving a RAS-mediated bradykinin storm. eLife 9:e59177. https://doi.org/10.7554/eLife.59177
Gerard L, Lecocq M, Bouzin C et al (2021) Increased angiotensin-converting enzyme 2 and loss of alveolar type II cells in COVID-19–related acute respiratory distress syndrome. Am J Respir Crit Care Med 204:1024–1034. https://doi.org/10.1164/rccm.202012-4461OC
Giacomelli A, Pezzati L, Conti F et al (2020) Self-reported olfactory and taste disorders in patients with severe acute respiratory coronavirus 2 infection: a cross-sectional study. Clin Infect Dis 71:889–890. https://doi.org/10.1093/cid/ciaa330
Gilbertson TA, Damak S, Margolskee RF (2000) The molecular physiology of taste transduction. Curr Opin Neurobiol 10:519–527. https://doi.org/10.1016/S0959-4388(00)00118-5
Gomez G, Rawson NE, Cowart B et al (2000a) Modulation of odor-induced increases in [Ca(2+)](i) by inhibitors of protein kinases A and C in rat and human olfactory receptor neurons. Neuroscience 98:181–189. https://doi.org/10.1016/s0306-4522(00)00112-3
Gomez G, Rawson NE, Hahn CG et al (2000b) Characteristics of odorant elicited calcium changes in cultured human olfactory neurons. J Neurosci Res 62:737–749. https://doi.org/10.1002/1097-4547(20001201)62:5%3c737::AID-JNR14%3e3.0.CO;2-A
Guan W, Ni Z, Hu Y et al (2020) Clinical characteristics of coronavirus disease 2019 in China. N Engl J Med NEJMoa2002032. https://doi.org/10.1056/NEJMoa2002032
Hamre D, Procknow JJ (1966) A new virus isolated from the human respiratory tract. Proc Soc Exp Biol Med 121:190–193. https://doi.org/10.3181/00379727-121-30734
Han AY, Mukdad L, Long JL, Lopez IA (2020) Anosmia in COVID-19: mechanisms and significance. Chem Senses 45:423–428. https://doi.org/10.1093/chemse/bjaa040
Hoffmann M, Kleine-Weber H, Pöhlmann S (2020a) A multibasic cleavage site in the spike protein of SARS-CoV-2 is essential for infection of human lung cells. Molecular Cell S1097276520302641. https://doi.org/10.1016/j.molcel.2020.04.022
Hoffmann M, Kleine-Weber H, Schroeder S et al (2020b) SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell. https://doi.org/10.1016/j.cell.2020.02.052
Hopkins C, Surda P, Kumar N (2020) Presentation of new onset anosmia during the COVID-19 pandemic. Rhin 58:295–298. https://doi.org/10.4193/Rhin20.116
Huang C, Wang Y, Li X et al (2020) Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 395:497–506. https://doi.org/10.1016/S0140-6736(20)30183-5
Konopka KE, Perry W, Huang T et al (2021) Usual interstitial pneumonia is the most common finding in surgical lung biopsies from patients with persistent interstitial lung disease following infection with SARS-CoV-2. eClin Med 42:101209. https://doi.org/10.1016/j.eclinm.2021.101209
Lai C-C, Shih T-P, Ko W-C et al (2020) Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) and coronavirus disease-2019 (COVID-19): the epidemic and the challenges. Int J Antimicrob Agents 55:105924. https://doi.org/10.1016/j.ijantimicag.2020.105924
Lamers MM, Beumer J, van der Vaart J et al (2020) SARS-CoV-2 productively infects human gut enterocytes. Science eabc1669. https://doi.org/10.1126/science.abc1669
Lane AP, Gomez G, Dankulich T et al (2002) The superior turbinate as a source of functional human olfactory receptor neurons. Laryngoscope 112:1183–1189. https://doi.org/10.1097/00005537-200207000-00007
Li ZS, Feng ZB, Luo H (1994) Distribution of rabies antigen in human tongue and salivary gland. Zhonghua Bing Li Xue Za Zhi 23:230–231
Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 25:402–408. https://doi.org/10.1006/meth.2001.1262
Matschke J, Lütgehetmann M, Hagel C et al (2020) Neuropathology of patients with COVID-19 in Germany: a post-mortem case series. Lancet Neurol 19:919–929. https://doi.org/10.1016/S1474-4422(20)30308-2
Mehta P, Rosas IO, Singer M (2022) Understanding post-COVID-19 interstitial lung disease (ILD): a new fibroinflammatory disease entity. Intensive Care Med 48:1803–1806. https://doi.org/10.1007/s00134-022-06877-w
Menni C, Valdes AM, Freidin MB et al (2020) Real-time tracking of self-reported symptoms to predict potential COVID-19. Nat Med 26:1037–1040. https://doi.org/10.1038/s41591-020-0916-2
Meunier N, Briand L, Jacquin-Piques A et al (2021) COVID 19-induced smell and taste impairments: putative impact on physiology. Front Physiol 11:625110. https://doi.org/10.3389/fphys.2020.625110
Monteil V, Kwon H, Prado P et al (2020) Inhibition of SARS-CoV-2 infections in engineered human tissues using clinical-grade soluble human ACE2. Cell S0092867420303998. https://doi.org/10.1016/j.cell.2020.04.004
Ozdener H, Spielman AI, Rawson NE (2012) Isolation and culture of human fungiform taste papillae cells. JoVE 3730. https://doi.org/10.3791/3730
Ozdener MH, Brand JG, Spielman AI et al (2011) Characterization of human fungiform papillae cells in culture. Chem Senses 36:601–612. https://doi.org/10.1093/chemse/bjr012
