
Abstract
When new individuals from a pest species are detected following eradication, identifying whether the new individuals are survivors from the eradication attempt, or reinvaders from another population, is important for management practices. Pearl Island (512 ha) in New Zealand was the first island in the world on which simultaneous eradication of all three invasive rat species was attempted. Rats were detected again 9 months after the eradication operation. We use genetic assignment methods to discriminate between survivor and reinvader hypotheses. All rats found on Pearl Island after eradication were likely to be reinvaders from an adjacent population on much larger Stewart Island (174,600 ha), suggesting that rats were swimming to the island at a rate much greater than anticipated, but that the original eradication was successful. Adequate genetic signal was obtained from opportunistically collected samples, making the method feasible for conservation managers with limited time and resources.
Similar content being viewed by others

Introduction
Invasive species are now a component of almost every ecosystem on earth (Vitousek et al. 1997; McKinney and Lockwood 1999), and are regarded as an important driver of species extinction (Blackburn et al. 2004). Eradication of invasive pests is the ultimate goal of pest management in terrestrial and marine systems (Myers et al. 2000; Simberloff 2003; Thresher and Kuris 2004), and is now a common management tool on islands (Donlan et al. 2003). However, pest individuals may reappear after the eradication attempt. If a long time has elapsed since eradication it may be assumed that this is an independent reinvasion, but when pests are rediscovered promptly it might not be clear whether the new individuals are survivors from the eradication attempt, or reinvaders (Abdelkrim et al. 2007). This has important management implications, because failed eradications may necessitate a change of protocol for the future (Abdelkrim et al. 2005), or alternatively, a revision of biosecurity measures to prevent reinvasion (Russell et al. 2008b).
Genetic techniques are now affordable and widely used in ecology (Selkoe and Toonen 2006), and include assignment methods for assigning individuals to a source population (Davies et al. 1999). Assignment methods compute the probability of finding the genotype of a given individual within each of a sampled set of populations, and can be used to select or exclude populations as possible sources for the individual (Manel et al. 2005). These methods have important applications in conservation (e.g. Manel et al. 2002; Berry et al. 2004; Robertson and Gemmell 2004). For invasive pest species following eradication, assignment methods can be used to select between the hypotheses of survival or reinvasion.
The invasive rat species Rattus exulans, R. norvegicus, and R. rattus are among many pest species now routinely eradicated from islands (Howald et al. 2007), but rat eradication attempts remain uncommon where there are multiple pest species or high reinvasion risk. R. norvegicus are known to be capable swimmers, and are considered the most likely invaders of offshore islands in New Zealand (Russell et al. 2005), and reinvading rats can be difficult to detect at low densities (Russell et al. 2008a).
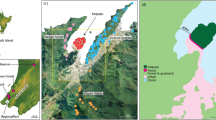
Pearl Island (512 ha; 47°11′S, 167°42′E; Fig. 1) was the first island in the world on which simultaneous eradication was attempted for all three invasive rat species (Clout and Russell 2006). Rats were eradicated during July and August 2005 by a standard aerial poison baiting program using brodifacoum 20R pellets (Animal Control Products Ltd, Whanganui, New Zealand). Low levels of reinvasion by swimming from the adjacent Stewart Island population were expected across Whale Passage and so four post-eradication monitoring lines each 1 km long were established on the north-eastern slopes by Whale Passage. Monitoring lines consisted of a combination of kill-traps, bait-stations, and monitoring tunnels. Monitoring took place every 3 months after eradication.

Pearl Island, north-east Port Pegasus, Stewart Island
No sign of rodents was detected until early May 2006, 9 months after eradication, when a rat was observed in the camp site. No sign of rodents in any monitoring devices had been found at this time, highlighting the difficulty of detecting rats at low density. Five rats were then caught in May 2006 (4 R. norvegicus, 1 R. rattus). All rodent monitoring to this point was confined to north-east Pearl Island adjacent to Whale Passage, so it remained unknown whether rats were widespread across the island or confined to this vicinity. In July eight more rats (unidentified species) were found in detection devices. A total of 13 rats had now been caught, consisting of both R. norvegicus and R. rattus and both sexes. No young rats indicative of recent breeding were caught. Whether the post-eradication rats were survivors of the eradication attempt, reinvaders across Whale Passage from Stewart Island, or a combination of both, was unknown. By April 2007, an R. norvegicus population was confirmed across the entire island.
