
Abstract
Oryctes monoceros is the most serious pest in coconut plantations, causing up to 40% damage in tropical Africa, especially in Ivory Coast. With a view to reducing pest populations by olfactory trapping, field trials were carried out to assess the efficiency of a synthetic aggregation pheromone: ethyl 4-methyloctanoate (1), 4-methyloctanoic acid (2), a related volatile produced by males, and decaying palm material, either oil palm empty fruit bunches (EFB) or pieces of coconut wood (CW) of various ages. Vertical polyvinyl chloride tube traps (2 × 0.16 m with two openings in the upper half), embedded in the soil, were more efficient than 30-L pail traps 1.5 m above ground. EFB, which were inactive alone, synergized captures with synthetic pheromone. CW was more effective than EFB in comparative trials. Compound 2 did not catch any beetles when assessed with EFB, and reduced catches by 1 + EFB when tested at >10% with the pheromone. Trapping over 6 mo in 2002 and 2003 in a 19-ha coconut plot inside a 4,000-ha oil palm estate reduced damage from 3.8% in 2001 to 0.5% in 2002, then to 0.2% in 2003. Damage was 0.0% in 2004 with routine trapping using 32 traps, which caught 3369 beetles in 9 mo. The results are discussed in relation to other Dynastid palm pests and coconut protection in Ivory Coast.
Similar content being viewed by others

Introduction
Oryctes monoceros Oliv. (Coleoptera, Scarabaeidae, Dynastinae) is one of the main pests in the coconut and oil palm plantations of tropical Africa (Lever, 1969; Mariau, 1981), particularly in Ivory Coast (Echimane et al., 1992). The Asian species, Oryctes rhinoceros L., which is found throughout the Asia-Pacific area, has a similar ecology and is equally harmful to cultivated palms (Waterhouse and Norris, 1987). Damage is generally caused by adults making feeding galleries in the apical section of young palms, but also of mature palms when populations are large. Larvae develop in decaying dead palm wood. Damage has increased over the last 20 yr as a result of large-scale replanting programs, which leave substantial numbers of felled stems on the ground, multiplying suitable sites for egg laying and larva development (Julia and Brunin, 1974). Periodic surveys carried out in plots at the Marc Delorme research station in Port Bouet (Ivory Coast) have revealed up to 40% damage in 0- to 5-yr-old oil palms (Mariau, 1981). In Africa, damage is worsened through attacks by Rhynchophorus phoenicis (Fabr.), which lays eggs in the galleries made by O. monoceros and often kills palms.
Removing or burning felled stems, or covering them with a legume, Pueraria javanica (Benth.), to prevent larval infestation has been recommended (Mariau and Calvez, 1973); however, these measures are rarely applied because of cost and the high planting density of palms (130–160/ha). Biological control has also been studied, but no pathogen (not even Baculovirus oryctes, which is quite active against O. rhinoceros) or parasitoid tested has provided long-lasting action against O. monoceros (Huger, 1966; Julia and Mariau, 1976a; Purrini, 1998; Seguni et al., 1999). The only control method currently used is manual extraction of adults from feeding galleries and insects (especially larvae) present in decaying old stems. This is highly labor intensive and is impossible in tall standing palms. Frequent insecticide applications are only possible in agroindustrial sectors, but they entail serious environmental risks (Echimane et al., 1992).
Other ways have been explored to control this pest, in particular mass trapping with attractants. Ethyl chrysanthemumate was rapidly abandoned because of insufficient catches (Julia and Mariau, 1976b). More recently, ethyl 4-methyloctanoate (1), reported as the male aggregation pheromone of O. monoceros (Gries et al., 1994) and O. rhinoceros (Hallett et al., 1995; Morin et al., 1996), proved to be a powerful attractant for O. rhinoceros (Purba et al., 2000). In a preliminary study, it also showed promise for O. monoceros (Allou et al., 2002).
