
Abstract
The Zagros Mountains represent a strategic geographic region near the junction of Africa, Europe, and Asia, and valleys within the mountain range likely correspond to migration corridors used by archaic and modern humans. Ghar-e Boof, a key archeological site in the southern Zagros, is located at an altitude of 905 m.a.s.l. in the Dasht-e Rostam region of the Fars Province, Iran. On the basis of lithic artifacts and radiocarbon and luminescence dating, the Late Pleistocene archeological record of the site spans from the Middle Paleolithic (MP) to the Late Epipaleolithic. Ghar-e Boof includes find horizons of the early Upper Paleolithic (UP) Rostamian dating to 42–35 ka cal. BP. These Rostamian-UP assemblages are typically attributed to modern humans. The high-resolution stratigraphy and microvertebrate record offer a unique opportunity for reconstructing the paleoenvironment of the southern part of the Zagros Mountains during the Late Pleistocene and contribute to a better understanding of long-term hominin settlement dynamics and cultural adaptations in this area. The current paper presents the first analysis of the microvertebrate remains recovered by the Tübingen-Iranian Stone Age Research Project (TISARP) team. All groups of small vertebrates, including fish, amphibians, squamate reptiles, birds, and micromammals, are present in the assemblage from Ghar-e Boof (NISP = 755). The key micromammal taxa used to reconstruct the paleoenvironment during the MP and UP include Libyan jird (Meriones cf. libycus), Vinogradov’s jird (Meriones vinogradovi), Indian gerbil (Tatera indica), Brandt’s hedgehog (Paraechinus cf. hypomelas), house mouse (Mus musculus), and mole vole (Ellobius sp.). Other taxa, such as amphibians (toad, Bufo/Bufotes sp.), fish (nase, Chondrostoma sp.), and squamate reptiles (agamids, lizards, Eastern Montpelier snake—Malpolon insignitus—and viper), also support the presence of an environment mainly characterized by warm, arid conditions with open, dry meadows, rocky terrain, and water sources close to the site throughout most of the sequence. During the shift from MP to UP, we record Afghan pika (Ochotona cf. rufescens), while all rodents, except very few remains of the Persian vole (Microtus cf. irani) and the Libyan jird (M. cf. lybicus) and some non-hibernating vertebrate species, are absent and reappear in later UP horizons. The presence of pikas could reflect a period of slightly colder and/or drier conditions. While the record from Ghar-e Boof is the only dataset from the southern Zagros, previous studies on microvertebrates from the central and northern Zagros show no clear change during the shift from the MP to the UP. Data from sediment cores and pollen analyses document numerous environmental and climatic fluctuations during the Late Pleistocene of the Zagros. The microvertebrate record from Ghar-e Boof improves our understanding of paleoenvironments during the Late Pleistocene and opens new opportunities to reconstruct the ecological conditions that helped to shape hominin settlement dynamics in the region.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The strategic location of the Zagros Mountains, near the intersection between Africa, Europe, and southwestern Asia, most probably represented a major biographical corridor used by archaic and anatomically modern humans (AMHs) during the Late Pleistocene (Field et al. 2006; Oppenheimer 2009; Boivin et al. 2013; Vahdati-Nasab et al. 2013; Shoaee et al. 2021). The anthropological and archeological potential of this geographic area for the understanding of human dispersals and cultural adaptations across Eurasia has been demonstrated with the discovery of Neanderthal and AMH remains associated with lithic industries at several localities in the Zagros (i.e., Solecki 1958; Solecki 1963; Trinkaus 1983, 2018; Trinkaus and Biglari 2006; Cowgill et al. 2007; Trinkaus et al. 2008; Churchill et al. 2009; Scott and Marean 2009; Tsanova 2013; Pomeroy et al. 2017, 2020; Trinkaus and Villotte 2017; Zanolli et al. 2019; Heydari-Guran et al. 2021a). Regardless of the precise timing and nature of the shift from Neanderthals to AMH, when different human groups first spread across Asia, they had to adapt to new and diverse environmental conditions (Boivin et al. 2013).
Local environmental reconstructions are especially relevant to understand the human response to landscape changes and, in particular, the Zagros Mountains contain numerous microenvironments and heterogeneous topography, so both MP and UP hunter-gatherers adopted various cultural adaptations and subsistence strategies in different landscapes and environments (Bretzke and Conard, 2017; Ghasidian et al. 2019; Heydari-Guran and Ghasidian 2020). Consequently, paleoenvironmental reconstructions of the Zagros during the MP and UP provide valuable new information for a better understanding of long-term hominin settlement dynamics and cultural adaptations.
Many environmental reconstructions available for the Zagros Mountains during the Late Pleistocene come from the analyses of sediment cores. Lake Urmia, located in the northern Zagros, has produced a long pollen record spanning 200 kya (Djamali et al. 2008a, 2008b). Moreover, botanists have also analyzed the pollen and seeds from Lake Zeribar, in the Western-Central Iran, with a record that covers the last 42 kya (van Zeist and Bottema 1977; Wasylikowa 2005). These studies have shown that during most of the Late Pleistocene, the environment of the Zagros Mountains was mainly characterized by the presence of shrub/grass steppe vegetation (i.e., Chenopodiaceae, Umbelliferae, Poaceae, and Artemisia sp.) with some scattered trees, such as junipers, deciduous oak, maple, and pistachio (van Zeist and Bottema 1977; Djamali et al. 2008a). Although this vegetation indicates a very arid landscape with colder temperatures than today, the environment fluctuated significantly during the Late Pleistocene between cold, warm, dry, and wet conditions (van Zeist and Bottema 1977; Wasylikowa 2005; Djamali et al. 2008a). Due to the complex topography of the Zagros and their altitudinal variations, the mountains offer a large variety of ecosystems with marked contrasts (Heydari and Mahdavi 2009; Noroozi et al. 2020; Mostafavi et al. 2021). Although the sediment cores probably reflect major environmental changes, they cannot be used for inferring a particular site environment.
It is well-known that small vertebrates represent good paleoenvironmental indicators and are very frequently found well-preserved and in relatively large quantities in archeological and paleontological sites, especially in rock shelters and caves (Andrews 1990; Avery 2007; Fernández-Jalvo et al. 2016). Small vertebrates are often used for inferring and reconstructing environmental conditions because, among other characteristics, they have limited geographic ranges, rapid turn-over rates, and unique habitat requirements, such as specific humidity conditions or levels of vegetation cover (Andrews and O’Brien 2000; Avery 2007; Blain et al. 2009; Belmaker and Hovers 2011).
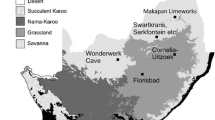
Since the second half of the twentieth century, researchers have studied numerous Late Pleistocene small vertebrate assemblages associated with MP and UP artifacts and human remains in the Zagros Mountains (Fig. 1A), including Shanidar Cave (Perkins 1964; Tilby et al. 2022); Gar Arjeneh, Kunji Cave and Pa-Sangar (Hole and Flannery 1967); Ghamari Cave (Hole and Flannery 1967; Bazgir et al. 2014); Yafteh Cave (Hole and Flannery 1967; Hashemi et al. 2006; Otte et al. 2007; Mashkour et al. 2009a); Palegawra Cave (Turnbull and Reed 1974; Asouti et al. 2020); Warwasi Cave (Turnbull 1975); Wezmeh Cave (Mashkour et al. 2009b); and Kaldar Cave (Bazgir et al. 2014, 2017; Rey-Rodríguez et al. 2020, 2021). However, with the exception of Rey-Rodríguez et al. (2020), who provided a thorough paleoenvironmental and paleoclimatic reconstruction for the MP and UP deposits of Kaldar Cave, these studies mainly consist of preliminary taxonomic lists and the majority of studied taxa correspond only to micromammals. Squamata reptiles, amphibians, birds, and fish have also been recovered at some sites (Turnbull and Reed 1974; Otte et al. 2007; Mashkour et al. 2009b; Bazgir et al. 2017; Asouti et al. 2020). Despite the limited data, small vertebrate assemblages are still the primary source of information for reconstructing the environment in which Neanderthals and AMHs lived during the MP and UP in the Zagros Mountains.

Map created by M. Zeidi. Source: Natural Earth 2. B Location of Ghar-e Boof Cave and its overview during the 2017 campaign (B. Figure created by M. Zeidi. Source: Bing Maps. Ghar-e Boof photo: M. Zeidi). C Ghar-e Boof excavation area divided into quadrants, with the cave entrance to the north (map created by M. Zeidi (modified after Conard and Ghasidian 2011)
A Location of archeological sites mentioned in the text: 1. Shanidar Cave; 2. Palegawra Cave; 3. Wezmeh Cave; 4. Warwasi Cave; 5. Gar Ajeneh; 6. Kaldar Cave, Pa-Sangar, Yafteh Cave; 7. Kunji Cave; 8. Ghamari Cave; 9. Ghar-e Boof.
Archeological sites with available environmental data obtained from the analysis of small vertebrates are mainly located in western/central and the northern Zagros, while little is known about the Late Pleistocene environmental conditions of the southernmost part of the mountain range. Although the southern Zagros is less studied, archeologists have long recognized the importance of this area for human and cultural evolution, with the documentation, for example, of MP and UP sites such as Eshkaft-e Ghadi Barmishur and Eshkaft-e Gavi (Piperno 1972, 1974; Ikeda 1979; Rosenberg 1979, 1985, 1988; Scott and Marean 2009).
Since 2004, the Tübingen-Iranian Stone Age research project (TISARP) team has conducted numerous surveys and excavations in the Fars Province (Iran) in the southern Zagros and documented more than 120 archeological sites (Conard et al. 2006, 2007, Zeidi et al. 2006; Conard and Ghasidian 2011; Heydari-Guran 2014). Among these sites, the most promising one was Ghar-e Boof, with a Late Pleistocene stratigraphic sequence spanning from the MP until the Late Epipaleolithic (Conard and Ghasidian 2011; Baines et al. 2014; Becerra-Valdivia et al. 2017; Conard and Zeidi 2019; Zeidi and Conard 2019; Heydari et al. 2021).
Based on the analysis of different lithic assemblages recovered at Ghar-e Boof, archeologists observed that there is no technological continuity between the MP and the UP, suggesting a population replacement or settlement discontinuity (Bretzke and Conard 2017). Furthermore, Ghar-e Boof is the type locality for the early UP Rostamian cultural group, which is characterized by unidirectional bladelet production and small platform cores (Conard and Ghasidian 2011; Ghasidian 2014). The definition of this cultural tradition has revealed a high degree of cultural diversity within the Zagros Mountains during the early UP, with archeologists observing different population and developmental trajectories (Conard and Ghasidian 2011; Ghasidian 2014; Ghasidian et al. 2017, 2019).
In this context, Ghar-e Boof also offers a unique opportunity for improving our understanding of the paleoenvironmental conditions in the southern Zagros and assessing possible correlations between cultural and demographic shifts and local environmental fluctuations. Thus, here we present a detailed taxonomic analysis of the small vertebrate assemblages from the MP and early UP sequence of Ghar-e Boof. Our main goals are the following: (1) the identification and description of the different small vertebrate taxa and (2) to infer the habitat requirements of the small vertebrates in order to reconstruct the paleoenvironment in which humans lived during the Late Pleistocene at Ghar-e Boof.
Ghar-e Boof
Ghar-e Boof (N 30.2839°, E 51.4352°) is a cave located in the Dasht-e Rostam region, on the northwestern edge of the Fars Province (Fig. 1A, B). The site lies at an altitude of 905 m.a.s.l., ca. 40 m above the valley bottom, and it is situated in the Yagheh Sangar, a narrow corridor in the topography that divides the Dasht-e Rostam region in half (Conard et al. 2006; Conard and Ghasidian 2011). From this location, the Solak seasonal stream is just 200 m away towards the northeast. The inside area of Ghar-e Boof is about 60 m2, and the entrance of the cave faces north (Fig. 1C, Conard and Ghasidian 2011).
Originally documented as Eshkaft-e Yagheh Sangar, Ghar-e Boof was visited for the first time by the TISARP team in 2005 (Conard et al. 2006; Zeidi et al. 2006). Since then, the team has carried out four campaigns of excavation at the site in 2006, 2007, 2015, and 2017 (Conard et al. 2006, 2007; Conard and Ghasidian 2011; Conard and Zeidi 2019; Zeidi and Conard 2019). The excavation area is 2 × 9 m, and it extends along the north–south axis of the cave, from near the dripline at the entrance toward the back of the cave (Fig. 2, Conard and Ghasidian 2011), and is approximately 6 m deep (Heydari et al. 2021). Archeologists divided each square meter into 50-cm sub-squares and excavated artificial spits of 2 to 3 cm deep following the slope of the geological deposits (Conard and Zeidi 2019). Overall, the stratigraphic sequence of Ghar-e Boof consists of six main archeological horizons (AHs) and 13 sub-horizons (Fig. 2). For the sake of clarity, we summarize the sedimentological characteristics, thickness, absolute dates, and cultural affiliations by AH in Table 1.

