
Abstract
Despite the soil could contain high amount of phosphorus (P), salinity reduce its availability for crop plants. Hence, farmers should practice several tactics to ameliorate P deficiency in soils. The current study aimed to assess the importance of zinc (Zn) supply for mitigating the deficiency of P for canola grown in saline soil. The effects of three Zn rates (0, 150 and 300 mg L−1, Zn0, Zn150 and Zn300, respectively) under three P rates (0, 36 and 72 kg P2O5 ha−1, P0, P36, and P72, respectively) on physiological status, yield and quality of canola were measured. Treatments were arranged in the strip plot design based on completely randomized blocks with three replicates. Findings exhibited that P36 recorded the highest values of membrane stability index in the 2nd season, while statistically leveled P72 for relative water content and chlorophyll fluorescence in both seasons. Zn300 exhibited potent effect on all canola physiological traits in both seasons. In both seasons, P36 × Zn300, P72 × Zn150 and P72 × Zn300 showed the maximum chlorophyll fluorescence and performance index values. Plots treated with P72 achieved 70.0% increase in canola seed yield, greater than the untreated ones. Seed yield obtained with Zn300 were higher than Zn0 and Zn150 by1.30 and 1.10 times in 2019/20 season and 1.23 and 1.05 times in 2020/21 season. The highest oil % was recorded with P0 × Zn150 and P72 × Zn0 in the 1st season and with P72 × Zn150 in the 2nd season.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Globally, canola or rapeseed (Brassica napus L.) is ordered as the third oil crop after oil palm and soybean for oil production with ~76.0 million tons, obtained from ~35.0 million hectares (FAO 2019). Since its oil is fortified by a high amount of oleic acid as a main unsaturated fatty acid, canola seeds are typified by high quality and quantity of oil (Zhou et al. 2019; Mamnabi et al. 2020). According to the cultivated genotype, canola oil comprises 7% saturated fatty acids, 66% monounsaturated fatty acids, and 27% polyunsaturated fatty acids (Safavi Fard et al. 2018). The quality of fatty acid in oilseed crops is mainly depends on the environment and genotype (Enjalbert et al. 2013; Safavi Fard et al. 2018).
Production and cultivation of agricultural crops are hugely influenced by numerous eco-stresses involving drought, salinity, extreme temperatures, and nutrient deficiency (Rady et al. 2020; Saudy et al. 2020a; Abd El-Mageed et al. 2021; El-Metwally and Saudy 2021a; Abou Tahoun et al. 2022; El-Bially et al. 2022a). It has been documented that such stresses adversely affect crop growth and development, hence the productivity and quality (Mubarak et al. 2021).
Soil salinity is one of the most distinctive abiotic stresses causes major reductions in cultivatable lands and crop yield and quality (Salem et al. 2021; Abd El-Mageed et al. 2022; Al-Elwany et al. 2022; Shaaban et al. 2022). It is forecasted that approximately 50% of all agricultural lands will be impacted by salinity by 2050 (Shrivastava and Kumar 2015). Consequently, owing to soil salinity, more than US$12 billion global losses yearly due to reduced crop productivity is expected (Jägermeyr and Frieler 2018). Thus, it is important to recognize the crop responses to salinity stress to reduce the economic loss and save food security. At the plant cellular level, salinization causes excessive accumulation of salt ions in soil resulting in toxicity and osmotic effects of the soil around the roots of the plant. Due to high osmotic potential in soil rhizosphere, the ability of the crop plants to absorb soil water is reduced (Machado and Serralheiro 2017). Furthermore, salt stress including osmotic and ionic stress interposes with cellular functions of plants owing to activating production of reactive oxygen species (ROS), which result in oxidative damage in various cellular complexes (Gupta and Huang 2014) with altering vital metabolic processes (Liang et al. 2018). Also, lipid peroxidation rated increased with salinity (Hernández 2019; Yu et al. 2020), hence higher membrane permeability and loss of ions from the cells occur (Gupta and Huang 2014). Accordingly, severe agricultural concerns can be emerged due to the limited plant productivity associated with soil salinity (Isayenkov and Maathuis 2019). To ameliorate the adverse impacts of salinity on crop plants, several tactics should be adopted.
Macronutrients play a crucial role in plant growth and development and productivity (Abd-Elrahman et al. 2022; Elgala et al. 2022; Saudy and El-Metwally 2022). Of these, phosphorus (P) is an essential element for enhancing yield and quality (Saudy and El-Metwally 2019; Saudy et al. 2020b). However, P availability in soil is substantially influenced by soil condition, resulting in low P accumulation in the economic product of the crops (Salem et al. 2022). The exposure of P to fixation in soils represents a critical issue. In this regard, conversion of about 80% of P fertilizers soil application was observed to be in unavailable form (Walpola and Yoon 2012). For instance, in calcareous or normal soils and acidic soils, P converted into an insoluble complex (Satyaprakash et al. 2017; Kumar et al. 2018), causing P deficiency (Saudy et al. 2022c). Lack of P affects the normal plant growth and brings about premature death of older leaves (Niu et al. 2013).