Under current rat eradication protocols, there is a management expectation of complete eradication success. A known eradication failure would instigate costly investigation and revision of protocols. We aimed to evaluate if genetic assignment methods could successfully discriminate between post-eradication survivors or re-invaders. Such a method is relevant for guiding ongoing invasive rat management on Pearl Island, and in other similar management scenarios.
Materials and methods
Study site
Pearl Island lies in north-east Port Pegasus, Stewart Island. At its closest point to Stewart Island it is separated by a 225 m channel across Whale Passage, which lies parallel to the coast of Pearl Island for about 800 m. The average water temperature around Stewart Island throughout the year is 12°C. Vegetation on the island is described by Harper (2006). Pearl Island is rarely visited by people or boats, and there are no easy landing points around the island. Boats occasionally pass through Whale Passage as they enter Port Pegasus. On neighbouring Stewart Island, all three species of invasive rat are present. Most common is R. rattus, while R. norvegicus and R. exulans are restricted to specific habitats (Harper et al. 2005; Russell and Clout 2004).
Field sampling
Rat tissue samples were acquired from the pre-eradication Pearl population (12 R. norvegicus and 11 R. rattus) the adjacent Stewart Island population (9 R. norvegicus and 8 R. rattus), and the post-eradication Pearl individuals (10 R. norvegicus and 1 R. rattus). All rat tissue samples were stored in 70% ethanol at room temperature. For the pre-eradication Pearl population, samples were gained opportunistically from a study of habitat use by all three rat species shortly before the eradication in June 2005 (Harper 2006). This sample size is constrained, as no further pre-eradication rats can now be sampled. For post-eradication Pearl individuals, tissue samples were available from 10 R. norvegicus and 1 R. rattus, comprising the first five rats caught in May 2006, three of eight rats caught in July 2006, and three more rats caught in August and September 2006. The three unidentified rats in this sample were identified as R. norvegicus by our DNA analysis. For the Stewart Island population, rats were obtained from September to November 2006 on the region of Stewart Island adjoining north-west Whale Passage, for the purpose of this study.
An additional 31 tissue samples of R. rattus were available from three other locations on Stewart Island, all captured in 2005 and 2006: 14 samples from Tupari; 11 from Rakeahua; and 6 from Ackers Point. These locations are, respectively, 18, 25, and 45 km from the region adjoining Whale Passage. We use these samples to give a wider context to our results when identifying possible source populations for some rats.
Genetics
DNA was extracted using the DNeasy Tissue Kit (Qiagen, Hilden, Germany). Eleven microsatellite markers characterised for R. norvegicus genome mapping were used on both rat species (Jacob et al. 1995; D10Rat20, D11Mgh5, D12Rat76, D15Rat77, D16Rat81, D18Rat96, D19Mit2, D20Rat46, D2Rat234, D5Rat83, D7Rat13). To avoid physical linkage, markers were chosen on different chromosomes. Each forward locus primer was labelled with fluorescent dyes before amplification by polymerase chain reaction (PCR). PCR was performed in 10 μl volumes, containing 10 ng DNA, 0.1 μM of forward primer labelled with 5′ fluorescent labels, 0.2 μM of reverse primer, 0.2 μM of each dNTP, 0.2 units Platinum Taq DNA polymerase (Invitrogen, Carlsbad, USA), and 1× reaction buffer with 1.5 mM MgCl2. PCR products were pooled with Genescan 400HD [ROX] size standard for a single run using an ABI Prism 3730 Genetic Analyzer capillary electrophoresis system (Applied Biosystems, Foster City, USA). Amplification size was scored using Genescan Analysis 3.7 and Genotyper 3.7.