Modified from Heydari et al. (2021)
Ghar-e Boof statigraphic sequence: west profile and part of the north. Radiocarbon dates are calibrated.
At the top of the sequence, AHs I and II comprise Holocene silts and ash deposits. Towards the center and bottom levels of AH II (sublayers IIa and IIb), there were many diagnostic Zarzian lithic artifacts (Zeidi and Conard 2019). However, the presence of a few pottery sherds and a grain of barley from AH IIb dated by 14C to ca. 1250 cal. BP still indicates some degree of disturbance with the uppermost Holocene sediments (Baines et al. 2014; Ghasidian 2014). The bottom of AH IIb also includes a layer of limestone cobbles with sizes from 5 to 25 cm in the central part of the excavation area, which seems to mark the distinction with the underlying undisturbed Paleolithic deposits (Conard and Ghasidian 2011).
The early UP sequence starts with AH III and spans downward until IVb. Here, OSL and radiocarbon dating analyses provided dates within the range of 42–35 kya cal. BP (Conard and Ghasidian 2011; Baines et al. 2014; Becerra-Valdivia et al. 2017; Heydari et al. 2021). These layers have yielded high concentrations of lithic artifacts representing the assemblage type used for defining the Rostamian technocomplex (Conard and Ghasidian 2011; Ghasidian 2014), which is characterized by a predominance of small bladelets, retouched bladelet tools, and abundant platform cores (Ghasidian 2014). Along with the lithic artifacts, excavators documented combustion features and uncovered personal ornaments, such as perforated shell beads and large amounts of well-preserved faunal and botanical remains (Conard and Zeidi 2019). During the Rostamian, Ghar-e Boof was used as a camp site, and hunter-gatherers primarily exploited caprines for meat and marrow, though gazelles, partridges, and tortoises might have been recurrent dietary supplements (Mata-González et al. 2022). Botanical studies also suggest that the accumulation of some large pulses might have resulted from dietary plant use by humans (Baines et al. 2014).
Currently, AHs IVc and IVd cannot be attributed with confidence to either the MP or UP due to the low find densities of lithic artifacts (Heydari et al. 2021). However, the layers lack Arjeneh points and perforated shells, which have been recorded in AHs IV to IVb and are considered characteristic components of the UP techno-cultural complexes in the Zagros. Consequently, based on an OSL date of 48–45 kya for AH IVd, Heydari et al. (2021) have proposed that AHs IVc and IVd may represent the transition period between the MP and UP at Ghar-e Boof. Moving deeper, AHs V to VI yielded MP industries, though find densities are still very low, reflecting ephemeral occupations at the site in the Dasht-e Rostam region (Bretzke and Conard 2017; Conard and Zeidi 2019; Zeidi and Conard 2019). The OSL chronology for AHs V to VI resulted in the range of 81–45 kya, at a 68% confidence interval (Heydari et al. 2021).
Zooarchaeological, archaeobotanical, and lithic analyses for these layers are ongoing. Nevertheless, the preliminary assessment of animal bones has already revealed that the humans at Ghar-e Boof were the main agent of bone accumulation in the cave, and that while they consumed mostly caprines, other small- to very large-bodied ungulates and small-game animals were also part of the diet.
Material and methods
The Ghar-e Boof small vertebrate assemblage used for this study includes a total of 755 identified specimens from the early UP and MP sequence (AHs III to VI). The material was recovered during the four excavation campaigns conducted at the site between 2006–2007 and 2015–2017 (Conard and Ghasidian 2011; Conard and Zeidi 2019). In order to collect small vertebrates, archeologists water-screened all sediments through superimposed 5- and 2-mm mesh (Ghasidian 2014; Conard and Zeidi 2019), processing in total more than 18,000 L of sediment. A few small vertebrates were also documented and recovered as single finds.
Taxonomy
In this work, we describe all recovered taxa of microvertebrates, though micromammals, amphibians, and squamate reptiles represent better proxies for paleoenvironmental reconstructions than fish or small birds, especially because their unique habitat and ecologic requirements limit their geographic range (Andrews and O’Brien 2000; Avery 2007; Blain et al. 2008; Blain 2009; Belmaker and Hovers 2011).
We identified the small vertebrate remains recovered at Ghar-e Boof mainly by using the modern reference collection at the University of Tübingen. When necessary, we also consulted several taxonomic atlases and published papers for the study of micromammals (Yusefi 2002; Pavlinov 2008; Kryštufek and Vohralík 2005, 2009; López-García 2011; Tesakov 2016; Rey-Rodríguez et al. 2020), amphibians and squamate reptiles (Delfino 2004; Blain 2009; De Lapparent et al. 2020; Biton et al. 2021), fish (Escala and Miranda 2002; Keivany et al. 2016), and small birds (Baumel et al. 1993; Wójcik 2002; Kessler 2015; Kaboli et al. 2016). We analyzed this assemblage using a ZEISS Stemi 305 stereo microscope and photographed the material using a Zeiss Stereo Discovery V8 and a Keyence VHX-500F. Moreover, the quantification of the Ghar-e Boof microvertebrate remains is solely based on the number of identified specimens (NISP), which refers to the total number of skeletal elements and/or fragments that have been identified to their taxonomic order, family, genus, or species, without involving any further derived calculation (Grayson 1984; Lyman 2008).
Taphonomy
In the present paper, we preliminarily analyzed a total of 514 micromammal remains including incisors, molars, and mandibles according to the criteria described in Andrews (1990) and Fernández-Jalvo et al. (2016), observing the degree of digestion marks. We also observed the possible compression and uniaxial mechanical deformation for the fish remains (Blanco-Lapaz et al. 2021; Frontini et al. 2021) and the possible digestion in 63 humeri and ilia belonging to Bufo/Bufotes sp. In this study, we also analyzed possible bite marks. The presence of bite marks, crushed bones, and digestive marks can give us insight into predatory activities carried out by terrestrial carnivores and birds of prey (Andrews 1990; Nicholson 1993). The preliminary observations could indicate if the predation activity by birds of prey or carnivores was the origin of at least part of the accumulation. We also analyzed the evidence of burning on microvertebrate remains, using the method of five stages of thermal-induced discoloration following Cáceres et al. (2002), which are based on heat-induced color alterations described by Shipman et al. (1984) and Stiner et al. (1995) for large mammal bones (see also Lloveras et al. 2009b; Medina et al. 2012; Walker et al. 2015, Fernández-Jalvo and Avery 2015 and Rhodes et al. 2016 for applications to microvertebrates). The stages correspond to 0 (no discoloration), 1 (yellowish with reddish-brown spots; < 100 to 300 °C), 2 (dark brown to black coloration; < 400 to 550 °C), 3 (charred bone, dark black or blue coloration over 50–100% of the surface; 500 to < 700 °C), 4 (gray-white coloration, partial calcination; 650 to < 950 °C), and 5 (calcined bone, white coloration over 50–100% of the surface; > 700 °C) (Shipman et al. 1984; Cáceres et al. 2002).
Paleoenvironmental reconstruction
Here, we use the habitat weighting method, also known as the taxonomic habitat index (Evans et al. 1981; Andrews 2006) in order to reconstruct the paleoenvironment and landscape of Ghar-e Boof. The method is based on the present distribution of each taxon in a given type of habitat where it is presently found (Evans et al. 1981; Andrews 2006; modified by Blain et al. 2008; López-García 2011; Rey-Rodríguez et al. 2020). The analysis of zooarchaeological remains recovered at Late Pleistocene sites in the Zagros Mountains has yielded taxa that are still extant in Iran (e.g., Otte et al. 2007; Mashkour et al. 2009a; Bazgir et al. 2014, 2017; Rey-Rodríguez et al. 2020). Therefore, due to the fact that there are no extinct species at Ghar-e Boof, we assume that the small vertebrate species identified in this assemblage had equivalent ecological and habitat requirements to their modern relatives.
For this study, we applied the habitat weighting method to small mammal taxa, which was adapted from Blain et al. (2008), López-García (2011), and Rey-Rodríguez et al. (2020). Here, we distinguish the following types of habitats: forest (Fo), shrubland (Sh), grassland (Gr), desert (De), wetland (We), and rocky (Ro). Each taxon has a score of 1.00, which is divided between the habitats where the species are found today (Table 2; Rey-Rodríguez et al. 2020).
The score and the habitat preference of each species were obtained from the IUCN Red List of Threatened Species (https://www.iucnredlist.org/resources/spatial-data-download). Due to the small sample size available for each particular layer, we combined all UP layers (III to IVb) and all MP layers (V to VI), excluding the presumable MP-UP shift period layers (IVc to VI AHs) since the number and type of recovered remains are not enough for applying the habitat weighting method. Nevertheless, we also examine and discuss species’ presence/absence and changes in relative species abundances by layer in order to identify possible environmental fluctuations over time at Ghar-e Boof.
Results
Taxonomic identifications, ecology, and current distribution
In the MP-UP Rostamian sequence of Ghar-e Boof, we identified a total of 498 micromammal remains to genus or species level, 16 micromammal remains to order/family level, 77 specimens identified as squamate reptiles, 75 remains of amphibians, 73 small bird remains, and 11 fish remains (Fig. 3, Table 3).

Some microvertebrate remains identified at Ghar-e Boof. A Elliobus sp. left m1 (Qu: 6/8 Nr: 939 AH: VI); B Microtus cf. irani left m1 (Qu: 6/7 Nr: 1213 AH: IVc); C Meriones vinogradovi right m1-m2 (Qu: 6/7 Nr: 625 AH: III); D Meriones lybicus (Qu: 6/7 Nr: 1223 AH: IVc); E Tatera indica right m1 (Qu: 6/9 Nr: 488 AH: III); F Mus musculus right m1 (Qu: 7/8 Nr: 136, AH: III); G Ochotona cf. rufescens fragment of left maxillary (P4-M1) and occlusal view (P4-M1) (Qu: 6/7 Nr: 151AH: IVc); H Paraechinus cf. hypomelas incomplete left mandible (p1, m1, and m2) (Qu: 6/2 Nr:196 AH: IV); I Chondrostoma sp. left branchial arch (Qu: 6/7 Nr: 743 AH: III); J Malpolon insignitus trunk vertebra (Qu: 6/9 Nr: 244 AH: VI); K Agamidae left dentary (Qu: 7/2 Nr: 76 AH: III); L Bufo/Bufotes sp. right ilium (Qu: 6/9 Nr: 252 AH: III). Qu: quadrant, Nr: register number, AH: archeological horizon
Micromammals
Order Lagomorpha Brandt, 1855.
Family Ochotonidae Thomas, 1897.
Genus Ochotona Link, 1795.
Ochotona cf. rufescens Gray, 1842.
Material
NISP = 3. All of these specimens were recovered from AH IVc and correspond to a left maxillary fragment. Only the P4 and the M1 are present, though the alveolus of the M2 is also visible.
Description and discussion
Generally, the key element to classify lagomorphs to species level is the p3, which is missing in our material. Our specimen shows the M1 and the P4 with the typical morphological characteristic to differentiate ochotonids from leporids: the absence of dentine bridges connecting anterior and posterior lobes (Laplana et al. 2015). Due the absence of the p3, we could not classify our specimen to the species level but in comparison with modern collections, the sample could be attribute to the Afghan pika (Ochotona cf. rufescens), the only representative of the Ochotonidae family in Iran (Firouz 2005; Čermák et al. 2006; Karami et al. 2016).
The record of Afghan pika at Ghar-e Boof would represent the southernmost appearance of this taxon in the Zagros. Besides Ghar-e Boof, archeologists tentatively documented Afghan pika in the MP-UP deposits of Warwasi Cave (Turnbull 1975), in the UP sequence of Yafteh Cave (Mashkour et al. 2009a), and during the Epipaleolithic of Palegawra Cave (Turnbull and Reed 1974). Hole and Flannery (1967) also reported the taxon in several other MP up to Epipaleolithic sites from the Khorramabad region, including Pa-Sangar, Ghar Arjaneh, Yafteh, Kunji, and Ghamari caves, but they did not specify from which sites Afghan pika were recovered.
Habitat and distribution
In Iran, the Afghan pika mostly lives in cold shrublands and grasslands of mountainous areas with a very wide altitudinal range, including parts of the Alborz and Zagros mountains higher than 3000 m.a.s.l. (Firouz 2005; Karami et al. 2016). Moreover, the species occurs in rocky terrain and steppes, where it either burrows or uses crevices as shelter (Firouz 2005). Although the Afghan pika inhabits areas with very low temperatures, it does not hibernate (Gromov and Erbajeva 1995; Karami et al. 2016).
Order Rodentia Bowdich, 1821.
Family Cricetidae Fisher, 1817.
Subfamily Arvicolinae Gray, 1821.
Material
NISP = 16. AH VI (NISP = 2), AH Vc (NISP = 3), AH Va (NISP = 1), AH IIIc (NISP = 2), AH IIIb (NISP = 1), AH IIIa (NISP = 4), and AH III (NISP = 3). These remains, all of them corresponding to fragmentary molars, were not identified to the genus level because of their poor preservation and a lack of key characteristics.