Being it is an activator of phosphoenolpyruvate carboxylase and ribulose diphosphate carboxylase and oxygenase, zinc (Zn) increased photosynthesis rate and photo-assimilates translocation and proteins synthesis (Olama et al. 2014; Ebrahimian et al. 2017; Manaf et al. 2019; Afsahi et al. 2020). Hence, protein and oil content of oilseed crops were stimulated by Zn supply (Weisany et al. 2014; Shahsavari and Dadrasnia 2016). The biosynthesis of enzymes such as carbonic anhydrase, alcohol dehydrogenase, and superoxide dismutase (Cakmak 2000) as well as indole-3-acetic acid (Fang et al. 2008) cannot be completed without Zn. A physiological stress was reported in plants subjected to Zn deficiency (Vojodi Mehrabani et al. 2018). While, under normal and stressed conditions Zn exhibited significant improvements in crop growth and yield (El-Metwally and Saudy 2021b; Saudy et al. 2021b, 2022a).
Despite the availability of P and Zn in most cultivated soils is low, such issue could be addressed by fertilization (Montalvo et al. 2016; Saudy et al. 2020b). We hypnotized that combined application of P and Zn could adjust the nutrient balance in canola plants grown in saline soil. Therefore, physiological status and productivity of canola as influenced by P plus Zn were assessed under soil salinity conditions.
Materials and Methods
Experimental Site Description
Field trials were done in two successive winter seasons (2019/20 and 2020/21) at the field crops research station located at El-Fayoum Governorate, Egypt (latitudes: 29° 02′ and 29° 35′ N, longitudes: 30° 23′ and 31° 05′ E, and altitude: +15 m.a.s. l.). Furthermore, the main physio-chemical characteristics of the soil were measured according to Page et al. (1982) and Klute and Dirksen (1986). The soil had a loamy sand texture (71.6%), silt (16.4%), clay (12.0%) with bulk density (1.56 g cm−3), pH (7.78), electrical conductivity of saturation extract (ECe = 6.24 dS m−1), cation exchange capacity (11.2 cmol kg−1), calcium carbonate (8.3%), organic carbon (0.86%), available N, (54.3 mg kg−1 soil), available P (4.3 mg kg−1 soil), available K (43.1 mg kg−1 soil) and available Zn (0.72 mg kg−1 soil). The experimental site located in an arid region with moderate winters and rare precipitation.
Experimental Treatments and Crop Husbandry
In our experiment, three P fertilizer rates, i.e., 0 (P0), 36 (P36), and 72 (P72) kg P2O5 ha−1 were supplied in the form of superphosphate (15.5% P2O5), which is commonly used as P fertilizer. The P fertilizer rates were incorporated into the soil prior to planting. Plants exogenously sprayed with three concentrations of Zn in the form of ZnSO4 namely, 0 (Zn0; tap water as a control), 150 (Zn150), and 300 (Zn300) mg L−1. The experiment was in a two-factor strip plot design based on completely randomized blocks with three replicates, where the horizontal factor included the P fertilization rates and the vertical factor included foliar-applied Zn levels. The Zn concentrations were added as foliar application twice at 45 and 60 days from planting (DFP). Foliar Zn application was applied by a fan nozzle on a hand sprayer to lessen solution drift. To guarantee optimal Zn absorption into canola plant leaves, 0.1%, v/v of the non-ionic surfactant Tween® 20 was added to the foliar-sprayed Zn solution. Healthy seeds of Brassica napus L. cultivar Serw4 were obtained from the Crop Research Institute, Agricultural Research Center, Egypt. Seeds were sown by hand on the 15th and 10th of November at 5 kg seed ha−1 in hills spaced by 0.2 m on one side of 0.6 m distanced rows. Each individual plot was 4 m in length × 3 m in width forming 12 m2 net area with five rows. The thinning process was carried out at the 4‑leaf stage (25 DFP) to maintain the strongest and healthful two canola plants per hill. The surface irrigation system was used, and canola plants needed six irrigations through the growing season according to the daily reference crop evapotranspiration, totaling 3066 m3 ha−1. During soil tillage and plant growth, the recommended nitrogen (N) and potassium (K) fertilizers in form of ammonium sulphate and potassium sulphate with rates of 108 kg ha−1 and 58 kg ha−1, respectively were applied. The total amount of N fertilizer was top-dressed in three split doses at 21, 35, and 50 DFP, while the K fertilizer, in two equal applications, was added directly during the soil tillage and after thinning process, respectively.
Crop Measurements
Physiological Traits
Membrane stability index (MSI%) and relative water content (RWC%) were measured according to Premachandra et al. (1990) and Hayat et al. (2007), respectively. Chlorophyll fluorescence and photosynthetic performance index as a convenient tool to assess photosynthetic efficiency, was determined according to Maxwell and Johnson (2000) and Clark et al. (2000) by Handy PEA, Hansatech Instruments (Ltd, Kings Lynn, UK).
Seed Yield and Protein and Oil Content
During the two canola cropping seasons, measurements of seed yield and its attributes and seed quality were recorded at harvest (162 DFP). Five guarded canola plants were randomly chosen and carefully harvested from each plot to count the siliques number plant−1. Canola yield was determined by manually harvesting all the plants of three inner rows from each plot. These harvested plants were field sun-dried for three days to reduce plant moisture to the greatest extent possible before oilseed separation by threshing. Canola seed yield and weight of 1000 seeds were recorded based on 12% seed moisture content. Canola seed representative subsamples (~ 200 g from each plot) were further purified to eliminate impurities or damaged seed for seed oil and protein determination using a Zeltex ZX-50 portable seed analyzer (Zeltex Inc., Hagerstown, Maryland, USA).