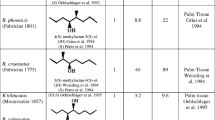
We conducted trials in Ivory Coast to determine if the advances made for O. rhinoceros control (Morin et al., 2001a; Sudharto et al., 2001) could be applied to O. monoceros. We focused our assessment on: (1) the plant material [oil palm empty fruit bunches (EFB) and coconut wood (CW)] as coattractants with 1, (2) the efficiency of two types of traps, and (3) the study of 4-methyloctanoic acid (2), an untested molecule regularly emitted by O. monoceros males (Gries et al., 1994; Rochat et al., 2004).
Materials and Methods
Attractants
Palm material: About 15 L of plant material were used per trap. We tested the following: (1) 1-wk-old oil palm EFB obtained after processing in the oil mills and (2) CW disks (6 cm thick and 30 cm diam) from 40- to 60-yr-old trees that we cut and left on the ground under natural conditions for 0.5, 1, 1.5, 2.5, and 3.5 mo before testing. The disks were sprayed with water from time to time during the dry season. For a given age category, three disks were cut into four pieces and placed in the trap. Plant material was not renewed once the trials had started.
Synthetic Chemicals
Ethyl 4-methyloctanoate: O. monoceros pheromone (1) and 4-methyloctanoic acid (2; 99% chemical purity by gas chromatography; racemic mixtures) were synthesized by EGNO-Chimie (Tancarville, France). The dispensers used were sachets made from high-density polyethylene (Alplast, Sainte-Marie-aux-Mines, France). Compound 1 was formulated in model D dispensers (25 × 50 mm, 200-μm-thick film) containing about 1 mL of pheromone, which emitted 13–14 mg/d under field conditions in an earlier assessment (Allou et al., 2002). Compound 2 was formulated in dispensers made of 100-μm-thick film. Four models were used: d1, 25 × 50 mm; d2, 12.5 × 50 mm; d3, 12.5 × 12.5 mm; and d4, 6.2 × 12.5 mm. To estimate average daily release rates, all dispensers were filled to last >1.5 mo and were weighed monthly and quarterly in trials 1–5, and 6, respectively.
Traps
Two types of trap were used. One is the pail trap that consisted of a covered 30-L plastic bucket with five holes (diam 5 cm) in the lid. The pheromone dispenser was suspended from the lid. Another type, the tube trap, based on the model described by Morin et al. (2001b), consisted of a vertical polyvinyl chloride (PVC) tube (height 2 m, diam 16 cm) with two apertures (20 × 10 cm) in its upper section 140 cm from the ground. The top of the tube was open while the base stood in a recipient with holes for rainwater drain-off. The dispenser was hung inside the tube, above the two apertures, and protected from direct sunlight with a piece of cardboard.
Trials
The study consisted of six trials conducted by using the same layout and procedure. They comprised eight randomized blocks 100 m apart, and within each block the traps, hung on poles 1.5 m from the ground, were spaced 50 m apart. They were inspected every 3 d and rerandomized every 6 d. Trial 5 compared both types of traps.
The majority of the trials were carried out at the Robert Michaux plantation, in Dabou, 65 km west of Abidjan. The traps were installed in a 19-ha plot, a coconut seed garden, located in the middle of a 4000-ha oil palm estate, planted in 1977 with the Malayan Yellow Dwarf variety. However, trial 3 was conducted in a 12-ha experimental coconut plantation in Assinie (30 km east of Abidjan on the coast) planted in 1999 with PB 121 hybrids combined with tree legumes, Acacia mangium and Acacia auriculiformis.
-
Trial 1 (effect of addition of EFB to 1) was carried out from 24 January to 21 February 2002, and compared the following three treatments: (1) 1 only, (2) 1 + EFB, and (3) EFB only.
-
Trial 2 (comparative effects of EFB and CW with 1) was conducted from 16 September to 16 October 2002, and comprised six treatments: (1) 1 only, (2) 1 + EFB, (3–6) 1 + 3.5-, 2.5-, 1.5-, and 0.5-mo-old CW, respectively.
