
Abstract
Relatively little is known about the direct influence of acid rain (AR) on pro- and antioxidative changes in plant cells. Intercompartmental differences between cytosol and mitochondria have not been studied before. Aboveground parts of plants were treated with different pH variants of AR and oxidative damages (lipid peroxidation) as well as antioxidative enzyme activities (superoxide dismutase, SOD; ascorbate peroxidase, APx; glutathione peroxidase, GSH-Px) in the cytosolic and mitochondrial fractions were examined. The character of changes in antioxidative enzyme activities and of oxidative damages was closely connected with the cell compartment as well as with pH and time after treatment. The activity of both APx and GSH-Px increased more intensively in cytosol. Contrastingly, strong induction of lipid peroxide formation was observed in the mitochondrial fraction. In both cell compartments SOD activity did not change significantly. The results suggest that cucumber mitochondria are more susceptible to oxidative damage caused by AR than cytosol. Antioxidative defence of cytosol appeared to provide sufficient protection against the oxidative stress imposed by AR.
Similar content being viewed by others


Introduction
Anthropogenic activities resulting in increased concentrations of sulphur dioxide (SO2) and nitrogen oxides (NOx) in the atmosphere are responsible for acid rain (AR). In spite of the restrictions on SO2 emission introduced in many countries, NOx contamination, which originates from motor vehicles and other combustion processes carried out at high temperature, still remains at a high level (Gimeno et al. 2001; Rosborg et al. 2003; Tran et al. 2011). AR is a major environmental threat for many regions in Europe, Asia and North America and especially concerns developing countries (Kulshrestha et al. 2003). In most regions its pH values range from 4.4 to 2.3 (Shan et al. 1996); however, Gravano reported AR value as low as 1.4 on San Rossore Estate (Pisa, Italy) (Gravano et al. 1999). Deleterious effects of AR on plant development have been studied for almost three decades and presented in several excellent articles (Ferenbaugh 1976; Haines et al. 1980; Neufeld et al. 1985). However, most of the experimental observations reported in the literature concern mainly changes in plant growth as well as histology and ultrastructure (Evans et al. 1984; Stoyanova 1997/98; Stoyanova and Velikova 1997/98; Sant’Anna-Santos et al. 2006). In recent years biochemical studies concerning photosynthesis intensity as well as pro- and antioxidative changes in plant tissue after AR application have not been numerous (Yu et al. 2002; Gabara et al. 2003; Wyrwicka and Skłodowska 2006; Choi et al. 2010). Our previous research (Gabara et al. 2003; Wyrwicka and Skłodowska 2006) showed that mitochondria were extremely susceptible to AR stress. After exposure of tomato plants (Lycopersicon esculentum Mill.) to AR, disturbances were observed only in 13 % of chloroplasts, but the population of deformed mitochondria increased up to 95 %. Due to that, we chose mitochondria for our biochemical research.
Mitochondria are an important cell compartment because the electron transport chain functions there and it harbours electrons with sufficient free energy to directly reduce molecular oxygen. It is considered the unavoidable primary source of mitochondrial reactive oxygen species (ROS) production, a necessary accompaniment to aerobic respiration (Rhoads and Subbaiah 2007). Formation of mitochondria-derived ROS takes place under normal respiratory conditions but can be enhanced in response to exposition to biotic and abiotic stress factors. Mitochondria and cells in general possess numerous ROS defence systems. Both enzymatic and nonenzymatic mechanisms have evolved to protect this cell compartment from oxygen toxicity. These include antioxidants such as ascorbate, glutathione, as well as ROS-scavenging enzymes such as superoxide dismutase (SOD; EC 1.15.1.1), ascorbate peroxidase (APx; EC 1.11.1.1) and glutathione peroxidase (GSH-Px; EC 1.11.1.9) (Willekens et al. 1994; Miszalski et al. 2001; Mittler 2002). It is suggested that the true source of oxidative stress is not the ROS generation per se but spatiotemporal imbalance of ROS production and detoxification (Andreyev et al. 2005). Until now the capacity of mitochondrial ROS defence is unknown.
Several lines of evidence indicate the importance of mitochondria in integration of cell signals. They point to a mitochondrion as a central player in the reception and amplification of cell-death signals. Levels of cellular messengers such as calcium, changes in cellular pH or changes in the levels of metabolites that reflect the energy status of the cell (ATP, ADP, NADH+H+, NADPH+H+) as well ROS concentration or carbon status can be signals for mitochondrion response (Lam et al. 2001).
Activities of the antioxidative enzymes such as SOD, APx and GSH-Px were investigated to compare the antioxidant systems operating in cytosol and mitochondria isolated from cucumber (Cucumis sativus L.) leaves subjected to different intensity of AR stress. In addition, to characterise the range of lipid oxidative damages, the concentration of 2-thiobarbituric acid reactive substances (TBARS) was measured. To the best of our knowledge, changes in pro- and antioxidative systems in plant cytosol and mitochondria caused by AR treatment have not been studied simultaneously before.