Statistical Analysis
Analysis of variance using InfoStat statistical software (Di Rienzo et al. 2013) was performed to determine the impacts of phosphorus and zinc levels and their interaction on canola performance according to the strip-plot design based on completely randomized blocks. Wherever, the F-test showed significant (p ≤ 0.05) differences among mean values, the differences among treatments were compared using Duncan’s test (Steel and Torrie 1980).
Results
Physiology of Canola
Data in Table 1 illustrated that under saline soil condition, without P or Zn application, reductions in membrane stability index, relative water content, chlorophyll fluorescence and performance index were recorded in both growing seasons of 2019/20 and 2020/21. On the contrary, P supply enhanced all canola physiological parameters. Herein, high rate of P (P72) showed the maximum values of membrane stability index in the 1st season as well as relative water content, chlorophyll fluorescence and performance index in both seasons. P36 recorded the highest values of membrane stability index in the 2nd season, while statistically leveled P72 for relative water content and chlorophyll fluorescence in both seasons.
Spraying of Zn at a rate of 300 mg L−1 (Zn300) exhibited potent effect on all canola physiological traits in both seasons. However, the differences between 300 mg L−1 and 150 mg L−1 Zn did not reach the level of significance (p ≥ 0.05) for membrane stability index in the 2nd season, chlorophyll fluorescence, in the 1st season as well as relative water content and performance index in both seasons.
The interaction revealed that except membrane stability index in the 1st season, all other canola physiological traits significantly (p ≤ 0.05) affected by P × Zn treatments (Table 1). Membrane stability index was higher with P0 × Zn300, P36 × Zn0, P36 × Zn150 and P72 × Zn300 than the other combinations in the 2nd season. The combinations of P0 × Zn300, P36 × Zn150 and P72 × Zn300 (in both season); P36 × Zn300 (in the 1st season) as well as P72 × Zn0 and P72 × Zn150 (in the 2nd season) were the most efficient for increasing relative water content. In both seasons, P36 × Zn300, P72 × Zn150 and P72 × Zn300 showed the maximum chlorophyll fluorescence and performance index values. However, these combinations were statistically at par with P0 × Zn150, P0 × Zn300 and P36 × Zn0 (for chlorophyll fluorescence) and with P0 × Zn150 and P36 × Zn150 (for performance index) in the 2nd season.
Canola Yield Traits
Siliques no. plant−1, seed index and seed yield of canola significantly (p ≤ 0.05) influenced by P and Zn and their interaction in both seasons (Table 2). The main effects of for each P and Zn clarified the progressive increase in all yield traits with increase the application rate. Accordingly, as averages of the two seasons, plots treated with P72 achieved increases of 35.6, 14.8 and 70.0% in siliques no. plant−1, seed index and seed yield, respectively, greater than the untreated ones.
Zn300 possessed the maximum increases in all yield traits however, significantly equaled Zn150 for siliques no. plant−1, in the 1st season and seed index n both seasons. Seed yield of Zn300 were higher than Zn0 and Zn150 by1.30 and 1.10 times in 2019/20 season and 1.23 and 1.05 times in 2020/21 season.
Concerning the interaction, seed index and seed yield of canola significantly responded to P × Zn, while siliques no. plant−1 did not affect (Table 2). All combinations between P and Zn showed similar improvements in seed index and seed yield in both seasons, except for P0 × Zn0 and P0 × Zn150, which recorded lower values.
Canola Protein and Oil
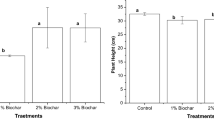
The responses of seed oil and protein content as affected by individual effects of P and Zn are illustrated in Fig. 1. P36 gave the maximum value of seed protein % in 2020/21 season. While, P72 showed the highest increase in seed oil % in both seasons statistically leveled with P36 in 2019/20 season.

Seed protein and oil content of canola plants grown in saline soil as affected by phosphorus and zinc in 2019/20 and 2020/21 seasons. P0, P36 and P72: 0, 36 and 72 kg P2O5 ha−1, respectively; Zn0, Zn150 and Zn300: 0, 150 and 300 Zn mg L−1, respectively. Values are the mean of 3 replicates ± standard errors. Means not sharing the common letters for each factor in each bar differ significantly at p ≤ 0.05
Addition of Zn300 caused the highest seed protein content in both seasons, without significant differences with Zn150 in the 2nd season. Also, Zn150 showed the maximum oil content in both seasons, significantly leveled with Zn300 in the 2nd season.
Remarkable effects of the interaction between P and Zn on seed and protein content of canola were obtained (Fig. 2). P0 × Zn150, P0 × Zn300 and P36 × Zn150 in both seasons, in addition to P72 × Zn150 and P72 × Zn300 in the 1st season as well as P36 × Zn0 and P36 × Zn300 in the 2nd season were the effective combination for increasing protein %. The highest oil % was recorded with P0 × Zn150 and P72 × Zn0 in the 1st season and with P72 × Zn150 in the 2nd season.

