
Abstract
Conversion of Norway spruce (Picea abies) plantations to more diverse and resilient forest types is an important task for European forest managers in the face of climate change and increased focus on ecosystem services beyond timber production. However, there is a lack of knowledge on how to cost-effectively restore such forests. This study reports the influence of vicinity (distance) of forest type (mixed or spruce), fencing and mechanical site preparation (MSP) on the early performance of planted beech (Fagus sylvatica) seedlings and natural regeneration of other tree species following clear-cuts of Norway spruce in southern Sweden. After 6 years, we found clear effects of fencing and MSP, but not of vicinity of forest type. Fencing had a positive effect on height growth of Scots pine (Pinus sylvestris) but not on height of planted beech and naturally regenerated birch (Betula pendula, B. pubescens). There was a positive effect of MSP on survival and height growth of planted beech, and on the amount of natural regeneration of Scots pine. We conclude that establishment of beech forest is greatly accelerated by active regeneration approaches such as planting. In addition, the combination of planting beech, natural regeneration of other species, fencing and MSP is effective to promote the transition to mixed and diverse stands with both broadleaves and conifers. Fencing represented the highest cost among the treatments, and its cost-effectiveness depends on the local ungulate browsing pressure. In our study, fencing was critical to protect natural regeneration of Scots pine from browsing. Finally, natural regeneration of birch was abundant in our study and relatively unaffected by fencing and MSP treatments. With time, pre-commercial thinning of the naturally regenerated birch will be needed to maintain a diverse mixture of tree species.
Similar content being viewed by others

Introduction
A large proportion of the original temperate broadleaved forest in southern Scandinavia and central Europe has been lost to other land uses during a long history of anthropogenic deforestation (Hannah et al. 1995; Gilliam 2016). Out of the remaining broadleaved forest areas, a considerable share has been replaced with conifer plantations during the past two centuries (Emmer et al. 1998; Kenk and Guehne 2001). At the beginning of the new millennium, Norway spruce (Picea abies) plantations, for example, occupied an estimated 6–7 million ha outside of its natural range, and a part of these plantations were on sites once dominated by broadleaves (Teuffel et al. 2004; Brunet et al. 2012).
Norway spruce has been the first choice for forest managers because of its easy establishment and management, rapid growth, and unpalatability to browsing animals (Emmer et al. 1998; Girdziusas et al. 2021). There is a high and stable demand for spruce and a large part of the forest industry has been based on spruce timber and pulp for many years (Eurostat 2018; Forest Europe 2020). This has resulted in much of the current forest landscape in central and northern Europe being dominated by Norway spruce (Lindbladh et al. 2014; Huuskonen et al. 2021).
In recent decades, however, Norway spruce plantations have been increasingly affected by both storm damage and severe drought, with tree mortality further increased by secondary damage caused by subsequent outbreaks of the bark beetle Ips typographus among others (Kazda and Pichler 1998; Löf et al. 2012a; Kamińska et al. 2020). Higher temperatures and associated drought due to anthropogenic climate change are probably a major factor behind the declining vitality of Norway spruce (Ols et al. 2020; Ols and Bontemps 2021). The resulting economic risks come together with increased concerns of negative effects of spruce monocultures on other ecosystem functions and services, such as soil protection, biodiversity conservation and social values, and have led to a revived interest for restoration of native broadleaved and mixed forests (Spiecker et al. 2004; Knoke et al. 2008, 2021; Holgén and Bostedt 2004; Felton et al. 2010; Löf et al. 2012a; Leidinger et al. 2021; Messier et al. 2022).
In a warmer future climate, several temperate broadleaved tree species will probably increase their performance and growth at the northern edge of their current range, while Scots pine (Pinus sylvestris) may become more competitive than Norway spruce due to its higher tolerance to drought (Thurm et al. 2018). Conversion of Norway spruce stands to more resilient forest types is thus an important task for European forest managers in the face of climate change (Spiecker et al. 2004; Messier et al. 2022; Ols and Bontemps 2021).
There are several active and passive silvicultural approaches for converting Norway spruce stands to broadleaved or mixed broadleaved-conifer stands (Kremer and Bauhus 2020). In areas where Norway spruce has been clear-cut or storm felled, wind-dispersed pioneer broadleaved tree species such as birch (Betula spp.) and aspen (Populus tremula) often establish vigorously (Karlsson 2001). However, the main target species in restoration of European hardwood forests, beech (Fagus sylvatica) and oak (Quercus robur or Q. petraea), have large and heavy seeds (acorns) which are dispersed by birds and rodents, but only to a very limited extent across longer distances. Therefore, these species are typically established through planting, either under shelterwood or nurse trees, or in larger clear-cut areas (Kenk and Guehne 2001; Lüpke et al. 2004; Ammer et al. 2008; Löf et al. 2010). However, active and passive approaches are seldom compared with regard to vicinity (distance) of seed sources, contributing to a general lack of knowledge on cost-efficient forest restoration. There is especially a lack of experiments in temperate forests regions in the subject (Reventlow et al. 2021). In addition, mechanical site preparation techniques have often been developed for plantation of conifers and more knowledge is needed for alternative tree species used during forest restoration (Löf et al. 2012b).
