
Abstract
Biochemical and blood cell parameters have commonly been employed as useful minimally invasive indicators of the health or nutritional status of many species. Here, we present data on a suite of commonly measured blood parameters from 342 White Stork Ciconia ciconia chicks of molecularly known sex. Samples were collected in western Poland during four breeding seasons (2005–2008). We examine whether sex, and also season (year), nesting locality, hatch date and brood size, contributed to variation in these blood parameters. There were significant seasonal differences in levels of the heterophil/lymphocyte ratio, red blood cell count, haematocrit, haemoglobin, mean corpuscular volume, mean corpuscular haemoglobin concentration, and particularly in white blood cell count. Significant nesting locality effects were detected in haemoglobin, mean corpuscular volume, mean corpuscular haemoglobin and mean corpuscular haemoglobin concentration. Hatch date, virtually synonymous with chick age, significantly negatively influenced (i.e. later hatch dates (younger chicks when sampled) had lower values of red blood cell count, haematocrit, haemoglobin and mean corpuscular haemoglobin concentration. Male chicks had significantly lower levels of red blood cell count, haematocrit, haemoglobin and mean corpuscular haemoglobin concentration. There was a significant sex/hatch date interaction for red blood cell count, haematocrit and haemoglobin. Brood size did not significantly affect any of the analysed parameters. Our work shows that blood parameters are influenced by many factors, including sex and age of individuals, and such factors may need to be taken into consideration when using blood parameters as indicators of health. Such considerations are especially critical in the establishment of reference ranges of blood parameters which may be of use in captive rearing of this endangered species.
Zusammenfassung
Geschlecht und andere Ursachender Variation hämatologischer Parameter bei Weißstorch-Nestlingen ( Ciconia ciconia )
Biochemische und blutzellspezifische Parameter werden häufig als hilfreiche minimalinvasive Indikatoren für den Gesundheits- oder Ernährungsstatus bei vielen Arten untersucht. In der vorliegenden Arbeit stellen wir die Werte von Standardblutparametern von 342 Nestlingen des Weißstorchs Ciconia ciconia vor, deren Geschlecht molekularbiologisch bestimmt wurde. Blutproben wurden von Nestlingen in Westpolen während vier Brutzeiten (2005-2008) entnommen. Untersucht wurde, inwieweit Geschlecht, Brutzeit (Jahr), Neststandort, Schlupftermin und Größe der Brut die Blutparameter beeinflussen. Es gab signifikante saisonale Unterschiede in der Lymphozytenkonzentration, in der Anzahl roter Blutkörperchen, im Hämatokrit- und Hämoglobinwert, im mittleren Blutzellvolumen, mittleren Blutzellhämoglobinwert und speziell in der Anzahl weißer Blutkörperchen. Der Neststandort beeinflusste den Hämoglobinwert, das mittlere Blutzellvolumen und den mittleren Blutzellhämoglobinwert. Das Schlupfdatum, nahezu gleichzusetzen mit dem Nestlingsalter, hatte eine negativen Einfluss auf die Anzahl roter Blutkörperchen, den Hämatokrit- und Hämoglobinwert und den mittleren Blutzellhämoglobinwert, d. h. später geschlüpfte Nestlinge hatten kleinere Werte. Männliche Nestlinge zeigten eine signifikant geringere Anzahl roter Blutkörperchen, einen niedrigeren Hämatokrit- und Hämoglobinwert und einen durchschnittlich kleineren Blutzellhämoglobinwert. Eine signifikante Interaktion von Geschlecht und Schlupfdatum ergab sich bei der Anzahl roter Blutkörperchen und dem Hämatokrit- und Hämoglobinwert. Die Anzahl Nestlinge pro Brut beeinflusste keinen der Parameter signifikant. Unsere Studie belegt, dass Blutparameter von vielen Faktoren beeinflusst werden (inklusive Geschlecht und Alter), die bei der Nutzung der Parameter als Indikatoren des Gesundheitszustands berücksichtigt werden müssen. Dies ist besonders wichtig, falls Referenzspannweiten von Blutparametern festgelegt werden, was hilfreich bei der Aufzucht dieser bedrohten Art in Menschenobhut sein kann.