-
Trials 3 and 4 assessed the effect of 2 with 1 + EFB. Trial 3 was carried out from 2 August to 1 September 2002, with five treatments: (1) 1 + EFB, (2–5) 1 + EFB + 2 emitted from dispensers d1, d2, d3, and d4, respectively. Trial 4 was conducted from 23 January to 22 February 2003, with the same protocol but with an additional treatment, (6) EFB + 2 from dispenser d1.
-
Trials 5 and 6 compared the effectiveness of both types of traps (pail and tube) with 1 + either EFB or CW. They comprised four treatments: (1) 1 + EFB in the pail trap, (2) 1 + EFB in the tube trap, (3) 1 + CW in the pail trap, and (4) 1 + CW in the tube trap. Trial 5 took place from 19 September to 22 December 2003, and plant material was not renewed. In trial 6, EFB and CW were renewed each month and every 6 mo, respectively. Pheromone dispensers were renewed every 3 mo in both trials. In trial 6, bait renewal was postponed by ±10 d on some occasions because of unforeseeable events. Trial 6, started on 7 April 2004, was set up for pest control and served, secondarily, to assess the monthly renewal of EFB (as compared to trial 5), and, as it is ongoing, we present 15 mo of results (last data from 1 July 2005).
Effect of Trapping on Coconut Damage
The coconut seed garden at the Dabou plantation had been planted to produce hybrid seeds but was abandoned in the 1990s. In 2000, it was decided to restore it to its initial function, and new management practices, including monthly inspections, were introduced to monitor the phytosanitary condition of the palms. A report is prepared yearly, including the percentage of palms killed by combined O. monoceros and R. phoenicis (Coleoptera, Curculionidae) attacks. From January 2000 to December 2001, no special effort was made to control the beetle due to the large size of the palms and the absence of traps. However, damage was high, with surrounding oils palms a constant source of beetles, so it was decided to start pheromone trapping as a control option with trials 1–2 and 4–6 starting in January 2002. At that time, 24–48 traps (1.2–2.5 traps/ha) were installed in the block, on three occasions for 1 mo, in 2002 and early 2003 (trials 1, 2, and 4). Thirty-two traps (1.7 per ha) were installed for another 12 mo, then permanently from April 2004 (trials 5–6). The effect of olfactory trapping was assessed by studying annual trends in coconut palm mortality over a 5-yr period (2000–2004).
Statistical Analyses
In all the trials, the individual in each trap were pooled together over 6-d periods. Trials 1–4: The effect of the treatments was assessed by ANOVAs on values transformed by (x + 0.5)1/2. Differences in average catches were assessed by Newman–Keuls multiple comparison tests at 5% (Statitcf Software, 1991). Trials 5–6: Data were analyzed by using a multifactor ANOVA with a GLIM procedure after transformation by Ln(x + 1). The experimental factors were trap type (pail or tube), the kind of palm coattractant (EFB or CW), and the block. The trapping period was an additional factor included in the model, with corresponding interactions for trial 5. The average ages of the baits after installation (EFB, CW, and pheromone dispensers) were introduced as covariables, as catches clearly fluctuated periodically in trial 6 (Fig. 4). Differences in average catches were assessed by Tukey multiple comparison tests (Minitab v12.2, 1998).
Results
Compound 1 was emitted at 12.7 ± 1.9 mg/d (mean ± S.D.; N = 384) over all trials, with a minimum of 9.1 mg/d in trial 5 to a maximum of 15.2 mg/d in trial 1. Compound 2 was emitted at 3.8 ± 0.5, 2.4 ± 0.6, 1.3 ± 0.4, and 0.6 ± 0.2 (N = 16) from dispensers d1, d2, d3, and d4, respectively.
More males than females were caught in trials 1–4 (64% of 794) and 6 (57% of 4,989), while in trial 5 both sexes were equally represented (51% of 704).