Samples with missing data were amplified and scored a second time to identify missing values where possible.
Statistical analysis
For the samples from the pre-eradication Pearl Island and Stewart Island populations, we calculated allele frequencies, observed and expected heterozygosities, and mean number of alleles per locus as summaries of genetic diversity within the two hypothesised source populations. Our assignment methods rely on assumptions of Hardy–Weinberg proportions and no linkage disequilibrium, so we checked these in each population using Fisher’s exact tests. We estimated F IS and F ST, with 95% confidence intervals from 10,000 bootstraps across loci, following Weir (1996). We did not treat the post-eradication rats as a population, because each individual could have been either a survivor or a reinvader. All calculations were done in R (Version 2.4.0).
The small sample sizes raised concerns that the observed differentiation between the assignment populations (i.e. the Pearl Island rats pre-eradication, and the rats from mainland Stewart Island) might arise through sampling variability alone even if the populations were genetically identical. To test for the possibility that the assignment populations were not genetically differentiable, we applied the global test for population differentiation using the log-likelihood G statistic (Goudet 1995) over 10,000 randomisations, available in the program FSTAT (Version 2.9.3.2).
For genetic assignment analysis, we used GeneClass2 (Piry et al. 2004), implementing the Bayesian assignment criterion of Rannala and Mountain (1997). This criterion was recommended by Cornuet et al. (1999) as the best of a set of assignment criteria, and they specifically demonstrated high assignment accuracy for samples as small as ours (n = 10). For each post-eradication Pearl Island rat, the criterion gives posterior probabilities of finding the rat’s genotype in the pre-eradication Pearl Island population (survivor hypothesis), and of finding the rat’s genotype in the Stewart Island population (reinvader hypothesis).
We present the assignment results graphically, using a scatterplot of the log posterior probabilities for the two assignment populations. The graphical method is a powerful tool for assessing the different hypotheses. The two-dimensional scatterplot displays the clustering in the samples and the orders of magnitude spanned by the posterior probabilities. This information is lost by compressing the two-dimensional points into the one-dimensional score values provided by GeneClass2 (Eq. 4 of Piry et al. 2004). If there are missing data at some loci, however, the log-posterior probabilities of all individuals are not directly comparable (Piry et al. 2004), so they can only be plotted on the same graph if an imputation algorithm is used. Missing data occur frequently in genetic analyses, especially when opportunistic samples include specimens that were not fresh when preserved in ethanol, and methods for coping with missing data are necessary.
To construct the log posterior probabilities for an individual with missing data, we calculated the log posterior probabilities based on only the loci that were complete for the individual in question, along with the corresponding probabilities using this reduced set of loci for all individuals with complete data records. We then ranked the probabilities for the individual in question among the probabilities for the other individuals. We used the rank of the individual based on the incomplete data to impute the missing value for its log posterior probabilities in the complete-data graph. This was achieved by generating an empirical density of the complete-data log posterior probabilities for all complete-data individuals. The imputed value for the individual with missing data was given by the same quantile in the complete-data distribution as its incomplete log probability achieved in the incomplete-data distribution. Imputing missing values is used only for the visual presentation of results, and does not alter any numerical summaries of the analysis or significance values.
To investigate the accuracy of using imputation for low levels of missing data, we present results from two passes at genotyping the samples: the first with up to 4% of missing data, and the second after most of the missing loci had been successfully retyped, leaving only 0.5% missing data.
To investigate quantitatively whether post-eradication individuals could plausibly have derived from each of the assignment populations, we used the algorithm of Paetkau et al. (2004), as implemented in GeneClass2. This algorithm provides percentile rankings of each individual’s log posterior probabilities in a distribution of log posterior probabilities simulated to represent the assignment population. Here we did not impute missing values. For a given assignment population, the output percentile gives the rank of the individual’s log posterior probability among 100,000 individuals simulated to have ‘real’ genotypes from this population. High values indicate good fit to the assignment population, while values of about 0.05 and less suggest that the rat has a marginal or poor fit to the population. The percentile rankings can be interpreted as P-values against the hypothesis that the individual originates from the population in question. We used 100,000 simulations to achieve consistent percentiles to two decimal places. The output from this algorithm complements the visual plots, and should give similar conclusions.
