
Abstract
Intratympanic (IT) gentamicin injections are effective in the control of episodic vertigo due to Ménière’s disease. Histological studies in animals have found that the loss of type I vestibular hair cells far exceeds that of type II cells after IT gentamicin treatment. The objective of this study was to determine whether this selective toxicity for type I hair cells might be due to selective concentration of the drug by these cells. Gentamicin was localized within the vestibular epithelium by both direct and indirect methods. Gentamicin conjugated to Texas Red® was used as a direct tracer, and anti-gentamicin antibody provided an indirect means of localization. Conjugated or unconjugated gentamicin was injected into the left tympanic space of chinchillas. The animals were killed and fixed 1 or 3 weeks post-treatment. Confocal fluorescence microscopy was used to determine the localization of gentamicin in semicircular canal cristae. Results from the animals killed within 1 week of administration showed that numerous type I hair cells still remained throughout the epithelium. The mean intensity in grayscale units (0–255) of anti-gentamicin labeling for type I hair cells was 28.14 (95% CI 24.60–31.69), for type II hair cells was 17.09 (14.99–19.20), and for support cells was 5.35 (5.34–5.46; p < 0.001, ANOVA). Anti-gentamicin antibody labeling appeared in the majority of type I hair cells throughout their cytoplasm, but with greater intensity at the apex (p < 0.001). Intensity of fluorescence with Texas-Red conjugated gentamicin was 25.38 (22.83–27.94) in type I hair cells, 15.60 (14.73–16.48) in type II cells, and 12.62 (12.06–13.17) in support cells (p < 0.001, ANOVA). These results suggest that type I hair cells are more susceptible to gentamicin because they more avidly take up or retain the drug in the early period after administration.
Similar content being viewed by others

INTRODUCTION
Aminoglycoside antibiotics are toxic to the cochlea and vestibular organs, affecting both hearing and balance (Hinshow and Feldmann 1945; Govaerts et al. 1990). Gentamicin has relatively greater toxicity for vestibular than cochlear hair cells (Webster et al. 1970). This vestibulotoxic effect has been used to advantage to control vertigo in unilateral Ménière’s disease refractory to medical management. Single intratympanic (IT) injections of gentamicin are often effective in reducing vestibular function and controlling vertigo (Chia et al. 2004; Cohen-Kerem et al. 2004).
The precise mechanism by which aminoglycosides exert their toxic effects on hair cells is yet unknown. Hair cells take up and retain aminoglycosides for extended periods (Hayashida et al. 1985; Dulon et al. 1993; Hashino et al. 1997). Intracellular levels of Ca++ and reactive oxygen species, which may be catalyzed by gentamicin–iron complexes, rise after aminoglycoside uptake (Schacht 1993; Clerici et al. 1996; Song et al. 1998; Hirose et al. 1999). Having taken up aminoglycosides, hair cells degenerate within the epithelium through apoptosis (Nakagawa et al. 1997; Forge and Li 2000; Matsui et al. 2002) and can be extruded into the endolymphatic space (Li et al. 1995).
Human physiological studies suggest that the lesion caused by IT gentamicin is partial in nature. The angular vestibulo-ocular reflex (AVOR) in response to rapid rotatory head thrusts is reduced in patients after IT gentamicin treatment, but the reduction in canal function is less severe than that seen after surgical labyrinthectomy (Carey et al. 2002). Recordings from vestibular nerve afferents in chinchillas after IT gentamicin treatment demonstrate that afferents continue to fire spontaneously and respond to galvanic stimulation, but not to head acceleration (Hirvonen et al. 2005).
Lindeman (1969) demonstrated differential sensitivity to aminoglycosides between type I and type II vestibular hair cells. These cells are distinguished by their morphologies and afferent innervations. Type I hair cells, which are more sensitive to aminoglycoside damage, have heterogeneous chromatin and constricted necks, are flask-shaped, and are surrounded by a single calyx afferent ending. Type II cells have homogeneous chromatin, are cylindrical without constricted necks, and are innervated with bouton afferent endings (Lysakowski and Goldberg 1997). Lopez et al. (1997) also found that type I hair cells are more sensitive to the effects of gentamicin than type II hair cells when the drug was administered directly to perilymph. Our laboratory found that a single dose of IT gentamicin in the chinchilla caused a 57% reduction in overall hair cell density, with a 99% reduction in type I hair cells but no significant reduction in type II cells (Hirvonen et al. 2005). We hypothesize that this selective toxicity for type I hair cells reflects a selective uptake or retention of the drug by type I hair cells relative to other cells in the neuroepithelium.