Materials and methods
Plant material
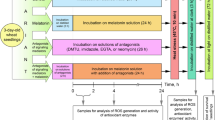
Cucumber seeds (Cucumis sativus L.) cv “Cezar” were germinated in Petri dishes for 5 days. Seedlings were planted into soil-containing plastic pots. Plants were grown in a controlled environment growth chamber under the temperature of 23 ± 0.5 °C with 16 h light/8 h dark photoperiod at light intensity of 150 μmol m−2 s−1 photon flux density and 60 % relative humidity. 5-week-old plants with four fully expanded leaves were used in the experiment. All biochemical analyses were carried out on the second, third and fourth leaves from the control and treated plants. The leaves were harvested on the first, third, fifth and seventh day after a single AR treatment. In each experiment the material was harvested from two plants, mixed and was treated as one sample.
AR treatment
Each cucumber plant was sprayed once with 10 cm3 of simulated AR or distilled water. The following variants were used:
-
1.
AR pH 4.4 containing 0.222 mM (NH4 +), 3.6 μM (NO2 −), 0.129 mM (NO3 −), 0.220 mM (Na+), 0.111 mM (SO4 2−), 0.750 mM (Cl−).
-
2.
AR pH 3.0 containing 0.222 mM (NH4 +), 2.4 μM (NO2 −), 0.353 mM (NO3 −), 0.190 mM (Na+), 0.111 mM (SO4 2−), 1.831 mM (Cl−).
-
3.
AR pH 1.8 containing 0.222 mM (NH4 +), 1.74 μM (NO2 −), 0.129 mM (NO3 −), 0.217 mM (Na+), 39.35 mM (SO4 2−), 0.219 mM (Cl−).
The sizes of minute droplets that appeared at the surface of leaves directly after spraying ranged between 0.02 and 0.005 cm3. All plants were sprayed in the middle of the day cycle. To avoid soil acidification it was covered with plastic foil during spraying. The control plants were sprayed with distilled water (pH 6.0).
Preparation of cytosolic fraction
Whole leaves were homogenised (1:6 w/v) in an ice-cold mortar using 50 mM sodium phosphate buffer pH 7.0 containing 0.5 M NaCl, 1 mM EDTA and 1 mM sodium ascorbate. After centrifugation (20,000×g for 15 min) the supernatant was used for determination of SOD, APx and GSH-Px activities as well as content of TBARS.
Isolation of leaf mitochondria
Mitochondria were isolated from the leaves by differential centrifugation method of Warrilow and Hawkesford (1998). The leaves (5 g) without the main midribs were cut into pieces, chilled in an ice bath for 20 min and homogenized two times, for 10 and 35 s in a blender in an ice-cold homogenisation medium (1:12 w/v) containing 50 mM TRIS–HCl, 0.33 M D-mannitol, 2.5 mM EDTA, 2.5 mM sodium ascorbate and 2 mM MgCl2 (pH 8.1). The homogenates were filtered through four layers of Miracloth (crude extract) and then centrifuged at 1,500×g for 3 min to remove chloroplasts. The supernatant was further centrifuged at 8,200×g for 10 min and the obtained supernatant was subsequently called “supernatant 8,200×g”. To obtain pure mitochondrial fraction the pellet (mitochondria and peroxisomes) was gently resuspended twice in a washing medium containing: 50 mM TRIS–HCl, 2 mM MgCl2 and 2 mM sodium ascorbate (pH 8.1) and centrifuged at 8,200×g for 10 min. The resulting pellet was the final mitochondrial fraction. All operations were carried out at 0–4 °C.
Organelle intactness and enzyme latency
The quality of mitochondrial preparation was monitored by measuring marker enzyme activities and chlorophyll content. All determinations were conducted in the cytosolic fraction in the supernatant obtained after centrifugation of the final mitochondrial fraction (isotonic medium) and in the mitochondrial extract achieved by homogenisation and centrifugation of the final mitochondrial fraction. To assess contamination with other organelles, the activities of the following enzymes were measured: fumarase (EC 4.2.1.2) (Hatch 1978) and glucose-6-phosphate dehydrogenase (EC 1.1.1.49) (using Sigma-Aldrich diagnostic kit; Steinheim, Germany), which are markers for mitochondria and cytosol respectively as well as catalase (EC 1.11.1.6) (Dhindsa et al. 1981) and hydroxypuryvate reductase (EC 1.1.1.29) (Tolbert 1971) which are specific enzymes for peroxysomes. Moreover, chlorophyll concentration (Porra et al. 1989),which is a characteristic compound of chloroplasts, was assessed.
The integrity of the outer mitochondrial membrane was estimated from the activity of fumarase (Hatch 1978), succinate:cytochrome c oxidoreductase (EC 1.3.99.1) and NADH+H+:cytochrome c oxidoreductase (EC 1.6.99.3) in the presence or absence of antimycyn A as described by Douce et al. (1972). The activities of both enzymes were assayed for the mitochondria in the isotonic medium and the mitochondrial extract. Organelle intactness was calculated according to the formula of Burgess et al. (1985) (Kuźniak and Skłodowska 2004); [100 − (activity associated with intact organelle/activity associated with permeabilized organelle) × 100].
Preparation of enzyme extracts from mitochondrial fraction
The final mitochondrial fraction was homogenised in an ice-cold mortar using 50 mM sodium phosphate buffer pH 7.0 containing 0.5 M NaCl, 1 mM EDTA and 1 mM sodium ascorbate. After centrifugation (20,000×g for 15 min) the supernatant (mitochondrial extract) was used for determination of SOD, APx and GSH-Px activities. To determine TBARS concentration homogenised mitochondrial pellet was taken to analyses before centrifugation.