Another challenge for restoration practice is that population sizes of wild ungulates are increasing in Europe and so are the effects of selective browsing by multi-species deer communities on forest regeneration. Important examples are browsing damage to saplings of broadleaves and Scots pine caused by moose (Alces alces), fallow deer (Dama dama) and roe deer (Capreolus capreolus) (Olesen and Madsen 2008; Gill and Morgan 2010; Pfeffer et al. 2021). On the opposite, Norway spruce is the least preferred tree species for ungulate browsers (Kullberg and Bergström 2001; Bergquist et al. 2009; Herfindal et al. 2015). However, fences are usually needed for the successful regeneration of browsing-sensitive broadleaves (Olesen and Madsen 2008; Bergquist et al. 2009) which increases management costs.
To successfully restore mixed broadleaved forests in a cost-efficient way, we need more knowledge about the influence of vicinity of seed sources in semi-natural forest remnants on the outcome of active and passive regeneration approaches. In addition, we need more knowledge on the combined effectiveness of treatments such as fencing and mechanical site preparation for regeneration.
Our research addresses the influence of vicinity (distance) of seed sources, fencing and mechanical site preparation treatment on the early performance (establishment, survival and growth) of planted beech seedlings and natural regeneration of other tree species. The hypotheses tested were as follows: (1) the performance of regeneration improves with vicinity to semi-natural mixed stands; (2) there is a need for protection against ungulate browsing for successful regeneration; (3) mechanical site preparation improves regeneration performance. Based on our findings, we discuss new management guidelines for cost-efficient restoration of mixed forests.
Methods
Study site and experimental design
The experiment was carried out in the Ecopark Raslången near Olofström in southern Sweden (56° 15ʹ N, 14° 24ʹ E, elevation 90 m), which was established by the Swedish state forest company Sveaskog in 2011. The park is situated at the border of the nemoral and hemiboreal vegetation zones where the current spontaneous distribution ranges of European beech and Norway spruce are overlapping (Lindbladh et al. 2014). Ecoparks are large contiguous landscapes, owned by Sveaskog, where nature conservation values are prioritized over commercial forestry although part of the forest land is used for production of wood and timber. The management of Ecoparks is regulated by a formal agreement between Sveaskog and the Swedish Forest Agency with a duration of 50 years. The park covers a land area of 1243 ha of which 1200 ha is forest, including large areas of Norway spruce plantations; the remaining areas are open pasture and wetland, rocky outcrops and forest roads (Sveaskog 2011). However, the long-term restoration goal is to diversify the forest landscape by increasing the share of broadleaved forests, mainly beech and oaks (Quercus robur or Q. petraea), from ca 15% to approximately 65% of the forest area to improve conditions for biodiversity and recreation (Sveaskog 2011). Parts of the pure spruce stands were therefore clear felled in the winter of 2010–2011 to initiate regeneration of broadleaved tree species, and together with Sveaskog we started an experiment to find cost-efficient strategies for conversion of pure Norway spruce to mixed-beech forests. The experiment ran for six growing seasons from spring 2011 until winter 2016/2017.
During this period, mean annual precipitation was 592 mm, and mean air temperatures were − 0.1 °C in January and 17.5 °C in July, respectively (SMHI 2022). There was little precipitation during April and the beginning of May 2011. Otherwise, there were no weather extremes compared to the 30-year-mean values during the growing seasons.
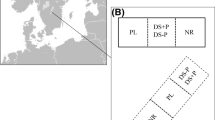
An experimental design including randomized blocks with two forest type treatments along with split-split plots in each of the four blocks was used for this study (Fig. 1). Each block was laid out in an area recently clear-cut of Norway spruce so that one side was adjacent to a semi-natural mixed broadleaved-conifer stand and the other side adjacent to a remaining pure Norway spruce stand. The size of each block on the clear-cut area was ca. 60 × 40 m (0.19 ha) and the distance between blocks was on average ca. 265 m. The adjacent mixed broadleaved stands were mostly over 60 years old and consisted mainly of birches (Betula pendula or B. pubescens), beech, European hornbeam (Carpinus betulus), Norway spruce, oaks and Scots pine (Jónsson 2016). The age of the adjacent pure spruce forest was about 35–45 years in 2011. Between the clear-cut experimental area and the mixed broadleaved stands was a gravel road in all four blocks, but the distance from the adjacent forests to the experimental areas was otherwise the same for both forest types (ca 10–15 m).