Genus Microtus Schrack, 1798.
Microtus sp.
Material
NISP = 24. Molars from Microtus were found in AH VI (NISP = 4), AH Vc (NISP = 1), AH IIIb (NISP = 11), and AH III (NISP = 8).
Description and discussion
Our material can only be attribute to genus level as they exhibit the basic morphology and characteristics for the Microtus genus, consisting of an anterior cap (AC) with five alternating and closed triangles. The second triangle (counting from the posterior end of the tooth) is not as wide (labiolingually) as the first (Bell and Bever 2006; López-García 2011).
Habitat and distribution
Due to the high diversity of species (about 62 are placed within this genus and are distributed across North America, Europe, and Asia) and habitats, it is not possible to attribute a specific habitat or distribution for this genus (Musser and Carleton 2005).
Microtus cf. irani.
Material
NISP = 1. The only remain belonging to this species corresponds to a left m1 from AH IVc.
Description and discussion
Four buccal and five lingual reentrant angles with a posterior lobe, seven triangles, and an AC characterize the Microtus m1 (Kryštufek and Vohralík 2009). In M. irani, triangles T1–T5 are closed, but T6–T7 are open and not parallel with one another (Rey-Rodríguez et al. 2021).
Habitat and distribution
The Persian vole is common in grasslands from Iran to the Caucasus (Karami et al. 2016).
Genus Ellobius Fischer, 1814.
Ellobius sp.
Material
NISP = 3. These specimens were recovered from AH III and correspond to a maxilla fragment showing just a left M3 and a fragmentary right M3. We did not distinguish specimens to species level due to the lack of discriminant key teeth, such as the M1 or m1 (Rey-Rodríguez et al. 2021).
Description and discussion
Remains were attributed to this genus by the presence of roots visible in adults. Molars lack cement in the re-entrant angles. Both M3 and m3 are reduced and smaller than the other molars, with three triangles on the labial side and two triangles on the lingual side (Gharkheloo and Kivanç 2003). In Iran, the genus Ellobius is currently represented by three species: E. fuscocapillus, E. lutescens, and E. talpinus (Kryštufek and Vohralík 2005; Karami et al. 2016).
Habitat and distribution
Mole-voles of the genus Ellobius are species specialized for subterranean life (Coşkun 2001; Coşkun and Ulutürk 2003). In general, Ellobius species can presently be found in steppes, grasslands, and semi-deserts from Eastern Europe to Central Asia (Kryštufek and Vohralík 2005).
Family Muridae Illiger, 1811.
Subfamily Gerbillinae Gray, 1825.
Genus Tatera Latesta, 1882.
Tatera indica Hardwicke, 1807.
Material
NISP = 1. We only identified a right lower m1 recovered from AH III.
Description and discussion
The Indian gerbil (Tatera indica) presents hypsodont molars but to a lesser degree than Meriones. The molars in adults consist of a series of plain straight plates, with three transverse plates in the first molars (Kryštufek and Vohralík 2009). The anteroconid in Gerbillinae includes initially three separate elements, exteroantroconid, interoanterioconod, and an accessory cusplet (Denys 2017), the protuconulid. Tatera shows the first two isolated cusps on minimally worn molars (Pavlinov 2008).
This gerbil has previously been documented in the MP deposits of Warwasi Cave (Turnbull 1975). According to Hashemi et al. (2006), the taxon is also present in the UP layers of Yafteh Cave, though it is not mentioned in later zooarchaeological studies about the site (see Otte et al. 2007; Mashkour et al. 2009a). In any case, our specimen constitutes the southernmost record of the Indian gerbil in the Zagros.
Habitat and distribution
The Indian gerbil is found from Mesopotamia (where it is known in Turkey, Syria, and Iraq), across Iran, Afghanistan, and Pakistan, to India and Sri Lanka (Kryštufek and Vohralík 2009). T. indica is the only species of this genus currently present in Iran (Karami et al. 2016). The species lives in steppe and desert habitats of eastern, central, and southern Iran. The Indian gerbil is less adapted to desert life than other gerbils and cannot tolerate frost (Kryštufek and Vohralík 2009; Karami et al. 2016).
Genus Meriones, Illiger 1811.
Meriones sp.
Material
NISP = 61. The molars from Meriones were not attribute to the species level because they were fragmentary or had no diagnostic characteristics They were recovered from AH VI (NISP = 1), AH Vd (NISP = 2), AH Vc (NISP = 4), AH Va (NISP = 6), AH V (NISP = 2), AH IIIb (NISP = 18), AH IIIa (NISP = 3), and AH III (NISP = 25).
Description and discussion
Typical morphology of Meriones includes semihypsodont molars with prismatic enamel triangles linked by a longitudinal crest and no trace of cusps (Dianat et al. 2017; Rey-Rodríguez et al. 2021). Although there are several species belonging to the genus Meriones, only some of them currently live in the southern Zagros Mountains (i.e., M. crassus, M. persicus, M. hurrianae, M. lybicus, and M. vinogradovi; Karami et al. 2016).
Habitat and distribution
The distribution of the genus Meriones encompasses North Africa, Central Asia, Transcaucasia, Turkey, and Pakistan (Yiğit et al. 2003; Darvish 2011; Darvish et al. 2014; Stoetzel et al. 2017). Its preferred habitats are clay and sandy deserts (Kryštufek and Vohralík 2009).
Meriones libycus Lichtenstein, 1823.
Material
NISP = 76. Molars and mandibles of this species were recovered from AH VI (NISP = 1), AH Vd (NISP = 3), AH Vc (NISP = 5), AH IIIc (NISP = 3), AH IIIb (NISP = 10), AH IIIa (NISP = 5), and AH III (NISP = 49).
Description and discussion
We provisionally attributed these specimens to the Libyn jird, but will investigate this further due to the lack of this species in our reference collection. Despite this, the Ghar-e Boof specimens attributed to M. lybicus show the typical morphology of the taxa, including semi-hypsodont molars with prismatic enamel triangles linked by a longitudinal crest and with no trace of cusps (Pavlinov 2008; Kryštufek and Vohralík 2009). We could distinguish M. lybicus from M. vinogradovi using the first lower molar (m1), since in occlusal view, the anteroconid shows an ascendent oblique elongation, but in M. lybicus, it is more narrow and flattened than in M. vinogradovi. The paraflexus-protoflexus and the metaflexus-hypoflexus pairs are more or less parallel in both species but it is clear that in M. libycus, the pairs are closer to each other than in M. vinogradovi. The second lower molar (m2) is also a key tooth to distinguish both species. M. vinogradovi presents an H-shaped m2, while in M. libycus, this molar is much more oblique (Mamkhair et al. 2007; Pavlinov 2008; Kryštufek and Vohralík 2009).
If confirmed, the presence of the Libyan jird at Ghar-e Boof would constitute the first record of this taxon during the MP in the Zagros Mountains. Until now, archeologists have only reported Libyan jird remains in the UP deposits of Yafteh Cave (Otte et al. 2007; Mashkour et al. 2009a).
Habitat and distribution
The Libyan jird is widely distributed, ranging from North Africa through Arabia, Jordan, Iraq, Syria, Transcaucasia, Iran, Turkmenistan, Afghanistan, Pakistan, and Kazakhstan to Xinjiang, in Western China (Kryštufek and Vohralík 2009). Overall, this species is well-adapted to arid conditions and inhabits mostly vegetated arid landscapes, including steppes, semi-deserts, and deserts, but is also associated with clay, loess, gravel, sandy, or hard soil substrates (Kryštufek and Vohralík 2009). Nevertheless, the Libyan jird prefers dry and exposed places (Pavlinov et al. 1990). In Iran, the species is distributed widely throughout the country, but lives mostly on Saxaul shrublands and the edges of cultivation fields and avoids Hyrcanian forests, high mountains, and coastal areas (Karami et al. 2016).
Meriones vinogradovi Heptner, 1931.
Material
NISP = 308. We identified Vinogradov’s jird molars, maxillae and mandibles in AH VI (NISP = 4), AH Vd (NISP = 9), AH Vc (NISP = 17), AH Va (NISP = 7), AH V (NISP = 4), AH IVc (NISP = 4), AH IV (NISP = 9), AH IIIc (NISP = 15), AH IIIb (NISP = 25), AH IIIa (NISP = 24), and AH III (NISP = 194).
Description and discussion
As mentioned previously, M. libycus can be distinguished from M. vinogradovi by using both the M/m1 and the M/m2 as key teeth. As a main characteristic, we observed in the m1 a less flattened AC and more distance between the paraflexus and protoflexus, which is also present between the metaflexus and protoflexus. Moreover, M. vinogradovi shows a typical H-shaped m2, unlike M. lybicus (Pavlinov 2008; Kryštufek and Vohralík 2009).
The Vinogradov’s jirds from our assemblage represent the first and oldest record of the taxon in the MP of the Zagros. So far, it has only been documented in the UP sequence of Yafteh Cave (Otte et al. 2007; Mashkour et al. 2009a).
Habitat and distribution
Presently, this species shows a range largely restricted to Transcaucasia and northern Iran, including also Syria and adjacent Turkey, but the fossil history of Vinogradov’s jird has not yet been well-documented (Kryštufek and Vohralík 2009). In Iran, the species is most common in the northwestern part of the country, and its typical habitats comprise cool semideserts and high elevation steppes (Kryštufek and Vohralík 2009). Furthermore, it requires deep soil for burrowing. Vinogradov’s jird prefers dry soils, and it does not occur on sands and on substrates of rocky debris (Kryštufek and Vohralík 2009).
Genus Mus Linnaeus, 1785.
Mus musculus Linnaeus, 1785.
Material
NISP = 4. A complete right hemi-mandible was recovered from AH III, containing all three molars (m1–m3).
Description and discussion
In our sample, the m1 shows the key characteristics for distinguishing this genus: the dental ends of mesial and central cusps on first lower molars fuse early and the mesiolabial cusp is small. The anterior half of the m1 also has a symmetrical X pattern formed at the anterior portion of the tooth (Siahsarvie and Darvish 2008; Rey-Rodríguez et al. 2020). Previously, in the Zagros, archeologists have documented this taxon in the MP and UP layers of Kaldar Cave (Bazgir et al. 2017; Rey-Rodríguez et al. 2020).
Habitat and distribution
Currently, the house mouse (M. musculus) is spread worldwide, living in close association with humans as a commensal animal, but it is also found in the wild (Firouz 2005; Karami et al. 2016). In Iran, the species is distributed throughout the whole country, with the exception of completely dry areas like the Dasht-e Kavir and the Dasht-e Lut, both located in the middle of the Iranian Plateau (Karami et al. 2016).
Order Eulipotyphla Waddell et al., 1999.
Family Erinaceidae Fischer, 1814.
Material
NISP = 3. The material comprises the distal epiphysis of a humerus, a small mandibular fragment, and an isolated molar recovered in AH IIIb (NISP = 1) and AH III (NISP = 2). Due to their fragmentary conditions and the lack of modern Iranian Erinaceidae taxa in our comparative collection, it was not possible to attribute them to genus or species.
Genus Paraechinus Trouessart, 1879.
Paraechinus cf. hypomelas Brandt, 1836.
Material
NISP = 13. The specimens consist of incomplete mandibles, with and without teeth, found in AH Vb (NISP = 1), AH IV (NISP = 8), and AH III (NISP = 4). The most complete mandible was from AH IV, which has the p1, m1, and m2 in situ.
Description and discussion
Today, four species of hedgehog are present in Iran: Erinaceus concolor, Hemiechinus auritus, Paraechinus aethiopicus, and Paraechinus hypomelas (Harrison and Bates 1991; Firouz 2005; Karami et al. 2016). The morphological characteristics of the mandible, such as the angle (close to 90°) between the ramus and the mandible body and the distribution of the cusps for m2 and m1 (Yusefi 2002; Yusefi et al. 2016), fit for the attribution of the Brandt’s hedgehog (P. hypomelas).
If our identification of these remains as Brandt’s hedgehog is correct, then it would be the first appearance of this taxon in the Paleolithic record of the Zagros Mountains. However, other species from the Erinaceidae family have been reported for this region. Archeologists found European hedgehogs (Erinaceus europaeus) or just Erinaceus sp. in the Late Pleistocene-Holocene levels of Wezmeh Cave (Mashkour et al. 2009b), in the UP layers of Gelimgoush Cave (Heydari-Guran et al. 2021b), and in the Epipaleolithic deposits of Palegawra Cave (Reed and Braidwood 1960; Turnbull and Reed 1974). Moreover, the long-eared hedgehog (Hemiechinus auritus) has also been documented in Palegawra (Turnbull and Reed 1974), and tentatively in some of the Pleistocene sites from the Khorramabad region studied by Hole and Flannery (1967). Finally, Bazgir et al. (2014) reported indeterminate Erinaceidae remains from the Pleistocene deposits of Ghamari Cave.
Habitat and distribution
Brandt’s hedgehog has been reported across Iran, except in northern forested areas and other northwestern regions of the country. This species usually lives in deserts and the foothills of steppe areas (Karami et al. 2016).