Similar content being viewed by others

Introduction
Biochemical and blood cell parameters have commonly been employed as useful indicators of the health or nutritional status of many species, and therefore influence the fitness of an organism, mainly via probability of survival (Jenni-Eiermann and Jenni 1998). For example, the most often used blood parameter, haematocrit, the percentage of the total blood volume occupied by erythrocytes upon centrifugation, has been used as an indicator of the health of both captive and free-living individuals (Dawson and Bortolotti 1997a, b; Moreno et al. 2002). There has been discussion of the true utility of this parameter in the case of birds (Dawson and Bortolotti 1997a; Bowerman et al. 2000). The haematocrit depends on the variation in plasma volume, the rate of erythrocyte production and destruction (a process in which haemolytic diseases or blood parasites may be involved), degree of hydration, toxins, and direct blood loss as a result of injury or blood-sucking ectoparasites (Dawson and Bortolotti 1997a; Campo and Davila 2002; Moreno et al. 2002; Sanz et al. 2004; Boughton et al. 2006; Jakubas et al. 2008), and it may thus be used as an index of the ‘health’ of the oxygen transport system. Haematological values of healthy individuals can be used to establish reference ranges which can subsequently be used as diagnostic tools to determine the health of free-living individuals or injured birds undergoing treatment in veterinary clinics (Tryjanowski et al. 2006). However, considerable within-population variation may exist in haematological traits and may need to be incorporated to provide useful reference values. Age has been identified as a major factor that can influence blood parameters, and others, such as sex or stage of the breeding cycle, may also be important (Kostelecka-Myrcha 1985; Pastor et al. 2001a, b; Campo and Davila 2002; Gayathri et al. 2004; Villegas et al. 2004; Baos et al. 2006b; Kasprzak et al. 2006; Jakubas et al. 2008). However, our knowledge of the influence of sex and other factors on haematological parameters, especially in chicks, is limited. It is also necessary to consider potential interactions between sex and age, both within and among life-history stages. To date, comparisons of haematology between adults and juveniles have suggested that there are age effects, at least in some haematological indices (Alonso et al. 1991). There is also often considerable unexplained variation among individuals (Dawson and Bortolotti 1997a; Campo and Davila 2002; Blas et al. 2006), potentially linked to sex, but whether this variation is indeed related to sex remains largely unknown. On the other hand, as has been seen in species with high sexual dimorphism (Kostelecka-Myrcha 1985; Bowerman et al. 2000; Gayathri et al. 2004; Villegas et al. 2004; Gil et al. 2007), we predict that some blood parameters of males and females will diverge with age due to the differential effects of sex-specific hormones. For instance, hormones may modulate aggressive behaviour among chicks, and therefore access to food provided by parents (Villegas et al. 2004; Tryjanowski et al. 2011).
In this paper, we will describe the range of haematological parameters and the factors that explain their variation within a population of wild White Stork Ciconia ciconia chicks. Previous haematological studies on the White Stork, both on free-living and captive birds, have generally been limited by sample size (Puetra et al. 1989; Alonso et al. 1991; Lashev et al. 2005) or, in a study with a more robust sample size (Montesinos et al. 1997), the authors did not investigate whether parameters differed between male and female chicks. This is understandable because, as a monomorphic species, female and male White Storks do not differ substantially in size and plumage, either during nestling development or as adults (Cwiertnia et al. 2006). Therefore, it would be difficult to detect clear sexual differences in haematological parameters, especially during the nestling stage. We recorded the following commonly analysed parameters: number of red blood cells (RBC), haematocrit (Ht), haemoglobin (Hb) content, some haematological indices (mean corpuscular volume, MCV; mean corpuscular haemoglobin, MCH; mean corpuscular haemoglobin concentration, MCHC) and number of white blood cells (WBC). We also prepared thin blood smears and determined the relative numbers of lymphocytes (L) and heterophils (H) (see “Methods”), and derived the H/L ratio. We examine how these haematological parameters vary with season (years), breeding locality, brood size, hatch date and sex.