Hayashida et al. (1985) used anti-gentamicin antibodies and immunofluorescent secondary antibodies to show selective staining of guinea pig type I vestibular hair cells after daily intraperitoneal gentamicin administration. Imamura and Adams (2003) found that the degree of gentamicin-induced damage in the cochlea and labyrinth depended on the overall degree of drug uptake by the endorgan as demonstrated by post-treatment labeling with anti-gentamicin antibody. Steyger et al. (2003) also localized the drug by using gentamicin conjugated to the fluorescent probe Texas Red® (GTTR) as an intracellular tracer in hair cells of the bullfrog sacculus in vitro. Dai et al. (2006) demonstrated uptake of GTTR by cochlear hair cells after subcutaneous injection in chicks, guinea pigs, and mice.
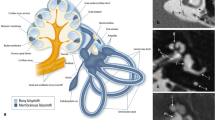
We use similar fluorescent labeling techniques in the present study to localize gentamicin in the chinchilla vestibular neuroepithelium after IT administration. We aimed to evaluate whether the type I hair cells demonstrate more intense labeling with fluorescent markers for gentamicin when compared to other cells in the vestibular neuroepithelium after IT administration of a single injection in a manner comparable to that used in the treatment of Ménière’s disease. Gentamicin was indirectly localized using an anti-gentamicin antibody and directly localized using gentamicin conjugated to Texas Red® (GTTR). Anti-Tenascin antibody was additionally used as a specific marker for afferent calyces to reliably identify type I hair cells. Tenascin is a glycoprotein found in the space extracellular between type I hair cells and calyx afferents (Swartz and Santi 1999).
MATERIALS AND METHODS
Animals
Semicircular canal cristae from seven adult chinchillas (Chinchilla laniger) weighing 350–550 g were studied for anti-gentamicin antibody immunofluorescence (anti-GT group) at one of two different survival periods after IT gentamicin injection: 7 days (four animals) or 21 days (three animals). An additional three animals received IT injections of gentamicin conjugated to Texas Red® (GTTR group); their cristae were harvested after a 5- to 7-day survival period. The survival period of 1 week (5–7 days) was chosen to study the acute period of uptake after drug administration; this is a point where obvious signs of vestibular hypofunction (head tilt and body sway) are not yet present. The 3-week time point was chosen due to our laboratory’s prior experience that the maximum head tilt, a sign of vestibular hypofunction, develops 2.5 to 3 weeks after IT gentamicin administration. In all treatment groups, the animal’s left ear was treated, and the right ear served as the control. Control ears for anti-GT localization were left untreated, as we have previously shown that the buffer carrier solution for gentamicin does not affect vestibular afferent function or vestibular endorgan histology (Hirvonen et al. 2005). For the GTTR control endorgans, buffered Texas Red® (Molecular Probes, Eugene, OR) solution without gentamicin was administered by intratympanic injection into the right middle ear space using the same protocol as for the treated ear.
Ten total treated semicircular canal cristae [three anterior canals (AC), four horizontal canals (HC) and three posterior canals (PC)] were suitable for quantitative analysis in the anti-GT group after 1 week, and seven cristae (2 AC, 3 HC and 2 PC) were suitable after 3 weeks. A total of ten untreated cristae were suitable for quantitative controls for these groups (four AC, three HC and three PC). The GTTR group yielded seven treated cristae (three AC, one HC and three PC) and three control cristae (two AC and one PC). All experimental protocols were performed in accordance with the National Institutes of Health guide for the care and use of laboratory animals (NIH Publications No. 80-23, revised 1978) and were approved by the Animal Care and Use Committee of the Johns Hopkins University School of Medicine (Baltimore, MD).
