
Abstract
Attacks by wolves (Canis lupus) on dogs (C. familiaris) presumably are motivated both by preying and elimination of potential competitors. Regardless of these alternative motivations in wolves, the risk of attacks might be higher when the density of primary prey is low. We examined how many dogs do territorial wolves in Finland kill in relation to the population density of the most abundant ungulates, moose (Alces alces), white-tailed deer (Odocoileus virginianus), and roe deer (Capreolus capreolus). Most attacks by wolves on dogs take place in hunting with dogs. The number of wolf-killed dogs was in highly significant negative relationship to the population density of white-tailed deer and to total ungulate biomass per unit area which is largely determined by the density of white-tailed deer. Our results indicate that abundant wild prey would decrease the risk at which wolves attack dogs. On the other side of the coin prevail two hard facts which wildlife managers had to take a notice. White-tailed deer, although a potential mitigator of wolf–human conflict, is an alien species and a partner in > 6000 traffic collisions annually in Finland. One factor that seemed to increase the risk of wolf attacks on dogs is the low ungulate density in regions where moose is the only remarkable ungulate prey. Higher moose densities could decrease the risk of attacks, but on the other hand, higher densities could increase the risk of serious traffic collisions and browsing damages in forests.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Attacks on domestic animals have stayed as one of the biggest global challenges to the successful conservation of large carnivore populations (Linnell et al. 2001; Cervasi et al. 2021). Wolf (Canis lupus) attacks on domestic dogs (C. familiaris) is a small-scale human-large carnivore conflict as compared to a conflict owing depredation on sheep and other livestock, but a remarkable conflict in some regions (Fritts and Paul 1989; Kojola and Kuittinen 2002; Backeryd 2007; Olson et al. 2015; Bassi et al. 2021; Tikkunen and Kojola 2020; Trbojevic et al. 2020).
The biological environment may affect the risk of carnivore-caused damages due to the potential relationship between the abundance of wild prey population and the magnitude of damages wolves might cause by killing domestic animals. Prey scarcity hypothesis (Nelson et al. 2016) assumes that livestock depredations by wolves may increase when wild prey exist at low densities in wolves’ home ranges. Some studies indicate that high rates of livestock depredations are associated with low density of wild ungulates (Meriggi et al. 1996; Cervasi et al. 2014; Khorozyan et al. 2015; Janeiro-Oteroa et al. 2020), thus providing empirical evidence that appears to support prey scarcity hypothesis.
Domestic dogs descend from the wolf and may breed with wolves (e.g., Salvatori et al. 2020; Harmoinen et al. 2021). Far more often, dog-wolf encounters end up wolves’ fatal attacks on dogs (Fritts and Paul 1989; Kojola & Kuittinen 2002; Kojola et al. 2004; Backeryd 2007; Bassi et al. 2021; Tikkunen and Kojola 2020). Domestic dogs differ from livestock animals both in ecological and social dimensions of wolf–human conflict. Dogs share the same carnivore guild with wolves, and therefore, their principal motivation to attack on dogs is not predation for food but the elimination of potential competitors although wolves usually also eat from the dog they have killed (Fritts and Paul 1989; Kojola and Kuittinen 2002; Backeryd 2007; Bassi et al. 2021). Occasionally, dogs might even constitute a remarkable portion of wolves’ diet (Trbojevic et al. 2020). Domestic dogs, irrespective of their concrete service roles (hunting, rescue operations, livestock guarding, etc.), are usually positioned as family members and therefore often regarded as priceless by their owners (Bisi et al. 2010).
Very little is known about potential influences of prey abundance on the risk of wolf attacks on domestic dogs. Results from Estonia and eastern Finland provided evidence that risk is highest in years when prey densities are low (Kojola et al. 2022). Herein, we evaluate the prey scarcity hypothesis at the wolf territory level in Finland. The examination of incidences at the territory level potentially provides us with a lot more detailed picture about the effects of prey abundancies, and in the current study, we can also take into account the estimated number of wolves occupying the territory.