Order Chiroptera Blumenbach, 1779.
Material
NISP = 2. Only two specimens of chiroptera were found in our assemblage, which consist of two distal fragments of humeri, one in AH IV, and another in AH III. Due to the wide diversity of different genera belonging to this order in Iran (e.g., Eptisicus, Hipposideros, Hypsugo, Myotis, Miniopterus, Nyctinomus, Nyctalus, Otonycteris, Pipistrellus, Rhinolophus, Rhinopoma, Tadarida, Vespertilio, to name just a few; Karami et al. 2016), and the lack of Iranian bats in modern comparative collections, it was not possible to classify the remains further.
Squamate reptiles
Order Squamata Oppel, 1811.
Family Agamidae Gray, 1827.
Material
NISP = 63. Except for AHs VI, Vd, and IIIa, dentaries belonging to this family are present throughout the entire sequence: AH Vc (NISP = 1), AH Vb (NISP = 4), AH Va (NISP = 4), AH V (NISP = 1), AH IVd (NISP = 2), AH IVc (NISP = 4), AH IVb (NISP = 5), AH IVa (NISP = 6), AH IV (NISP = 15), AH IIIc (NISP = 8), AH IIIb (NISP = 8), and AH III (NISP = 5). The main characteristic of the Agamidae dentaries is the presence of an acrodont dentition showing a general triangular shape and the teeth are laterally flattened (Blain 2009). Once again, based on the wide diversity of this family in Iran and the lack of specimens in modern comparative collections, we could not determine the Ghar-e Boof specimens to genus or species level. Several genera are present in Iran, such as Calotes, Laudakia, Paralaudakia, Phrynocephalus, and Trapelus, with each of the genera containing several species (Mozaffari et al. 2016).
Family Lacertidae Oppel, 1811.
Material
NISP = 1. We only found one fragmentary trunk vertebra that belongs to the family Lacertidae, recovered from AH IV, but a more precise identification was not possible. Overall morphology of this vertebra is consistent with an attribution to this family excluding other groups of Squamata such as Agamidae or Ophidia.
Suborder Ophidia Linnaeus, 1758.
Material
NISP = 4. Here, we include very fragmentary trunk vertebrae from AH Vb (NISP = 1), AH V (NISP = 1), and AH III (NISP = 2).
Family Lamprophiidae Fitziger, 1843.
Genus Malpolon Fitzinger, 1826.
Malpolon insignitus Geoffroy St. Hilarire, 1809.
Material
NISP = 7. Isolated trunk vertebrae were present in AH VI (NISP = 1), AH Vb (NISP = 1), and AH III (NISP = 5).
Description and discussion
We attributed these vertebrae to the Eastern Montpellier snake (M. insignitus) on the basis of the haemal keel, which is thin and sharp before the condyle (Bendrey et al. 2020). This is the first record of M. insignitus during the MP in the Zagros Mountains. Previously, Mashkour et al. (2009b) had only identified the Western Montpellier snake (M. monspessulanus) in Wezmeh Cave. The authors, probably used to the old taxonomy when they studyied those remains. Our current knowledge about the distribution of species of the genus Malpolon dictates that the species M. monspessulanus is not distributed eastward beyond northwestern Italy and central Algeria, so its identification outside of this region is not correct nowadays and the individuals considered before to be M. monspessulanus are now attributed to the new species M. insignitus (Mozzaffari et al. 2016).
Habitat and distribution
The Eastern Montpellier snake is widely distributed across Eurasia (Mozaffari et al. 2016). The species lives in diverse types of habitats, such as forest, shrublands, grasslands, foothills, rocky areas, and semi-desert (Firouz 2005; Mozaffari et al. 2016).
Family Viperidae Oppel, 1811.
Material
NISP = 2. We only identified two isolated trunk vertebrae as Viperidae, one from AH Va and another one from AH III. Although the shape of the caudal area of the postzygapophysis is typical for the genus Vipera (Markert 1978; Blain 2009), we could not attribute these specimens with confidence to the genus level. The remains show poor preservation and there is a lack of Iranian taxa in the modern comparative collections. Moreover, the family shows a great diversity in Iran, with 13 different species belonging to the genera Cerastes, Echis, Eristocophis, Gloydius, Macrovipera, Montivipera, Pseudocerastes, and Vipera (Mozaffari et al. 2016).
Amphibians
Order Anura Duméril, 1806.
Material
NISP = 12. Here, we included several fragmentary postcranial elements such as vertebrae, humeri, and ilium that lack discriminant traits. We found indeterminate Anura specimens in AH IVc (NISP = 1), AH IVa (NISP = 1), AH IIIb (NISP = 2), and AH III (NISP = 8).
Family Bufonidae Gray, 1825.
Genus Bufo/Bufotes Garsault, 1764.
Bufo/Bufotes sp.
Material
NISP = 63. Most of the toad (Bufo/Bufotes sp.) remains that correspond to ilia and humeri were present in AH Vc (NISP = 4), AH Vb (NISP = 5), AH Va (NISP = 3), AH IV (NISP = 2), AH IIIc (NISP = 1), AH IIIb (NISP = 20), AH IIIa (NISP = 4), and AH III (NISP = 24).
Description and discussion
For anurans, the ilium is widely considered to be the single best element upon which to base fossil identifications when isolated bones are all that are available (Bever 2005). The Bufo/Bufotes genus determination is based on the absence of a dorsal crest and the presence of an upper tuberosity that is flat and exhibits a rounded dorsal edge (Blain 2009). In Iran, there are several species that belong to the genus Bufo/Bufotes, such as B. bufo, B. eichwaldi, B. kavirensis, B. luristanicus, B. oblongus, B. olivaceus, B. stomaticus, B. surda, and B. viridis (Yousefi-Siahkalroodi et al. 2013). Due to this wide diversity, and the lack of Iranian toads in our comparative collection, it was not possible to attribute the remains recovered at Ghar-e Boof to the species level.
Habitat and distribution
Bufo/Bufotes represent the most numerous and wide spread genus of amphibians in Iran, most likely because of their ecological tolerance (Firouz 2005). Overall, amphibians are very dependent on humid habitats (Blain et al. 2008); however, toads have a particularly dry skin through which water is not easily lost, enabling them to live and survive in more arid areas than other amphibians (Firouz 2005). Therefore, although most toads need to have access to water resources and thus inhabit areas near marshes, shallow ponds, streams, and rivers, they can also live in meadows, rocky areas, and dry areas (Yousefi-Siahkalroodi et al. 2013).
Fish
Order Cypriniformes Bleeker, 1859.
Family Leusciscinae Bonaparte, 1835.
Material
NISP = 10. All of these specimens correspond to precaudal and caudal vertebrae from AH IIIb (NISP = 8), AH IIIa (NISP = 1), and AH III (NISP = 1). We could not identify the remains up to the genus or species level because of the lack of Iranian taxa in our comparative collection, as well as the wide diversity of the family Leuciscinae in Iran (Keivany et al. 2016).
Genus Chondrostoma Agassiz, 1832.
Chondrostoma sp.
Material
NISP = 1. This genus is represented in our assemblage by just a left pharyngeal arch fragment from AH III.
Description and discussion
We determined this branchial arch as nase (Chondrostoma sp.) based on the compressed hook shape of the branchial teeth and the presence of just one tooth row (Escala and Miranda 2002). Currently, only two species are presently living in Iran: C. cyri and C. regium (Keivany et al. 2016). However, due to the lack of these species in our comparative collection, as well as the fragmentary state of this specimen, we could not attribute it to either of the two species.
Our study represents the first record of this genus in a Paleolithic site of the Zagros. Overall, fish remains in MP and UP Iranian sites are very rare. So far, archeologists have only reported the presence of cyprinids and chubs (Leuciscus sp.) in Yafteh Cave (Otte et al. 2007; Mashkour et al. 2009a) and some undetermined fish remains in Palegawra Cave, which still need to be analyzed (Asouti et al. 2020).
Habitat and distribution
Kura undermouth (C. cyri) is located exclusively in the Caspian Sea basin (Aras River) and lives mainly in streams and rivers. As for the Mesopotamian nase (C. regium), it is more widely distributed and is present in the Tigris (Karun, Marun, Bazoft, Karkheh), Isfahan (Zayandehrud River), and Fars (Kor River) basins. The Mesopotamian nase lives in lakes, reservoirs, and rivers with stone grounds and still waters (Keivany et al. 2016).
Small birds
Order Passeriformes Linnaeus, 1758.
Material
NISP = 73. The small bird assemblage of Ghar-e Boof consists mostly of postcranial elements such as humeri, tibiotarsus, tarsmetatarsus, or vertebrae recovered from almost all layers, except for AHs V and IIIa: AH VI (NISP = 4), AH Vd (NISP = 2), AH Vc (NISP = 2), AH Vb (NISP = 7), AH Va (NISP = 4), AH IVd (NISP = 3), AH IVc (NISP = 10), AH IVb (NISP = 8), AH IVa (NISP = 1), AH IV (NISP = 14), AH IIIc (NISP = 9), AH IIIb (NISP = 4), and AH III (NISP = 5). In Iran, the order Passeriformes encompasses more than 200 species (Kaboli et al. 2016). Due to the great diversity of species and the lack of Iranian taxa in our collection, we could not go deeper in the taxonomical classification of these remains. Therefore, this material would be considered a subject of study for future researchers.
Taxonomic remarks
In the MP sequence of Ghar-e Boof (Table 3, Fig. 3), the most abundant identified taxa are jirds (Meriones sp. and M. vinogradovi, with a few examples of M. cf. lybicus). Among other genera and species of micromammals, we also documented the presence of voles (Microtus sp.), mole-voles (Ellobius sp.), and Brandt’s hedgehog (Paraechinus cf. hypomelas). Moreover, we identified squamate reptiles and amphibians, such as agamids, lizards, vipers, Eastern Montpellier snake (M. insignitus), toad (Bufo/Bufotes sp.), and other indeterminate Anura (Table 3).
Except for AH V, small birds or Passeriformes were recorded throughout the entire MP sequence. Overall, rodents are well-represented in AHs VI to V, with the exception of AH Vb. However, they are practically missing in AHs IVd and IVc, and also in AHs IVb to IV. The drastic reduction of rodents corresponds with the transition period between the MP and the beginning of the early UP Rostamian techno-complex (Heydari et al. 2021; see Table 1). The only small mammals recovered in these layers are the Afghan pika (O. cf. rufescens, NISP = 3), the Persian vole (M. cf. irani, NISP = 1), and the Libyan jird (M. cf. lybicus, NISP = 4), all of them in AH IVc, along with a few squamate reptiles and amphibians (agamids, lizards, toads, and indeterminate Anura) and small birds.
In the early UP Rostamian layers of Ghar-e Boof, the majority of the rodents reappear in AH IIIc (Table 3). From AH IIIc upwards, the small vertebrate assemblage is once again dominated by jirds (Meriones sp. and M. vinogradovi, M. cf. libycus) followed by voles (Microtus sp.), mole-voles (Ellobius sp.), Indian gerbil (T. indica), and the house mouse (M. musculus). Alongside these rodents, we also recorded insectivores, including the Brandt’s hedgehog (P. cf. hypomelas), and other indeterminate Erinaceidae and bats. Regarding squamate reptiles, amphibians, and small birds, we did not observe any apparent changes with previous horizons. However, for the first time, we documented fish remains in AHs IIIb to III, represented by indeterminate cyprinids and nase (Chondrostoma sp.).
Taphonomic observations
According to our preliminary observations of different degrees of digestion in our assemblage of micromammals and Bufo/Bufotes remains (mostly very light or light; < 20% of the bone surface is damaged), the possible predator responsible for part of the accumulation could be a category 1 predator such as the Barn owl (Tyto alba, Fernández-Jalvo et al. 2016). This species, with a generalist diet in areas where prey is scarce, is currently present in the area. Barn owls are also cavity nesters and are sedentary, indicating that their prey spectrum is assumed to be a good representation of the ecosystem in which they are present (Kaboli et al. 2016). No mechanical deformation, breakage, or polishing on the bone surface is present in cranial or postcranial elements (Fernández-Jalvo et al. 2016; Frontini et al. 2021).
We also document the presence of one individual in layer III in anatomical connection (skull, mandibles, and cervical vertebrae) likely indicating in situ mortality, although some birds of prey such as the Eurasian eagle owl could produce pellets with skulls in connection with mandibles (Andrews 2006; Fernández-Jalvo et al. 2016). In this case, the presence of cervical vertebrae in anatomical connection and lack of digestion marks on teeth further indicates that the animals were not introduced to the site by predators. Other microvertebrates seemed to have accumulated similarly, such as agamids, which can die of thermal stress during hibernation (Rastegar-Pouyani and Torki 2007). Bite marks, crush marks, or digestive marks were also absent on the fish remains, which means that carnivores or birds can be ruled out as accumulative agents for fish specimens (Nicholson 1993).