Treatment and surgical procedures
The animals were anesthetized using intramuscular ketamine 40 mg/kg and xylazine 0.5 mg/kg (Phoenix Pharmaceuticals, St. Joseph, MO). The external auditory canal was sterilized with povidone–iodine solution. For anti-GT groups, gentamicin at a concentration of 26.7 mg/ml in pH 7-buffered solution (American Pharmaceutical Partners, Schaumberg, IL) was injected into the tympanic cavity after puncturing the tympanic membrane using a 25-gauge spinal needle. The treatment solution was injected until the middle ear space was visibly full (0.2–0.6 ml). The animal’s head remained stationary with the treated ear and nose turned towards the ceiling so as to bathe the round window niche in solution for 30 min. For the GTTR group, the gentamicin conjugated to Texas Red® was administered in the same fashion. Dimethyl formamide was used to suspend succinimidyl esters of Texas Red® (Molecular Probes; 5 mg/ml) to make a 2.0 mg/ml Texas Red® (TR) stock solution. For conjugation, 2.2 ml of gentamicin sulfate solution (Sigma, St. Louis, MO; 50 mg/ml in 0.1 M K2CO3, pH 9) was added to 0.3 ml of TR stock solution (300:1 molar ratio) and agitated overnight at 4C (Steyger et al. 2003). The GTTR conjugate solution (44 mg/ml) was then diluted to a concentration of 26.7 mg/ml using sterile phosphate buffer.
Tissue harvesting
After each respective survival period, deep anesthesia was induced with intraperitoneal 5,5 diallylbarbituric acid 40 mg/kg (Sigma), and the animals were killed using a transcardiac perfusion of 0.9% saline followed by fixative (4% paraformaldehyde, 1% acrolein, 1% picric acid, 5% sucrose in 0.1M PO4 buffer). After decapitating and opening both bullae, the skull was immersed in the fixative for up to 16 h. The otic capsule was drilled away from the vestibular endorgans, which were then harvested, washed in phosphate buffer, dehydrated in graded ethanol solutions and cryoprotected using graded sucrose solutions.
Immunohistochemistry
Whole cristae were blocked for nonspecific immunoreactivity (0.5% fish gelatin and 1% bovine serum albumin in 0.4% Triton). Treated and control whole endorgans from the anti-GT groups were incubated with anti-gentamicin monoclonal antibodies (QED Biosciences, San Diego, CA), 1:200 in blocking solution for 48 h, and secondarily labeled with Alexa Fluor® 555-conjugated goat antimouse antibodies (Molecular Probes) for 24 h. Subsequently, all specimens were labeled with polyclonal rabbit antichicken tenascin antibodies, using a 1:200 dilution of antibody reactive for tenascin C with some crossreactivity to tenascin R (Chemicon International, Temecula, CA). Anti-tenascin antibody was applied for 48 h, and then specimens were secondarily labeled by Alexa Fluor® 488-conjugated goat antirabbit antibodies (Molecular Probes) for 24 h. Tissue was embedded in Tissue Tek OCT compound (Miles, Elkart, IN) by cooling with 2-methylbutane brought to approximately −60°C with dry ice, and transversely cut into 10-μm thick sections using a cryostat (four to seven sections per endorgan). Sections were mounted in Vectashield mounting medium with 4′,6-Diamidino-2-phenylindole (DAPI) to label nuclei (Vector Labs, Burlingame, CA) and sealed with coverslips.
Confocal microscopy
Specimen sections from the midportion of the cristae (four to seven sections per endorgan) were scanned using a Zeiss LSM 510 Meta confocal microscope (Carl Zeiss, Thornwood, NY). Serial optical image slices (0.5–1.0 μm) through the 10-μm physical sections were obtained. For all the specimens, laser power settings and pinhole were not varied. Amplifier offset and detector gain were optimized for each endorgan to prevent saturation of pixels in the images. Images were recorded with Plan-Apochromat 40×/1.00 NA and 63×/1.40 NA oil lenses at a minimum 0.14 × 0.14 μm/pixel resolution. To avoid cross talk between the different probes, specimens were sequentially excited with the respective lasers and recorded. Minimal adjustments to photodetector gain were necessary between different specimens to prevent saturation of pixels in any specimen, but gain was held constant for all optical image slices in a given tissue section.