Study area
Our study area are wolf territories located in Finland outside the reindeer husbandry region from winter 2016/2017 to winter 2019/2020 (Fig. 1). Predominant landscape is a coniferous boreal forest where the primary tree species are Scots pine (Pinus sylvestris) and Norway spruce (Picea abies). Topography outside the reindeer husbandry region is relatively flat; elevation ranges between 0 and 354 m above sea level. Of wolves’ most numerous prey, moose (Alces alces) exist throughout the study area while white-tailed deer (Odocoileus virginianus) only in the southwest. Roe deer (Capreolus capreolus) distribution is wider than that of white-tailed deer, but population densities are low in the east.

Wolf-killed dogs (black dots) and territories of wolves in Finland outside the reindeer husbandry region in 2016–2020
Data
Wolves
Wolf territory boundaries (Fig. 1) were assessed from point observations for packs and twosomes (pairs) recorded by about 2000 voluntary large carnivore contact persons and locations by GPS-collared territorial wolves, and non-invasive genetics were used for separation of neighboring packs and pairs from each other (Kojola et al. 2018; Heikkinen et al. 2020). The estimated wolf population size in March increased from 150–165 (2017) to 216–246 (2020) during our study period (Heikkinen et. 2018, Kojola et al. 2018; Heikkinen et al. 2019, 2020).
Genetic monitoring was performed annually between 56 and 76% of all territories in 2017–2020 to distinguish neighboring territories from each other (Heikkinen et al. 2018, 2019, 2020; Kojola et al. 2018).
Territory delineations were multiple convex polygons (MCP) because only GPS locations would provide data convenient for more advanced home range estimates. The estimates for pack size are November estimates based on both genetic analyses from scats collected by Luke’s field technicians and volunteers and the maximal number of wolves in point observations made during winter before the end of February. The known mortality was taken into account in pack size estimation. Genetic analyses and observational data provide highly correlated estimates for the pack size (Kojola et al. 2018).
Dogs killed by wolves
We considered casualties taken place between June 1 and March 31 in 2016–2020. We ignored a few cases that took place in April and May because most born in the previous year depart their natal territory by the end of April (Kojola et al. 2006), and thereby, the number of wolves occupying the territory during these months was not possible to estimate. The conclusion is based on movement data for 29 pups collared in winter (Kojola et al. 2006). Female wolf gives birth to new pups in late April and early May (Kaartinen et al. 2010) which results in the formation of a family pack. Only the damages accepted in a formal inspection by local officials were included in our study. The cases were stored in the official register (“Riistavahinkorekisteri”; the register of wildlife damages) with dates and geographic coordinates (Fig. 1). When we related damages to prey abundance within wolf territories, we accepted only the cases that took place within the delineated wolf territories (n = 91). These cases comprised of 58% of all casualties (n = 156). Nearly all fatal attacks on dogs by wolves (97%) took place during autumn hunting seasons (2018).
Abundance of wild prey
Population densities of moose, white-tailed deer, and roe deer were autumn post-hunt estimates by hunting clubs. Prey biomass available for the wolves per unit area were estimated based on fawn/adult (deer) ratios in hunting bag and mean dressed weights of adults and calves or fawns in autumn hunting seasons. The resultant biomass estimates were coarse because the age structure in the living population is not known. Because wolf territories were manually delineated MCPs, we used biomass estimates only per areal unit while not for the total area of the territory.
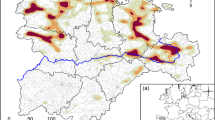
The population density of moose varied less among the wolf territories than did the density of roe deer especially the density of white-tailed deer. White-tailed deer population was highly concentrated in the southwestern wolf territories (Fig. 2). Moose existed in almost all study territories (Fig. 2). White-tailed deer was highly concentrated in southwestern Finland while roe deer was more evenly distributed in western Finland (Fig. 2). Only moose population density was normally distributed among wolf territories (Fig. 2). The mean density of moose was ca. 3.0 moose per 1000 ha. The density distribution of white-tailed deer and roe deer were biased (Fig. 2) because these deer were absent in most territories in eastern Finland (Fig. 2).