Seed protein and oil content of canola plants grown in saline soil as affected by the interaction of phosphorus (P) and zinc (Zn) in 2019/20 and 2020/21 seasons. P0, P36 and P72: 0, 36 and 72 kg P2O5 ha−1, respectively; Zn0, Zn150 and Zn300: 0, 150 and 300 Zn mg L−1, respectively. Values are the mean of 3 replicates ± standard errors. Means not sharing the common letters for each factor in each bar differ significantly at p ≤ 0.05
Discussion
There is no doubt that the crop plants that subjected to environmental stresses cannot grow and develop normally (El-Bially et al. 2018, 2022b; El-Metwally et al. 2022). Under abiotic stresses such as salinity, drought and heat, disturbance in the physiological status is realized (Semida et al. 2015; Saudy et al. 2021a; Abd El-Mageed et al. 2020; Makhlouf et al. 2022). Consequently, crop productivity and quality adversely influenced (El-Metwally et al. 2021; Saudy et al. 2022b). Specifically, salinity has multiple adverse effects start in the soil and extend to plant metabolism. Despite the large-scale supply of P fertilizer could increase the total quantity of P in arable lands, large amount of P is fixed in the saline soil, which is difficult to move to the crop rhizosphere. Hence, P utilization rate did not exceed 25% (Perassi and Borgnino 2014). Soil environment was affected by salinity causing nutrient lack and affected the content of available phosphorus in soil (Xie et al. 2022). In salt-affected soils (salinity or alkalinity impacts) the applied phosphorus to the soil transforms into insoluble form of phosphate with low availability (Bruland and DeMent 2009). Saline soil is typified by low nutrient ion activity, involving P and Zn, due to intemperate ratios of Na+/Ca2+, Na+/K+, Ca2+/Mg2+, and Cl−/NO3− in the soil solution, which affected the plant growth and nutrient uptake (Grattan and Grieve 1992; Bidalia et al. 2019). In sodium chloride medium, significant reduction in nitrogen, potassium and zinc was obtained (Murat et al. 2007). Additionally, high osmotic pressure and increase Na+ and Cl− inflow into root cells, generated by salinity, create shortage in the vital nutrient uptake, causing ionic imbalance in plant cells (Wang et al. 2017).
Accordingly, salinity generates another abiotic stress expressed in P deficiency. Due to such stress, membrane stability index, relative water content, chlorophyll fluorescence and performance index of canola were deteriorated under salinity. Unlike, providing the saline soil with P improved the physiological state of canola.
On the other site, by adjusting the permeability of cell membrane zinc quenches excessive Na uptake under salinity (Aktaş et al. 2006). Also, zinc decreased Na accretion and improved K/Na ratio in plants under salinity (Saleh et al. 2009; Nadeem et al. 2020). Therefore, canola cell membranes exhibited high permeability, hence leakage of some compounds from the roots under Zn deficiency. While, Zn supply improved the physiological state of canola expressed in enhancing membrane stability index, relative water content, chlorophyll fluorescence and performance index. Consequently, yield traits and seed quality were increased.
Since phosphorus level in soil is one of the significant factors affecting the zinc mobility and uptake (Wei et al. 2007), foliar application of Zn was more efficient for enhancing canola physiology and yield under P supply. Also, Zn has a crucial act in indole acetic acid biosynthesis, beginning of the primordia of the reproductive organs and metabolic reactions (Brown et al. 1993; Rehman et al. 2012), hence canola yield traits were improved.
Conclusion
Since salinity has disturbed the physiological processes of the plant, there have been quantitative and qualitative losses in the canola yield. According to the findings of the current study, the integration between P and Zn helped canola plants grown in saline soil to produce high yield with good quality oil and protein. It is worthily to note that supplying canola grown in saline soils with Zn can compensate the deficiency in P and vice versa.