Ozdener MH, Donadoni M, Cicalese S et al (2020) Zika virus infection in chemosensory cells. J Neurovirol 26:371–381. https://doi.org/10.1007/s13365-020-00835-2
Ozdener MH, Rawson NE (2011) Culture and maintenance of taste cells in vitro. In Vitro Cell DevBiol-Animal 47:513–514. https://doi.org/10.1007/s11626-011-9437-0
Parma V, Ohla K, Veldhuizen MG et al (2020) More than smell—COVID-19 is associated with severe impairment of smell, taste, and chemesthesis. Chem Senses 45:609–622. https://doi.org/10.1093/chemse/bjaa041
Pérez CA, Huang L, Rong M et al (2002) A transient receptor potential channel expressed in taste receptor cells. Nat Neurosci 5:1169–1176. https://doi.org/10.1038/nn952
Raghu G, Wilson KC (2020) COVID-19 interstitial pneumonia: monitoring the clinical course in survivors. Lancet Respir Med 8:839–842. https://doi.org/10.1016/S2213-2600(20)30349-0
Ralli M, Di Stadio A, Greco A et al (2020) Defining the burden of olfactory dysfunction in COVID-19 patients. Eu Rev Med Pharmacol Sci 24:3440–3441. https://doi.org/10.26355/eurrev_202004_20797
Rawson NE, Gomez G, Cowart B et al (1997) Selectivity and response characteristics of human olfactory neurons. J Neurophysiol 77:1606–1613. https://doi.org/10.1152/jn.1997.77.3.1606
Ren L-L, Wang Y-M, Wu Z-Q et al (2020) Identification of a novel coronavirus causing severe pneumonia in human: a descriptive study. Chin Med J 133:1015–1024. https://doi.org/10.1097/CM9.0000000000000722
Rothan HA, Byrareddy SN (2020) The epidemiology and pathogenesis of coronavirus disease (COVID-19) outbreak. J Autoimmun 109:102433. https://doi.org/10.1016/j.jaut.2020.102433
Ryu S, Chun BC (2020) An interim review of the epidemiological characteristics of 2019 novel coronavirus. Epidemiol Health 42:e2020006. https://doi.org/10.4178/epih.e2020006
Shiwa N, Kimitsuki K, Manalo DL et al (2018) A pathological study of the tongues of rabid dogs in the Philippines. Arch Virol 163:1615–1621. https://doi.org/10.1007/s00705-018-3785-y
Srinivasan M (2021) Taste dysfunction and long COVID-19. Front Cell Infect Microbiol 11:716563. https://doi.org/10.3389/fcimb.2021.716563
Tian S, Hu N, Lou J et al (2020) Characteristics of COVID-19 infection in Beijing. J Infect 80:401–406. https://doi.org/10.1016/j.jinf.2020.02.018
Tyrrell DA, Bynoe ML (1965) Cultivation of a novel type of common-cold virus in organ cultures. Br Med J 1:1467–1470. https://doi.org/10.1136/bmj.1.5448.1467
Wang W, Tang J, Wei F (2020) Updated understanding of the outbreak of 2019 novel coronavirus (2019-nCoV) in Wuhan, China. J Med Virol 92:441–447. https://doi.org/10.1002/jmv.25689
Xu W, Sunavala-Dossabhoy G, Spielman AI (2022) Chemosensory loss in COVID-19. Oral Dis Odi 14300. https://doi.org/10.1111/odi.14300
Zazhytska M, Kodra A, Hoagland DA et al (2022) Non-cell-autonomous disruption of nuclear architecture as a potential cause of COVID-19-induced anosmia. Cell 185:1052-1064.e12. https://doi.org/10.1016/j.cell.2022.01.024
Zhao GQ, Zhang Y, Hoon MA et al (2003) The receptors for mammalian sweet and umami taste. Cell 115:255–266. https://doi.org/10.1016/s0092-8674(03)00844-4
Zhou P, Yang X-L, Wang X-G et al (2020) A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 579:270–273. https://doi.org/10.1038/s41586-020-2012-7
Ziegler CGK, Allon SJ, Nyquist SK et al (2020) SARS-CoV-2 receptor ACE2 is an interferon-stimulated gene in human airway epithelial cells and is detected in specific cell subsets across tissues. Cell 181:1016-1035.e19. https://doi.org/10.1016/j.cell.2020.04.035
Acknowledgements
The authors wish to thank past and present members of the Department of Neuroscience/Center for Neurovirology and Gene Editing for sharing of reagents and ideas.
Funding
This work was supported, in part, by grant (NIH-R01DC014286-05S1) (PI Margolskee) awarded by the NIH to RM, MHO, and IKS. This study utilized services offered by core facilities of the Comprehensive NeuroHIV Center (CNHC NIMH Grant Number P30MH092177-11). The imaging experiments were performed at the Monell Chemical Senses Center “The Histology and Cellular Localization Core,” which is supported, in part, by funding from the NIH-NIDCD Core Grant P30DC011735.
Author information
Authors and Affiliations
Contributions
Conceived and designed the experiments: MHO and IKS. Performed the experiments: MD, RK, and SL. Analyzed the data: MD, MHO, and IKS. Contributed reagents/materials/analysis tools: RM, MHO, and IKS. Wrote the paper: MD, MHO, and IKS.
Corresponding authors
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Donadoni, M., Kaminski, R., Liao, S. et al. Coronavirus infection in chemosensory cells. J. Neurovirol. 29, 35–44 (2023). https://doi.org/10.1007/s13365-023-01117-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13365-023-01117-3