Enzyme assay
Total SOD activity was determined by the method of Minami and Yoshikawa (1979) with 50 mM TRIS-cacodylic buffer pH 8.2 containing 0.1 mM EDTA, 1.4 % (v/v) Triton X-100, 0.055 μM NBT, 16 μM pyrogallol and enzyme extract. The concentration of reduced form of NBT was measured at 540 nm. The activity unit (50 % inhibition) was defined according to McCord and Fridovich (1969). SOD activity was expressed in μg enzyme mg−1 protein.
APx activity was assayed following the oxidation of ascorbate to dehydroascorbate at 265 nm (ε = 13.7 mM−1 cm−1) by a method of Nakano and Asada (1981) modified by Kuźniak and Skłodowska (1999). The assay mixture contained 50 mM sodium phosphate buffer pH 7.0, 0.25 mM sodium ascorbate, 25 μM H2O2 and enzyme extract (5–10 μg protein). Addition of H2O2 started the reaction. Rates were corrected for the nonenzymatic oxidation of ascorbate by the inclusion of reaction mixture without enzyme extract. Enzyme activity was expressed in μmol ascorbate min−1 mg−1 protein.
To detect GSH-Px activity, the method of Hopkins and Tudhope (1973) with t-butyl hydroperoxide as a substrate was used. The reaction mixture comprised 50 mM potassium phosphate buffer pH 7.0, 2 mM EDTA, 0.28 μM NADPH+H+, 0.13 μM GSH, 0.16 U GR, 0.073 μM t-butyl hydroperoxide and enzyme extract (50 μg protein). GSH-Px activity was expressed in μmol NADPH+H+ min−1 mg−1 protein.
Determination of lipid peroxidation
The concentration of lipid peroxides was estimated spectrofluorometrically according to Yagi (1982) by measuring the content of 2-thiobarbituric acid reactive substances (TBARS). The concentration of lipid peroxides was calculated in terms of 1,1,3,3-tetraethoxypropane which was used as a standard and expressed in nmol mg−1 protein.
Other determinations
The protein content was determined by Bradford’s method (1976) with standard curves prepared using bovine serum albumine.
All assays were performed spectrophotometrically (UNICAM UV 300 UV–visible spectrometer) at 25 °C.
Statistical analysis
All measurements were performed in two replicates in four to five independent experiments (n = 8–10). Sample variability was calculated as the standard deviation (SD) of the means. Values of particular parameters were given as percentage of control. The significance of differences between mean values was determined by a nonparametric Mann–Whitney Rank Sum Test. Differences at P < 0.05 were considered significant. All statistical estimates were performed using the STATISTICA 10 software.
Results
Some visible symptoms were observed already on the first day after a single treatment with AR pH 3.0 and 1.8 (Fig. 1). However, at pH 3.0 small brown spots appeared after 24 h only on the oldest leaves and white greater ones—after 48 h. Such symptoms were also observed after pH 1.8 treatment but they were more intensive and appeared on all leaves. At the latter case bleaching of the leaf surface within the ribs was observed. During the first hours after AR pH 1.8 spraying the plant leaves had reduced turgor, but it was recovered 24 h after treatment.

Visible injuries in cucumber leaves after the exposure to AR pH 4.4; pH 3.0 and pH 1.8
In the purified mitochondria obtained from both the control and AR-treated leaves, the activity of glucose-6-phosphate dehydrogenase was negligible indicating high purity of the collected fraction (Table 1). The mitochondrial extract did not contain chlorophyll. The activities of catalase and hydroxypyruvate reductase indicated that the isolated mitochondria could be slightly contaminated with peroxisomes (Table 1).
The calculated latency values for fumarase, succinate:cytochrome c oxidoreductase and NADH+H+:cytochrome c oxidoreductase were 98, 87 and 90 %, respectively, and indicated high integrity of outer mitochondrial membrane (Table 2).
The character of changes of antioxidative enzyme activities was closely connected with the cell compartment as well as with pH of the applied AR and time after treatment. Significant differences in the total SOD activity between AR-treated and the control plants were noticed only in the cytosolic fraction (Fig. 2). Increase up to 150 % (P < 0.05) of the control value was observed in this cell compartment on the third day of experiment after exposure to AR pH 1.8. In the cytosolic fraction significant differences in the total SOD activities were found also between different experimental variants. In mitochondrial fraction gradual lowering of SOD activity was observed in each variant of AR-treated plants; on the fifth and seventh day it was lower than that in the control. However, this tendency was not statistically important. In the control the mean value of SOD activity in the cytosolic fraction was 32 μg SOD mg−1 protein and in the mitochondrial fraction—251 μg SOD mg−1 protein.