Results
A total of 51 rat samples of the two species of interest were collected from the pre-eradication and post-eradication Pearl Island populations, and from the adjacent Stewart Island population. Trapping success was <10% per 100 trap nights. Harper (2006) describes the pre-eradication distribution of all three rat species on Pearl Island. Table 1 shows samples collected and the pattern of missing data from each of the two genotyping sessions. In the first session for R. rattus, all genotypes were missing for one locus and for one individual, so this locus and this individual were excluded from the results for this session, leaving 3.3% missing data (18/540). In the second session previous missing data (including the entire missing locus and individual) were retyped successfully, leaving 0.5% missing data (3/561). All numerical results reported use the genotype data from the second session.
Microsatellite data were polymorphic with mean numbers of alleles per locus between 3.1 and 4.0, and heterozygosity between 0.49 and 0.60, for pre-eradication Pearl and Stewart Island populations. Genetic differentiation was found between pre-eradication Pearl Island and the adjacent Stewart Island population for both species (F ST > 0.05; Table 2), including alleles exclusive to single populations. No significant departure from Hardy–Weinberg equilibrium (P > 0.05) and only minor levels of linkage disequilibrium between some locus pairs (3/181, P < 0.05) were found in populations.
The global test for population differentiation provided very strong evidence that the level of genetic differentiation observed between the pre-eradication Pearl Island and Stewart Island samples did not arise by chance alone (P < 0.0001 for both R. norvegicus and R. rattus). This gives confidence that the observed genetic differentiation is real for the purposes of discriminating between the survivor and reinvader hypotheses.
Figures 2 and 3 show the log posterior genotype probability plots, using the imputation algorithm for missing data as featured in Table 1. For each rat, the log posterior probability of finding its genotype in the pre-eradication Pearl population (‘survivor hypothesis’) is plotted against the log posterior probability of finding its genotype in the Stewart Island population (‘reinvader hypothesis’). High values on either axis indicate a good fit of the rat to the corresponding population. The plots show all samples of known origin (pre-eradication Pearl and Stewart Island samples) as well as those of unknown origin (post-eradication Pearl samples), so that the placement of the unknown samples can be seen with respect to that of the known samples. A leave-one-out procedure is used to place the samples of known origin, so that they also serve to cross-validate the assignment procedure.

Log posterior probability plot for R. norvegicus from Pearl Island before and after eradication, and on adjacent Stewart Island. Crosses indicate post-eradication rats to be assigned. Plots show a results from genotyping session 1, where the imputation algorithm was applied with 4% missing data involving 6 individuals (Table 1); and b results from genotyping session 2, where the imputation algorithm was used for one missing locus of one Stewart Island individual (marked with a tag on plot b). Points below the solid diagonal line have greater posterior probability of belonging to Pearl Island than to Stewart Island. Points outside the dashed diagonal lines have over nine times greater posterior probability of belonging to one population than the other

Log posterior probability plot for R. rattus from Pearl Island before and after eradication, and on adjacent Stewart Island. The cross indicates the post-eradication rat to be assigned. Plots show a results from genotyping session 1, where the imputation algorithm was used for three missing loci of three individuals (Table 1); and b results from genotyping session 2, where the imputation algorithm was used for two missing loci of two Stewart Island individuals (marked with tags on plot b). Plot b involves one extra locus and one extra individual missing from plot a. Points below the solid diagonal line have greater posterior probability of belonging to Pearl Island than to Stewart Island. Points outside the dashed diagonal lines have over nine times greater posterior probability of belonging to one population than the other
Rats below the solid diagonal lines in the plots have greater posterior probability of occurring in the pre-eradication Pearl population than occurring in the Stewart Island population, and rats above the solid lines have greater posterior probability of occurring in the Stewart Island population. Points lying outside the dashed diagonal lines have over nine times greater posterior genotype probability from one population than the other. Under the Bayesian assignment criterion of Rannala and Mountain (1997) implemented in GeneClass2, all points outside the dashed lines have a GeneClass2 assignment score of 0.9 or above for the favoured population, and 0.1 or below for the other population, except for small discrepancies that might arise from using the imputation algorithm. The range of scores from 0.1 to 0.9 covers the very narrow region of the log posterior plot contained within the dashed lines, and corresponds to posterior probabilities within a single order of magnitude of each other. This is put into context by the two-dimensional plot which covers about 20 orders of magnitude, revealing that a seemingly high score of 0.9 is in fact barely discriminative between the two hypotheses.