Effect of Addition of Palm Material to Compound 1
A total of 292 beetles was captured in trial 1. The majority were caught in EFB + 1 traps, and while there were significantly fewer in 1 traps, 1 alone was more effective than EFB alone (Fig. 1; Newman–Keuls tests, P < 0.05).

Comparative catches of O. monoceros in 30-L pail traps baited with synthetic pheromone: ethyl 4-methyloctanoate (1) diffused at a rate of 15 mg/day, 1-wk-old oil palm EFB, or the two combined (trial 1). Treatments with different letters differed significantly (Newman–Keuls test at α = 0.05 on (x + 0.5)1/2 transformed data)
A total of 340 beetles was caught in trial 2. Whereas there were no significant differences between the treatments that included a source of palm volatiles, traps baited with the pheromone 1 and either 3.5- or 2.5-mo-old CW captured significantly more than 1 alone (Fig. 2).

Comparative catches of O. monoceros in 30-L pail traps using synthetic pheromone: ethyl 4-methyloctanoate (1) without plant material (control) or with various palm coattractants: 1-wk-old oil palm EFB, CW after aging outdoors for 0.5, 1.5, 2.5, or 3.5 mo (trial 2). Treatments with different letters differed significantly [Newman–Keuls test at α = 0.05 on (x + 0.5)1/2 transformed data]
Effect of Compound 2
In 2002, the release rate of 2 did not significantly affect the trap catch. However, at the two highest release rates, the number of beetles captured was significantly lower than in traps without 2 (Fig. 3a). In 2003, very few beetles were captured, due in part to heavy rainfall (some traps were flooded). However, as in the previous year, traps with EFB + 1 captured more beetles than EFB + 1 with 2 at the highest release rate although lower rates gave variable results (Fig. 3b). EFB + 2 captured no beetles at all.

Effect of 4-methyloctanoic acid (2), a volatile emitted by males, assessed at different ratios with synthetic pheromone: ethyl 4-methyloctanoate (1) in 30-L pail traps containing 1-wk-old oil palm EFB on O. monoceros catches in (a) trial 3, 2002, and (b) trial 4, 2003. Treatments with different letters differed significantly [Newman–Keuls test at α = 0.05 on (x + 0.5)1/2 transformed data]
Comparison of Trap Types with EFB or CW
Trap type and the kind of palm coattractant added to 1 significantly affected catches (trap: F 1,41 = 21.8; F 1,24 = 141; palm: F 1,41 = 16.1; F 1,24 = 15.0 in trials 5 and 6, respectively; P < 0.001). Tube traps caught more beetles than pail traps, whatever the bait, whereas CW + 1 was more attractive than EFB + 1 regardless of trap type and renewal frequency (Table 1).
A significant “palm × trap” interaction was observed in trial 6 (F 1,24 = 4.44; P < 0.05; Table 1) due to the unexpectedly high performance of EFB in tube traps, although globally CW was better than EFB, irrespective of trap type. The presence of a significant interaction in trial 6 but not in trial 5 could be the result of renewing EFB monthly in trial 6, combined with other unknown factors.
Seasonal Variation in Catches in Trial 6
About 5000 beetles were removed from the plot in 15 mo. The sex ratio of the beetles fluctuated from 26% to 70% female, with an average value of 43%. Overall, no significant decrease in catches (C as the mean catch per period) was recorded over time (T as the 6-d period numbered from 1 to 75; C = −0.008 + 2.38T; F 1,73 = 2.50; P = 0.12). Catches ranged from 0.2 to 4.8 beetles/trap/6 d, with an average of 2.1 (Fig. 4). These fluctuations were not correlated to the age of the palm material (EFB: F 1,24 = 0.40 and CW: F 1,24 = 0.22; P > 0.5) but significantly decreased with pheromone dispenser age (F 1,24 = 16.06; P < 0.001). On two occasions, when bait renewal was unavoidably delayed, most of the dispensers ran out and resulted logically in lower catches (Fig. 4).