An example layout of one out of four blocks at Raslången near Olofström in southern Sweden. Main treatments in each block were forest type, i.e. mixed broadleaved-conifer (top) or pure Norway spruce (bottom). Split plots were fenced (A) or unfenced (B) and split-split plots were with mechanical site preparation (MSP) or without soil disturbance (C)
For each forest type treatment (i.e. adjacent forest type, main plots), a fenced and non-fenced treatment (split plots) and plots with and without mechanical site preparation treatment (split-split plots) were included (Fig. 1). A steel-wire fence (2 m height) was put up surrounding one-half of each block to exclude hares (Lepus timidus and Lepus europaeus) and larger herbivores such as roe deer and moose. In each split plots, mechanical site preparation (MSP) was carried out (split-split plots). For practical reasons, this was done before the fence was set up, and the fence was thereafter installed in the middle of the MSP area so that the MSP plots were always located close to the fence and therefore not completely randomized. The MSP treatment was done as inverting site preparation (Löf et al. 2012b) and carried out with an excavator, turning the soil profile upside down so that the humus layer was buried by mineral soil to a depth of 20–30 cm over a 40 × 40 cm treated spot. These split-split plots varied in size but were ca 10 × 10 m in most cases, and the MSP spots at ca. 2 m spacing. A similar-sized untreated control plot (C) was placed near each MSP plot.
Bare-rooted beech saplings were manually planted using hand tools in the beginning of May 2011. The cost per beech sapling in 2022 is 0.57 euro (planting procedure excluded). The saplings were 30–60 cm tall and 2 years old, and planted in the middle of the MSP spots and in the C plots at ca. 2 m spacing. Before fencing, mechanical site preparation and planting, slash from fallen and harvested trees was removed.
Almost no ground vegetation or natural regeneration of trees and other woody vegetation was present in the clear-cuts when the experiment started in 2011. During the study period, the ground vegetation developed and was mainly dominated by pill sedge (Carex pilulifera), wavy hair-grass (Deschampsia flexuosa), willow herb (Epilobium angustifolium), hairy wood-rush (Luzula pilosa), raspberry (Rubus idaeus) and heath speedwell (Veronica officinalis), indicating acidic and well-drained soil conditions (Jónsson 2016).
Data collection
In June 2011, ca. four weeks after planting, a first inventory of beech sapling performance (survival and growth) was carried out. Planted saplings in each treatment were observed, and we determined if they were living or dead. A sub-group of living saplings was randomly chosen for further measurements. In most plots ca. 15 living saplings were chosen, but depending on the size of each treatment plot, the numbers varied from 6 to 31 living saplings. Height (stem height from ground to highest living bud in cm) of all living saplings that were selected was then determined (also in June), and remeasured one, two, three and six growing seasons after planting in each treatment and block. Any mortality since the last measurement was noted. The final measurements of the beech were taken in January 2017, and by then, there were 524 saplings in total in the whole experiment.
Together with the last measurements of beech saplings in January 2017, we also performed measurements of the natural regeneration of all tree species. In all the split-split plots, we counted the individual saplings of Scots pine and oak and measured their individual heights. For other tree species, we used a randomly-placed 4 × 4 m subplot within each split-split plot, and in these, we measured the number of individual saplings per tree species and their individual heights. During these measurements, we noted if they had been browsed by hares or ungulates.
Calculations and statistical analysis
For fencing costs, we used a standard cost of 11.60 euro per meter (Skogforsk 2022), and assumed that one square-shaped hectare was fenced. In the calculations, we used a standard density of 2800 seedlings or seeding points per ha (Bogghed 2018). We assumed relatively high labor costs for the planting procedure, 1266 euro per ha, since larger bare-rooted saplings of broadleaves are somewhat more difficult to handle than conifer saplings (Bogghed 2018). We also assumed relatively high costs for mechanical site preparation, 387 euro per ha, since we used inverting site preparation and not normal disk trenching (Bogghed 2018). Except for fencing, no additional costs were included for natural regeneration.
The mixed procedure for tests of fixed effects in SAS was used to test differences in saplings’ height growth, survival of beech saplings and number of naturally regenerated plants after calculating treatment averages (SAS Institute Inc., Cary, NC, USA). Prior to analyses, the survival proportions of beech saplings were transformed using the formula:
where \(p^{\prime}\) is the transformed frequency, X denotes the number of living beech saplings at the end of the experiment, and n is the number of planted beech saplings at the beginning of the experiment. Differences among treatments were tested six growing seasons following planting. Prior to analyses of the browsed proportions of saplings, a similar transformation was carried out but where X denotes the number of browsed individuals, and n the total amount of individuals in 2016. Where significant F-values were calculated for browsing, the ANOVA was followed by a Tukey’s multiple range test (α = 0.05).
Results
Natural regeneration
Eight different tree species were established in the natural regeneration. The most abundant was birch, followed by Scots pine, Norway spruce, goat willow (Salix caprea), common hazel (Corylus avellana), European hornbeam, oak and gray willow (Salix cinerea). However, only birch and Scots pine were present in the majority of the experimental plots (Table 1). Therefore, the other tree species are only included in the analysis as total number of naturally regenerated trees. The average densities across all treatments of the unevenly distributed Norway spruce and goat willow were approximately 1800 and 1000 saplings per ha, respectively. The densities for the also unevenly distributed common hazel, European hornbeam, oak and gray willow were less than 100 saplings per ha in all cases. We did not find any naturally regenerated beech saplings.