Influence of AR stress on SOD activity in the cucumber cytosol and mitochondria. (Plain box) AR pH 4.4; (wavy box) AR pH 3.0; (checked box) AR pH 1.8. Symbols *, ** and *** indicate values that differ significantly from control at P < 0.05, P < 0.01 and P < 0.001, respectively. Symbols (a), (aa) and (aaa) indicate values that differ significantly between variants pH 4.4 and pH 3.0; symbols (b), (bb) and (bbb) indicate values that differ significantly between variants pH 4.4 and pH 1.8; symbols (c), (cc) and (ccc) indicate values that differ significantly between variants pH 3.0 and pH 1.8 at P < 0.05, P < 0.01 and P < 0.001, respectively. SOD activity expressed as percentage of control value
On the first day, exposure to AR pH 1.8 decreased mitochondrial APx activity to 68 % (P < 0.01) of the control (Fig. 3). On the following days increase in APx activity in pH 1.8 treated plants was significant both in the cytosolic and mitochondrial fractions; however, it was more intensive in the former one. On the third, fifth and seventh day after AR treatment the activity of cytosolic APx increased up to 350 (P < 0.01), 267 (P < 0.001) and 299 % (P < 0.001) of the control and that of mitochondrial APx up to 155 (P < 0.05), 175 (P < 0.01) and 155 % (P < 0.05) of the control, respectively (Fig. 3). On the first day AR pH 3.0 significantly increased cytosolic APx activity (to 176 % of the control value, P < 0.05) but caused few changes in mitochondria where the statistically important rise was observed only on the third day of experiment and reached 137 % (P < 0.05) of the control. Additionally, on the seventh day of experiment in the mitochondrial fraction significant decreases in APx activity were observed after the application of AR: for pH 3.0–72 % (P < 0.05) and for pH 4.4–77 % (P < 0.05) of the control value. In the control plants the mean value of cytosolic APx activity was 0.463 μmol ascorbate min−1 mg−1 protein and of mitochondrial, 0.651 μmol ascorbate min−1 mg−1 protein.

Influence of AR stress on APx activity in the cucumber cytosol and mitochondria. APx activity expressed as percentage of control value. For further explanations, see Fig. 2
Changes of GSH-Px activity differed between the cytosolic and mitochondrial fractions (Fig. 4). However, significant differences were observed only in AR pH 1.8-treated plants. In contrast to the mitochondrial GSH-Px activity which in all pH variants was similar to the control value, the activity of the cytosolic enzyme isolated from the leaves treated with AR pH 1.8 was significantly increased to 207 (P < 0.05), 128 (P < 0.05) and 169 % (P < 0.01) of the control on the third, fifth and seventh day, respectively. Differences in GSH-Px activity were found not only between the experimental variants and control but also among the variants themselves. The mean value of GSH-Px activity in the control plants was 97.7 μmol NADPH+H+ min−1 mg−1 protein in cytosol and 94.8 μmol NADPH+H+ min−1 mg−1 protein in mitochondria.

Influence of AR stress on GSH-Px activity in the cucumber cytosol and mitochondria. GSH-Px activity expressed as percentage of control value. For further explanations, see Fig. 2
The present investigations revealed great enhancement of TBARS concentrations in the mitochondrial fraction of AR-treated plants (especially of those that sprayed with pH 1.8 and 3.0 AR) in comparison with the control while the respective values for cytosol were similar to the control (Fig. 5). On the first day of experiment all applied pH variants caused significant increase in lipid peroxidation to 333 (P < 0.01), 233 (P < 0.001) and 150 % (P < 0.05) of the control after pH 1.8, 3.0 and 4.4 treatment, respectively. Contrary to mitochondria, on the first day increase in TBARS concentrations in cytosol was observed only after AR pH 3.0 application (132 % of control; P < 0.05). While in the mitochondrial compartment the largest content of lipid peroxides was noticed on the first day, in all experimental series in cytosol the highest concentration of this compound was found on the third day following exposition to AR pH 1.8 (183 % of control, P < 0.05). Moreover, in pH 1.8 variant enhanced TBARS concentration in mitochondria persisted throughout the experimental time; however, it was slowly decreasing so that on the seventh day the increase was not statistically significant and in pH 3.0 variant it was also enhanced to the end of experimental period. Exposure to AR pH 1.8 increased the value of this parameter to 219 % (P < 0.05) and 220 % (P < 0.01) of the control value on the third and fifth day, respectively. The value of TBARS concentration in the mitochondrial fraction on the third, fifth and seventh day after AR pH 3.0 spraying reached 182 % (P < 0.05), 186 % (P < 0.01) and 139 % (P < 0.05) of the control, respectively. Furthermore, in the cytosolic fraction decrease in TBARS concentration to 67 % (P < 0.05) on the fifth day after AR pH 4.4 exposure was discovered. Similarly as in the case of GSH-Px, differences in lipid peroxidation were found not only between the experimental variants and control but also among the variants themselves, especially in the mitochondrial fraction. Mean value for TBARS concentration in the control plants was 0.940 nmol mg−1 protein in cytosol and 10.2 nmol mg−1 protein in mitochondria.