Methods
Our study was performed from 2005 to 2008 at 65 nest sites in western Poland encompassing an area between 51°40′ and 54°38′N and 14°42′ and 17°33′E, in habitats with a relatively high density of breeding White Storks (for more details, see Tryjanowski et al. 2005; Kulczykowska et al. 2007). Nest sites were situated along the flood plain of the Oder river, and we have allocated nest sites to five contiguous downstream nesting locality categories from near Glogów in the southeast (coded 1) to near Kłopot in the northwest (coded 5). All White Storks bred in open nests on roofs, in the tops of trees, or on electricity posts. Chicks hatched after approximately 1 month of incubation, but remained in the nest and relied on both parents for food and protection for the next 70–80 days. Our study was carried out between 23 June and 4 July in each year. To control for possible diurnal variation in blood measures (see Kulczykowska et al. 2007), data were collected only during the late morning (0900–1200 hours). Mean hatch dates per year varied between May 18 and 24. Samples were collected from a total of 342 chicks from 124 broods: 44 chicks from 18 broods in 2005, 84 chicks from 34 broods in 2006, 111 chicks from 36 broods in 2007, and 103 chicks from 36 broods in 2008.
Blood sampling and laboratory procedures
White Stork chicks are very quiet and display only very little physiological response to handling (Blas et al. 2006), and the opportunity was taken to examine their physical health using traditional veterinary diagnostics (e.g. leg development, eye brightness, plumage condition, breast muscle development). All chicks appeared to be healthy; indeed, only four deaths were recorded between sampling and fledging during the 4 years of study. Blood samples were collected from nestlings (342 chicks in total, 150 females and 192 males) by brachial vein-puncture with a vacationer. Each chick was removed from the nest and placed individually in cotton bags, and blood samples were collected as soon as possible. Body mass of chicks averaged 2,860 g and was determined using a Pesola spring balance (to the nearest 50 g). Age (with accuracy 1–2 days) was estimated by measurement of the bill length following the method of Kania (1988) and hence hatch date was found by back calculation. The mean (±SE) estimated age of chicks during sampling was 37.1 ± 0.48 days, range 15–62 days, with no significant difference between years and sites. Because blood sampling was carried out during a short time window each year, hatch date is almost synonymous with chick age (r = −0.928, P < 0.001).
Immediately after blood sampling, a small amount of each sample was used to prepare a blood smear that was air dried the remainder of the sample (5 ml) was immediately mixed with the anticoagulant, dipotassium ethylenediaminetetraacetic acid (1.5 mg/dl). The samples were maintained at 4–6 °C and in the dark. Upon return to the laboratory, blood samples were processed within 12 h, and haematological analyses were performed on the day of collection by the same person (M.K.).
The number of RBC and WBC were counted in a Bürcer chamber after the blood was diluted 200 times in Natt and Herrick’s solution using haematological pipettes. Haematocrit (Ht) was determined using heparinized capillary tubes and centrifuging the blood in a microhaematocrit centrifuge at 12,000 rpm for 3 min, and expressed as the percentage of the length of the part of the capillary tube occupied by RBC in relation to the total length of the capillary tube occupied by both the cellular fraction and plasma. Haemoglobin (Hb) concentration was measured by Drabkin’s colorimetric method. Well-mixed whole blood (0.02 ml) was added to 5 ml of Drabkin’s reagent. After 10 min, this was centrifuged at 3,500 rpm to remove RBC nuclei and membranes, and the Hb was measured in the supernatant in a spectrophotometer at 540 nm wavelength (van Kampen and Zijlstra 1961).
The determination of MCV, MCHC and MCH require the determination of the packed cell volume (PCV) of RBC (see Campbell 1995). This was accomplished by measuring the height of the RBC fractions of heparinised capillary tubes with digital calipers to the nearest 0.1 mm after 10 min of centrifugation at 10,000 rpm. MCV, MCHC and MCH were then calculated from the RBC count, PCV and Hb concentration, using the following equations from Campbell (1995): MCV = (PCV/RBC) × 10; MCH = (Hb/RBC) × 10; MCHC = (Hb/PCV) × 100.
To analyse the relative abundance of L and H, the heterophil/lymphocyte ratio was calculated. The heterophil/lymphocyte (H/L) ratio is widely used as an indicator of stress in poultry and is known to increase as a response to various stressors, including infectious diseases, starvation and physiological disturbance (Maxwell 1993; Moreno et al. 2002; Sanz et al. 2004; Jakubas et al. 2008).
All parameters, except H and L levels, were collected over the whole 4-year period; H and L levels were measured only in 2005 and 2007. Due to problems with some laboratory analyses (one sample with too little blood), sample sizes differed slightly between parameters.