None of the 10 R. norvegicus caught on Pearl Island following eradication were grouped genetically with the pre-eradication Pearl population (Fig. 2). Similarly, the single post-eradication R. rattus specimen was not grouped with the pre-eradication Pearl population (Fig. 3). The pre-eradication Pearl Island specimens were grouped closely together for both species, indicative of a strong genetic signal from the island populations. Stewart Island rats were more dispersed but nearly always had posterior probabilities to the left of the Pearl Island rats. Sensitivity analysis, leaving one locus out at a time, demonstrated robustness in these groupings (results not shown). Based on these plots, we believe that all individuals trapped on Pearl Island after the eradication were reinvaders.
The graphical findings corroborate the percentile rankings of Paetkau et al. (2004) given by GeneClass2 (Table 3). These rankings are P-values against the corresponding survivor or reinvader hypothesis, and should not be interpreted as membership probabilities. For all post-eradication rats, there was strong evidence against the survivor hypothesis (P < 0.01 in every case; Table 3). Evidence that the reinvading rats derived from the Stewart Island population was more ambiguous, with only five of the post-eradication R. norvegicus samples given percentile rankings greater than 0.10 (Table 3). The ambiguous fit is likely to be due to small sample sizes, and interpretation is aided by the graphical results. Figure 2b shows that the remaining five post-eradication R. norvegicus had scores on the vertical axis that were comparatively low for the Stewart Island population, but still clearly grouped together with Stewart Island rats. By contrast, Fig. 3b shows that the single post-eradication R. rattus was not grouped with either the Stewart Island or the Pearl Island samples.
We widened the spatial sampling scale for Stewart Island using data from another study to see if the single R. rattus could have come from a more distant geographical location. Figure 4 shows the results of augmenting the Stewart Island sample by 31 extra samples, from three locations between 18 and 45 km distant from Whale Passage. All conclusions from Fig. 3 remain the same when the larger sample from Stewart Island is used in Fig. 4. In particular, the small sample (n = 8) from the Whale Passage region of Stewart Island appears to capture much of the range and extent of the larger Stewart Island sample (n = 39). The pre-eradication Pearl Island population retains its genetic distinctiveness. The single post-eradication R. rattus sample remains close to the lower extreme of Stewart Island rats, but it now appears consistent with the general profile from Stewart Island.

Log posterior probability plot for R. rattus when 31 additional samples from three dispersed locations on Stewart Island are included in the Stewart Island sample. Diagonal lines are as for Fig. 2
Comparison of plots (a) and (b) in Figs. 2 and 3 demonstrates that the imputation algorithm has worked effectively, with substantive conclusions remaining the same from the first genotyping session to the second. The transition from Fig. 3a to b is more strongly affected by the addition of an extra locus and an extra individual than by the imputation algorithm.
Discussion
We have shown that genetic assignment methods can enable conclusive discrimination between survivor and reinvader hypotheses following an eradication attempt. Even with small opportunistic samples, excluding the survivor hypothesis with a high level of confidence was possible. Percentile rankings under the survivor hypothesis were zero in nearly every case, and graphical results showed that none of the post-eradication rats appeared to associate with the pre-eradication Pearl Island population. The strong genetic signal from the pre-eradication Pearl Island rats allowed for clear differentiation between Pearl and Stewart Island populations. This is an important result for managers who generally do not have the resources for extensive genetic sampling and profiling.