Population densities of primary prey species (ind./1000 ha) and wolf territory boundaries A and the distribution of population densities of prey species within wolf territories in 2020 B, Finland outside the reindeer husbandry region
Dogs in wolf territories
The casualties were tightly connected to hunting seasons. The number of dogs within wolf territories was not known. The registry for wildlife damages does not provide the breed of wolf-killed dog or the personal information about the owner of the dog, but an inquiry to communal authorities provided us with details for 32 cases taken place in 2017–2020. Only one of these dogs was killed by a wolf in house-yard and others in hunting with the dogs. In 20 casualties (63.2%), the wolf-killed dog was a breed used in moose hunting. Local hunting clubs report the number of moose-hunting dogs used in moose hunting per season (from Oct. 9 through Jan. 15).
Statistical analysis
A generalized linear mixed model with a negative binomial distribution assumption and log-link function was constructed to model the count response (number of wolf-killed dogs in the territory i). The hierarchy of the model consisted of two levels: wolves’ territory as the highest level and years nested within the territories. The number of years varied by the territories, and no serious temporal autocorrelations were found between the adjacent years. Thus, the residuals could be treated as uncorrelated errors instead of AR(1). The model was computed using R-package glmmTMB (Brooks et al. 2017).
In the R function glmmTMB, two variance functions were tried: NB1 \(\mathrm{variance}=\mu \left(1+\alpha \right)\) and NB2 \(\mathrm{variance}= \mu (1+\frac{\mu }{\theta })\) (Brooks et al. 2017). NB1-parametrization suggests the linear mean–variance relationship, and NB2-parametrization suggests the quadratic relationship. Both parametrizations gave almost the same AIC values in the models. The NB2-parametrization was used in the models, and the dispersion parameters of NB2 models were used in the simulations of model fit, giving directly the \(\theta\)-parameter for the simulations using R-function dnbinom.
R-package glmmTMB allowed also the zero-inflation modeling, but the zero-inflation was not an obvious problem in the data. It was tested using R-package DHRMa (Hartig 2021). The test indicated that negative binomial distribution worked without any zero-inflation coefficient in all the models. The p-values of zero-inflation tests (function testZeroInflation) remained far higher from the limit of significance at a 5% risk level. The test compares the number of simulated zeroes (function simulateResiduals) against the observed ones.
Variance inflation factor (VIF) was used to check a possible multicollinearity of the main effects of the explanatory variables. The VIF was computed using R package “performance” (Lüdecke et al. 2021) and its function check_collinearity. The predictions with standard errors were computed using R-package ggeffects (Lüdecke 2021) and the effects plots using R-package ggplot2 (Wickham 2016).
All candidate models included the estimated number of wolves occupying the territory and the number of moose-hunting dogs per area of the territory. Moose-hunting dogs are the most common breeds among attacked dogs during hunting (Kojola and Kuittinen 2002). The model-specific variable for prey abundance was (1) the population density of moose, (2) population density of white-tailed deer, (3) population density of roe deer, (4) common density of these three ungulates and (5) ungulate biomass per unit area, and (6) population densities of all these 3 ungulates. All candidate models addressed the status of the territory (pair/pack), pack size estimate (2 for pairs), and the number of moose-hunting dogs per unit area (“dog index”). Because the area of the territory was not a significant variable, it was excluded from the final models We evaluated model fit by using the Akaike information criterion (AIC) (Burnham and Anderson 2002). Models with a ΔAIC < 2 were considered equally competitive.
Results
Of six candidate models (Table 1), the annual number of wolf-killed dogs was significantly related to the number of moose-hunting dogs per area of the territory (“dog index”) in two models: the model where the pack size was treated as independent variable together with the white-tailed deer density or with densities of all three ungulates (Table 1, Fig. 3). The number of wolf-killed dogs was related positively to the pack size in all six models.