Variation in O. monoceros catches at the Robert Michaux Plantation over 15 mo with 32 (16 pail plus 16 tube) traps baited with synthetic pheromone: ethyl 4-methyloctanoate (1) and 15 L of either oil palm EFB (renewed monthly) or coconut stem wood (CW renewed quarterly) from April 2004 to July 2005 (trial 6). Arrows indicate actual renewal of CW, EFB, and pheromone (P) and unavoidable run out of most of the pheromone dispensers (Pro)
Effectiveness of Olfactory Trapping in the 19-ha Seed Garden at the Robert Michaux Plantation
Damage (dead coconut palms) caused by O. monoceros + R. phoenicis in the coconut seed garden at the Dabou plantation was 3.5% in 2000 and 3.8% in 2001 (Fig. 5). No insect control (neither insecticide applications nor particular cultural practice) was carried out in those 2 yr, except the elimination of dead coconut palms. Trapping clearly had an effect as palm mortality dropped to 0.5%, 0.2%, and 0.0% in 2002, 2003, and 2004, respectively (Fig. 5).

Reduction in damage (dead palms) in the 19-ha coconut seed garden at the Robert Michaux plantation (Dabou) after 15 mo of olfactory trapping from January 2002 to December 2004 without any modification to the other plant protection measures. The dotted line indicates when trapping was initiated and the gray bars when trials 1, 2, and 4–6 were run
Discussion
Synergistic Effects of Decaying Palm Material with Pheromone
Our results show that decaying palm material, either EFB or CW, is a powerful coattractant with the aggregation pheromone 1 of O. monoceros, as previously reported for O. rhinoceros (Hallett et al., 1995; Alfiler, 1999; Sudharto et al., 2001). The effect of these host plant odors is another similarity of the two Oryctes species, which use the same aggregation pheromone. Although both odors are synergistic with 1, CW was more efficient than EFB. CW can be prepared shortly before trapping starts, and is effective for 6 mo in both trap types. In contrast, EFB only works in a confined environment (straight pipe) and must be replaced each month.
Although the use of readily available plant material will enhance trap efficacy, the need to change it on a regular basis is labor intensive for vast trap networks. For instance, about 1 m3 of EFB is needed to service one trap for 1 yr. However, in using CW that only needs to be changed every 3 mo instead of monthly, one needs a chainsaw to prepare the pieces of CW effectively. Thus, the financial investment of this approach limits the availability to owners of small holdings.
The synergistic effect of plant odors on pheromone communication is common in insects, particularly in long-lived Coleoptera (Landolt and Phillips, 1997). This has been well documented for palm weevils Rhynchophorinae (e.g., Rhynchophorus and Metamasius spp.; Giblin-Davis et al., 1996; Rochat et al., 2000; Avand-Faghih 2004; Saïd et al., 2005) and certain scarabs, Dynastinae (e.g., Scapanes australis Bsdv. and Oryctes elegans: Rochat et al., 2002, 2004), where volatiles from freshly cut host plant tissues serve as cosignals with the pheromones for beetle aggregation on sites where both feeding, mating, and egg laying take place.
However, O. rhinoceros and O. monoceros feed individually rather than in aggregations on fresh tissues of living palms, although a palm may be attacked by several insects in succession (Morin, 1999). Thus, in these species, the synergistic effect of volatiles from decaying palm woody material (CW or EFB) with the pheromones is only important for aggregation at mating and egg-laying sites (Morin and Ollivier, 2001)
Inhibiting Effect of Compound 2
It is clear that when 4-methyloctanoic acid is combined with the aggregation pheromone (∼30:70 acid/ester), there is a significant decline in the number of O. monoceros captured. This is in marked contrast with the findings for two other Oryctes species. In the case of O. elegans, 4-methyloctanoic acid serves as an aggregation attractant when combined with odors of freshly cut date palm stem (Rochat et al., 2004), whereas at 30 mg/d, either evaluated alone or combined with the ethyl ester, it has no effect on O. rhinoceros (Hallett et al., 1995). Thus, Oryctes species appear to use 4-methyloctanoic acid and the ethyl ester derivative differently.