Because White Storks do not exhibit strong sexual dimorphism (Schulz 1998; Cwiertnia et al. 2006), we used the cellular fraction of the blood as a source of DNA to determine the sex of all birds. Whole genomic DNA was extracted using DNAeasy Blood and Tissue Kit (Qiagen, Hilden, Germany) following the manufacturer’s standard protocol. CHD-W and CHD-Z gene fragments were PCR-amplified with primers 2550F and 2718R (Fridolfsson and Ellegren 1999; and see Tryjanowski et al. 2011 for more technical details).
Statistical analyses
All measured variables were correlated to show potential relationships between study parameters, and to provide some information for comparison with older papers where the number of measured variables was more limited. Subsequently, a linear mixed model with Gaussian error structure and identity link was used to analyse each of the haematological parameters. This model included year, sex, hatch date and brood size as fixed effects and an interaction between sex and hatch date. Brood, nest site and nest region (locality) were included as random effects in a nested structure. The models were run with maximum likelihood estimation, and fixed effects were sequentially deleted from the model using likelihood ratio tests, with a threshold for inclusion of 0.05. Partial R 2 values for each fixed effect were calculated using the methods described in Edwards et al. (2008) with the Kenward–Rogers approximation of degrees of freedom (Kenward and Roger 1997). The final model was re-run using restricted maximum likelihood estimation to accurately estimate the variance associated with the random effects. H, L and H/L were analysed in 153 birds with information complete for the three variables; all other analyses included 341 birds with complete information for the remaining variables. Statistical analyses were conducted using the statistical package R (R Development Core Team 2010) and packages lme4 (Bates and Maechler 2010) and doBy (Højsgaard et al. 2010).
Results
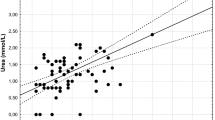
Correlations between the haematological parameters are presented in Tables 1 and 2. Large coefficients (arbitrarily taken here as >|0.8|) occurred for five pairs of indices (H and L, H and H/L, L and H/L, Hb and MCHC, MCV and MCH). For four variables (RBC, Ht, Hb and MCHC), sex was a statistically significant factor and for all of these variables females had significantly higher mean values than males. For these four parameters, sex explained 1–5 % of the variation, and for the six parameters for which sex was not significant, it explained 1 % or less of the observed variation. Hatch date was a statistically significant factor for the same four haematological parameters for which sex was significant, and the interaction between hatch date and sex was significant for three of these. Since hatch date was confounded with chick age a positive effect of hatch date could represent a negative effect of age, and vice versa. RBC, Ht, Hb and MCHC all had significant negative effects of hatch date, and, for the first three of these, the negative effect was more pronounced for female chicks than male chicks (Fig. 1). Hatch date explained 3–19 % of variation in these four parameters. A significant effect of brood size was not detected for any of the haematological parameters investigated. Year was modelled as a factor, and was a significant explanatory variable for eight of the haematological parameters: H/L, RBC, WBC, Ht, Hb, MCV, MCH and MCHC. For each of these, year explained the most variation of all of the explanatory variables.

Relationship between red blood cell count (RBC 106/μl; top) and haematocrit [Ht (%); lower)] and White Stork Ciconia ciconia hatch date (females, circles and solid lines, males, crosses and dotted lines). Fitted lines are modelled means, averaged over each year
Discussion
In our study, RBC, Ht, Hb and MCHC decreased significantly with hatch date (i.e. increased with chick age) and were lower in males than females. For the first three of these, a significant sex/hatch date interaction suggests that higher values for females are exaggerated in older chicks. These are novel results, as this is to date the only report of sex differences in the blood of young chicks. These results may be partly explained by the well-known adaptation of birds developing haematological functions (such as oxygen transport) during their developmental stage to prevent age-related tissue damage caused by changes in their blood (Holmes et al. 2001). It can also be hypothesised that the degree to which birds can change oxygen transport in blood, and therefore change haematological parameters, is connected with a response to potential stress, as described by Blas et al. (2006) for the White Stork. However, age and sex effects were small compared to the effects of year and nesting locality. These presumably reflect, respectively, the direct or indirect effects of that year’s weather (e.g. on physiological condition or prey abundance), and on (unrecorded) differences in the local environment. It has previously been shown that year and environment affect chick production and/or population changes in White Stork (e.g. Tryjanowski et al. 2005, 2006), but effects on haematological parameters of chicks have not been previously reported.