Vicinity of seed sources
Six growing seasons after the start of the experiment, there were no effects of forest type (vicinity of seed sources) on performance of regeneration for any of the variables measured in this study, i.e. height growth, survival of planted beech (Table 2), amount or height of naturally regenerated seedlings (Table 3, Table S1), or browsing damage (Table S2).
Fencing
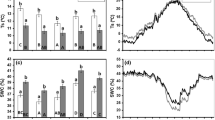
There was no effect of fencing on the height of beech saplings (Fig. 2A, Table 2). In both fenced and unfenced plots, beech saplings were on average ca. 175 cm tall following the 2016 growing season. Nor did fencing influence survival of beech saplings (Fig. 2B). Fencing did not influence the amount of natural regeneration, but 6 years after the start of the experiment, there was a positive trend (p = 0.06) of fencing on the height of natural regeneration of all species combined (Table S1). For height of birch, there was no significant effect of fencing (Fig. 3A, Table S1), whereas pine saplings were on average ca. 70 cm taller (p < 0.05) when fenced (Fig. 3B).

Mean height of beech saplings (A) and their survival (B) in four treatment combinations following planting in 2011 until 2016 in Raslången, n = 4. For description of treatments, see Fig. 1 and text

Mean height of naturally regenerated birch (A) and of naturally regenerated pine saplings (B) with or without fence and mechanical site preparation (MSP) treatments at Raslången in 2016. Mean ± SE, n = 3−4
There was a significant protective effect of fencing on browsing damage for beech (p < 0.05), and pine (p < 0.05) but not for birch (Table S2). In the unfenced plots, far more pine saplings (ca. 63%) were browsed compared to beech and birch (p < 0.05; data not shown). Only ca. 6% of the beech, and almost no birch saplings were browsed.
Mechanical site preparation
There was a positive effect (p < 0.05) of MSP treatment on beech height; saplings in MSP spots were ca. 20 cm taller than saplings planted in untreated soil (Fig. 2A, Table 2). There was also a clear positive effect (p < 0.05) from MSP treatment on beech survival (Fig. 2B, Table 2). If planted in MSP spots, approximately 75% of the beech survived the first 6 years whereas only ca. 60% survived without this treatment. This corresponds to ca. 1900 and 1500 surviving beech saplings per ha with and without MSP treatment, respectively. The positive effect from MSP treatment on survival of beech was obvious from the beginning of the experiment. For beech, there was also a significant protective effect of MSP where fewer saplings (p < 0.01) were browsed.
Concerning the amount of natural regeneration, the MSP treatment was only positive (p < 0.05) for pine (Tables 1, 3). There was no significant effect of MSP treatment on the overall height of the natural regeneration or on the height of individual species (Table S1).
Treatment costs
The alternative with only natural regeneration (no planting of bare-rooted beech saplings, fencing or MSP treatment) cost the least in our study (i.e. no additional costs; Table 4). The cost per hectare increased substantially when fencing was included whereas the contribution of the MSP treatment to overall costs was minor. Planting beech (in combination with the use of natural regeneration) including fencing and MSP treatment was the most expensive regeneration technique, at almost 8000 euro per hectare. Planting beech without fencing reduced costs by approximately 60%.
Discussion
Vicinity of seed sources
In contrast to our hypothesis, the vicinity of seed sources (older mixed broadleaf forest) did not affect the natural regeneration in our experiment. The absence of such an effect may be explained by several reasons. The mixed forest patches adjacent to the treatment were rather small and thus the amount of dispersal-limited tree species with relatively large seeds (beech, oak, hornbeam, hazel) may have been too small to produce spatial gradients during the first years. More distant larger patches of broadleaf forest may slowly contribute to seed dispersal from other directions. The time period studied, 6 years, may also have been too short for distribution gradients to develop. In addition, we cannot exclude that seed dispersal by small mammals may have been inhibited by the gravel roads located between the mixed forest patches and the study sites.
Nevertheless, our results suggest that establishment of beech forest is greatly accelerated by active regeneration approaches such as planting, probably by decades. Although there are seed sources surrounding our study sites, we did not find any natural regeneration of beech. In other cases with natural regeneration, beech typically establishes as scattered individuals under species like birch, but this process had not yet started in our experiment after 6 years.
Fencing and ungulate browsing
High browsing pressure from ungulates affects species composition and forest structure, and poses one of the greatest challenges to cost-effective restoration of mixed forests (Côté et al. 2004). Selective ungulate browsing often reduces woody species diversity, favoring a few unpalatable (e.g. Norway spruce), or browse-tolerant (e.g. hornbeam) species over most other trees and shrubs (Kuijper et al. 2010; Metslaid et al. 2013). Protection of regeneration by fencing is usually effective but comes at a high cost.
Surprisingly, fencing did not influence performance of beech in our study. This indicates a low to moderate browsing pressure on beech by deer in the study area. Browsing pressure from deer (roe, fallow or red deer) shows a strong spatial and temporal variation in southern Sweden and many other parts of Europe (Jedrzejewska et al. 1997; Wallgren et al. 2013; Kardell 2016; Pfeffer et al. 2021). Therefore, the browsing on naturally regenerated beech in an area can be an indication whether fencing may be worth the relatively high cost investment (Olesen and Madsen 2008).