Interestingly, O. monoceros males emit variable amounts and ratios of both compounds (Gries et al., 1994; Morin et al., 1996). Although the origin and meaning of this variability are unknown, males may modulate the amount of acid emitted under certain circumstances and this may act as an antiaggregation signal to regulate densities at breeding sites (decaying woody material). Further research is required to clarify the situation, including the importance of chirality on the responses observed.
Comparison of Trap Types
The tube trap was more efficient than the pail trap for O. monoceros, as it was for O. rhinoceros (Morin et al., 2001b). The height and shape appear to be more adapted to the flight and approach behavior of the beetle, and insects entering the trap are prevented from escaping, without the need for insecticide or soapy water. The shape also helps keep the plant material moist for longer than in the more aerated pails, thereby lengthening the amount of time the pheromone coattractants remains effective. Thus, as long as the bait remains active, the tube trap, which is sturdier that the pail trap in field conditions, could stay in the field for a long time without inspection or maintenance, making it suitable for use in coconut small holdings. The disadvantage remains to be its higher initial cost, and the availability of PVC tubes in remote areas.
Effect of Trapping on Damage to Coconut Palms
Despite continuous and stable catches over 3 yr, the damage caused by Oryctes (+ Rhynchophorus) decreased substantially in the 19-ha plot. The impact of olfactory trapping regimes have generally examined the reduction in a pest populations and this is, to our knowledge, the first comprehensive study showing reduced damage, as earlier reports analyzing 1–2 yr of trapping, reported mitigated effects (Ho 1996; Chung 1997). The insects caught since 2004 probably came from the oil palm plots surrounding the nursery, and the olfactory trapping clearly reduced attacks on palms whose considerable height (>8 m) prevented manual extraction of the insects from the galleries and insecticide applications. Our work has shown that olfactory trapping can be beneficial, and trials are under way to assess whether trapping intensity can be reduced while maintaining the benefit. This could be performed by either decreasing the trap density or by carrying out intermittent trapping at regular intervals. Studies are also in progress to evaluate the effect of trapping in immature crops.
References
Alfiler, A. R. 1999. Increased attraction of Oryctes rhinoceros aggregation pheromone, ethyl 4-methyloctanoate, with coconut wood. Coconut Res. Dev. (CORD) 15:106–114.
Allou, K., Morin, J.-P., and Rochat, D. 2002. Amélioration du piégeage olfactif de Oryctes monoceros (Olivier) (Coleoptera, Dynastidae), ravageur du cocotier et du palmier à huile en Côte d'Ivoire. Proceedings of AFPP-6e Conférence Internationale sur les Ravageurs en Agriculture. Montpellier, France, pp. 295–302.
Avand-Faghih, A. 2004. Identification et application agronomique de synergistes végétaux de la phéromone du charançon Rhynchophorus ferrugineus (Olivier) 1790. Thèse de Doctorat de l'INA-PG, Paris. 171 p.
Chung, G. F. 1997. The bioefficacy of the aggregation pheromone in mass trapping of rhinoceros beetles (Oryctes rhinoceros L.) in Malaysia. The Planter (Kuala Lumpur) 73:119–127.
Echimane, M., Amani, I. C, Lanon, Y. S., Gnigou, J. S., Zakra, A. N., and Blehou, N. 1992. Les ennemis du cocotier. Les cultures d'exploitation. Le guide de l'agriculteur en Côte d'Ivoire. Vol II, pp. 101–110.
Giblin-Davis, R. M., Peña, J. E., Oehlschlager, A. C., and Perez, A. L. 1996. Optimisation of semiochemical-based trapping of Metamasius hemipterus sericeus (Ollivier) (Coleoptera: Curculionidae). J. Chem. Ecol. 22:1389–1410.