References
Abd El-Mageed TA, Shaaban A, Abd El-Mageed SA, Semida WM, Rady MO (2020) Silicon defensive role in maize (Zea mays L.) against drought stress and metals-contaminated irrigation water. Silicon 13:2165–2176. https://doi.org/10.1007/s12633-020-00690-0
Abd El-Mageed TA, Rady MO, Semida WM, Shaaban A, Mekdad AA (2021) Exogenous micronutrients modulate morpho-physiological attributes, yield, and sugar quality in two salt-stressed sugar beet cultivars. J Soil Sci Plant Nutr 21:1421–1436. https://doi.org/10.1007/s42729-021-00450-y
Abd El-Mageed TA, Mekdad AAA, Rady MOA, Abdelbaky AS, Saudy HS, Shaaban A (2022) Physio-biochemical and agronomic changes of two sugar beet cultivars grown in saline soil as influenced by potassium fertilizer. J Soil Sci Plant Nutr 22:3636–3654. https://doi.org/10.1007/s42729-022-00916-7
Abd-Elrahman SH, Saudy HS, El-Fattah ADA, Hashem FA (2022) Effect of irrigation water and organic fertilizer on reducing nitrate accumulation and boosting lettuce productivity. J Soil Sci Plant Nutr 22:2144–2155. https://doi.org/10.1007/s42729-022-00799-8
Abou Tahoun AMM, Abou El-Enin MM, Mancy AG, Sheta MH, Shaaban A (2022) Integrative soil application of humic acid and foliar plant growth stimulants improves soil properties and wheat yield and quality in nutrient-poor sandy soil of a semiarid region. J Soil Sci Plant Nutr 22:2857–2287. https://doi.org/10.1007/s42729-022-00851-7
Afsahi K, Nazari M, Omidi H, Shekari F, Bostani AA (2020) The effects of different methods of zinc application on canola seed yield and oil content. J Plant Nutr 43:1070–1079. https://doi.org/10.1080/01904167.2020.1724299
Aktaş H, Abak K, Öztürk L, Çakmak I (2006) The effect of zinc on growth and shoot concentrations of sodium and potassium in pepper plants under salinity stress. Turk J Agric 30:407–412. https://doi.org/10.3906/tar-0609-2
Al-Elwany OA, Hemida KA, Abdel-Razek MA, El-Mageed ATA, El-Saadony MT, AbuQamar SF, Tarabily KA, Taha RS (2022) Impact of folic acid in modulating antioxidant activity, osmoprotectants, anatomical responses, and photosynthetic efficiency of Plectranthus amboinicus under salinity conditions. Front Plant Sci 13:887091. https://doi.org/10.3389/fpls.2022.887091
Bidalia A, Vikram K, Yamal G, Rao KS (2019) Effect of salinity on soil nutrients and plant health. In: Salt stress, microbes, and plant interactions: causes and solution. Springer, Singapore, pp 273–297
Brown PH, Cakmak I, Zhang Q (1993) Form and function of zinc in plants. Chap. 7. In: Bobson AD (ed) Zinc in soils and plants. Kluwer, Dordrecht, pp 90–106
Bruland GL, DeMent G (2009) Phosphorus sorption dynamics of Hawaii’s Coastal Wetlands. Estuaries Coast 32:844–854. https://doi.org/10.1007/s12237-009-9201-9
Cakmak I (2000) Possible roles of zinc in protecting plant cells from damage by reactive oxygen species. New Phytol 146:185–205. https://doi.org/10.1046/j.1469-8137.2000.00630.x
Clark AJ, Landolt W, Bucher JB, Strasser RJ (2000) Beech (Fagus sylvatica) response to ozone exposure assessed with a chlorophyll a fluorescence performance index. Environ Pollut 109:501–507. https://doi.org/10.1016/s0269-7491(00)00053-1
Di Rienzo JA, Casanoves F, Balzarini MG, Gonzalez L, Tablada M, Robledo CW (2013) InfoStat versión. Grupo InfoStat, FCA, Universidad Nacional de Córdoba, Córdoba (http://www.infostat.com.ar)
Ebrahimian E, Bybordi A, Seyyedi SM (2017) How nitrogen and zinc levels affect seed yield, quality, and nutrient uptake of canola irrigated with saline and ultra-saline water. Commun Soil Sci Plant Anal 48:345–355. https://doi.org/10.1080/00103624.2016.1269804
El-Bially MA, Saudy HS, El-Metwally IM, Shahin MG (2018) Efficacy of ascorbic acid as a cofactor for alleviating water deficit impacts and enhancing sunflower yield and irrigation water-use efficiency. Agric Wat Manag 208:132–139. https://doi.org/10.1016/j.agwat.2018.06.016
El-Bially MA, Saudy HS, El-Metwally IM, Shahin MG (2022a) Sunflower response to application of L‑ascorbate under thermal stress associated with different sowing dates. Gesunde Pflanz 74:87–96. https://doi.org/10.1007/s10343-021-00590-2
El-Bially MA, Saudy HS, Hashem FA, El-Gabry YA, Shahin MG (2022b) Salicylic acid as a tolerance inducer of drought stress on sunflower grown in sandy soil. Gesunde Pflanz 74:603–6013. https://doi.org/10.1007/s10343-022-00635-0
El-Metwally IM, Saudy HS (2021a) Interactional impacts of drought and weed stresses on nutritional status of seeds and water use efficiency of peanut plants grown in arid conditions. Gesunde Pflanz 73:407–416. https://doi.org/10.1007/s10343-021-00557-3
El-Metwally IM, Saudy HS (2021b) Interactive application of zinc and herbicides affects broad-leaved weeds, nutrient uptake, and yield in rice. J Soil Sci Plant Nutr 21:238–248. https://doi.org/10.1007/s42729-020-00356-1