For fluorescence intensity measurements, one image slice from each z-stack was randomly selected (Urbaniak and Plous 2006). Intensity of the respective fluorescent probe within the cells of the neuroepithelium for the GTTR and 1 week anti-GT groups was quantified using the image analysis software of the Zeiss LSM 510 Meta confocal microscope (Carl Zeiss, Thornwood, NY). The software reported the intensity as a grayscale value (0–255). Type I hair cells were identified by the basal location of their nuclei within the sensory cell layer, their flask shape with constricted necks, and anti-tenascin labeling of their associated calyx afferent endings. Type II hair cells were identified by the more apical location of their nuclei within the sensory cell layer and lack of associated anti-tenascin labeling. Support cells were identified by their dense heterochromatin and nuclear location adjacent to the basal lamina (Lysakowski and Goldberg 1997). The operator outlined the borders of the cell so that the intensity was measured on the area including the cytoplasm and nucleus. This was facilitated by scrolling through the surrounding optical slices of the z-stack and selectively activating the different color laser channels.
Additionally, for type I hair cells of the 1-week anti-GT localization group, apex intensity of the probe was measured and compared to the intensity of the probe at the base. The apex was defined as the area from the top of the cell to the nearest constriction of the neck of that cell. The base was defined as the area from the point of the apical neck constriction to the furthest basal extent of the cytoplasm of the cell. The nucleus was excluded to ensure that a difference in intensity of the two regions was not biased by decreased concentration of fluorescent tracers by the nucleus. By 3 weeks the disruption of the boundaries of remaining cells did not allow for reliable outlining of the cytoplasm to compare intensity of labeling between different cellular sites.
Cell counts
Sensory and support cells were quantified in the three groups: controls with no gentamicin treatment, 1-week post IT gentamicin treatment, and 3-weeks post treatment. Cell counts from the randomly selected optical slice were normalized by dividing by the length of the neuroepithelium in that slice to generate a linear density measurement. Because of the disruption of the epithelial architecture appearing 3 weeks after IT gentamicin treatment, cells were simply categorized as being in the “support cell layer” (cells with denser chromatin and contiguous with the basal lamina) or in the “sensory cell layer” (located more apically in the neuroepithelium). Cells were counted if the section of the nucleus seen was at least 50% of the 6–7 μm-diameter of the typical nucleus. To avoid double counting of nuclei, we ensured that the distance between the selected optical slices in adjacent tissue sections was at least 6 μm. Counts were obtained in the same way between control and treated sides. To control for a nuclear shrinkage effect on linear density, the degree and possible effects of nuclear shrinkage were obtained by measuring nuclear diameters in randomly selected cells of the three cell types from control and treated tissues. If shrinkage was found to be a factor, appropriate adjustments to the changes in linear cell density were made to account for shrinkage effect.
Comparisons and statistics
The measured outcome variables were the intensity of the fluorescent labels for gentamicin and the densities of hair cells and support cells. Means and 95% confidence intervals were calculated for these measures. General linear model (GLM) analyses of variance were performed on these outcomes to determine the significance of the input variables of treatment, animal, canal, and cell type. Variables thus identified as significant were further analyzed through the application of post hoc ANOVA and t tests. To control for differences in the efficacy of anti-GT antibody labeling between different endorgans and different sections, we also calculated the mean ratio of type I to type II hair cell intensity for each analyzed optical slice.
RESULTS
Distribution of gentamicin within the vestibular neuroepithelium
Anti-gentamicin antibody labeling
Anti-GT antibodies tagged with the red Alexa Fluor® 555 fluorochrome intensely labeled hair cells in the treated crista 1 week post drug administration, whereas the control specimen displayed negligible red fluorescence (Fig. 1). It is also evident that at this time, there is some loss of type I hair cells from the central zone (apex and adjacent areas) of the treated crista, and this area is thinner when compared with that of the control specimen. Intense pan-cellular (cytoplasmic and nuclear) anti-GT labeling was seen only in treated type I hair cells, identifiable by their flask shapes and by the thin rim of anti-tenascin labeling (green) associated with their calyces. Type II hair cells, without associated tenascin-labeled afferent endings and with more apically located nuclei, showed less anti-gentamicin antibody labeling. Support cells showed negligible intensity.