Influence of AR stress on lipid peroxidation in the cucumber cytosol and mitochondria. Concentration of 2-thiobarbituric acid reactive substances is expressed as percentage of control value. For further explanations, see Fig. 2
Discussion
ROS are generated by mitochondria, chloroplasts and other organelles during their normal activities, and at the same time these compartments are sensitive to ROS (Jones 2000). Mitochondria, like other compartments of a plant cell, house both enzymatic and non-enzymatic antioxidants to prevent ROS accumulation. Mitochondria isolated from the plants subjected to AR stress showed significantly higher TBARS concentration than those isolated from the control plants. This suggests that AR stress results in an increased rate of ROS production and in consequence a higher degree of lipid oxidation in mitochondrial membranes. The fact that mitochondria are the site of mitochondrial electron transport chain reactions and that they harbour a developed membrane system forming cristae makes their membrane lipid compounds, especially predisposed to oxidative damage (Rhoads et al. 2006). In the present work, the extent of oxidative lipid damage in mitochondria depended on the pH value of AR. The lowest pH value caused the highest lipid peroxidation in this cell compartment. These observations are in agreement with our previous work (Gabara et al. 2003), in which we reported that the main ultrastructural changes in mesophyll cells of tomato (Lycopersicon esculentum Mill.) leaves treated with AR pH 1.8 were located in mitochondria. Four days following a single spraying the lesions were observed only in a few chloroplasts (13 %) and almost in al mitochondria (95 %). Bartoli et al. (2004) found out that in wheat plants (Triticum aestivum L.) growing under non-stressful conditions, mitochondria contained 9- to 28-fold higher concentrations of oxidatively modified proteins than chloroplasts or peroxisomes. They pointed out that the mitochondria were extremely susceptible to oxidative damage and were the main target under the oxidative stress conditions. Moreover, many mitochondrial proteins, e.g. those involved in cellular respiration are susceptible to oxidative modifications (Møller 2001). It is thought that damage of mitochondrial membranes increases their permeability and leakage of lipid peroxides to cytoplasm (Gidrol et al. 1989). It is suggested that these compounds may participate in signal transduction between mitochondria and other organelles and nucleus. Lipid peroxides may also trigger antioxidative response in other cell compartments as well as programmed cell death if stress is intensive (Spiteller 2003).
In comparison with the mitochondrial fraction of cucumber leaves, in cytosol increase in TBARS concentration was less dynamic in all experimental variants. Considering our earlier results (Wyrwicka and Skłodowska 2006), triple AR treatment caused much stronger lipid oxidation in cytosolic fraction than a single one in the present work. Lower TBARS level in cytosol than in mitochondrial fraction after AR application could be caused by lower ROS production in this cell compartment, in comparison with mitochondria which are one of the main site of ROS production and/or better antioxidative protection of this cell compartment. However, it is known that tissue capability of buffering acidic input is also associated with buffering capacity of cell sap (Soares et al. 1995). The latter is strongly connected with leaf internal pH and leaf cation content. Therefore, the degree of oxidative damage could be linked not only to pH but to ion composition of AR as well.
In our current investigation the activity of SOD in cytosol of AR treated cucumber leaves (with one exception) was not changed or was lower than in the control plants. Moreover, decreasing trend of this enzyme activity was observed in mitochondria throughout the experiment. Similar tendency was observed in other works, e.g., in tomato leaves (Gabara et al. 2003) where exposition of plants to AR pH 1.8 triggered induction of SOD activity only shortly after stress appearance (0.5–3 h) later on this activity decreased. Reduction of SOD activity after AR treatment was also observed in other plants and presented in several works (He et al. 2011; Tong et al. 2005). On the other hand, there are some articles in which increases in SOD activity were observed as the effects of AR treatment (Dolatabadian et al. 2013). Chen et al. (2013) reported much higher constitutive SOD activity in Schima superb, a plant species tolerant to AR, than in AR-sensitive plants and this activity did not change considerably after appearance of AR stress factor. On the contrary, decreasing SOD activity after AR treatment is characteristic of AR-sensitive plant species, e.g. Liquidambar formosana. Several SOD forms (MnSOD, FeSOD, CuZnSOD) are known to occur in different plant cell compartments (Rabinowith and Fridovich 1983). While CuZnSOD is found primarily in the cytosol (but also in chloroplasts and mitochondria), MnSOD is located primarily in mitochondria (and peroxisomes) (Gupta et al. 1993). CuZnSOD which accounts for the majority of all SOD activity is comparatively stable in a wide pH range (5–9.5) (Ellerby et al. 1996). However, the metal ion affinity of the native zinc binding site drops abruptly but reversibly at low pH and a zinc ion is consequently released which might influence of the enzyme activity (Pantoliano et al. 1979). It cannot be excluded that a similar effect is triggered by AR-induced acidification. Our results seem to suggest that mitochondrial forms of SOD are susceptible to AR treatment.