El-Metwally IM, Saudy HS, Abdelhamid MT (2021) Efficacy of benzyladenine for compensating the reduction in soybean productivity under low water supply. Ital J Agrometeorol 2:81–90. https://doi.org/10.36253/ijam-872
El-Metwally IM, Geries L, Saudy HS (2022) Interactive effect of soil mulching and irrigation regime on yield, irrigation water use efficiency and weeds of trickle-irrigated onion. Arch Agron Soil Sci 68:1103–1116. https://doi.org/10.1080/03650340.2020.1869723
Elgala AM, Abd-Elrahman ShH, Saudy HS, Nossier MI (2022) Exploiting Eichhornia crassipes shoots extract as a natural source of nutrients for producing healthy tomato plants. Gesunde Pflanz 74:457–465. https://doi.org/10.1007/s10343-022-00622-5
Enjalbert J‑N, Zheng S, Johnson JJ, Mullen JL, Byrne PF, McKay JK (2013) Brassicaceae germplasm diversity for agronomic and seed quality traits under drought stress. Ind Crop Prod 47:176–185 (https://linkinghub.elsevier.com/retrieve/pii/S0926669013001179)
Fang Y, Wang L, Xin Z, Zhao L, An X, Hu Q (2008) Effect of foliar application of zinc, selenium, and iron fertilizers on nutrients concentration and yield of rice grain in China. J Agric Food Chem 56:2079–2084. https://doi.org/10.1021/jf800150z
FAO (2019) Food and Agriculture Organization of the United Nations Statistical Database, FAOSTAT production statistics of crops. http://www.fao.org/faostat/en/#data/QC. Accessed June 2022
Grattan SR, Grieve CM (1992) Mineral element acquisition and growth response of plants grown in saline environments. Agric Ecosyst Environ 38:275–300. https://doi.org/10.1016/0167-8809(92)90151-Z
Gupta B, Huang B (2014) Mechanism of salinity tolerance in plants: physiological, biochemical, and molecular characterization. Int J Genomics. https://doi.org/10.1155/2014/701596
Hayat S, Hayat Q, Alyemeni MN, Wani AS, Pichtel J, Ahmad A (2007) Role of proline under changing environments: a review. Plant Signal Behav 7:1456–1466. https://doi.org/10.4161/psb.21949
Hernández JA (2019) Salinity tolerance in plants: trends and perspectives. Int J Mol Sci 20:2408. https://doi.org/10.3390/ijms20102408
Isayenkov SV, Maathuis FJM (2019) Plant salinity stress: many unanswered questions remain. Front Plant Sci 10:80. https://doi.org/10.3389/fpls.2019.00080
Jägermeyr J, Frieler K (2018) Spatial variations in crop growing seasons pivotal to reproduce global fluctuations in maize and wheat yields. Sci Adv 4:eaat4517. https://doi.org/10.1126/sciadv.aat4517
Klute A, Dirksen C (1986) Hydraulic conductivity and diffusivity: laboratory methods. In: Methods of soil analysis: part 1‑physical and mineralogical methods. Soil Science Society of America, American Society of Agronomy, pp 687–734
Kumar A, Kumar A, Patel H (2018) Role of microbes in phosphorus availability and acquisition by plants. Int J Curr Microbiol Appl Sci 7:1344–1347. https://doi.org/10.20546/ijcmas.2018.705.161
Liang W, Ma X, Wan P, Liu L (2018) Plant salt-tolerance mechanism: a review. Biochem Biophys Res Commun 495:286–291. https://doi.org/10.1016/j.bbrc.2017.11.043
Machado R, Serralheiro R (2017) Soil salinity: effect on vegetable crop growth. Management practices to prevent and mitigate soil salinization. Horticulturae 3:30. https://doi.org/10.3390/horticulturae3020030
Makhlouf BSI, Khalil SRA, Saudy HS (2022) Efficacy of humic acids and chitosan for enhancing yield and sugar quality of sugar beet under moderate and severe drought. J Soil Sci Plant Nutr 22:1676–1691. https://doi.org/10.1007/s42729-022-00762-7
Mamnabi S, Nasrollahzadeh S, Ghassemi-Golezani K, Raei Y (2020) Improving yield-related physiological characteristics of spring rapeseed by integrated fertilizer management under water deficit conditions. Saudi J Biol Sci 27:797–804. https://doi.org/10.1016/j.sjbs.2020.01.008
Manaf A, Raheel M, Sher A, Sattar A, Ul-Allah S, Qayyum A, Hussain Q (2019) Interactive effect of zinc fertilization and cultivar on yield and nutritional attributes of canola (brassica napus L.). J Soil Sci Plant Nutr 19:671–677. https://doi.org/10.1007/s42729-019-00067-2
Maxwell K, Johnson GN (2000) Chlorophyll fluorescence—a practical guide. J Exp Bot 51:659–668. https://doi.org/10.1093/jexbot/51.345.659
Montalvo D, Degryse F, Silva RC, Baird R, Mclaughlin MJ (2016) Chapter five—agronomic effectiveness of zinc sources as micronutrient fertilizers. Adv Agron 139:215–267. https://doi.org/10.1016/bs.agron.2016.05.004
Mubarak M, Salem EMM, Kenawey MKM, Saudy HS (2021) Changes in calcareous soil activity, nutrient availability, and corn productivity due to the integrated effect of straw mulch and irrigation regimes. J Soil Sci Plant Nutr 21:2020–2031. https://doi.org/10.1007/s42729-021-00498-w