The majority of rats were positioned outside the dashed diagonal lines on Figs. 2 and 3, indicating that their GeneClass2 assignment scores exceed 0.9 in favour of one population or the other. The graphs show that a GeneClass2 assignment score of about 0.9 can be misleading, because it might be interpreted as strong evidence in favour of the corresponding assignment population, whereas in reality the evidence is weak or absent. Posterior probabilities that are within the same order of magnitude for both populations do not present strong evidence in favour of either, when seen in the context of about 20 orders of magnitude in the overall span of the posterior probabilities (about 10−25–10−5; Figs. 2, 3). For example, posterior probabilities from genuine pre-eradication Pearl rats cover about five orders of magnitude (10−10–10−5) when assigned to their correct population, and despite this they are reasonably tightly clustered on the graph. Additionally, it is possible for individuals to gain high GeneClass2 scores in favour of one population when the visual results suggest that their fit to both populations is poor. An example is the post-eradication R. rattus sample (Fig. 3b), which gains an assignment score of 1.0 in favour of the Stewart Island population, but does not appear to be associated with this population from these data. Information is lost by compressing the two-dimensional points into a single score value, while the graphical presentation of results retains this information.
The graphical method provides a more informative and more interpretable output than numerical summaries, and can simplify decision making for managers. Its disadvantage is the need for imputation in order to display all individuals on the same plot. However, we have shown that imputation is effective for low levels of missing data (<5% in Fig. 2). We suggest that the benefit of the visual presentation of results outweighs the disadvantage of applying imputation for low levels of missing data. Imputation should only be used where missing data are sparse and distributed at random among samples and loci. If an entire locus fails for one or more of the populations, as happened in our first genotyping session for R. rattus, the locus should be excluded from the analysis.
The majority of R. norvegicus gained moderate to high percentile rankings under the hypothesis that they were reinvaders from the adjacent population on Stewart Island (Table 3). The remaining R. norvegicus, and the single R. rattus, gained ambiguous percentile rankings for the adjacent Stewart Island population. Graphical results suggested that these rats were nonetheless consistent with the Stewart Island population profile. The hypothesis of reinvasion by swimming across Whale Passage is supported by the ecological evidence that rats were only detected on the part of Pearl Island closest to Whale Passage and most accessible by swimming. An alternative explanation is that rats were transported to Pearl Island by boat after the eradication. However, Pearl Island is a protected reserve in a remote location, and is rarely visited, having no easy landing spots. It seems most likely that all post-eradication rats were reinvaders by swimming across Whale Passage. For the R. rattus sample, Fig. 4 demonstrates that the larger Stewart Island R. rattus population is genetically diffuse and spatially homogeneous. The R. rattus reinvader could therefore have been geographically close to Pearl Island despite its observed genetic profile.
Immediate post-eradication monitoring is not usually undertaken for rat populations, but was conducted on Pearl Island because of the perceived risk of reinvasion. Survivors of rat eradications are plausible, especially in large populations where occasional individuals might withstand LD99 doses of poison (Airey and O’Connor 2003). The Allee effect suggests low-density survivors may fail to locate each other for breeding (Courchamp et al. 2008), but alternatively there is evidence that rats adapt their behaviour at low densities to overcome this effect (Russell et al. 2005). As larger and riskier eradications are undertaken, with higher probabilities of survivors or reinvasion, immediate post-eradication monitoring will become increasingly necessary.
The genetic differentiation observed between the two possible source populations suggests they were isolated to some degree prior to eradication, despite the high reinvasion rate after the eradication. This might be explained by an incumbent advantage, whereby resident animals repelled invaders prior to eradication (Granjon and Cheylan 1989). Genetic methods of assigning post-eradication individuals would have been less successful if there had been more extensive population mixing prior to eradication.