Data presented in this paper indicate that cucumber plants tried to cope with the oxidative stress induced by AR treatment by strengthening their antioxidant capabilities since the APx activity in both cytosolic and mitochondrial fractions were significantly increased under stress condition. In the present work, enhanced cytosolic APx activity especially after using pH 1.8 solution was observed. Increased level of APx activity was also observed in tomato plants treated with AR with the same pH (Gabara et al. 2003). Similar tendency regarding changes in APx activity was noticed in birch seedlings even though the pH of AR was not very low—4.0 (Koricheva et al. 1997). Some authors claim that APx play a special role in modulation of H2O2 level appearing after exposition of plants to AR stress (Wyrwicka and Skłodowska 2006; Kováčik et al. 2011; Chen et al. 2013). After AR application to AR-sensitive plants (Liquidambar formosana) markedly increased protein expression of APx was noted. On the contrary, in AR-tolerant plants (Schima superba) there were no significant changes in APx protein expression. Moreover, AR-sensitive plants showed decrease in activities of other antioxidative enzymes (SOD, catalase) and simultaneously enhanced H2O2 level (Chen et al. 2013). This results seem to indicate that APx plays the crucial role in the defence against the AR-induced oxidative stress. In present work, AR-induced enhancement of the APx activity in cucumber plants is greater in the cytosolic fraction than in the mitochondrial one. Additionally, activity of mitochondrial APx seems to be more susceptible to AR stress than that of the enzyme present in the cytosolic fraction. It cannot be excluded that after strong acidification of intracellular space which can take place after AR pH 1.8 exposition, mitochondrial metabolism slowed down. Decreased APx activity in the mitochondrial fraction at the beginning of the experiment might be the result of the above changes. Taking into consideration as strong changes in mitochondria ultrastructure (mentioned above) after AR pH 1.8 application may occur (Gabara et al. 2003), slowing down of activity of this antioxidant enzyme is not unexpected.
Our results also support the involvement of GSH-Px in the defence mechanisms in cytosol since the activity of this glutathione-dependent enzyme increased markedly in AR pH 1.8 treated plants. Increase in GSH-Px activity may be connected with sufficient GSH pool in this cell compartment. The supply of cysteine, the end product of sulphur assimilation, is the main factor influencing GSH levels (Mullineaux and Rausch 2005). Environmental stresses associated with atmospheric pollution are among the factors enhancing GSH in plant tissues (Tausz and Grill 2000). It is suggested that increased GSH concentration in plant cells after SO2 fumigation is related more to enlarged uncontrolled sulphur assimilation than to direct GSH reaction with ROS (Schulz and Härtling 2001). A similar situation cannot be excluded in the cytosol isolated from cucumber leaves treated with AR containing high concentration of SO4 2−. It is worth noting that in plants treated with AR pH 3.0 and 4.4, which did not contain so high SO4 2− concentration, there was no increase in GSH-Px activity in the cytosolic fraction. In contrast to the above mentioned cell compartment exposure of the cucumber leaves to AR pH 1.8 did not change mitochondrial GSH-Px activity. Both enzymes, γ-glutamylcysteine synthetase and glutathione synthetase, which are responsible for GSH synthesis, are found in cytosol and plastids, but apparently not in mitochondria (May et al. 1998). Transport of GSH from cytosol to mitochondria is required to ensure sufficient concentrations of this molecule for GSH-Px action in this compartment. It is possible that under stress conditions GSH is consumed by enzymatic and/or non-enzymatic antioxidative reactions in cytosol before it is transported to the mitochondrial compartment. On the other hand, GSH transport through mitochondrial membranes whose lipid compounds undergo peroxidation may be disturbed. Decrease in GSH concentrations may also be caused by inhibition of the enzymes involved in GSH synthesis due to acidification of cell sap.
The present study suggests that even the single AR treatment caused oxidative stress-related changes in pro- and antioxidative systems within cytosolic and mitochondrial cell compartments in a pH-dependent manner. The collected data point out that in comparison with cytosol, plant mitochondria are especially susceptible to AR stress. It is important since they play many crucial functions in plant metabolism, for example, supply of ATP and carbon skeletons for the biosynthesis of several compounds, participation in photorespiration (van Lis and Atteia 2004 and references therein) and programmed cell death (Jones 2000), optimization of photosynthesis and synthesis of ascorbate (Bartoli et al. 2004). Further investigations on antioxidant systems are necessary to understand various plant defence responses. The role of the oxidative damage as an alarm signal triggering plant defence responses under stress conditions seems to be an interesting topic of further investigations.
Author contribution
AW carried out the experiment and wrote this manuscript. MS helped in conception and design of the experiment. Both authors have read and approved the final manuscript.