Murat AT, Katkat V, Suleyman T (2007) Variations in proline, chlorophyll and mineral elements contents of wheat plants grown under salinity stress. J Agron 6:137–141. https://doi.org/10.3923/ja.2007.137.141
Nadeem F, Azhar M, Anwar-ul-Haq M, Sabir M, Samreen T, Tufail A, Awan HUM, Juan W (2020) Comparative response of two rice (Oryza sativa L.) cultivars to applied zinc and manganese for mitigation of salt stress. J Soil Sci Plant Nutr 20:59–2072. https://doi.org/10.1007/s42729-020-00275-1
Niu YF, Chai RS, Jin GL, Wang H, Tang CX, Zhang YS (2013) Responses of root architecture development to low phosphorus availability: a review. Ann Bot 112:391–408. https://doi.org/10.1093/aob/mcs285
Olama V, Ronaghi A, Karimian N, Yasrebi J, Hamidi R, Tavajjoh M, Kazemi MR (2014) Seed quality and micronutrient contents and translocations in rapeseed (brassica napus L.) as affected by nitrogen and zinc fertilizers. Arch Agron Soil Sci 60:423–435. https://doi.org/10.1080/03650340.2013.796588
Page AL, Miller RH, Keeney DR (1982) Methods of soil analysis part 2. Chemical and microbiological properties. American Society of Agronomy, Madison, pp 225–246
Perassi I, Borgnino L (2014) Adsorption and surface precipitation of phosphate onto CaCO3–montmorillonite: effect of pH, ionic strength and competition with humic acid. Geoderma 232–234:600–608. https://doi.org/10.1016/j.geoderma.2014.06.017
Premachandra GS, Saneoka H, Ogata S (1990) Cell membrane stability, an indicator of drought tolerance, as affected by applied nitrogen in soyabean. J Agric Sci 115:63–66. https://doi.org/10.1017/S0021859600073925
Rady MO, Semida WM, El-Mageed ATA, Howladar SM, Shaaban A (2020) Foliage applied selenium improves photosynthetic efficiency, antioxidant potential and wheat productivity under drought stress. Int J Agric Biol 24:1293–1300. https://doi.org/10.17957/IJAB/
Rehman HU, Aziz T, Farooq M, Wakeel A, Rengel Z (2012) Zinc nutrition in rice production systems: a review. Plant Soil 361:203–226. https://doi.org/10.1007/s11104-012-1346-9
Safavi Fard N, Heidari SAH, Shirani Rad AH, Majidi Heravan E, Daneshian J (2018) Effect of drought stress on qualitative characteristics of canola cultivars in winter cultivation. Ind Crop Prod 114:87–92 (https://linkinghub.elsevier.com/retrieve/pii/S0926669018300955)
Saleh J, Maftoun M, Safarzadeh S, Gholami A (2009) Growth, mineral composition, and biochemical changes of broad bean as affected by sodium chloride and zinc levels and sources. Comm Soil Sci Plant Anal 40:3046–3060. https://doi.org/10.1080/00103620903261619
Salem EMM, Kenawey MKM, Saudy HS, Mubarak M (2021) Soil mulching and deficit irrigation effect on sustainability of nutrients availability and uptake, and productivity of maize grown in calcareous soils. Comm Soil Sci Plant Anal 52:1745–1761. https://doi.org/10.1080/00103624.2021.1892733
Salem EMM, Kenawey MKM, Saudy HS, Mubarak M (2022) Influence of silicon forms on nutrient accumulation and grain yield of wheat under water deficit conditions. Gesunde Pflanz 74:539–548. https://doi.org/10.1007/s10343-022-00629-y
Satyaprakash M, Nikitha T, Reddi EUB, Sadhana B, Vani SS (2017) A review on phosphorous and phosphate solubilizing bacteria and their role in plant nutrition. Int J Curr Microbiol Appl Sci 6:2133–2144. https://doi.org/10.20546/ijcmas.2017.604.251
Saudy HS, El-Metwally IM (2019) Nutrient utilization indices of NPK and drought management in groundnut under sandy soil conditions. Commun Soil Sci Plant Anal 50:1821–1828. https://doi.org/10.1080/00103624.2019.1635147
Saudy HS, El-Metwally IM (2022) Effect of irrigation, nitrogen sources and metribuzin on performance of maize and its weeds. Commun Soil Sci Plant Anal. https://doi.org/10.1080/00103624.2022.2109659
Saudy HS, El-Metwally IM, Abd El-Samad GA (2020a) Physio-biochemical and nutrient constituents of peanut plants under bentazone herbicide for broad-leaved weed control and water regimes in dry land areas. J Arid Land 12:630–639. https://doi.org/10.1007/s40333-020-0020-y
Saudy HS, Noureldin NA, Mubarak M, Fares W, Elsayed M (2020b) Cultivar selection as a tool for managing soil phosphorus and faba bean yield sustainability. Arch Agron Soil Sci 66:414–425. https://doi.org/10.1080/03650340.2019.1619078
Saudy HS, El-Bially MA, El-Metwally IM, Shahin MG (2021a) Physio-biochemical and agronomic response of ascorbic acid-treated sunflower (Helianthus annuus) grown at different sowing dates and under various irrigation regimes. Gesunde Pflanz 73:169–179. https://doi.org/10.1007/s10343-020-00535-1
Saudy HS, El-Metwally IM, Shahin MG (2021b) Co-application effect of herbicides and micronutrients on weeds and nutrient uptake in flooded irrigated rice: Does it have a synergistic or an antagonistic effect? Crop Prot 149:105755. https://doi.org/10.1016/j.cropro.2021.105755