Model-based relationships of the number of wolf-killed dogs to the estimated number of wolves occupying the territory and prey abundances in Finnish wolf territories, 2016–2020. Models are shown in Table 1
The model where white-tailed deer density was the only variable for prey abundance fitted best with data (Fig. 3, model 2 in Table 1). The model where the density of white-tailed deer was recovered by the total ungulate density (model 4 in Table 1) had only a slightly poorer fit (ΔAIC = 3.2), largely because white-tail density was the primary determinant of ungulate density. The model with total ungulate biomass was not weaker than a model with a total density of ungulates (model 5 in Table 1, ΔAIC = 4.4).
The model where densities of all three ungulates were treated as independent variables (model 6 in Table 1) accounted for 47.7% of the variation in the number of dogs killed by wolves. The model with white-tailed deer density as the only variable for prey abundance accounted for 38.4% of the variation, but fit to the results was remarkably better (ΔAIC = 28.0) than that of the model with densities of all three ungulate species.
In a model where densities of all three ungulate species were entered as independent variables (model 6 in Table 1), the number of wolf-killed dogs was related significantly to the density of white-tailed deer but not to the density of moose and roe deer.
Discussion
The rate of fatal attacks by wolves on domestic dogs was related to the abundance of wild ungulate prey in Finland’s wolf territories. To what extent the risk of attacks is related to ungulate abundance remains, however, somewhat questionable because the number of dogs that are accessible to wolves within wolf territories is not explicitly known. The abundance of dogs is probably correlated with human densities that are far higher in southwestern than in eastern Finland. Hunting with dogs is a key issue here because almost all casualties were connected to hunting with dogs. Unleashed dogs are used in many kinds of hunting in Finland, most commonly hunting for moose, hare, and grouse. For roe deer, dogs are not used that often, and white-tailed deer are mostly shot from blinds at agricultural fields and feeding sites without using dogs in hunting.
Although the attacks were highly related to white-tailed deer abundance, we had to be cautious with conclusions about the risk because the data on the number of dogs that can be attacked by wolves are incomplete. The density of human residences is higher in wolf territories located in regions where white-tailed deer is abundant, and consequently, dogs are very likely less abundant outside the distribution range of this deer. Unfortunately, the official register for damages launched in 2010 did not provide records of whether the attack had taken place in house yards or in hunting situations. To have an idea of how often wolves kill dogs just in house yards in a situation where they are dependent on low-density moose population for food (Gade-Jörgensen and Stagegaard 2000; Kojola and Kuittinen 2002), we explored data recorded in North Karelia during 1998–2009 by Finnish Wildlife Agency (J. Kuittinen, unpublished). In 30 cases (26%), wolves had killed a dog in a house yard. When related to the number of wolf packs in North Karelia (cf. Kojola et al. 2014), the mean number of dogs killed in house yards was 0.61. The annual mean cases/packs ratio that took into account all cases in southwestern Finland during 2017–2022, regardless of whether they had occurred in house-yards or hunting situations, was 0.39 (I. Kojola unpublished). Although practices in the performance of hunting may differ regionally, we do not have a reason to suggest that the lower use of hunting dogs could explain why so much fewer dogs were killed by wolves within southwestern wolf territories where white-tailed deer was abundant because the use of hunting dogs differ by target species but probably much less by the region. Landscapes in southwestern wolf territories are most fragmented by farmland and highly trafficked roads (Kaartinen et al. 2015). Fragmentation may ease the location of wolves in winter, but on the other hand, the prevailing dense network of forest roads in eastern Finland which wolves commonly used as their movement routes (Gurarie et al. 2011) probably promotes the detection of wolf tracks.
We focused on casualties taken place within wolf territories. Vagrant wolves may, however, be more motivated to kill dogs than territorial wolves. The proportion of kills outside the delineated wolf territories was 42%. We do not have data on the proportion of vagrants in autumn, but their proportion in wolf populations is 10–20% without a specification for the season (Fuller et al. 2003).
Our results provided evidence that the risk to lose the dog to wolves is impacted by an abundant prey, white-tailed deer. This pattern could be explained by interference competition that increases wolves’ motivation to eliminate dogs within their territory. The attacks on dogs take place near territory boundaries much more often than expected based on the position of GPS-collared territorial wolves (Tikkunen and Kojola 2019). Such a spatial pattern fits to an explanation that intraguild competition is an important motivator for wolves to kill dogs. Regardless of the primary reason (competition, preying) moose being the only important prey could result in a higher risk of attacks.