On the other hand, fencing clearly favored growth of naturally regenerated pine, which supports our hypothesis concerning that species. Scots pine is a main forage for moose, and browsing decreases height growth and vitality (Edenius et al. 2002; Bergqvist et al. 2014). Our results indicate abundant natural establishment of pine on clear-cuts, further increased by MSP, but that most saplings are kept in a moose browsing trap in unfenced areas. The probability of pine escaping from browsing damage may further decrease when it is overtopped by birch (Bergqvist et al. 2014). Overtopping was common at our study sites where birch was ca. 2.5 m and pine was only ca. 60 cm tall outside fences 6 years into the experiment.
Where pine is more abundant, it is less likely to be browsed (Wallgren et al. 2013; Herfindal et al. 2015). However, in our study area, young pine is rare, and although there is abundant alternative browse which may relieve pine from being browsed (Herfindal et al. 2015), most of this alternative browse is birch which is only marginally preferred over pine by moose (Månsson et al. 2007; Herfindal et al. 2015).
Natural regeneration of birch was abundant in our study and relatively unaffected by silvicultural treatments. However, this exemplifies the need for more intensive management, i.e. pre-commercial thinning of the naturally regenerated birch (Karlsson 2001; Löf et al. 2010). Otherwise, planted seedlings of other species will risk being overtopped at an early stage and will eventually be outcompeted by the naturally regenerated birch. This is likely to be a particular problem for the more light-demanding species, such as oak and pine (Bergqvist et al. 2014), and will be reinforced by ungulate browsing as pine and oak are preferred over spruce (Edenius et al. 2002; Palmer et al. 2004; Herfindal et al. 2015).
Mechanical site preparation
Supporting our hypothesis, MSP had clear positive effects on early survival and growth of beech which is in line with previous research (Löf 2000). The spring season of 2011, when seedlings were planted, was very dry. This year resulted in a considerably higher mortality of planted saplings in untreated soil, probably due to weak root–soil contact and higher competition from ground vegetation for soil water (Löf et al. 2012b). A wet planting season would probably have resulted in a different outcome. The modest cost of soil preparation may motivate this treatment by lowering the risk of sapling mortality and further increasing cost efficiency due to accelerated sapling growth.
Climate change predictions suggest that several broadleaved tree species and also pine will become more competitive relative to Norway spruce (Thurm et al. 2018; Ols and Bontemps 2021). Management strategies to reduce and spread abiotic and biotic risks need to take this into account by implementing a mix of different site-specific management methods and silvicultural systems including the use of several tree species instead of a one-size-fits-all solution (Millar et al. 2007; Bolte et al. 2009). In our study, the negative effect of the drought after planting in spring 2011 is such an example as MSP clearly reduced mortality of the planted beech saplings.
Treatment costs
Restoration costs vary depending on methods used, from lower-cost alternatives using natural regeneration with native tree species to higher-cost approaches for active restoration using planting, mechanical site preparation and fencing. Recently, there has been a debate regarding advantages, disadvantages and the degree of success with these approaches (Meli et al. 2017; Löf et al. 2019; Banin et al. 2022). Methods need to be adapted to the local context, e.g. in terms of seed sources and browsing pressure to accomplish the restoration target. If appropriate seed sources are available, natural regeneration may represent a cost-effective option.
Implications for practice
To summarize, our results show that a combination of planting beech, mechanical site preparation and high-quality fencing effectively promotes the transition from Norway spruce plantations to mixed stands with both broadleaves and conifers, and including both light-demanding (birch, pine) and shade-tolerant (beech, spruce) tree species. Such mixed forest stands, including beech-pine mixtures as in our study, are probably less sensitive to disturbance and may recover more rapidly after storms, drought and insect outbreaks than spruce monocultures (Pretzsch et al. 2016; Huuskonen et al. 2021).
However, by planting beech supported with mechanical site preparation, mixed forests of beech and birch can be promoted without fencing and at a much lower cost. Our data suggest that the low-cost alternative without planting, and without mechanical site preparation or fencing would develop into birch-dominated forest. In such forests, spruce would eventually establish and grow, probably together with some naturally regenerated beech, while other tree species such as pine and oak would be suppressed by ungulate browsing.
We have demonstrated that it is possible to establish diverse mixed forests where Norway spruce monocultures once stood using standard silvicultural tools such as mechanical site preparation and fencing. However, higher tree diversity comes with higher costs. Our results further suggest that under high browsing pressure, pine is quickly outcompeted by naturally regenerated birch. In our fenced treatments, pine was able to remain in the developing canopy. To secure the regeneration of pine, fences need to be monitored over several years and repaired if damaged, as even brief intrusions of moose can cause great damage. On the other hand, beech regeneration does not require costly fences, but it is important to remember that this is probably very context dependent. In other areas, beech may be more difficult to establish. In clear-cuts, birch regenerated naturally and grew quickly. Birch regeneration may have some positive shelter effect for planted beech and other tree species during the first years after planting. With time, however, active management is needed to avoid negative effects of competition. The further development of the studied stands will thus depend on the choices made during pre-commercial thinning. In our case, high priority should be given to favoring promising beech and pine over birch. In addition, rare species such as oak, hornbeam, sallow and hazel should be favored during pre-commercial thinning to further increase species and habitat diversity.