Gries, G., Gries, R., Perez, A. L., Oehlschager, A. C., Gonzalez, L. M., Pierce, H. D. Jr, Zebeyou, M., and Kouame, B. 1994. Aggregation pheromone of the African rhinoceros beetles, Oryctes monoceros (Olivier) (Coleoptera, Dynastidae). Z. Naturforsch. 49c:363–366.
Hallett, R. H., Perez A. L., Gries, G., Gries, R., Pierce, H. D. Jr, Yue, J., Oehlschlager, A. C., Gonzalez, L. M., and Borden, J. H. 1995. Aggregation pheromone of the coconut rhinoceros beetle, Oryctes rhinoceros (L.) (Coleoptera, Scarabaeidae). J. Chem. Ecol. 21:1549–1570.
Ho, C. T. 1996. The integrated management of Oryctes rhinoceros (L.) populations in the zero burning environment. Proceedings of the PORIM international oil palm congress, Kuala Lumpur, Malaysia. pp. 336–368.
Huger, A. M. 1966. A virus disease of the Indian rhinoceros beetle Oryctes rhinoceros caused by a new type of insect virus Rhabdionvirus oryctes. J. Invertebr. Pathol. 8:38–51.
Julia, J.-F. and Brunin, C. 1974. Recherches sur Oryctes monoceros en Côte d'Ivoire : Enquête sur les attaques d'Oryctes dans les jeunes plantations villageoises. Oléagineux 29:239–242.
Julia, J.-F. and Mariau, D. 1976a. Recherches sur Oryctes monoceros Ol. en Côte d'Ivoire. II. Essais de lutte biologique à l'aide du virus Rhabdionvirus oryctes. Oléagineux 31:113–117.
Julia, J.-F. and Mariau, D. 1976b. Piégeage olfactif à l'aide du chrysanthémate d'éthyle. Recherches sur Oryctes monoceros (Olivier) en Côte d'Ivoire. Oléagineux 31:263–276.
Landolt, P. J. and Phillips, T. W. 1997. Host plant influences on sex pheromone behaviour of phytophagous insects. Annu. Rev. Entomol. 42:371–391.
Lever, R. J. A. 1969. Les ravageurs du cocotier. FAO Bull. 77:120–185.
Mariau, D. 1981. Les ravageurs du palmier à huile et du cocotier en Afrique Occidentale. Oléagineux 36:169–227.
Mariau, D. and Calvez, C. 1973. Méthode de lutte contre l'Oryctes en replantations de palmier à huile. Oléagineux 28:215–218.
Minitab, 1998. Minitab for Windows Version 12.2 in French, Minitab SARL, 1 Cité Paradis, 75010 Paris.
Morin, J.-P., 1999. Study mission in Indonesia for Improvement of Oryctes rhinoceros trapping. Rapport de mission. Cirad, Montpellier, France. Doc CP 1086.
Morin, J.-P. and Ollivier, L. 2001. New technology of pest management against insect pests of oil palm and coconut crops: Research on and development of selective trapping using synthetic attractants. INCO project ERB-IC18CT970199, Annual report 4, November, 2001, DOC CP-SCI no. 1419, Cirad-cp, Montpellier, France.
Morin, J.-P., Rochat, D., Malosse, C., Lettere, M., Desmier de Chenon, R., Wibowo, H., and Descoins, C. 1996. Ethyl 4-methyloctanoate, major component of Oryctes rhinoceros (L.) (Coleoptera, Dynastidae) pheromone. C. R. Acad. Sci., Ser. 3 Sci. vie 319:595–602.
Morin, J.-P., Prawirosukarto, S., Purba, R., Beaudoin-Ollivier, L., Kakul, T., Aldana, R., Philippe, R., Dery, S., Desmier de Chenon, R., and Rochat, D. 2001a. La protection des jeunes palmiers. Un moyen prometteur: le piégeage sélectif de masse des coléoptères ravageurs. Proceedings of the International Conférence “L'Avenir des Cultures Pérennes”, Yamoussoukro, Ivory Coast, pp. 95–96.