In terms of burning, we recovered just a dozen burnt remains, all of them from the upper layers of the sequence (AH III). All of these specimens show stage 3 of thermal discoloration, characterized by blackening across the entire bone surface. This stage indicates a fire temperature from 500 to < 700 °C (Cáceres et al. 2002).
In general, our observations are preliminary and further ongoing taphonomic studies will provide further data about the origin of the microvertebrate accumulation and post-depositional modifications.
Paleoenvironmental reconstructions
We present the results of our paleoenvironmental reconstructions obtained with the habitat weighting method in Fig. 4. These data, based exclusively on micromammals, indicate that the paleoenvironment of Ghar-e Boof was mainly characterized by warm, arid conditions, with the presence of shrublands and rocky terrain. These environmental conditions are mostly derived from the predominance of Vinogradovi’s jird throughout the entire stratigraphic sequence (Tables 2 and 3). The results also provide evidence of wetlands and permanent water sources close to the site, which is inferred from the presence of Lybian jird in both MP and UP layers and the house mouse in the UP. To a lesser extent, the surrounding landscape of the cave included grasslands, open forests, and desert or semi-desert areas, though their relative proportions fluctuated between the MP and UP (Fig. 4).

Results of the habitat weighting method for MP and UP layers based on NISP. Forest (Fo), shrubland (Sh), grassland (Gr), desert (De), wetland (We), and rocky (Ro)
Although desert and semi-desert conditions decrease during the UP, there is also an increase in grassland-adapted species, in parallel with a decline in forest habitats (Fig. 4). Overall, our data seem to suggest a slight increase in aridity at Ghar-e Boof during the UP in comparison to the MP.
Some squamate reptiles, amphibians, and fish support the general paleoenvironmental conditions inferred from small mammal taxa. Among all the squamate reptiles recovered at Ghar-e Boof, we were only able to identify the Eastern Montpellier snake (M. insignitus) to the species level, while we classified the rest to family (Agamidae, Lacertidae, and Viperidae). The Eastern Montpellier snake is not a good paleoenvironmental indicator, since it inhabits diverse types of habitats (i.e., forest, shrublands, grasslands, rocky areas, and semi-deserts, Firouz 2005; Mozaffari et al. 2016). However, most Viperidae and Agamidae taxa in Iran live in warm, rocky outcrops, while there are some species that can be found near small bushes, or in arid, semi-desert, or desert areas (Firouz 2005; Mozaffari et al. 2016). Toads (Bufo/Bufotes sp.) in the assemblages suggest the presence of meadows and wetlands, such as marshes and ponds (Yousefi-Siahkalroodi et al. 2013). Moreover, the recovery and identification of fish remains, such as Leuciscinae cyprinids and nase, reflect the proximity of the cave to rivers with rocky beds and still waters (Keivany et al. 2016). As mentioned above, Ghar-e Boof is located about 200 m away from the Solak seasonal stream. Besides this stream, the Dasht-e Rostam region is drained by the Fahliyan River, with a perennial flow, and the Shiv stream, also seasonal and contains more than 50 springs (Conard and Ghasidian 2011; Heydari-Guran 2014). Therefore, despite the predominance of warm, arid conditions during most of the MP and UP at the site, hunter-gatherers had access to permanent and abundant water resources during most of the year.
Discussion
Ghar-e Boof: paleoenvironmental reconstructions and archeological implications
Our results provide new data for a better understanding of the paleoenvironmental context of Ghar-e Boof based on microvertebrates. The combination of data belonging to the micromammals, reptiles, amphibians, and fish record is an important tool to characterize the MP-UP landscape around Ghar-e Boof as dominated by warm, arid conditions, with the presence of shrublands and rocky terrain. Amphibians and fish also indicated the presence of water sources. Available archaeobotanical and zooarchaeological data describe a similar landscape characterized by dry/arid conditions with the presence mostly of grass and scrubland (Baines et al. 2014), inhabited by sheep/goat and gazelles (Mata-González et al. 2022).
A second goal of this study was to assess chronologically whether the cultural and/or demographic shifts documented at Ghar-e Boof correlate with local environmental fluctuations. Find densities of lithic artifacts were low in AHs IVd and IVc (Conard and Zeidi 2019; Heydari et al. 2021), so any cultural attribution to either the MP or UP has to be made with caution. However, these layers lack characteristic components associated with the Zagros UP techno-cultural complexes, such as Arjeneh points and perforated shells, which indeed have been observed in the subsequent upper layers (AHs IVb–IV and above; Heydari et al. 2021). Whereas OSL ages for AH IVd fall in the range of 48–45 kya, AHs IV to IVa have been dated between 42 and 40 kya (Heydari et al. 2021). As a result, in the same study, Heydari and colleagues suggested that AHs IVc–IVd represent the period between the MP and the Rostamian, while AHs IVb–IV correspond to the onset of the UP at Ghar-e Boof and in the Zagros Mountains as a whole. Based on the analysis of lithic artifacts, Bretzke and Conard (2017) hypothesized that the early UP occupation in the southern Zagros might have been related to a displacement of small groups carrying MP industries. There is the question as to whether paleoenvironmental fluctuations contributed to such a cultural and/or demographic replacement at Ghar-e Boof or in the Dasht-e Rostam region more broadly. Sample size is relatively small for AHs IVd to IV in comparison with other MP and UP layers. Nonetheless, the analysis of species presence/absence, along with the discussion of their habitat preferences and requirements, also allows us to draw qualitative paleoenvironmental inferences for these layers. In general, as mentioned above, rodents (mostly murids) are well-represented in AHs VI to V (MP) and AHs IIIc to III (UP). However, one of the most striking results of our study is that during the shift from the MP to the UP (AHs IVd to IV, Table 3), we document a temporary drastic reduction of rodents and the first and only record of Afghan pika (O. cf. rufescens) at the site occurs in AHs IVc.
Although some authors associate remains of lagomorphs with the presence of large birds of prey (Redpath et al. 2009; Lloveras et al. 2009a; Lloveras et al. 2017), our preliminary taphonomic observations would indicate that only the barn owl (T. alba) could be the main accumulating agent of at least part of the microvertebrate assemblage in Ghar-e Boof, excluding the activity of large birds of prey. Archeologists have interpreted the presence of pikas as a possible sign of colder climatic conditions and rugged, open landscapes (Turnbull and Reed 1974; Laplana et al. 2015; Kandel et al. 2017). The drastic reduction of rodents and the presence of the Afghan pika could indicate colder and/or drier conditions than today (Laplana et al. 2015). In other studies, remains of pikas are only recovered from northern sites in Zagros Mountains such as Warwasi Cave (Turnbull 1975) or Yafteh Cave (Mashkour et al. 2009a). Thus, the available data indicate a transitionary time period with colder and/or drier conditions between MP and UP (AHs IVd–IV), which seems to correlate with a change in the techno-cultural industries recovered at Ghar-e Boof.
Our results would support a plausible scenario in which scattered groups of MP hunter-gatherers had to face harsher environmental conditions, visiting Ghar-e Boof very sporadically until their disappearance from the Dasht-e Rostam region ca. 48–45 kya. Chronological investigations carried out at the site have suggested the arrival of AMHs ca. 45–42 kya (Becerra-Valdivia et al. 2017; Heydari et al. 2021). This shift was accompanied by a major technological change and the rise of symbolic artifacts, such as personal ornaments (Bretzke and Conard 2017; Conard and Zeidi 2019; Heydari et al. 2021). Despite the continuity of relative cold, arid conditions and an open landscape until 40–39 kya, AMHs were able to populate the intermountain valleys of the southern Zagros Mountains, possibly thanks to new and more efficient hunting technologies as indicated by the predominance of bladelets in the early UP Rostamian assemblages (Bretzke and Conard 2017). After the temporary cold phase, we observe an increase in occupation intensity at Ghar-e Boof during the early UP, reflecting more frequent visits to the site, longer occupation periods, and/or even larger human populations spread across the region (Mata-González et al. 2022). It is likely that relatively better and more stable environmental conditions led to a more intense use of Ghar-e Boof a few millennia after the initial arrival of AMHs, as we recently hypothesized (Mata-González et al. 2022).
Ghar-e Boof microvertebrate record in the context of the Zagros Mountains
In Table 4, we compare the identified species from Ghar-e Boof with other MP and UP small vertebrate assemblages recovered in the Zagros Mountains, including Kaldar Cave (Bazgir et al. 2017; Rey-Rodríguez et al. 2020, 2021), Shanidar Cave (Perkins 1964; Tilby et al. 2022), Warwasi Cave (Turnbull 1975), Yafteh Cave (Hashemi et al. 2006; Otte et al. 2007; Mashkour et al. 2009a), and Palegawra Cave (Turnbull and Reed 1974; Asouti et al. 2020).
In addition to small vertebrates having the potential to inform us about local paleoenvironmental conditions, the establishment of inter-site comparisons, along with the combination of different sets of paleoenvironmental data, can also provide a better understanding of regional- and trans-regional population dynamics and human habitat preferences. It is worth noting that microvertebrate remains have also been reported in Gar Arjeneh, Kunji Cave, Pa-Sangar and Ghamari Cave (Hole and Flannery 1967; Bazgir et al. 2014), and Wezmeh Cave (Mashkour et al. 2009b). Nonetheless, these assemblages are excluded from our comparisons because either the available species lists include very few taxa, as is the case of Ghamari Cave (Hole and Flannery 1967; Bazgir et al. 2014), or there is evidence of disturbances, such as at Wezmeh Cave (Mashkour et al. 2009b).
The most striking difference between the MP small vertebrate assemblages of Kaldar Cave, Shanidar Cave, Warwasi Cave, and Ghar-e Boof is the relatively high richness of micromammals observed at Kaldar Cave in comparison with the other two sites, although it could be related to sample size bias. Rey-Rodríguez et al. (2020) reported 10 different genera of rodents, some of them even with more than one species (i.e., Persian vole, M. irani; social vole, M. socialis; Southern mole-vole, E. fuscocapillus; and Transcaucasian mole-vole, E. lutescens). Kaldar Cave is also rich in squamate reptiles and amphibians, though so far only toad (Bufo/Bufotes sp.) and sand boa (Eryx sp.) were identified to the genus level, while the rest were assigned to family (Bazgir et al. 2017). Despite that, the local environmental conditions were similar at the four sites. Like the situation at Ghar-e Boof, the small vertebrate assemblage of Kaldar Cave indicates that during the MP, the surrounding landscape of the site was characterized by dry, open steppe with some vegetation cover and active water resources nearby, but lower temperatures and more arid conditions than today (Bazgir et al. 2017; Rey-Rodríguez et al. 2020). Turnbull (1975) also argued that Warwasi Rockshelter had a rocky, open environment, covered sparsely with bushes, and with colder and drier conditions than at present. Tilby et al. (2022) observed similar drier and colder conditions at Shanidar Cave during the MP, although the authors also indicate relatively warm, wet conditions at the base of the sequence.
As for the UP, species representation at Kaldar and Warwasi were more or less the same as the MP, with a few exceptions: first, Persian jird (Meriones cf. persicus) is only present in the UP record of Warwasi (Turnbull 1975); second, mouse-tailed dormouse (Myomimus sp.) and Persian vole (M. irani) were not reported in the UP layer of Kaldar, whereas archeologists recorded Günter’s vole (M. guentheri), which were absent in the MP (Rey-Rodríguez et al. 2020); and finally, the UP sequence of Kaldar also show a greater diversity of squamate reptiles (Bazgir et al, 2017). However, Turnbull (1975) did not mention any environmental shifts during the complete stratigraphic sequence of Warwasi (from the MP to the Epipaleolithic). Likewise, Rey-Rodríguez et al. (2020) did not observe any major paleoenvironmental and paleoclimatic changes, though the MP was slightly colder and drier. The UP small mammal assemblages of Yafteh Cave also show strong similarities with Ghar-e Boof, including the presence of Lybian jird (M. libycus), Vinogradov’s jird (M. vinogradovi), Indian gerbil (Tatera indica), voles (Microtus sp.), and mole voles (Ellobius sp.) (Hashemi et al. 2006; Otte et al. 2007; Mashkour et al. 2009a). Indeed, archeologists have suggested a landscape composed of arid lowlands, piedmont and cool uplands, and some woodlands (Otte et al. 2007). The Epipaleolithic deposits of Palewgara Cave (Turnbul and Reed 1974) have also yielded small vertebrate remains, which indicate a steppe environment, with colder temperatures than today in northern Iraq (Turnbull and Reed 1974). Recently, Asouti et al. (2020) re-excavated the site and confirmed previous environmental data, highlighting the human exploitation of grasslands and shrublands, and some sparsely forested areas.
In addition, there are fish remains at Yafteh (indeterminate cyprinids and Leuciscus sp.), which seem to have been accumulated by raptors and/or carnivores, but archeologists do not completely rule out a possible anthropogenic origin as a result of fishing activities (Otte et al. 2007; Mashkour et al. 2009a). Similarly, it is possible that the fish remains at Ghar-e Boof were accumulated anthropogenically, as there are no bite or digestion marks implicating raptors and/or carnivores in the accumulation of the remains.