References
Andreyev AYu, Kushnareva YuE, Starkov AA (2005) Mitochondrial metabolism of reactive oxygen species. Biochemistry (Moscow) 70:200–214
Bartoli CG, Gómez F, Martínez DE, Guiamet JJ (2004) Mitochondria are the main target for oxidative damage in leaves of wheat (Triticum aestivum L.). J Exp Bot 55:1663–1669
Bradford MM (1976) A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72:248–254
Burgess N, Beakes GW, Thomas DR (1985) Separation of mitochondria from microbodies of Pisum sativum L. cv. Alaska cotyledons. Planta 166:151–155
Chen J, Wang W-H, Liu T-W, Wu F-H, Zheng H-L (2013) Photosynthetic and antioxidant responses of Liquidambar formosana and Schima superba seedlings to sulphuric-rich and nitric-rich simulated acid rain. Plant Physiol Biochem 64:41–51
Choi EY, Moon JH, Lee WM, Son SH, Lee SG, Cho IH (2010) The response of antioxidant enzyme activity, growth and yield of pepper and watermelon plants to a single application of simulated acid rain. J Food Agric Environ 8:1265–1271
Dhindsa RS, Plumb-Dhindsa P, Thorpe TA (1981) Leaf senescence: correlated with increased levels of membrane permeability and lipid peroxidation, and decreased levels of superoxide dismutase and catalase. J Exp Bot 32:93–101
Dolatabadian A, Sanavy SAMM, Gholamhoseini M, Joghan AK, Majdi M, Kashkooli AB (2013) The role of calcium in improving photosynthesis and related physiological and biochemical attributes of spring wheat subjected to simulated acid rain. Physiol Mol Biol Plants 19:189–198
Douce R, Christensen EL, Bonner WD Jr (1972) Preparation of intact plant mitochondria. Biochim Biophys Acta 275:148–160
Ellerby LM, Cabelli DE, Graden JA, Valentine JS (1996) Copper-zinc superoxide dismutase: why not pH-dependent? J Am Chem Soc 118:6556–6561
Evans LS, Lewin KF, Patt MJ (1984) Effects of simulated acidic rain on yields of field-grown soybeans. New Phytol 96:207–213
Ferenbaugh RW (1976) Effects of simulated acid rain on Phaseolus vulgaris L. (fabaceae). Am J Bot 63:283–288
Gabara B, Skłodowska M, Wyrwicka A, Glińska S, Gapińska M (2003) Changes in the ultrastructure of chloroplasts and mitochondria and antioxidant enzyme activity in Lycopersicon esculentum Mill. leaves sprayed with acid rain. Plant Sci 164:507–516
Gidrol X, Serghini H, Noubhani A, Mocquot B, Mazliak P (1989) Biochemical changes induced by accelerated aging in sunflower seeds. I. Lipid peroxidation and membrane damage. Physiol Plant 76:591–597
Gimeno L, Marín E, del Teso T, Bourhim S (2001) How effective has been the reduction of SO2 emissions on the effect of acid rain on ecosystems? Sci Total Environ 275:63–70
Gravano E, Ferretti M, Bussotti F, Grossoni P (1999) Foliar symptoms and growth reduction of Ailanthus altissima Desf. in an area with high ozone and acidic deposition in Italy. Water Air Soil Pollut 116:267–272
Gupta AS, Webb RP, Holaday AS, Allen RD (1993) Overexpression of superoxide dismutase protects plants from oxidative stress. Plant Physiol 103:1067–1073
Haines B, Stefani M, Hendrix F (1980) Acid rain: threshold of leaf damage in eight plant species from a southern Appalachian forest succession. Water Air Soil Pollut 14:403–407
Hatch MD (1978) A simple spectrophotometric assay for fumarate hydratase in crude tissue extracts. Anal Chem 85:271–275
He S-Y, Gao Y-J, Shentu J-L, Chen K-B (2011) Combined effects of copper and simulated acid rain on copper accumulation, growth and antioxidant enzyme activities of Rumex acetosa. Chin J Appl Ecol 22:481–487
Hopkins J, Tudhope GR (1973) Glutatione peroxidase in human red cells in health and disease. Br J Haematol 25:563–575
Jones A (2000) Does the plant mitochondrion integrate cellular stress and regulate programmed cell death? Trends Plant Sci 5:225–230
Koricheva J, Roy S, Vranjic JA, Haukioja E, Hughes PR, Hänninen O (1997) Antioxidant responses to simulated acid rain and heavy metal deposition in birch seedlings. Environ Pollut 95:249–258
Kováčik J, Klejdus B, Bačkor M, Štork F, Hedbavny J (2011) Physiological responses of root-less epiphytic plants to acid rain. Ecotoxicol 20:348–357
Kulshrestha UC, Kulshrestha MJ, Sekar R, Sastry GSR, Vairamani M (2003) Chemical characteristics of rainwater at an urban site of south-central India. Atm Environ 37:3019–3026
Kuźniak E, Skłodowska M (1999) The effect of Botrytis cinerea infection on ascorbate-glutathione cycle in tomato leaves. Plant Sci 148:69–76
Kuźniak E, Skłodowska M (2004) Comparison of two methods for preparing mitochondria from tomato leaves to study the ascorbate-glutathione cycle activity. Biol Plant 48:537–542
Lam E, Kato N, Lawton M (2001) Programmed cell death, mitochondria and the plant hypersensitive response. Nature 411:848–853
May MJ, Vernoux T, Leaver C, Van Montagu M, Inzé D (1998) Glutathione homeostasis in plants: implications for environmental sensing and plant development. J Exp Bot 49:649–667
McCord JM, Fridovich I (1969) Superoxide dismutase: an enzymic function for erythrocuprein (hemocuprein). J Biol Chem 244:6049–6050
Minami M, Yoshikawa H (1979) A simplified assay method at superoxide dismutase activity for clinical use. Clin Chem Acta 92:337–342
Miszalski Z, Niewiadomska E, Ślesak I, Lüttge U, Kluge M, Ratajczak R (2001) The effect of irradiance on carboxylating/decarboxylating enzymes and fumarase activities in Mesembryanthemum crystallinum L. exposed to salinity stress. Plant Biol 3:17–23