For this study, the genetic differentiation was evident even with small, opportunistically collected samples. Small samples may generate spurious allele frequency distributions, or fail to detect rare alleles which are important for assignment, however the analysis methods used here are designed to incorporate variability due to sample size, and to prevent population exclusion due to unsampled alleles. Cornuet et al. (1999) found by simulation that the Bayesian assignment criterion yielded over 80% correct assignment with samples of size n = 10, ten loci, and F ST around 0.1, for the much more difficult task of distinguishing between ten source populations. Our data have similar characteristics and only two source populations.
Sampling prior to the eradication was essential for the genetic assignment method. We urge all pest managers undertaking eradication to collect a representative spatial sample of pre-eradication individuals (Abdelkrim et al. 2007), something which is not currently included in best-practice protocols. Genetic labwork need not be undertaken unless a reinvasion occurs, in which case the genetic costs can defray the much higher costs of remedial action with unknown focus. To minimise effects of genetic drift, source populations are best sampled as soon as post-eradication individuals are detected, from as many different source locations as possible. In some cases it might not be possible to identify the exact reinvasion source, but it might still be possible to exclude other sources. In our example, exclusion of the survivor hypothesis was the result most fundamentally useful to managers.
References
Abdelkrim J, Pascal M, Calmet C, Samadi S (2005) Importance of assessing population genetic structure before eradication of invasive species: examples from insular Norway rat populations. Conserv Biol 19:1509–1518
Abdelkrim J, Pascal M, Samadi S (2007) Establishing causes of eradication failure based on genetics: Case study of ship rat eradication in Ste. Anne archipelago. Conserv Biol 21:719–730
Airey AT, O’Connor CE (2003) Consumption and efficacy of rodent baits to Norway rats. DOC Science Internal Series 148. Department of Conservation, Wellington
Berry O, Tocher MD, Sarre SD (2004) Can assignment tests measure dispersal? Mol Ecol 13:551–561
Blackburn TM, Cassey P, Duncan RP, Evans KL, Gaston KJ (2004) Avian extinction and mammalian introductions on oceanic islands. Science 305:1955–1958
Clout MN, Russell JC (2006) The eradication of mammals from New Zealand islands. In: Koike F, Clout MN, Kawamichi M, De Poorter M, Iwatsuki K (eds) Assessment and control of biological invasion risks. IUCN, Gland, Switzerland and Cambridge, U.K., and Shoukadoh Book Sellers, Kyoto, Japan, pp 127–141
Cornuet J-M, Piry S, Luikart G, Estoup A, Solignac M (1999) New methods employing multilocus genotypes to select or exclude populations as origins of individuals. Genetics 153:1989–2000
Courchamp F, Berec L, Gascoigne J (2008) Allee effects in ecology and conservation. Oxford University Press, Oxford
Davies N, Villablanca FX, Roderick GK (1999) Determining the source of individuals: multilocus genotyping in nonequilibrium population genetics. Trends Ecol Evol 14:17–21
Donlan CJ, Tershy BR, Campbell K, Cruz F (2003) Research for requiems: the need for more collaborative action in eradication of invasive species. Conserv Biol 17:1850–1851
Goudet J (1995) FSTAT (Version 1.2): a computer program to calculate F-Statistics. J Hered 86:485–486
Granjon L, Cheylan G (1989) Le sort des rats noirs (Rattus rattus) introduits sur une île, tévélé par radio-tracking. Comptes Rendus de l’Académie des Sciences, série III 309:571–575
Harper GA (2006) Habitat use by three rat species (Rattus spp.) on an island without other mammalian predators. N Z J Ecol 30:321–333
Harper GA, Dickinson KJM, Seddon PJ (2005) Habitat selection by three rat species (Rattus spp.) on Stewart Island/Rakiura, New Zealand. N Z J Ecol 29:251–260