The mean haematological values presented here are very similar to those presented by Puetra et al. (1989) and Montesinos et al. (1997) from their studies of White Stork chicks in the wild in Spain, and are also similar to values obtained from chicks of the closely related Black Stork C. nigra (Lanzarot et al. 2005). We also found that Ht, Hb, RBC and MCHC parameters changed with age of chicks, which is a well-known pattern in birds, reflecting increased efficiency of oxygen transport in blood with age (Gayathri et al. 2004; Eklom and Lill 2006; Simmons and Lill 2006), including White Stork chicks (Montesinos et al. 1997). The latter study, which detected no differences between captive and free-living birds, found that RBC, PCV and Hb increased with age, whilst WBC and lymphocytes decreased with age. These changes may reflect sibling competition between chicks, especially at a very young age, but after day 40 of their life nestling mortality is very low, and almost all chicks survive to fledging (Tortosa and Castro 2003; Andrzejewska et al. 2004). In our study, we also detected increases in RBC and Hb with age, but the change in WBC was not significant.
Interestingly, sexual differences were found in four parameters; however, the differences were relatively small, not only in absolute values but also relative to other sources of variation. Although the White Stork is a monomorphic species, and morphological differences between chicks of different sex do not exist, we suspect that changes in haematological parameters are probably due to changes in activity of sex steroid hormones in chicks during development (suggestion in Gil et al. 2007). On the other hand, Dawson and Bortolotti (1997a), in a study on American Kestrels Falco sparverius, found no sexual differences in either Ht or plasma protein levels, although nestling kestrels show sexual differences in plumage and size. Therefore, these differences may be related to the age of chicks and the hierarchy of chicks in particular broods. In the current study, for reasons that are not understood, there was a gender bias among chicks to males, and older chicks also tended to be male (Tryjanowski et al. 2011). As is the case for most species, the youngest White Stork chicks (i.e. <20 days of age) are more sensitive to environmental perturbations or extremes (Tortosa and Castro 2003; Jovani and Tella 2004), whereas as they age they become more capable of thermoregulation (Tortosa and Castro 2003). Given that such changes are likely mediated in part by changes to the circulatory systems, it is reasonable that changes in haematological parameters also change as nestlings become older.
The White Stork is an icon of nature protection, and injured and orphaned young birds are frequently taken into captivity to ensure their survival or to provide offspring for future release programmes (Tryjanowski et al. 2006). We have shown that some haematological parameters vary by age and sex (but more so by year and locality) and that all these factors need to be considered in any diagnostic role in studies of this and possibly other species. Additionally, the values presented here may be useful for comparisons with blood parameters from chicks in polluted areas (Pastor et al. 2001a; Baos et al. 2006a, b). Knowledge of reference values of their blood characteristics will be useful in ensuring their general health and also in post-mortem analyses of birds that are found dead in the wild.
References
Alonso JC, Huecas V, Alonso JA, Abelenda M, Muñoz-Pulido R, Puerta ML (1991) Hematology and blood chemistry of adult white storks (Ciconia ciconia). Comp Biochem Physiol A 98:395–397
Andrzejewska I, Tryjanowski P, Zduniak P, Dolata PT, Ptaszyk J, Ćwiertnia P (2004) Toxoplasma gondii antibodies in the white stork Ciconia ciconia. Berl Münch Tierärztl Wschr 117:274–275
Baos R, Blas J, Bortolotti GR, Marchant TA, Hiraldo F (2006a) Adrenocortical response to stress and thyroid hormone status in free-living nestling white storks (Ciconia ciconia) exposed to heavy metal and arsenic contamination. Environ Health Persp 114:1497–1501