Data availability
Readers may contact the corresponding author for getting access to data.
References
Ammer C, Bickel E, Kölling C (2008) Converting Norway spruce stands with beech—a review of arguments and techniques. Aust J For Sci 125:3–26
Banin LF, Raine EH, Rawland LM et al (2022) The road to recovery: a synthesis of outcomes from ecosystem restoration in tropical and sub-tropical Asian forests. Phil Trans R Soc B 378:20210090. https://doi.org/10.1098/rstb.2021.0090
Bergquist J, Löf M, Örlander G (2009) Effects of roe deer browsing and site preparation on performance of planted broadleaved and conifer seedlings when using temporary fences. Scand J For Res 24:308–317. https://doi.org/10.1080/02827580903117420
Bergqvist G, Bergström R, Wallgren M (2014) Recent browsing damage by moose on Scots pine, birch and aspen in young commercial forests—effects of forage availability, moose population density and site productivity. Silva Fenn 48:13. https://doi.org/10.14214/sf.1077
Bogghed A (2018) Skogsbrukets kostnader 2018. Norra, mellersta och södra Sverige. Rapport 2018:2, Lantmäteriet
Bolte A, Ammer C, Löf M et al (2009) Adaptive forest management in central Europe: climate change impacts, strategies and integrative concept. Scand J For Res 24:473–482. https://doi.org/10.1080/02827580903418224
Brunet J, Felton A, Lindbladh M (2012) From wooded pasture to timber production—changes in a European beech (Fagus sylvatica) forest landscape between 1840 and 2010. Scand J For Res 27:245–254. https://doi.org/10.1080/02827581.2011.633548
Côté SD, Rooney TP, Tremblay J-P et al (2004) Ecological impacts of deer overabundance. Annu Rev Ecol Evol Syst 35:113–147. https://doi.org/10.1146/annurev.ecolsys.35.021103.105725
Edenius L, Bergman M, Ericsson G et al (2002) The role of moose as a disturbance factor in managed boreal forests. Silva Fenn 36:57–67. https://doi.org/10.14214/sf.550
Emmer IM, Fanta J, Kobus AT et al (1998) Reversing borealization as a means to restore biodiversity in Central-European mountain forests—an example from the Krkonose Mountains, Czech Republic. Biodiv Conserv 7:229–247
Eurostat (2018) Industrial roundwood by species: Export in Euro
Felton A, Lindbladh M, Brunet J et al (2010) Replacing coniferous monocultures with mixed-species production stands: an assessment of the potential benefits for forest biodiversity in northern Europe. For Ecol Manag 260:939–947. https://doi.org/10.1016/j.foreco.2010.06.011
Forest Europe (2020) State of Europe’s Forests 2020. Ministerial Conference on the Protection of Forests in Europe. Bratislava
Gill RMA, Morgan G (2010) The effects of varying deer density on natural regeneration in woodlands in lowland Britain. Forestry 83:53–63. https://doi.org/10.1093/forestry/cpp031
Gilliam FS (2016) Tansley review: forest ecosystems of temperate climatic regions: from ancient use to climate change. New Phytol 212:871–887. https://doi.org/10.1111/nph.14255
Girdziusas S, Löf M, Hanssen KH et al (2021) Forest regeneration management and policy in the Nordic-Baltic region since 1900. Scand J For Res 36:513–523. https://doi.org/10.1080/02827581.2021.1992003
Hannah L, Carr JL, Lankerani A (1995) Human disturbance and natural habitat: a biome level analysis of a global data set. Biodiv Conserv 4:128–155
Herfindal I, Tremblay J-P, Hester AJ et al (2015) Associational relationships at multiple spatial scales affect forest damage by moose. For Ecol Manag 348:97–107. https://doi.org/10.1016/j.foreco.2015.03.045
Holgén P, Bostedt G (2004) Should planting of broad-leaved species be encouraged at the expense of spruce? An economic approach to a current southern Swedish forestry issue. J For Econ 10:123–134. https://doi.org/10.1016/j.jfe.2004.07.001
Huuskonen S, Domisch T, Finér L et al (2021) What is the potential for replacing monocultures with mixed-species stands to enhance ecosystem services in boreal forests in Fennoscandia? For Ecol Manag 479:118558. https://doi.org/10.1016/j.foreco.2020.118558
Jedrzejewska B, Jedrzejewski W, Bunevich AN et al (1997) Factors shaping population densities and increase rates of ungulates in Białowieza Primeval Forest (Poland and Belarus) in the 19th and 20th century. Acta Theriol 42:399–451
Jónsson JA (2016) Natural regeneration on clear-cuts in Southern Sweden—how fencing, soil treatment and distance from broad-leaved forest affect tree regeneration and ground vegetation. SLU, master thesis no 249, Southern Swedish Forest Research Centre, Alnarp
Kamińska A, Lisiewicz M, Kraszewski B et al (2020) Habitat and stand factors related to spatial dynamics of Norway spruce dieback driven by Ips typographus (L.) in the Białowieża Forest District. For Ecol Manag 476:118432. https://doi.org/10.1016/j.foreco.2020.118432