In general, paleoenvironmental reconstructions drawn on the basis of small vertebrate analyses from MP and UP sites in Zagros support other paleoenvironmental data obtained from sediment cores and archaebotanical studies (i.e., van Zeist and Bottema 1977; Wasylikowa 2005; Djamali et al. 2008a, 2011; Baines et al. 2014; Fiacconi and Hunt 2015; Allúe et al. 2018). Therefore, both MP and UP humans inhabited and exploited mosaic landscapes with relatively similar environmental conditions across different regions of the Zagros Mountains.
The number of sites in the Zagros with both well-dated MP and UP deposits has increased considerably during the last few years (Becerra-Valdivia et al. 2017; Heydari et al. 2021; Heydari-Guran et al. 2021a, b). Nevertheless, Kaldar Cave represents the only site at which archeologists have evaluated the possible influence of paleoenvironmental conditions on cultural changes in Neanderthal and AMH populations (Rey-Rodríguez et al. 2020), though they did not find any major environmental or climatic changes during the MP and UP.
The small vertebrate assemblages of Ghar-e Boof indicate a relatively short phase with colder and/or drier environmental conditions during the shift from the MP to UP. The palynological study of sediment cores from Lake Urmia (northern Zagros), along with loess sequences from the Basin of Persepolis (southern Zagros), have documented several environmental and climatic changes during the Late Pleistocene (Kehl et al. 2005; Djamali et al. 2008a, 2008b). For instance, Lake Urmia shows evidence of an increase in lake-level corresponding to the middle part of last glacial period (MIS 3), suggesting lower evaporation rates caused by a decrease in summer temperatures (Djamali et al. 2008b). Thus, the MP and early UP deposits of Ghar-e Boof could reflect changes in the local environmental signal.
Our results are quite similar in comparison with the taphonomic analysis of Kaldar Cave. Rey-Rodríguez et al. (2020) observed that the predation activity was the origin of at least part of the accumulation of the microvertebrate remains in this site. The authors hypothesized that a category 3 predator, such as the tawny owl (Strix aluco) or the Eurasian eagle owl (Bubo bubo), accumulated the remains. Based on our samples, a category 1 predator such as the Barn owl (Tyto alba) could be the accumulation agent of at least part of the assemblage in Ghar-e Boof. In Shanidar Cave, the results are different and Tilby et al. (2022) observed a constant presence of digestion marks, a fact that would indicate an important accumulation of the remains by birds of prey. The authors also highlighted a gradual increase in rockfall damage towards the upper layers of the site, indicating a slight shift produce by a relatively wet, warm condition in the lower levels. In Shanidar Cave (Tilby et al. 2022), like in Ghar-e Boof, there are also some burnt specimens present, suggesting proximity to hearths, although those remains appear sporadically mixed with the majority of unburnt bones. In terms of taphonomy, further quantitative analyses would need to be conducted to more thoroughly clarify the origin of the microvertebrate accumulation in Ghar-e Boof.
Conclusions
Our study represents the first taxonomic analysis of the small vertebrate assemblages from the MP and early UP sequence of Ghar-e Boof, along with paleoenvironmental reconstructions. We recovered and studied an NISP of 755 of micromammals, squamate reptiles, amphibians, fish, and small birds. Among these taxa, this study shows the first record of Brandt’s hedgehog (P. cf. hypomelas) and nase (Chondrostoma sp.) in a Paleolithic locality of the Zagros Mountains, the southernmost record of the Indian gerbil (T. indica) and the Afghan pika (O. cf. rufescens) of this region, and, finally, the first record of two jird species (M. cf. libycus and M. vinogradovi) in the MP of the southern Zagros.
The results of our paleoenvironmental reconstructions suggest a landscape mainly characterized by warm, arid conditions, with dry, open meadows, a rocky terrain, and water sources close to the site during most of the sequence. Alongside the strategic location of Ghar-e Boof within the Dasht-e Rostam region, these environmental conditions favored frequent visits to the site, which provided hominins access to different terrestrial and aquatic, plant, and animal resources (Mata-González et al. 2022).
During the shift from the MP to the UP, we document a short period with colder and/or drier conditions, based on the presence of the Afghan pika and the temporary drastic reduction of rodents. This cold and relatively dry phase seems to correlate with a change in the techno-cultural record at Ghar-e Boof, which might reflect a population shift and the arrival of AMHs. Adverse environmental conditions during the shift from the MP to UP appear to have affected MP hominins in the region. During this period of environmental change, groups occupied the site sporadically. Despite the presence of relatively cold, arid conditions, which lasted until ca. 40–39 kya, AMHs were able to spread across the intermountain valleys of the southern Zagros Mountains, possibly due to more efficient hunting technologies (Bretzke and Conard, 2017). Ongoing archaeobotanical analyses will soon allow us to test this hypothesis further. At present, the microvertebrate record provides the most reliable insights into the environmental conditions that prevailed during the late MP and early UP of the southern Zagros, a period of radical change in hominin population dynamics and cultural adaptations.
Change history
09 October 2022
Missing Open Access funding information has been added in the Funding Note.
References
Allúe E, Expósito I, Tumung L, Ollé A, Bazgir B (2018) Early evidence of Prunus and Prunus cf. amygdalus from Palaeolithic sites in the Khorramabad valley, western Iran. CR Palevol 17:335–345
Andrews P (1990) Owls, caves and fossils: predation, preservation, and accumulation of small mammal bones in caves, with an analysis of the Pleistocene cave faunas from Westbury-sub-Mendip, Somerset. The University of Chicago Press, Chicago, UK, p 231
Andrews P (2006) Taphonomic effects of faunal impoverishment and faunal mixing. Palaeogeogr, Palaeclimatol, Palaeoecol 241:572–589
Andrews P, O’Brien EM (2000) Climate, vegetation, and predictable gradients in mammal species richness in southern Africa. J Zool 251:205–231
Asouti E, Baird D, Kabukcu C, Swinson K, Martin L, García-Suárez A, Jenkins E, Rasheed K (2020) The Zagros Epipalaeolithic revisited: new excavations and 14C dates from Palegawra cave in Iraqi Kurdistan. PLoS ONE 15(9):e0239564
Avery D (2007) Micromammals as palaeoenvironmental indicators of the southern African quaternary. Trans Royal Soc South Afr 62(1):17–23
Baines JA, Riehl S, Conard NJ, Zeidi M (2014) Upper Palaeolithic archaeobotany of Ghar-e Boof cave, Iran: a case study in site disturbance and methodology. Archaeol Anthropol Sci 7:245–256
Baumel JJ, King AS, Breazile JE, Evans HE, Vanden Berge JC (1993) Handbook of avian anatomy, 2nd edn. Massachusetts, Cambridge, p 775
Bazgir B, Otte M, Tumung L, Ollé A, Deo SG, Joglekar P, López-García JM, Picin A, Davoudi D, van der Made J (2014) Test excavations and initial results at the Middle and Upper Paleolithic sites of Gilvaran, Kaldar, Ghamari Caves and Gar Arjene Rockshelter, Khorramabad Valley, Western Iran. CR Palevol 13:511–525
Bazgir B, Ollé A, Tumung L, Becerra-Valdivia L, Douka K, Higham T, van der Made J, Picin A, Saladié P, López-García JM, Blain HA, Allué E, Fernández-García M, Rey-Rodríguez I, Arceredillo D, Bahrololoumi F, Azimi M, Otte M, Carbonell E (2017) Understanding the emergence of modern humans and the disappearance of Neanderthals: insights from Kaldar Cave (Khorramabad Valley, Western Iran). Sci Rep 7:43460
Becerra-Valdivia L, Douka K, Comeskey D, Bazgir B, Conard NJ, Marean CW, Ollé A, Otte M, Tumung L, Zeidi M, Higham TFG (2017) Chronometric investigations of the Middle to Upper Paleolithic transition in the Zagros Mountains using AMS radiocarbon dating and Bayesian regression modelling. J Hum Evol 109:57–69
Bell C, Bever GS (2006) Description and significance of the Microtus (Rodentia: Arvicolinae) from the type Irvington fauna, Alameda County, California. J Vertebr Paleontol 26(2):371–380
Belmaker M, Hovers E (2011) Ecological change and the extinction of the levantine Neanderthals: implications from a diachronic study of micromammals from Amud cave, Israel. Quat Sci Rev 30:319–3209
Bendrey R, Van Neer W, Bailon S, Rofes J, Herman J, Morlin M, Moore T (2020) Animal remains and humananimal-environment realationships at Early Neolithic Bestansur and Shimshara. In: Matthews R, Matrrhews W, Raheem KR, Richardson A (eds) The early neolithic of the eastern fertile crescent: Excavations at Bestansur and Shimshara, Iraqui Kurdistan. Central Zagros Archaeological Project CZAP Reports Volume 2, pp 311–352
Bever GS (2005) Variation in the ilium of North American Bufo (Lissamphibia: Anura) and its implications for species-level identification of fragmentary anuran fossils. J Vertebr Paleontol 25(3):548–560
Biton R, Bailon S, Birkenfeld M, Bridault A, Khalaily H, Valla FR, Rabinovich R (2021) The anurans and squamates assemblage form Final Natufian Eynan (Ain Mallaha, Israel) with an emphasis on snake-human interactions. PLoS ONE 16(2):e0247283
Blain HA (2009) Contribution de la paléoherpétofaune (Amphibia & Squamata) à la connaissance de l’évolution du climat et du paysage du Pliocène supérieur au Pléistocènce du climat et du paysage du Pliocènce supérieur au Pléistocènce moyen d’Espagne. Treballs Del Museu De Geologia De Barcelona 16:39–170
Blain HA, Bailon S, Cuenca-Bescós G (2008) The Early-Middle Pleistocene palaeoenvironmental change based on the squamate reptile and amphibian proxies at the Gran Dolina site, Atapuerca, Spain. Palaeogeogr Palaeclimatol Palaeoecol 261:177–192
Blain HA, Bailon S, Cuenca-Bescós G, Arsuaga JL, Bermúdez de Castro JM, Carbonell E (2009) Long-term climate record inferred from early-middle Pleistocene amphibian and squamate reptile assemblages at the Gran Dolina Cave, Atapuerca, Spain. J Hum Evol 56:55–65
Blanco-Lapaz A, Martínez-Monzón A, Blain H-A, Cuenca-Bescós G (2021) Early-Middle Pleistocene freshwater ecosystems in the Sierra de Atapuerca (northern Iberia) based on the Gran Dolina fish record. Palaeogeogr Palaeoclimatol Palaeoecol 574(2):11044
Boivin N, Fuller DQ, Dennell R, Allaby R, Petraglia MD (2013) Human dispersal across diverse environments of Asia during the Upper Pleistocene. Quatern Int 300:32–47
Bretzke K, Conard NJ (2017) Not just a crossroad: population dynamics and changing material culture in southwestern Asia during the Late Pleistocene. Curr Anthropol 58(17):449–462
Cáceres I, Bravo P, Esteban M, Expósito I, Saladié P (2002) Fresh and heated bones breakage. An experimental approach. In: De Renzi M, Pardo M, Belinchón M, Peñalver E, Montoya P, Márquez-Aliaga A (eds) Current topics on taphonomy and fossilization, Special Issue International Conference Taphos 2002. 3rd Meeting on Taphonomy and Fossilization: 471–479
Čermák S, Obuch J, Benda P (2006) Notes on the genus Ochotona in the Middle East (Lagomorpha: Ochotonidae). Lynx 37:51–66
Churchill SE, Franciscus RB, McKean-Peraza HA, Daniel JA, Warren BR (2009) Shanidar 3 Neanderthal rib puncture wound and Paleolithic weaponry. J Hum Evol 57:163–178
Conard NJ, Ghasidian E (2011) The Rostamian cultural group and the taxonomy of the Iranian Upper Paleolithic. In: Conard N, Drechslerr P, Morales A (eds) Between sand and sea: the archaeology and human ecology of southwestern Asia. Kerns Verlag, Tübingen, pp 33–52