Cross-sections of the crista ampullaris from control, 1-week, and 3-week treated chinchillas labeled with anti-gentamicin (anti-GT) and anti-tenascin antibodies. Each lower panel magnifies the boxed area in the respective upper panel. Red represents anti-GT antibody, blue represents DAPI labeling of ds-DNA in the nuclei of cells and green represents anti-tenascin labeling between calyces and type I hair cells. CONTROL: Image of an untreated right anterior canal crista demonstrating the utility of anti-tenascin label in the identification of calyces and type I hair cells. Anti-gentamicin antibody labeling is absent in the control crista. ONE WEEK: Image of a left horizontal canal crista 1 week after IT gentamicin treatment. Anti-GT labeling is diffuse and intense in type I hair cells (I) but is less intense in type II hair cells (II) and not visible in support cells (S). Thinning of the central region of the neuroepithelium is evident (compare thickness at arrows). THREE WEEKS: Left horizontal canal crista three weeks after IT gentamicin exposure. Type I hair cells do not appear to be present, based on the absence of diffuse and intense anti-gentamicin labeling and calyx-associated anti-tenascin labeling. The entire neuroepithelium is thinned. A clear support cell layer persists (S). Above this layer, occupying the neuroepithelium’s sensory layer (Se), are cells with ill-defined shape and weak, inhomogeneous anti-gentamicin labeling. Anti-tenascin labeling of the basement membrane is variable across specimens.
Quantitative analysis of the anti-GT intensity data demonstrated that type I hair cells concentrated more gentamicin 1 week after treatment than did other cell types (Fig. 2). To determine whether cell type had a significant influence on anti-GT labeling intensity in all 1-week specimens, a GLM analysis was done using the following input factors: intervention (df = 1: treatment vs control), cell type (df = 2: type I vs type II vs support), and canal (df = 2: AC vs HC vs PC). This model predicted 63% of the adjusted sum square error from the mean anti-GT intensity (i.e., r 2 = 0.63). Intervention alone accounted for 26% of the model’s predicted error from the mean (p < 0.0001), and post hoc t tests showed that type I and II hair cells labeled more intensely in treated specimens than in controls (p < 0.001). Cell type and the interaction of intervention × cell type accounted for 71% of the GLM model’s predicted error from the mean (p < 0.0001 for each), leading to a rejection of the null hypothesis that all cell types label to the same degree with gentamicin treatment. Post hoc ANOVA demonstrated that in treated cristae the anti-GT fluorescence of type I hair cells exceeded that of type II hair cells, which in turn exceeded that of support cells (p < 0.001). There were also significant (p < 0.0001) interactions in the GLM model of intervention × canal and intervention × canal × cell, suggesting that canals did not take up gentamicin equally and that some cells in some canals took up gentamicin especially avidly. This was confirmed by ANOVA, which demonstrated that at 1 week after treatment, fluorescence was lower for remaining type I hair cells in the HC when compared to those of the AC and PC (23.6 ± 13.8, 28.7 ± 16.8 and 29.5 ± 15.8, respectively; p = 0.005). For type II hair cells intensities in the HC, AC, and PC were 11.9 ± 3.6, 15.2 ± 4.3, and 14.9 ± 6.2, respectively (p < 0.0001). However, these canal effects accounted for only 2% of the GLM model’s predicted error from the mean.