Kardell Ö (2016) Swedish forestry, forest pasture grazing by livestock, and game browsing pressure since 1900. Environ Hist 22:561–587. https://doi.org/10.3197/096734016X14727286515817
Karlsson M (2001) Natural regeneration of broadleaved tree species in southern Sweden. Ph.D. thesis, Faculty of Forest Sciences, Swedish Univ. of Agricultural Sciences
Kazda M, Pichler M (1998) Priority assessment for conversion of Norway spruce forests through introduction of broadleaf species. For Ecol Manag 102:245–258. https://doi.org/10.1016/S0378-1127(97)00166-7
Kenk G, Guehne S (2001) Management of transformation in central Europe. For Ecol Manag 151:107–119. https://doi.org/10.1016/S0378-1127(00)00701-5
Knoke T, Ammer C, Stimm B et al (2008) Admixing broadleaved to coniferous tree species: a review on yield, ecological stability and economics. Eur J For Res 127:89–101. https://doi.org/10.1007/s10342-007-0186-2
Knoke T, Gosling E, Thom D et al (2021) Economic losses from natural disturbances in Norway spruce forests—a quantification using Monte-Carlo simulations. Ecol Econ 185:107046. https://doi.org/10.1016/j.ecolecon.2021.107046
Kremer KN, Bauhus J (2020) Drivers of native species regeneration in the process of restoring natural forests from mono-specific, even-aged tree plantations: a quantitative review. Restor Ecol 28:1074–1086. https://doi.org/10.1111/rec.13247
Kuijper DP, Jędrzejewska B, Brzeziecki B et al (2010) Fluctuating ungulate density shapes tree recruitment in natural stands of the Białowieża primeval forest, Poland. J Veg Sci 21:1082–1098. https://doi.org/10.1111/j.1654-1103.2010.01217.x
Kullberg Y, Bergström R (2001) Winter browsing by large herbivores on planted deciduous seedlings in southern Sweden. Scand J For Res 16:371–378. https://doi.org/10.1080/02827580117385
Leidinger J, Blaschke M, Ehrhardt M et al (2021) Shifting tree species composition affects biodiversity of multiple taxa in Central European forests. For Ecol Manag 498:1195552. https://doi.org/10.1016/j.foreco.2021.119552
Lindbladh M, Axelsson A-L, Hultberg T et al (2014) From broadleaves to spruce—the borealization of southern Sweden. Scand J For Res 29:686–696. https://doi.org/10.1080/02827581.2014.960893
Löf M (2000) Influence of patch scarification and insect herbivory on growth and survival in Fagus sylvatica L., Picea abies L. Karst. and Quercus robur L. seedlings following a Norway spruce forest. For Ecol Manag 134:111–123. https://doi.org/10.1016/S0378-1127(99)00250-9
Löf M, Bergquist J, Brunet J et al (2010) Conversion of Norway spruce to broadleaves—regeneration systems, fencing and performance of planted seedlings. Ecol Bull 53:165–173
Löf M, Brunet J, Hickler T et al (2012a) Restoring broadleaved forests in southern Sweden as climate changes. In: Stanturf J, Madsen JP, Lamb D (eds) A goal-oriented approach to forest landscape restoration. World forests, vol 16. Springer, Dordrecht, pp 373–391
Löf M, Dey DC, Navarro RM et al (2012b) Mechanical site preparation for forest restoration. New For 43:825–848. https://doi.org/10.1007/s11056-012-9332-x
Löf M, Madsen P, Metslaid M et al (2019) Restoring forests: regeneration and ecosystem function for the future. New For 50:139–151. https://doi.org/10.1007/s11056-019-09713-0
Månsson J, Kalén C, Kjellander P et al (2007) Quantitative estimates of tree species selectivity by moose (Alces alces) in a forest landscape. Scand J For Res 22:407–414. https://doi.org/10.1080/02827580701515023
Meli P, Holl KD, Rey Benayas JM et al (2017) A global review of past land use, climate, and active vs. passive restoration effects on forest recovery. PLoS ONE 12:e0171368. https://doi.org/10.1371/journal.pone.0171368
Messier C, Bauhus J, Sousa-Silva R et al (2022) For the sake of resilience and multifunctionality, let’s diversify planted forests. Conserv Lett 15:e12829. https://doi.org/10.1111/conl.12829
Metslaid M, Palli T, Randveer T et al (2013) The condition of Scots pine stands in Lahemaa National Park, Estonia 25 years after browsing by moose (Alces alces). Boreal Environ Res 18:25–34
Millar CI, Stephenson NL, Stephens SL (2007) Climate change and forests of the future: managing in the face of uncertainty. Ecol Appl 17:2145–2151. https://doi.org/10.1890/06-1715.1
Olesen CR, Madsen P (2008) The impact of roe deer (Capreolus capreolus), seedbed and seed fall on natural beech (Fagus sylvatica) regeneration. For Ecol Manag 255:3962–3972. https://doi.org/10.1016/j.foreco.2008.03.050