Saudy HS, Abd El-Samad GA, El-Temsah ME, El-Gabry YA (2022a) Effect of iron, zinc and manganese nano-form mixture on the micronutrient recovery efficiency and seed yield response index of sesame genotypes. J Soil Sci Plant Nutr 22:732–742. https://doi.org/10.1007/s42729-021-00681-z
Saudy HS, El-Bially MA, Hashem FA, Shahin MG, El-Gabry YA (2022b) The changes in yield response factor, water use efficiency, and physiology of sunflower owing to ascorbic and citric acids application under mild deficit irrigation. Gesunde Pflanz. https://doi.org/10.1007/s10343-022-00736-w
Saudy HS, Salem EMM, Abd El-Momen WR (2022c) Effect of potassium silicate and irrigation on grain nutrient uptake and water use efficiency of wheat under calcareous soils. Gesunde Pflanz. https://doi.org/10.1007/s10343-022-00729-9
Semida WM, El-Mageed ATA, Howladar SM, Mohamed GF, Rady MM (2015) Response of solanum melongena seedlings grown under saline calcareous soil conditions to a new organo-mineral fertilizer. J Anim Plant Sci 25:485–493
Shaaban A, Al-Elwany OAAI, Abdou NM, Hemida KA, El-Sherif AMA, Abdel-Razek MA, Semida WM, Mohamed GF, El-Mageed ATA (2022) Filter mud enhanced yield and soil properties of water-stressed Lupinus termis L. in saline calcareous Soil. J Soil Sci Plant Nutr 22:1572–1588. https://doi.org/10.1007/s42729-021-00755-y
Shahsavari N, Dadrasnia A (2016) Effect of zeolites and zinc on the physiological characteristics of canola under late-season drought stress. Commun Soil Sci Plant Anal 47:2077–2087. https://doi.org/10.1080/00103624.2016.1228940
Shrivastava P, Kumar R (2015) Soil salinity: a serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation. Saudi J Biol Sci 22:123–131. https://doi.org/10.1016/j.sjbs.2014.12.001
Steel RGD, Torrie JH (1980) Principles and procedures of statistics. A biometral approach. McGraw-Hill, New York
Vojodi Mehrabani L, Hassanpouraghdam MB, Shamsi-Khotab T (2018) The effects of common and nano-zinc foliar application on the alleviation of salinity stress in Rosmarinus officinalis L. Acta Sci Polono Horto Cult 17:65–73. https://doi.org/10.24326/asphc.2018.6.7
Walpola BC, Yoon M (2012) Prospectus of phosphate solubilizing microorganisms and phosphorus availability in agricultural soils: a review. Afr J Microbiol Res 6:6600–6605. https://doi.org/10.5897/AJMR12.889
Wang Y, Stevanato P, Yu L, Zhao H, Sun X, Sun F, Li J, Geng G (2017) The physiological and metabolic changes in sugar beet seedlings under different levels of salt stress. J Plant Res 130:1079–1093. https://doi.org/10.1007/s10265-017-0964-y
Wei Y, Bai Y, Jin J, Yang L, Yao Z, Xu S, Luo G, Song W, Zhu C (2007) Sufficiency and deficiency indices of soil available zinc for rice in the alluvial soil of the coastal yellow sea. Rice Sci 14:223–228. https://doi.org/10.1016/S1672-6308(07)60031-6
Weisany W, Sohrabi Y, Heidari G, Siosemardeh A, Badakhshan H (2014) Effects of zinc application on growth, absorption and distribution of mineral nutrients under salinity stress in soybean (glycine max L.). J Plant Nutr 37:2255–2269. https://doi.org/10.1080/01904167.2014.920386
Xie W, Yang J, Gao S, Yao R, Wang X (2022) The effect and influence mechanism of soil salinity on phosphorus availability in coastal salt-affected soils. Water 14:2804. https://doi.org/10.3390/w14182804
Yu D, Boughton BA, Hill C, Feussner I, Roessner U, Rupasinghe TWT (2020) Insights into oxidized lipid modification in barley roots as an adaptation mechanism to salinity stress. Front Plant Sci 11:1. https://doi.org/10.3389/fpls.2020.00001
Zhou YJ, Chang YN, You JQ, Li SZ, Zhuang W, Cao CJ (2019) Cold-pressed canola oil reduces hepatic steatosis by modulating oxidative stress and lipid metabolism in KM mice compared with refined bleached deodorized canola oil. J Food Sci 84:1900–1908. https://doi.org/10.1111/1750-3841.14504
Funding
Open access funding provided by The Science, Technology & Innovation Funding Authority (STDF) in cooperation with The Egyptian Knowledge Bank (EKB).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
A. Shaaban, T.A.A. El-Mageed, W.R.A. El-Momen, H.S. Saudy and O.A.A.I. Al-Elwany declare that they have no competing interests.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Shaaban, A., El-Mageed, T.A.A., El-Momen, W.R.A. et al. The Integrated Application of Phosphorous and Zinc Affects the Physiological Status, Yield and Quality of Canola Grown in Phosphorus-suffered Deficiency Saline Soil. Gesunde Pflanzen 75, 1813–1821 (2023). https://doi.org/10.1007/s10343-023-00843-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10343-023-00843-2