Baos R, Jovani R, Pastor N, Tella JL, Jiménez B, Gómez G, González MJ, Hiraldo F (2006b) Evaluation of genotoxic effects of heavy metals and arsenic in wild nestling white storks (Ciconia ciconia) and black kites (Milvus migrans) from southwestern Spain after a mining accident. Environ Toxicol Chem 25:2794–2803
Bates D, Maechler M (2010) lme4: linear mixed-effects models using S4 classes. R package version 0.99375-35. http://CRAN.R-project.org/package==lme4
Blas J, Baos R, Bortolotti GR, Marchant TA, Hiraldo F (2006) Related variation in the adrenocortical response to stress in nestling white storks (Ciconia ciconia) supports the developmental hypothesis. Gen Comp Endocrinol 148:172–180
Boughton RK, Atwell JW, Schoech SJ (2006) An introduced generalist parasite, the sticktight flea (Echidnophaga gallinacea), and its pathology in the threatened Florida scrub-jay (Aphelocoma coerulescens). J Parasitol 92:941–948
Bowerman WW, Stickle JE, Sikarskie JG, Giesy JP (2000) Hematology and serum chemistries of nestling bald eagles (Haliaeetus leucocephalus) in the lower peninsula of MI, USA. Chemosphere 41:1575–1579
Campbell TW (1995) Avian hematology and cytology, 2nd edn. Iowa State University Press, Ames
Campo JL, Davila SG (2002) Estimation of heritability for heterophil: lymphocyte ratio in chickens by restricted maximum likelihood. Effects of age, sex, and crossing. Poult Sci 81:1448–1453
Ćwiertnia P, Kwieciński Z, Kwiecińska H, Wysocki A, Tryjanowski P, Ollson O (2006) Sexing of white stork Ciconia ciconia based on biometric measurements. In: Tryjanowski P, Sparks TH, Jerzak L (eds) The white stork in Poland: studies in biology, ecology and conservation. Bogucki WN, Poznań, pp 423–429
Dawson RD, Bortolotti GR (1997a) Variation in hematocrit and total plasma proteins of nestling American kestrels (Falco sparverius) in the wild. Comp Biochem Physiol A 117:383–390
Dawson RD, Bortolotti GR (1997b) Are avian haematocrits indicative of condition? American Kestrels as a model. J Wildl Manag 61:1297–1306
Edwards LJ, Muller KE, Wolfinger RD, Qaqish BF, Schabenberger O (2008) An R 2 statistic for fixed effects in the linear mixed model. Stat Med 27:6137–6157
Eklom K, Lill A (2006) Development of parameters influencing blood oxygen-carrying capacity in nestling doves. Emu 106:283–288
Fridolfsson A-K, Ellegren H (1999) A simple and universal method for molecular sexing of non-ratite birds. J Avian Biol 30:116–121
Gayathri KL, Shenoy KB, Hegde SN (2004) Blood profile of pigeons (Columba livia) during growth and breeding. Comp Biochem Physiol A 138:187–192
Gil D, Biard C, Lacroix A, Spottiswoode CN, Saino N, Puerta M, Møller AP (2007) Evolution of yolk androgens in birds: development, coloniality, and sexual dichromatism. Am Nat 169:802–819
Højsgaard S, Wright K, Leidi AA (2010) doBy: groupwise computations of summary statistics, general linear contrasts and other utilities. http://CRAN.R-project.org/package=doBy
Holmes DJ, Flückiger R, Austad SN (2001) Comparative biology of ageing in birds: an update. Exp Gerontol 36:869–883
Jakubas D, Wojczulanis-Jakubas K, Kreft R (2008) Sex differences in body condition and hematological parameters in Little Auk Alle alle during the incubation period. Ornis Fenn 85:90–97
Jenni-Eiermann S, Jenni L (1998) What can plasma metabolites tell us about the metabolism, physiological state and condition of individual birds? An overview. Biol Cons Fauna 102:312–319
Jovani R, Tella JL (2004) Age-related environmental sensitivity and weather mediated nestling mortality in white storks Ciconia ciconia. Ecography 27:611–618
Kania W (1988) Investigations of white stork (Ciconia ciconia) hatching phenology based on bill measurements of nestlings. Ring 134–135:13–19
Kasprzak M, Hetmański T, Kulczykowska E (2006) Changes in hematological parameters in free-living pigeons (Columba livia f. urbana) during the breeding cycle. J Ornithol 147:599–604
Kenward MG, Roger JH (1997) Small sample inference for fixed effects from restricted maximum likelihood. Biometrics 53:983–997
Kostelecka-Myrcha A (1985) Hematological values and respiratory function of a unit of blood volume of birds during their development. Zesz nauk Filii Univ Warsaw 48 Biol 10:11–30