A holistic approach in wildlife management that encompasses food chains and species interactions may limit damages large carnivores cause to domestic animals (Meriggi et al. 1996; Cervasi et al. 2014; Khorozyan et al. 2015; Janeiro-Oteroa et al. 2020). Presently high densities of ungulates cause substantial damages to forestry and agriculture in many locations in Europe (Valente et al. 2015), but at the same time, positive effects of their abundance on the viability of large carnivore populations are obvious (Chapron et al. 2014).
Our results provide an example of how a high deer density could influence the risk at which wolves attack domestic dogs. The negative relationship between the deer density and the number of dogs killed by wolves might not hold true only in our study system. In years when roe deer and wild boar (Sus scrofa) populations are down, the risk of wolves attacking dogs in house yards has been shown to be high in Estonia (Kojola et al. 2022).
Management implications
It is notifiable that high deer density might decrease wolf attacks on dogs. The overall applicability might, however, remain limited because the key species in our study system, white-tailed deer, is an alien species whose present high densities are largely owing to winter feeding by hunters. It comes expensive to the society at its present density due to damage to forestry and agriculture and being annually a partner in > 6000 traffic collisions (Statistics Finland 2022).
Decreasing the risk of wolf attacks on dogs in regions of single-species prey population in eastern Finland presumably would be a key in the mitigating conflict between wolves and hunters. Higher moose densities which could be achieved through decreasing the hunting harvest of the moose population would, on the other hand, increase the risks of traffic collisions and browsing damages to forestry.
References
Backeryd J (2007) Wolf attacks on dogs in Scandinavia 1995–2005. Will wolves in Scandinavia go extinct if dog owners are allowed to kill a wolf attacking a dog? Dissertation, University of Uppsala. https://www.slu.se/globalassets/ew/org/inst/ekol/forskning/projekt/skandulv/publikationer/studentarbeten/backeryd-2007-wolf-attacks-on-dogs-in-scandinavia-1995-2005.pdf
Bassi E, Pervan I, Ugarković D et al (2021) Attacks on hunting dogs: the case of wolf-dog interactions in Croatia. Eur J Wildl Res 67:4. https://doi.org/10.1007/s10344-020-01451-5
Bisi J, Liukkonen T, Mykrä S et al (2010) The good bad wolf – evaluation reveals the roots of the Finnish wolf conflict. Eur J Wildl Res 56:771–779. https://doi.org/10.1007/s10344-010-0374-0
Brooks ME, Kristensen K, van Benthem KJ (2017) glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R Journal 9(2):378–400
Burnham KP, Anderson DR (2002) Model selection and inference: a practical information-theoretic approach. 2nd edn. Springer-Verlag, New York. https://doi.org/10.1007/b97636
Chapron G, Kaczensky P, Linnell JDC (2014) Recovery of large carnivores in Europe’s modern human-dominated landscapes. Science 19:1517–1519. https://doi.org/10.1126/science.12575
Cervasi V, Nilsen EB, Odden J et al (2014) The spatio-temporal distribution of wild and domestic ungulates modulates lynx kill rates in a multiple-use landscape. J Zool 292:175–183. https://doi.org/10.1111/jzo.12088
Cervasi J, Linnell JDC, Boitani L et al (2021) Ecological correlates of large carnivore depredation on sheep in Europe. Global Ecol Cons 30. e0179. https://doi.org/10.1016/j.gecco.2021.e01798
Fritts SH, Paul WJ (1989) Interactions of wolves and dogs in Minnesota. Wildl Soc Bull 17:121–123. https://www.jstor.org/stable/3782636