Intensity of anti-gentamicin fluorescent antibody labeling one week after intratympanic gentamicin treatment. Fluorescence intensity is expressed in grayscale values from 0 to 255. Intensity of the total neuroepithelium in each optical slice was obtained by simultaneously measuring all of the neuroepithelial elements above the basement membrane. Average intensities for each of the indicated cell types are also shown. Error bars are 1 SD. Asterisks indicate p values of <0.001 that were obtained for post hoc t tests comparing control (gray bars) and treated (black) values. Numbers of specimens were: for total neuroepithelia: 10 controls, 7 treated; for type I hair cells: 593 controls and 367 treated; for type II hair cells: 445 controls and 302 treated; and for support cells: 877 controls and 697 treated. Also indicated (arrows) is the significant gradient of label in treated type I hair cells > treated type II hair cells > treated support cells (ANOVA, p < 0.001).
There was also variation between animals in anti-GT intensity 1 week after treatment. This was demonstrated by a GLM analysis of intensity of treated cells using factors of animal, cell type, and their interaction. This model accounted for 52% of the error from the mean (r 2 = 0.52). The largest predictive factor remained cell type (65% of the model’s predicted error from the mean), but the effects of animal (18%) and the animal × cell type interaction (17%) were also significant (p < 0.0001). Thus, as an additional test, to control for the effects of inter-animal variation as well as inter-specimen variation, the ratio of intensity of fluorescence of anti-GT antibody in type I hair cells to that in type II hair cells was determined for each optical slice to control for differences in antibody labeling efficacy. The mean ratio was 2.08 (95% CI 1.91–2.24) in treated specimens, but 1.08 (95% CI 1.00–1.15) in controls.
Among type I hair cells, a zonal gradient of gentamicin fluorescence was observed in treated cristae 1 week after drug exposure. In some specimens central zone type I hair cells were still present, and anti-GT fluorochrome was distributed throughout their cytoplasm and nuclei. In the same specimens, the labeling seen in peripheral zone type I hair cells was punctate, not diffuse (Fig. 3a). In other 1-week specimens damage appeared to have caused of loss of central zone type I hair cells. In these specimens, the peripheral type I hair cells did not show punctate labeling. Rather, the fluorochrome was diffusely distributed (Fig. 3b). In such hair cells, there was an apical predominance of the label, with the apex/base intensity ratio being 1.59 (95% CI 1.17–2.01, N = 26).

The spectrum of type I hair cell damage 1 week after IT gentamicin treatment. A Anti-GT labeled type I hair cells are identifiable throughout the crista, but those in the central zone already appear to be extruding. Note that labeling in peripheral type I cells is mostly punctate, with a low level of cytoplasmic intensity (Identified by arrows). B Type I hair cell damage appears more advanced in this specimen: The epithelium is thinner in the central zone, where fewer type I cells and associated tenascin labeling can be identified. Those remaining in the peripheral zone now show diffuse anti-GT labeling (A is a cross section of a left anterior canal crista and B is the cross section of a left posterior canal crista).
By the survival point of 3 weeks, the neuroepithelium was markedly altered by IT gentamicin treatment. Type I hair cells, which had demonstrated significant concentration of the drug by 1 week, were no longer identifiable in the neuroepithelium (Fig. 1). Calyces, previously identifiable with the tenascin label, could no longer be found. Nuclei of cells were still identifiable in the sensory layer of the neuro-epithelium (Se). Distinct anti-GT fluorescent labeling of hair cells was not seen, but less intense punctate labeling was detected. Boundaries of remaining cells in the sensory layer could not be reliably outlined, thus intra- and intercellular intensity measurements were not obtained at this time point.
Direct localization with gentamicin conjugated to Texas Red®
After intratympanic treatment with GTTR and a survival period of 5–7 days, the red-labeled drug was sequestered primarily in remaining type I hair cells of the neuroepithelium, which were again identified with the aid of anti-tenascin antibody (Fig. 4b). Unconjugated Texas Red® (TR) fluorochrome used in control labyrinths did not remain bound (Fig. 4a). In contrast to the result obtained with the anti-GT antibody, GTTR fluorescence was punctate, not uniform, in treated type I hair cells. However, as with anti-GT antibody labeling, the GTTR fluorescence was localized primarily at the apical region of the type I cells. The loss of type I hair cells appeared to be extensive in some specimens 1 week after intratympanic GTTR treatment (Fig. 4c), giving a much more variable labeling result than that obtained after IT gentamicin treatment and labeling with anti-GT antibody (Fig. 1). Type I hair cells that were identifiable in the GTTR-treated preparations had greater intensity of the drug-fluorochrome conjugate than did type II hair cells or support cells.