Kulczykowska E, Kasprzak M, Kalamarz H, Kuriata M, Nietrzeba M, Jerzak L, Kamiński P (2007) Melatonin and thyroxine response to pollution in white stork nestlings (Ciconia ciconia): aspects of rhythmicity and age. Comp Biochem Physiol C 146:392–397
Lanzarot MP, Barahona MV, San Andrés MI, Fernández-Garcia M, Rodríguez C (2005) Hematologic, protein electrophoresis, biochemistry, and cholinesterase values of free-living black stork nestlings (Ciconia nigra). J Wildl Dis 41:379–386
Lashev L, Lasheva V, Daskalov Y, Chakarova V (2005) Values of some haematological indices in white storks (Ciconia ciconia), reared in captivity. Bulg J Vet Med 8:265–267
Maxwell MH (1993) Avian blood leukocyte responses to stress. World’s Poult Sci J 49:34–43
Montesinos A, Sainz A, Pablos MV, Mazzucchelli F, Tesouro MA (1997) Hematological and plasma biochemical reference intervals in young white storks. J Wildl Dis 33:405–412
Moreno J, Merino S, Sanz JJ, Arriero E (2002) An indicator of maternal stress is correlated with nestling growth in pied flycatchers Ficedula hypoleuca. Avian Sci 2:175–182
Pastor N, López-Lázaro M, Tella JL, Baos R, Forrero MG, Hiraldo F, Cortés F (2001a) DNA damage in birds after the mining waste spill in southwestern Spain: a comet assay evaluation. J Environ Pathol Toxicol Oncol 20:317–324
Pastor N, López-Lázaro M, Tella JL, Baos R, Hiraldo F, Cortés F (2001b) Assessment of genotoxic damage by the comet assay in white storks (Ciconia ciconia) after the Doñana ecological disaster. Mutagenesis 16:219–223
Puetra ML, Muñoz-Pulido R, Huecas V, Abelenda M (1989) Hematology and blood chemistry of chicks of white and black storks (Ciconia ciconia and Ciconia nigra). Comp Biochem Physiol A 94:201–204
R Development Core Team (2010) R: a language and environment for statistical computing. R foundation for statistical computing. Vienna, Austria. http://www.r-project.org
Sanz JJ, Moreno J, Merino S, Tomás G (2004) A trade-off between two resource-demanding functions: post-nuptial moult and immunity during reproduction in male pied flycatchers. J Anim Ecol 73:441–447
Schulz H (1998) Ciconia ciconia White Stork. BWP Update 2:69–105
Simmons P, Lill A (2006) Development of parameters influencing blood oxygen carrying capacity in the welcome swallow and fairy martin. Comp Biochem Physiol A 143:459–468
Tortosa FS, Castro F (2003) Development of thermoregulatory ability during ontogeny in the white stork Ciconia ciconia. Ardeola 50:39–45
Tryjanowski P, Jerzak L, Radkiewicz J (2005) Effect of water level and livestock on the productivity and numbers of breeding white storks. Waterbirds 28:378–382
Tryjanowski P, Sparks TH, Jerzak L (eds) (2006) The white stork in Poland: studies in biology, ecology and conservation. Bogucki WN, Poznań
Tryjanowski P, Sparks TH, Bochenski M, Dabert M, Kasprzak M, Kaminski P, Mroczkowski S, Wisniewska E, Jerzak L (2011) Do males hatch first and dominate sex ratios in White Stork Ciconia ciconia chicks? J Ornithol 152:213–218
van Kampen E, Zijlstra WG (1961) Standardization of hemoglobinometry. II The hemiglobincyanide method. Clin Chim Acta 6:538–544
Villegas A, Sánchez Guzmán JM, Corbacho C, Corbacho P, Vargas JM (2004) Blood values of bald Ibis (Geronticus eremita) in captivity: comparative ranges and variability with age, sex and physical condition. J Ornithol 145:98–104
Acknowledgments
We thank all farmers and students, especially A. Steliga and A. Ponińska, who participated in this study. Special thanks to B. Kavanagh, J. Blas, R. Nager and S. J. Schoech for comments. The study was undertaken following the Guidelines of the European Union Council and the current laws in Poland, according to the Ethical Commission (permission number: 05/2005).
Conflict of interest
The authors declare that they have no conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by N. Chernetsov.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution License which permits any use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
About this article
Cite this article
Kaminski, P., Jerzak, L., Sparks, T.H. et al. Sex and other sources of variation in the haematological parameters of White Stork Ciconia ciconia chicks. J Ornithol 155, 307–314 (2014). https://doi.org/10.1007/s10336-013-1016-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10336-013-1016-6