Fuller T, Mech LD, Cochrane JF (2003) Wolf Population Dynamics. In: Mech LD, Boitani L (eds) Wolves: behavior, ecology, and conservation. University of Chicago Press, Chicago
Gade-Jørgensen I, Stagegaard R (2000) Diet composition of wolves Canis lupus in east-central Finland. Acta Theriol 45:537–547. https://rcin.org.pl/ibs/publication/29013
Gurarie E, Suutarinen J, Kojola I et al (2011) Summer movements, predation and habitat use of wolves in human modified boreal forests. Oecologia 165:891–903. https://doi.org/10.1007/s00442-010-1883-y
Harmoinen J, von Thaden A, Aspi J et al (2021) Reliable wolf-dog hybrid detection in Europe using a reduced SNP panel developed for non-invasively collected samples. BMC Genomics 22:473. https://doi.org/10.1186/s12864-021-07761-5
Hartig F (2021) DHARMa: residual diagnostics for hierarchical (multi-level / mixed) regression models. Rpackage version 0.4.4. https://CRAN.R-project.org/package=DHARMa
Heikkinen S, Kojola I, Mäntyniemi S et al (2018) Susikanta Suomessa maaliskuussa 2018. Luonnonvara- ja biotalouden tutkimus 27/2018. Natural Resources Institute Finland. http://wordpress1.luke.fi/riistahavainnot-suurpedot/wp-content/uploads/sites/4/2018/06/Susi_luke-luobio_27_2018.pdf
Heikkinen S, Kojola I, Mäntyniemi S et al (2019) Susikanta Suomessa maaliskuussa 2019. Luonnonvara- ja biotalouden tutkimus 35/2019. Natural Resources Institute Finland. https://jukuri.luke.fi/bitstream/handle/10024/544158/luke-luobio_35_2019.pdf?sequence=1&isAllowed=y
Heikkinen S, Kojola I, Mäntyniemi S et al (2020) Susikanta Suomessa maaliskuussa 2020. Luonnonvara- ja biotalouden tutkimus 37/2020. Natural Resources Institute Finland. https://jukuri.luke.fi/bitstream/handle/10024/545931/luke-luobio_37_2020.pdf?sequence=4&isAllowed=y
Janeiro-Otero A, Newsome TM, van Eeden LM (2020) Grey wolf (Canis lupus) predation on livestock in relation to prey availability. Biol Cons 243:108433. https://doi.org/10.1016/j.biocon.2020.108433.
Kaartinen S, Luoto M, Kojola I (2010) Selection of den sites by wolf in Finland. J Zool 281:99–104. https://doi.org/10.1111/j.1469-7998.2009.00678.x
Kaartinen S, Antikainen H, Kojola I (2015) Habitat model for a recolonizing wolf population in Finland. Ann Zool Fenn 52:77–89. https://doi.org/10.5735/086.052.0207
Khorozyan I, Ghoddousi A, Scoofi M et al (2015) Big cats kill more livestock when wild prey reaches a minimum threshold. Biol Cons 192:268–275. https://doi.org/10.1016/j.biocon.2015.09.031
Kojola I, Kuittinen J (2002) Wolf attacks on dogs in Finland. Wildl Soc Bull 30:498–501. http://www.jstor.org/stable/3784509
Kojola I, Ronkainen S, Hakala A, Heikkinen S, Kokko S (2004) Interactions between wolves Canis lupus and dogs C. familiaris in Finland. Wildl Biol 10:101–105. https://doi.org/10.2981/wlb.2004.014
Kojola I, Aspi J, Hakala A (2006) Dispersal in expanding wolf population in Finland. J Mamm 87:1–86. https://doi.org/10.1644/05-MAMM-A-061R2.1
Kojola I, Helle P, Heikkinen S, Linden H, Paasivaara A, Wikman M (2014) Tracks in snow and population size estimation: the wolf Canis lupus in Finland. Wildlife Biol 20:279–284. https://doi.org/10.2981/wlb.00042
Kojola I, Heikkinen S, Holmala K (2018) Balancing costs and confidence: volunteer-provided point observations, GPS telemetry and genetic monitoring of Finland’s wolves. Mamm Res 63:415–423. https://doi.org/10.1007/s13364-018-0371-3
Kojola I, Hallikainen V, Kübarsepp M, Männil P, Tikkunen M, Heikkinen S (2022) Does prey scarcity increase the risk of wolf attacks on domestic dogs? Wildlife Biol 2022:e01038. https://doi.org/10.1002/wlb3.01038