Conard NJ, Zeidi M (2019) New research on the Palaeolithic occupation of Ghar-e Boof, Fars Province. Archaeol Spring 2:1–10
Conard NJ, Ghasidian E, Heydari S, Zeidi M (2006) Report on the 2005 survey of the Tübingen-Iranian Stone Age Research Project in the provinces of Esfahan, Fars and Kohgiluyeh-Boyerahmad. In: Azarnoush M (ed) Archaeological reports 5. ICAR, Tehran, pp 9–34
Conard NJ, Ghasidian E, Heydari S, Naderi R, Zeidi M (2007) The 2006 season of the Tübingen-Iranian Stone Age Research Project in the provinces of Fars and Markazi. In: Fazeli H (ed) 9th Annual Report of Archaeological Organisation. ICAR, Tehran, pp 43–67
Coşkun Y (2001) On distribution, morphology and biology of the mole vole, Ellobius lutescens Thomas, 1897 (Mammalia: Rodentia) in eastern Turkey. Zool Middle East 23:5–12
Coşkun Y, Ulutürk S (2003) Observations on the mole vole, Ellobius lutescens Thomas 1897, (Mammalia: Rodentia) in Turkey. Turk J Zool 27:81–87
Cowgill LW, Trinkaus E, Zeder MA (2007) Shanidar 10: a Middle Paleolithic immature distal lower limb from Shanidar Cave, Iraqi Kurdistan. J Hum Evol 53:213–233
Darvish J (2011) Morphological comparison of fourteen species of the genus Meriones Illiger, 1811 (Rodentia: Gerbillinae) from Asia and North Africa. Iran J Anim Biosystematic 7:49–74
Darvish J, Mohammadi Z, Mahmoudi A, Siahsarvie R (2014) Faunistic and taxonomic study of rodents from northwestern Iran. Iran J Anim Bisoyst 10:119–136
Delfino M (2004) The Middle Pleistocene herpetofauna of Valdemino Cave (Liguria, North-Western Italy). Herpetol J 14:113–128
Denys C (2017) Subfamily Deomynae, Gerbillinae, Ieimacomynae, Iophimynae species accounts. In: Wilson DE, Lacher TE, Mittermeier RA (eds) Handbook of the mammals of the world, vol 7. Rodents II. Lynx Edicions, Barcelona, pp 598–650
Dianat M, Darvish J, Cornett R, Aliabadian M, Nicolas V (2017) Evolutionary history of the Persian Jird, Meriones persicus, based on genetics, species distribution modelling and morphometric data. J Zool Syst Evol Res 55:29–45
Djamali M, de Beaulieu JL, Shah-Hosseini M, Andrieu-Ponel V, Ponel P, Amini A, Akhani H, Leroy SAG, Stevens L, Lahijani H, Brewer S (2008) A Late Pleistocene long pollen record from Lake Urmia, NW Iran. Quatern Res 69:413–420
Djamali M, Kürschner H, Akhani H, de Beaulieu JL, Amini A, Andrieu-Ponel V, Ponel P, Stevens L (2008) Palaeoecological significance of the spores of the liverwort Riella (Riellaceae) in a late Pleistocene long pollen record from the hypersaline Lake Urmia, NW Iran. Rev Palaeobot Palynol 152:66–73
Djamali M, Biglari F, Abdi K, Andrieu-Ponel V, de Beaulieu JL, Mashkour M, Ponel P (2011) Pollen analysis of coprolites from a late Pleistocene-Holocene cave deposit (Wezmeh Cave, west Iran): insights into the late Pleistocene and late Holocene vegetation and flora of the central Zagros Mountains. J Archaeol Sci 38:3394–3401
Escala MC, Miranda R (2002) Guía de Identificación de restos óseos de los ciprínidos presentes en España. Escamas, opérculos, cleitros y arcos faríngeos. Serie Zoología 28:1–239
Evans EMN, Van Couvering JAH, Andrews P (1981) Palaeoecology of Miocene sites in western Kenya. J Hum Evol 10:99–116
Fernández-Jalvo Y, Avery DM (2015) Pleistocene micromammals and their predators at Wonderwerk Cave, South Africa. Afr Archaeol Rev 32:752–791
Fernández-Jalvo Y, Andrews P, Denys C, Sesé C, Stoetzel E, Marin-Monfort D, Pesquero D (2016) Taphonomy for taxonomists: implications of predation in small mammal studies. Quat Sci Rev 139:138–157
Fiacconi M, Hunt CO (2015) Pollen taphonomy at Shanidar Cave (Kurdish Iraq): an initial evaluation. Rev Palaeobot Palynol 223:87–93
Field JS, Petraglia MD, Mirazón-Lahr M (2006) The southern dispersal hypothesis and the South Asian archaeological record: examination of dispersal routes through GIS analysis. J Anthropol Archaeol 26:88–108
Firouz E (2005) The complete fauna of Iran. Tauris & Co Ltd, London, p 322
Frontini R, Roselló-Izquierdo E, Morales-Muñiz A, Denys C, Guillaud E, Fernández-Jalvo Y, Pesquero-Fernández MD (2021) Compression and digestion as agents of vertebral deformation in Sciaenidae, Merlucidae and Gadidae remains: an experimental study to interpret archaeological assemblages. J Archaeol Method Theory. https://doi.org/10.1007/s10816-021-09527-5
Gharkheloo M, Kivanç E (2003) A study on the morphology, karyology and distribution of Ellobius Fischer, 1814 (Mammalia: Rodentia) in Iran. Turk J Zool 27(4):281–292
Ghasidian E (2014) The Early Upper Paleolithic occupation at Ghar-e Boof Cave: a reconstruction of cultural tradition in the southern Zagros Mountains of Iran. Kerns Verlag, Tübingen, p 304
Ghasidian E, Bretzke K, Conard NJ (2017) Excavations at Ghar-e Boof in the Fars Province of Iran and its bearing on models for the evolution of the upper Paleolithic in the Zagros Mountains. J Anthropol Archaeol 47:33–49
Ghasidian E, Heydari-Guran S, Mirazón-Lahr M (2019) Upper Paleolithic cultural diversity in the Iranian Zagros Mountains and the expansion of modern humans into Eurasia. J Hum Evol 132:101–118
Grayson DK (1984) Quantitative zooarchaeology. Academic Press, Orlando, p 202
Gromov IM, Erbajeva MA (1995) The mammals of Russia and adjacent territories: lagomorphs and rodents. Russian Academy of Sciences, Zoological Institute, St. Petersburg, p 522
Harrison DL, Bates PJJ (1991) The mammals of Arabia. Harrison Zoological Museum, Sevenoaks, p 354
Hashemi N, Darvish J, Mashkour M, Biglari F (2006) Rodents and lagomorphs remains from late Pleistocene and early Holocene caves and rockshelter sites in the Zagros region, Iran. Iran J Anim Biosyst 2(1):25–33
Heydari M, Mahdavi A (2009) The survey of plant species diversity and richness between ecological species groups (Zagros Ecosystem, Ilam). J Appl Sci 9(4):745–751
Heydari M, Guérin G, Zeidi M, Conard NJ (2021) Bayesian luminescence dating at Ghar-e Boof, Iran, provides a new chronology for Middle and Upper Paleolithic in the southern Zagros. J Hum Evol 151:102926
Heydari-Guran S (2014) Paleolithic landscapes of Iran. BAR international series, Oxford, p 296
Heydari-Guran S, Ghasidian E (2020) Late Pleistocene hominin settlement patterns and population dynamics in the Zagros Mountains: Kermanshah region. Archaeol Res Asia 21:100161
Heydari-Guran S, Benazzi S, Talamo S, Ghasidian E, Hariri N, Oxilia G, Asiabani S, Azizi F, Naderi R, Safaierad HJJ, Foley RA, Mirazón-Lahr M (2021a) The discovery of an in situ Neanderthal remain in the Bawa Yawan Rockshelter, West-Central Zagros Mountains, Kermanshah. Plos ONE 16(8):e0253708
Heydari-Guran S, Douka K, Higham T, Münzel SC, Deckers K, Hourshid S, Nadari R, Asiabani S, Ghasidian E (2021b) Early Upper Palaeolithic occupation at Gelimgoush Cave, Kermanshah; West-Central Zagros mountains of Iran. J Archaeol Sci Report 38:103050
Hole F, Flannery KV (1967) The prehistory of southwestern Iran: a preliminary report. Proc Prehist Soc 33:147–206
Ikeda J (1979) Preliminary report of an archaeological survey in Arsanjan Area, Fars Province, Iran, 1977. Kyoto University, Kyoto
Kaboli M, Aliabadian M, Tohidifar M, Hashemi A, Musavi SB, Roselaar CC (2016) Atlas of birds of Iran. Department of Environment of Iran, Tehran, p 617
Kandel AW, Gasparyan B, Allué E, Bigga G, Bruch AA, Cullen VL, Frahm E, Ghukasyan R, Gruwier B, Jabbour F, Miller CE, Taller A, Vardazaryan V, Vasilyan D, Weissbrod L (2017) The earliest evidence for Upper Paleolithic occupation in the Armenian Highlands at Aghitu-3 Cave. J Hum Evol 110:37–68
Karami M, Ghadirian T, Faizolahi K (2016) The atlas of mammals of Iran. Iran Department of the Environment, Tehran, p 292
Kehl M, Frechen M, Skowronek A (2005) Paleosols derived from loess and loess-like sediments in the Basin of Persepolis, Southern Iran. Quatern Int 140–141:135–149
Keivany Y, Nasri M, Abbasi K, Abdoli A (2016) Atlas of inland water fishes of Iran. In: Daneshgahi J, Branch K (eds) Biodiversity and Wildlife Division of Iranian Department of Environment, Teheran, p 186
Kessler JE (2015) Osteological guide of songbirds from Central Europe. Ornis Hungarica 23(2):62–155
Kryštufek B, Vohralík V (2005) Mammals of Turkey and Cyprus. Rodentia I: Sciuridae, Dipodidae, Gliridae, Arvicolinae. University of Primorska, Science and Research Centre Koper, Koper, p 292. https://doi.org/10.1644/10-MAMM-R-221.1
Kryštufek B, Vohralík V (2009) Mammals of Turkey and Cyprus, Rodentia II: Cricetinae, Muridae, Spalacidae, Calomyscidae, Capromyidae, Hystricidae. University of Primorska, Science and Research Centre Koper, Koper, Castoridae, p 374
Laplana C, Sevilla P, Arsuaga JL, Arriaza MC, Baquedano E, Pérez-González A, López-Martínez N (2015) How far into Europe did pikas (Lagomorpha: Ochotonidae) go during the Pleistocene? New Evidence from Central Iberia. PlosOne 10(11):e0140513
Lapparent FD, Bailon S, Augé ML, Rage JC (2020) Amphibians and reptiles from the Neogene of Afghanistan. Plazi.org taxonomic treatments database. Geodiversitas. https://doi.org/10.5252/geodiversitas2020v42a22
Lloveras L, Cosso A, Solé J, Claramunt-López B, Nadal J (2017) Taphonomic signature of Golden eagles (Aquila chrysaetos) on bone prey remains. Hist Biol. https://doi.org/10.1080/08912963.2017.1319830
Lloveras L, Moreno-García M, Nadal J (2009) The eagle owl (Bubo bubo) as a leporid remains accumulator. Taphonomic analysis of modern rabbit remains recovered from nests of this predator. Int J J Osteoarchaeol 19:573–592
Lloveras L, Moreno-García M, Nadal J (2009) Butchery, cooking and human consumption marks on rabbit (Oryctolagus cuniculus) bones: an experimental study. Journal of Taphonomy 7:179–201
López-García JM (2011) Los micromamíferos del Pleistoceno superior de la Península Ibérica: Evolución de la diversidad taxonómica y cambios paleoambientales y paleoclimáticos. Editorial Académica Española, Tarragona, p 416
Lyman RL (2008) Quantitative paleozoology. In: Cambridge Manuals in Archaeology. Cambridge University Press, New York, p 388
Mamkhair H, Samara FF, Shehab AH (2007) Morphological characteristics of the Libyan Jird, Meriones libycus Lichtenstein, 1823 (Rodentia: Gerbillinae), in Syria. Zool Middle East 42(1):37–46. https://doi.org/10.1080/09397140.2007.10638244
Markert D (1978) Erstmalige verwengund quartärer reptilreste bei paleökologischen rekonstruktionsversuchen am beispiel des oberen Donauraumes um die wende Pleistozän/Holozän. Archaeologica Venatoria, Tübingen, p 97
Mashkour M, Radu V, Mohaseb A, Hashemi N, Otte M, Shidrang S (2009a) The Upper Paleolithic faunal remains from Yafteh cave (central Zagros) 2005 campaign. A preliminary study. In: Otte M, Biglari F, Jaubert J (eds) Le Paleolithique D’Iran. Archaeopress, Oxford, pp 73–85
Mashkour M, Moncho H, Trinkaus E, Reyss JL, Biglari F, Bailon S, Heydari S, Abdi K (2009b) Carnivores and their prey in the Wezmeh Cave (Kermanshah, Iran): a late refuge in the Zagros. Int J Osteoarchaol 19:678–694
Mata-González M, Starkovich BM, Zeidi M, Conard NJ (2022) New zooarchaeological perspectives on the early Upper Paleolithic Rostamian sequence of Ghar-e Boof (southern Zagros Mountains). Quatern Sci Rev 279:107350
Medina ME, Teta P, Rivero D (2012) Burning damage and small-mammal human consumption in Quebrada del Real 1 (Córdoba, Argentina): an experimental approach. J Archaeol Sci 39:737–743
Mostafavi H, Reza-Merhabian A, Teimori A. Shafizade-Moghadam H. Kambouzia J (2021) The ecology and modelling of the freshwater ecosystems in Iran. In: Jawad LA (Ed.), Tigris and Euphrates Rivers: their environment from headwaters to mouth. Aquat Ecol Ser 11:1143–1200