Morin, J.-P., Sudharto, Ps., Purba, R., Desmier de Chenon, R., Kakul, T., Laup, S., Beaudoin-Ollivier, L., and Rochat, D. 2001b. A new type of trap for capturing Oryctes rhinoceros (Scarabaeidae, Dynastidae), the main pest in young oil palm and coconut plantings. Coconut Res. Dev. (CORD) 17:13–22.
Purba, R., Prawirosukarto, S., Desmier de Chenon, R., Morin, J.-P., and Rochat, D. 2000. Effect of Oryctes rhinoceros pheromone (Ethyl 4-methyloctanoate) diffusion rate on the size of pest catches. Proceedings of XXIth International Congress of Entomology. Foz do Iguaçu, Brazil, vol. I, p. 181.
Purrini, K. 1998. Baculovirus oryctes release into Oryctes monoceros populations in Tanzania with special reference to the interaction of virus isolates used in our laboratory infections experiments. J. Invertebr. Pathol. 3:285–300.
Rochat, D., Ramirez-Lucas, P., Malosse, C., Aldana, R., Kakul, T., and Morin, J.-P. 2000. Role of solid phase micro-extraction in the identification of highly volatile pheromones of two Rhinoceros beetles Scapanes australis and Strategus aloeus (Coleoptera, Scarabaeidae, Dynastinae). J. Chromatogr. A 885:433–444.
Rochat, D., Morin, J., Kakul, T., Beaudoin-Ollivier, L., Prior, R., Renou, M., Malosse, I., Stathers, T., Embupa, S., and Laup, S. 2002. Activity of male pheromone of Melanesian rhinoceros beetle Scapanes australis. J. Chem. Ecol. 28:479–500.
Rochat, D., Malosse C., Lettere, M., Beauhaire, J., Morin J-P., Pezier A., Renou, M., and Abdollahi, G. A. 2004. Male aggregation pheromone of date palm fruit stalk borer Oryctes elegans. J. Chem. Ecol. 30:387–407.
Saïd, I., Renou, M., Morin, J.-P., Ferreira, J. M. S., and Rochat, D. 2005. Interactions between acetoin, a plant volatile and pheromone in Rhynchophorus palmarum: Behavioral and olfactory neuron responses. J. Chem. Ecol. 31:1789–1805.
Seguni, Z., Lohr, B., and Mwaiko, W. 1999. Introducing Baculovirus oryctes Huger into Oryctes monoceros Oliv. populations in Tanzania. J. Appl. Entomol. 123:427–431.
Statitcf, 1991. Statistical Software, version 5. Institut technique des céréales et des fourrages, Paris.
Sudharto, P. S., Purba, R. Y., Rochat, D., and Morin, J-P. 2001. Synergy between empty oil palm fruit bunch and synthetic aggregation pheromone (ethyl 4-methyloctanoate) for mass trapping of Oryctes rhinoceros beetles in the oil palm plantation in Indonesia. Proceedings of the PORIM International Palm Oil Congress, Kuala Lumpur, Malaysia. Aug. 20–23, 10 p.
Waterhouse, D. F. and Norris, K. R. 1987. Oryctes rhinoceros (L.) In Biological Control Prospects. ACIAR, Inkata Press. Melbourne, Australia, pp. 101–317.
Acknowledgments
We are grateful to Peter Biggins from CIRAD for the English revision of the manuscript, and to two anonymous reviewers and Jeremy McNeil for helpful criticism.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Allou, K., Morin, JP., Kouassi, P. et al. Oryctes monoceros Trapping with Synthetic Pheromone and Palm Material in Ivory Coast. J Chem Ecol 32, 1743–1754 (2006). https://doi.org/10.1007/s10886-006-9106-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10886-006-9106-z