Linnell JDC, Swenson JE, Anderson R (2001) Predators and people: conservation of large carnivores is possible at high human densities if management policy is favourable. Anim Cons 4:345–349. https://doi.org/10.1017/S1367943001001408
Lüdecke D, Ben-Shachar MS, Patil I, Waggoner P, Makowski D (2021) Performance: an R package for assessment, comparison and testing of statistical models. J Open Sour Softw 6(60):3139. https://doi.org/10.21105/joss.03139
Meriggi A, Brangi A, Matteucci C (1996) The feeding habits of wolves in relation to large prey availability in northern Italy. Ecography 19:287–295. https://doi.org/10.1111/j.1600-0587.1996.tb01256.x
Nelson AA, Kauffman MJ, Middleton AD, Jimenez MD, McWrither DE, Cerow K (2016) Native prey distribution and migration mediates wolf (Canis lupus) predation on domestic livestock in the Greater Yellowstone Ecosystem. Can J Zool 94:291–299. https://doi.org/10.1139/cjz-2015-0094
Olson ER, Van Deelen TR, Wydeven AP, Ventura SJ, MacFarland DM (2015) Characterizing wolf human-conflict in Wisconsin. USA Wildl Soc Bull 39:676–688. https://doi.org/10.1002/wsb.606
Salvatori V, Donfrancesco V, Trouwborst A, Boitani L, Linnell JDC, Alvares F et al (2020) European agreements for nature conservation need to explicitly address wolf-dog hybridisation. Biol Cons 248:108525. https://doi.org/10.1016/j.biocon.2020.108525
Statistics Finland (2022) Riistaonnettomuudet (Traffic accidents with game animals). https://www.stat.fi/tup/kokeelliset-tilastot/riistaonnettomuudet/index.html. Accessed 8 September 2022
Tikkunen M, Kojola I (2019) Hunting dogs are at biggest risk to get attacked by wolves near wolves’ territory boundaries. Mamm Res 64:581–586. https://doi.org/10.1007/s13364-019-00444-3
Tikkunen M, Kojola I (2020) Does public information about wolf (Canis lupus) movements decrease wolf attacks on hunting dogs (C. familiaris)? Nat Conserv 42:33–49. https://doi.org/10.3897/natureconservation.42.48314
Trbojevic I, Penezic A, Kusak J, Stevanovic O, Cirovic D (2020) Wolf diet and livestock depredation in North Bosnia and Herzegovina. Mamm Biol 100:499–504. https://doi.org/10.1007/s42991-020-00053-7
Valente AM, Acevedo P, Figueiredo AM (2015) Overabundant wild ungulate populations in Europe: management with consideration of socio-ecological consequences. Mamm Rev 50:343–366. https://doi.org/10.1111/mam.12202
Wickham H (2016) ggplot2: Elegant Graphics for Data Analysis. Springer-Verlag, New York, p 211. ISBN: 978–0–387–08140–6
Acknowledgements
We are grateful to field technicians Antti Härkälä, Leo Korhonen, Esa Leinonen, Reima Ovaskainen, Seppo Ronkainen, and Tapio Visuri for collaring wolves and coordinating DNA sampling. Volunteering people are acknowledged for DNA sampling and wolf observations. We also want to thank Finnish hunting clubs for ungulate density estimates.
Funding
Open access funding provided by Natural Resources Institute Finland (LUKE).
Author information
Authors and Affiliations
Contributions
All authors contributed to the study’s conception and design. Material preparation, data collection, and analysis were performed by Ville Hallikainen, Vesa Nivala, Samuli Heikkinen, Mari Tikkunen, Esa Huhta, Leena Ruha, and Jyrki Pusenius. The first draft of the manuscript was written by Ilpo Kojola, and all authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Kojola, I., Hallikainen, V., Nivala, V. et al. Wolf attacks on hunting dogs are negatively related to prey abundance in Finland: an analysis at the wolf territory level. Eur J Wildl Res 69, 26 (2023). https://doi.org/10.1007/s10344-023-01652-8
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10344-023-01652-8