Mozaffari O, Kamali K, Fahimi H (2016) The atlas of reptiles of Iran. Department of Environment of Iran, Tehran, p 361
Musser GG, Carleton MD (2005) Superfamily Muroidea. In: Wilson DE, Reeder DM (eds) Mammal species of the world: a taxonomic and geographic reference. Johns Hopkins University Press, pp 894–1531
Nicholson RA (1993) An investigation into the effects on fish bone on passage through the human gut: some experiments and comparisons with archaeological material. Circaea 10:38–51
Noroozi J, Talebi A, Doostmohammadi M, Bagheri A (2020) The Zagros Mountain range. In: Noroozi J (eds) Plant biogeography and vegetation of high mountains of Central and South-West Asia. Plant and Vegetation. 17:185–214
Oppenheimer S (2009) The great arc of dispersal of modern humans: Africa to Australia. Quat Int 202:2–13
Otte M, Biglari F, Flas D, Shidrang S, Zwyns N, Mashkour M, Naderi R, Mohaseb A, Hashemi N, Darvis J, Radu V (2007) The Aurignacian in the Zagros region: new research at Yafteh Cave, Lorestan. Iran Antiquity 81:82–96
Pavlinov I (2008) A review of phylogeny and classification of Gerbillinae (Mammalia: Rodentia). Moscow University Publishing, Moscow, p 74
Pavlinov I, Dubrovsky YA, Rossolimo OL, Potapova EG (1990) Gerbils of the world. Nauka, Moscow, p 363
Perkins D (1964) Prehistoric fauna from Shanidar, Iraq. Science 144(3226):1565–1566
Piperno M (1972) Jahrom, a Middle Palaeolithic site in Fars, Iran. East West 22:183–197
Piperno M (1974) Upper Paleolithic caves in southern Iran, preliminary report. East West 24:9–13
Pomeroy E, Mirazón-Lahr M, Crivellaro F, Farr L, Reynolds T, Hunt CO, Barker G (2017) Newly discovered Neanderthal remains from Shanidar Cave, Iraqi Kurdistan, and their attribution to Shanidar 5. J Hum Evol 111:102–118
Pomeroy E, Bennett P, Hunt CO, Reynolds T, Farr L, Frouin M, Holman J, Lane R, French C, Barker G (2020) New Neanderthal remains associated with the ‘flower burial’ at Shanidar Cave. Antiquity 94(373):11–26
Rastegar-Pouyani N, Torki F (2007) Spermatogenesis without spermiogenesis in Laudakia caucasia (Reptilia: Agamidae): the role of hibernation. Iran J Anim Biosyst 3(1):37–42
Redpath S, Clarke R, Madders M, Thirgood SJ (2009) Assessing raptor diet: comparing pellets, prey remains, and observational data at Hen Harrier nests. Condor 103:184–188
Reed CA, Braidwood RJ (1960) Toward the reconstruction of the environmental sequence of northern Iraq. Stud Anc Oriental Civiliz 31:169–174
Rey-Rodríguez I, López-García JM, Blain HA, Stoetzel E, Denys C, Fernández-García M, Tumung L, Ollé BB (2020) Exploring the landscape and climatic conditions of Neanderthals and anatomically modern humans in the Middle East: the rodent assemblage from the late Pleistocene of Kadar Cave (Khorramabad Valley, Iran). Quatern Sci Rev 236(2):106278
Rey-Rodríguez I, Arnaud J, López-García JM, Stoetzel E, Denys C, Cornette R, Bazgir B (2021) Distinguish between three modern Ellobius species (Rodentia, Mammalia) and identification of fossil Ellobius from Kaldar Cave (Iran) using geometric morphometric analyses of the first lower molar. Palaentol Electron 24(1):01. https://doi.org/10.26879/1122palaeo-electronica.org/content/2021/3265-ellobius-and-gmm
Rhodes SE, Walker MJ, López-Jiménez A, López-Martínez M, Hber-Uriarte M, Fernánez-Jalvo Y, Chazan M (2016) Fire in the Early Palaeolithic: Evidence from burnt small mammal bones at Cueva Negra del Estrecho del Río Quñipar, Murcia, Spain. J Archaeol Sci Rep 9:427–436
Rosenberg MS (1979) Eshkaft-e Gavi (The Malyan Project), Iran. J Br Inst Persian Stud 17:148–149
Rosenberg MS (1985) Report on the 1978 sondage at Eshkaft-e Gavi, Iran. J Br Inst Persian Stud 23:51–62
Rosenberg MS (1988) Paleolithic settlement pattern in the Marv Dasht, Fars Province, Iran. Unpublished Ph.D. dissertation. University of Pennsylvania, p 798
Scott JE, Marean CW (2009) Paleolithic hominin remains from Eshkaft-e Gavi (southern Zagros Mountains, Iran): description, affinities, and evidence for butchery. J Hum Evol 57:248–259
Shipman P, Foster G, Shoeninger M (1984) Burnt bones and teeth: an experimental study of color, morphology, crystal structure and shrinkage. J Archaeol Sci 11(4):307–325
Shoaee MJ, Vahdati-Nasab H, Petraglia MD (2021) The Paleolithic of the Iranian Plateau: hominin occupation history and implications for human dispersals across southern Asia. J Anthropol Archaeol 62:101292
Siahsarvie R, Darvish J (2008) Geometric morphometric analysis of Iranian wood mice of the genus Apodemus (Rodentia, Muridae). Mammalia 72:109–115
Solecki, RS (1958) The Baradostian industry and the Upper Palaeolithic in the near east. Unpublished PhD dissertation. Columbia University, p 174
Solecki RS (1963) Prehistory in Shanidar Valley northern Iraq. Science 139:179–193
Stiner MC, Kuhn SL, Weiner S, Bar-Yusef O (1995) Differential burning, recrystallization and fragmentation of archaeological bone. J Archaeol Sci 22:223–237
Stoetzel E, Cornette R, Lalis A, Nicolas V, Cucchi T, Denys C (2017) Systematics and evolution of the Meriones shawii/grandis complex (Rodentia, Gerbillinae) during the Late Quaternary in northwestern Africa: exploring the role of the environmental and anthropogenic changes. Quat Scien Rev 164:199–216
Tesakov AS (2016) Early middle Pleistocene Ellobius (Rodentia, Cricetidae, Arvicolinae) from Armenia. Russian Journal of Theriology 15(2):151–158
Tilby E, Miracle P, Barker G (2022) The microvertebrates of Shanidar Cave: preliminary taphonomic findings. Quaternary 5:4. https://doi.org/10.3390/quat5010004
Trinkaus E (1983) The Shanidar Neanderthals. Academic Press, New York, p 502
Trinkaus E (2018) Pleistocene human remains from Iran. In: Biglari F, Shidrang S, Mashkour M (eds) The Pleistocene archaeology of the Iranian Plateau, Iraq and the Caucasus. National Museum of Iran, Tehran, pp 53–63
Trinkaus E, Biglari F (2006) Middle Paleolithic human remains from Bisitun Cave. Iran Paléorient 32:105–111
Trinkaus E, Villotte S (2017) External auditory exostoses and hearing loss in the Shanidar 1 Neanderthal. PLoS ONE 12(10):e0186684
Trinkaus E, Biglari F, Mashkour M, Monchot H, Reyss JL, Rougier H, Heydari S, Abdi K (2008) Late Pleistocene human remains from Wezmeh Cave, western Iran. Am J Phys Anthropol 135:371–378
Tsanova T (2013) The beginning of the Upper Paleolithic in the Iranian Zagros. A taphonomic approach and techno-economic comparison of Early Baradostian assemblages from Warwasi and Yafteh (Iran). J Hum Evol 65:39–64
Turnbull F (1975) The mammalian fauna of Warwasi rock shelter. West-Central Iran Fieldiana 33(8):141–155
Turnbull PF, Reed CA (1974) The fauna from the terminal Pleistocene of Palegawra Cave, a Zarzian occupation site in northeastern Iraq. Fieldiana. 63(3):81–146
Vahdati-Nasab H, Clark GA, Torkamandi S (2013) Late Pleistocene dispersal corridors across the Iranian Plateau: a case study from Mirak, a Middle Paleolithic site on the northern edge of the Iranian Central Desert (Dasht-e Kavir). Quatern Int 300:267–281
van Zeist W, Bottema S (1977) Palynological investigations in western Iran. Palaeohistoria 19:19–85
Walker MJ, Anesin D, Angelucci DE, Avilés-Fernández A, Berna F, Buitrago-López A, Fernández-Jalvo Y, Haber-Uriarte M, López-Jiménez A, López-Martínez M, Martín-Lerma I, Ortega-Rodrigáñez J, Polo-Camacho JL, Rhodes SE, Richter D, Rodríguez-Estrella T, Schwenninger JL, Skinner AR (2015) Findings, context, and significance of combustión at the late Early Pleistocene Palaeolithic site of Cueva Negra del Estrecho del Río Quípar (Caravaca de la Cruz, Spain). Antiquity 90(351):571–589
Wasylikowa K (2005) Palaeocology of Lake Zeribar, Iran, in the Pleniglacial, Lateglacial and Holocene, reconstructed from plant macrofossils. Holocene 15(5):720–735
Wójcik J (2002) The comparative osteology of the humerus in European thrushes (Aves: Turdus) including a comparison with other similarly sized genera of passerine birds- preliminary results. Acta Zool Cracov 45:369–381
Yiğit N, Çolak E, Sözen M, Özkurt Ş (2003) A study on the geographical distribution along with habitat aspects of rodent species in Turkey. Bonn Zool Bull 50:355–368
Yousefi-Siahkalroodi S, Saeedi H, Sadat-Behfar M, Fallahi R, Izadian M (2013) Atlas of amphibians of Iran. Department of Environment of Iran, Tehran
Yusefi GH (2002) Taxonomic study of the Brandt’s hedgehog Paraechinus hypomelas (three islands and one mainland populations). MSc thesis, Shahid Beheshti Universit
Yusefi GH, Kiabi BH, Khalatbari L, Faizolahi K, Monteiro NM (2016) Morphological analysis of Brandt’s hedgehog (Paraechinus hypomelas) reflects the isolation history of Persian Gulf islands and has implications for taxonomy. Biol J Lin Soc 119(2):497–510
Zanolli C, Biglari F, Mashkour M, Abdi K, Monchot H, Debue K, Mazurier A, Bayle P, Leyur ML, Rougier H, Trinkaus E, Macchiarelli R (2019) A Neanderthal from the Central Western Zagros, Iran. Structural reassessment of the Wezmeh 1 maxillary premolar. J Human Evol 135:102643
Zeidi M, Conard NJ (2019) The fourth season of excavation at Ghar-e Boof, Rostam County, Fars Province. In: Shirazi RA (ed) 16th Annual Symposium of the Iranian Archaeology. Iranian Center for Archaeological Research, Tehran, pp 234–238 ((in Farsi))
Zeidi M, McCall B, Khosrowzadeh A (2006) Survey of Dasht-e Rostam-e Yek and Dasht-e Rostam-e Do. In: Potts DT, Roustaei K (eds) The Mamasani Archaeological Project Stage One: a report on the first two seasons of the ICAR – University of Sydney Expedition to the Mamasani District, Fars Province, Iran. Iranian Center for Archaeological research, Tehran, pp 147–168
Acknowledgements
Many people deserve thanks for their contributions in this study. We are grateful to the editors and the anonymous reviewers for providing constructive feedback that improved our manuscript. The authors would like to thank the Iranian Center for Archaeological Research (ICAR) and the Research Institute of Cultural Heritage and Tourism (RICHT). We are thankful to all members of the TISARP team for their contribution during the excavation, water-screening, and management of the recovered material, especially Saman Hamzavi. We also express our gratitude to Dr. Hugues-Alexandre Blain, Dr. Elisa Luzi, Dr. Iván Rey-Rodríguez, and Dr. Yusefi for their comments and suggestions.
Funding
Open Access funding enabled and organized by Projekt DEAL. MMG is the beneficiary of a research grant awarded to NJC from the Ministerium für Wissenschaft, Forschung und Kunst Baden-Württemberg as part of the Evolution of Cultural Modernity Project at the University of Tübingen. Excavations at Ghar-e Boof were funded by the Deutsche Forschungsgemeinschaft (DFG) grant CO 226/30–1. We would like to thank the ROCEEH, SHEP, and members of the University of Tübingen who supported us during our work.
Author information
Authors and Affiliations
Contributions
A. Blanco-Lapaz and M. Mata-González analyzed the microvertebrate assemblage. B.M. Starkovich, N.J. Conard, and M. Zeidi provided archeological and geological context. The manuscript was written by A. Blanco-Lapaz and M. Mata-González. All authors contributed to all sections of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This article is part of the Topical Collection on Microvertebrate Studies in Archaeological Contexts: Middle Paleolithic to early Holocene past environments.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Blanco-Lapaz, A., Mata-González, M., Starkovich, B.M. et al. Late Pleistocene environments in the southern Zagros of Iran and their implications for human evolution. Archaeol Anthropol Sci 14, 161 (2022). https://doi.org/10.1007/s12520-022-01615-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12520-022-01615-1