Mittler R (2002) Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci 7:405–410
Møller IM (2001) Plant mitochondria and oxidative stress: electron transport, NADPH turnover, and metabolism of reactive oxygen species. Ann Rev Plant Physiol Plant Mol Biol 52:561–591
Mullineaux PM, Rausch T (2005) Glutathione, photosynthesis and the redox regulation of stress-responsive gene expression. Photosynth Res 86:459–474
Nakano Y, Asada K (1981) Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol 22:867–880
Neufeld HS, Jernstedt JA, Haines BL (1985) Direct foliar effect of simulated acid rain. I. Damage, growth and gas exchange. New Phytol 99:389–405
Pantoliano MW, McDonnell PJ, Valentine JS (1979) Reversible loss of metal ions from the zinc binding site of copper-zinc superoxide dismutase. The low pH transition. J Am Chem Soc 101:6454–6456
Porra RJ, Thompson WA, Kriedmann PE (1989) Determinate of accurate extinction coefficients and simultaneous equations for assaying chlorophylls a and b extracted with four different solvents: verification of the concentration of chlorophyll standards by atomic absorption spectroscopy. Biochim Biophys Acta 975:384–394
Rabinowith HD, Fridovich I (1983) Superoxide radicals, superoxide dismutases and oxygen toxicity in plants. Photochem Photobiol 37:679–690
Rhoads DM, Subbaiah CC (2007) Mitochondrial retrograde regulation in plants. Mitochondrion 7:177–194
Rhoads DM, Umbach AL, Subbaiah CC, Siedow JN (2006) Mitochondrial reactive oxygen species. Contribution to oxidative stress and interorganellar signaling. Plant Physiol 141:357–366
Rosborg I, Nihlgard B, Gerhardsson L (2003) Inorganic constituents of well water in one acid and one alkaline area of south Sweden. Water Air Soil Pollut 142:261–277
Sant’Anna-Santos BF, Silva LC, Azevedo AA, Aguiar R (2006) Effects of simulated acid rain on leaf anatomy and micromorphology of Genipa americana L. (Rubiaceae). Braz Arch Biol Technol 49:313–321
Schulz H, Härtling S (2001) Biochemical parameters as biomarkers for the early recognition of environmental pollution on Scots pine trees. II. The antioxidative metabolites ascorbic acid, glutathione, α-tocopherol and the enzymes superoxide dismutase and glutathione reductase. Z Naturforsch 56c:767–780
Shan YF, Feng ZW, Izuta T, Aoki M, Totsuka T (1996) The individual and combined effects of ozone and simulated acid-rain on growth, gas exchange rate and water-use efficiency of Pinus armandi Franch. Environ Pollut 91:355–361
Soares A, Ming YJ, Pearson J (1995) Physiological indicators and susceptibility of plants to acidifying atmospheric pollution: a multivariate approach. Environ Pollut 87:159–166
Spiteller G (2003) The relationship between changes in the cell wall, lipid peroxidation, proliferation, senescence and cell death. Physiol Plant 119:5–18
Stoyanova D (1997/98) Effects of simulated acid rain on anatomy of primary leaves of Phaseolus vulgaris. Biol Plant 40:581–588
Stoyanova D, Velikova V (1997/98) Effects of simulated acid rain on chloroplast ultrastructure of primary leaves of Phaseolus vulgaris. Biol Plant 40:589–595
Tausz M, Grill D (2000) The role of glutathione in stress adaptation of plants. Phyton 40:111–118
Tolbert NE (1971) Isolation of leaf peroxisomes. In: Colowick SP, Kaplan NO (eds) Methods in Enzymology, Photosynthesis, part A, vol XXIII. Academic Press, New York, London, pp 665–682
Tong G-H, Liu T-J, Huang W (2005) Effect of simulated acid rain and its acidified soil on lipid peroxidation of wheat seedlings. Acta Ecol Sin 25:1509–1516
Tran TT, Newby G, Mölders N (2011) Impacts of emission on sulfate aerosols in Alaska. Atm Environ 45:3078–3090
van Lis R, Atteia A (2004) Control of mitochondrial function via photosynthetic redox signals. Photosynth Res 79:133–148
Warrilow AGS, Hawkesford MJ (1998) Separation, subcellular location and influence of sulphur nutrition on isoforms of cysteine synthase in spinach. J Exp Bot 49:1625–1636
Willekens H, Van Camp W, Van Montagu M, Inze D, Langebartels C, Sanderman H (1994) Ozone, sulfur dioxide and ultraviolet B have similar effects on mRNA accumulation of antioxidant genes in Nicotiana plumbaginifolia L. Plant Physiol 106:1007–1014
Wyrwicka A, Skłodowska M (2006) Influence of repeated acid rain treatment on antioxidative enzyme activities and on lipid peroxidation in cucumber leaves. Environ Exp Bot 56:198–204
Yagi K (1982) Assay for serum lipid peroxide level its clinical significance. In: Yagi K (ed) Lipid Peroxides in Biology and Medicine. New York Academic Press Inc., London, pp 223–241
Yu JQ, Ye SF, Huang LF (2002) Effects of simulated acid precipitation on photosynthesis, chlorophyll fluorescence, and antioxidative enzymes in Cucumis sativus L. Photosynthetica 40:331–335
Acknowledgments
This work was partly supported by University of Łódź Grant No 505/429.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Z. Miszalski.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution License which permits any use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
About this article
Cite this article
Wyrwicka, A., Skłodowska, M. Intercompartmental differences between cytosol and mitochondria in their respective antioxidative responses and lipid peroxidation levels in acid rain stress. Acta Physiol Plant 36, 837–848 (2014). https://doi.org/10.1007/s11738-013-1462-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11738-013-1462-z