Ols C, Bontemps J-D (2021) Pure and even-aged forestry of fast-growing conifers under climate change: on the need for a silvicultural paradigm shift. Environ Res Lett 16:024030. https://doi.org/10.1088/1748-9326/abd6a7
Ols C, Hervé J-C, Bontemps J-D (2020) Recent growth trends of conifers across Western Europe are controlled by thermal and water constraints and favored by forest heterogeneity. Sci Total Environ 742:140453. https://doi.org/10.1016/j.scitotenv.2020.140453
Palmer SCF, Mitchell RJ, Truscott A-W et al (2004) Regeneration failure in Atlantic oakwoods: the roles of ungulate grazing and invertebrates. For Ecol Manag 194:251–265. https://doi.org/10.1016/j.foreco.2004.01.038
Pfeffer SE, Singh NJ, Cromsigt JPGM et al (2021) Predictors of browsing damage on commercial forests—a study linking nationwide management data. For Ecol Manag 479:118597. https://doi.org/10.1016/j.foreco.2020.118597
Pretzsch H, del Rio M, Schuetze G et al (2016) Mixing of Scots pine (Pinus sylvestris L.) and European beech (Fagus sylvatica L.) enhances structural heterogeneity, and the effect increases with water availability. For Ecol Manag 373:149–166. https://doi.org/10.1016/j.foreco.2016.04.043
Reventlow DOJ, Nord-Larsen T, Biber P et al (2021) Simulating conversion of even-aged Norway spruce into uneven-aged mixed forest: effects of different scenarios on production, economy and heterogeneity. Eur J For Res 140:1005–1027. https://doi.org/10.1007/s10342-021-01381-0
Skogforsk (2022) Skogskunskap www.skogskunskap.se. Accessed 28 Aug 2022
SMHI (2022) Swedish meteorological and hydrological institute. Open data retrieved from https://www.smhi.se/data 2022-08-05 and climate station in Kristianstad ca 30 km southwest from the experimental site
Spiecker H, Hansen J, Klimo E et al (eds) (2004) Norway spruce conversion—options and consequences. European Forest Inst. Research Report 18. Brill Academic Publ
Sveaskog (2011) Ekoparksplan Raslången. Sveaskog. PDF-file retrieved from https://www.sveaskog.se 2022-10-29
Thurm EA, Hernandez L, Baltensweiler A et al (2018) Alternative tree species under climate warming in managed European forests. For Ecol Manag 430:485–497. https://doi.org/10.1016/j.foreco.2018.08.028
von Lüpke B, Ammer C, Bruciamacchie M et al (2004) Silvicultural strategies for conversion. In: Spiecker H et al (eds) Norway spruce conversion—options and consequences. European Forest Inst. Research Report 18. Brill Academic Publ., pp 121–164
von Teuffel K, Heinrich B, Baumgarten M (2004) Present distribution of secondary Norway spruce in Europe. In: Spiecker H et al (eds) Norway spruce conversion—options and consequences. European Forest Inst. Research Report 18. Brill Academic Publ., pp 63–96
Wallgren M, Bergström R, Bergqvist G et al (2013) Spatial distribution of browsing and tree damage by moose in young pine forests, with implications for the forest industry. For Ecol Manag 305:229–238. https://doi.org/10.1016/j.foreco.2013.05.057
Acknowledgements
We greatly appreciate the work with soil preparation, fencing and planting undertaken by the staff of Sveaskog AB. We thank Jan-Eric Englund for help with the statistical analyses, and Carl Salk for revising the language.
Funding
Open access funding provided by Swedish University of Agricultural Sciences. This study was supported by the Broadleaves for the Future research program within SLU. Sveaskog AB financed the establishment of the field experiment.
Author information
Authors and Affiliations
Contributions
ML and JB developed the original idea for the study. All authors jointly collected the field data. ML analyzed the data with contributions from the other authors. ML and JB wrote the manuscript, including comments by ESF and MS.
Corresponding author
Ethics declarations
Conflict of interest
The author declares that they have no competing of interest.
Additional information
Communicated by Thomas Knoke.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Table S1
Statistical analysis of effects on height of naturally regenerated saplings at Raslången in 2016 (Mixed-procedure, type 3 tests of fixed effects) (DOC 37 KB)
Table S2
Statistical analysis of effects on browsed proportions of beech, birch and pine saplings at Raslången in 2016 (Mixed-procedure, type 3 tests of fixed effects) (DOC 36 KB)
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Löf, M., Sandell Festin, E., Szydło, M. et al. Restoring mixed forests through conversion of Norway spruce stands: effects of fencing and mechanical site preparation on performance of planted beech and natural tree regeneration. Eur J Forest Res 142, 763–772 (2023). https://doi.org/10.1007/s10342-023-01554-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10342-023-01554-z