Howald GR, Donlan CJ, Galván JP, Russell JC, Parkes J, Samaniego A, Wang Y, Veitch CR, Genovesi P, Pascal M, Saunders A, Tershy B (2007) Invasive rodent eradication on islands. Conserv Biol 21:1258–1268
Jacob HJ, Brown DM, Bunker RK, Daly MJ, Dzau VJ, Goodman A, Koike G, Kren V, Kurtz T, Lernmark Å, Levan G, Mao Y-P, Pettersson A, Pravenec M, Simon JS, Szpirer C, Szpirer J, Trolliet MR, Winer ES, Lander ES (1995) A genetic linkage map of the laboratory rat, Rattus norvegicus. Nat Genet 9:63–69
Manel S, Berthier P, Luikart G (2002) Detecting wildlife poaching: identifying the origin of individuals with Bayesian assignment tests and multilocus genotypes. Conserv Biol 16:650–659
Manel S, Gaggiotti OE, Waples RS (2005) Assignment methods: matching biological questions with appropriate techniques. Trends Ecol Evol 20:136–142
McKinney ML, Lockwood JL (1999) Biotic homogenization: a few winner replacing many losers in the next mass extinction. Trends Ecol Evol 14:450–453
Myers JH, Simberloff D, Kuris AM, Carey JR (2000) Eradication revisited: dealing with exotic species. Trends Ecol Evol 15:316–320
Paetkau D, Slade R, Burden M, Estoup A (2004) Genetic assignment methods for the direct, real-time estimation of migration rate: a simulation-based exploration of accuracy and power. Mol Ecol 13:55–65
Piry S, Alapetite A, Cornuet J-M, Paetkau D, Baudouin L, Estoup A (2004) GENECLASS2: a software for genetic assignment and first-generation migrant detection. J Hered 95:536–539
Rannala B, Mountain JL (1997) Detecting immigration by using multilocus genotypes. Proc Natl Acad Sci USA 94:9197–9201
Robertson BC, Gemmell NJ (2004) Defining eradication units to control pests. J Appl Ecol 41:1042–1048
Russell JC, Clout MN (2004) Modelling the distribution and interaction of introduced rodents on New Zealand offshore islands. Glob Ecol Biogeogr 13:497–507
Russell JC, Towns DR, Anderson SH, Clout MN (2005) Intercepting the first rat ashore. Nature 437:1107
Russell JC, Beaven BM, MacKay JWB, Towns DR, Clout MN (2008a) Testing island biosecurity systems for invasive rats. Wildl Res 35:215–221
Russell JC, Towns DR, Clout MN (2008b) Review of rat invasion biology: implications for island biosecurity. Science for Conservation 286. Department of Conservation, Wellington
Selkoe KA, Toonen RJ (2006) Microsatellites for ecologists: a practical guide to using and evaluating microsatellite markers. Ecol Lett 9:615–629
Simberloff D (2003) Eradication - preventing invasions at the outset. Weed Sci 51:247–253
Thresher RE, Kuris AM (2004) Options for managing invasive marine species. Biol Invasions 6:295–300
Vitousek PM, D’Antonio CM, Loope LL, Rejmanek M, Westbrooks R (1997) Introduced species: a significant component of human-caused global change. N Z J Ecol 21:1–16
Weir BS (1996) Genetic data analysis II: methods for discrete population genetic data. Sinauer Associates, Sunderland
Acknowledgments
This research was funded by New Zealand Department of Conservation Science Advice Funding. James Russell and Steven Miller were supported by Top Achiever Doctoral scholarships from the NZ Tertiary Education Commission, and Rachel Fewster and Hamish MacInnes were supported by a Royal Society of New Zealand Marsden grant. Thanks to Matt Hare, Richard Clayton, Brent Beaven, Miriam Ritchie, Finn Buchanan and Clint Brown of the New Zealand Department of Conservation for their support. Andy Cox, Andy Roberts and two anonymous referees provided valuable feedback on earlier versions of the manuscript.
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License (https://creativecommons.org/licenses/by-nc/2.0), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Russell, J.C., Miller, S.D., Harper, G.A. et al. Survivors or reinvaders? Using genetic assignment to identify invasive pests following eradication. Biol Invasions 12, 1747–1757 (2010). https://doi.org/10.1007/s10530-009-9586-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-009-9586-1