Gentamicin conjugated to Texas Red® (GTTR) localized in the neuroepithelium of three anterior semicircular canal cristae. A Control specimen, left anterior canal crista, one week post IT treatment with Texas Red® fluorochrome in buffer. Note the lack of red fluorescence within the crista. Tenascin labeling (green) is associated with type I hair cells. B Crista 1 week post treatment with IT GTTR. Punctate GTTR labeling is most intense in the apices of type I hair cells, as indicated by arrow. C Another 1-week GTTR treated specimen. Note the decreased number of type I hair cells in the sample. The fluorescence is still, however, concentrated in the type I hair cell that is present.
A GLM analysis for the GTTR fluorescence was performed using the following input factors: intervention (df = 1: treatment vs control), cell type (df = 2: type I vs type II vs support) and animal (df = 2; canal could not be treated as a separate factor because of a rank error deficiency). This model predicted 70% of the adjusted sum square (Adj SS) error from the mean GTTR intensity (r 2 = 0.70). Intervention alone accounted for 21% of the model’s predicted error from the mean, and post hoc t tests showed that type I and II hair cells labeled more intensely in treated specimens than in controls. Cell type and the interaction of intervention × cell type accounted for 10% of the GLM model’s predicted error from the mean, a smaller effect than was seen with anti-GT labeling, but still significant (p < 0.0001) effect than was seen with anti-GT labeling. This would lead to rejection of the null hypothesis that all cell types label to the same degree after intratympanic GTTR treatment. A post hoc ANOVA demonstrated that in treated cristae GTTR labeling of type I hair cells (25.38 ± 6.33, n = 26), exceeded that of type II hair cells (15.60 ± 5.19, n = 137), which in turn exceeded that of support cells (12.62 ± 4.10, n = 212; p < 0.001; Fig. 5). The variability across animals seen in Figure 4 was reflected in the large effects of animal and intervention × animal in the GLM model, accounting for 26 and 31% of the model’s predicted error from the mean, respectively (p < 0.0001). To control for such variability, the intensity of fluorescence of type I hair cells was also expressed as a ratio to the intensity of fluorescence of type II hair cells in the same optical section. The mean ratio of type I to type II fluorescence with GTTR was 1.66 (95% CI 1.36–1.95), again demonstrating greater affinity of type I cells for gentamicin labeled with TR. The ratio within control specimens was 1.02 (95% CI 1.00–1.05).

Intensity of gentamicin-Texas Red® conjugate labeling one week after IT gentamicin treatment. Fluorescence intensity is expressed in grayscale values from 0 to 255. Intensity of the total neuroepithelium in each optical slice was obtained by simultaneously measuring all of the neuroepithelial elements above the basement membrane. Average intensities for each of the indicated cell types are also shown. Error bars are 1 SD. Asterisks indicate p values of <0.001 that were obtained for post hoc t tests comparing control (gray bars) and treated (black) values. The diamond indicates a p value of 0.003 (post hoc t test). Numbers of specimens were: for total neuroepithelia—3 controls, 7 treated; for type I hair cells—54 controls and 26 treated; for type II hair cells—57 controls and 137 treated; and for support cells—92 controls and 212 treated. Also indicated (arrows) is the significant gradient of label in treated type I hair cells > treated type II hair cells > treated support cells (ANOVA, p < 0.001).
Changes in density of sensory and support cells
Nuclear size changes associated with gentamicin treatment were measured. In control specimens, mean nuclear diameters were: 6.42 ± 0.71 μm (type I hair cells), 7.04 ± 0.99 μm (type II hair cells), and 6.02 ± 0.72 μm (support cells). A significant decrease in diameter in gentamicin-treated specimens was noted only for nuclei of type I hair cells at 1 week after treatment (6.00 ± 0.68 μm, a 6.5% decrease, p < 0.0001). No other significant changes were found in the residual cells in the 1- and 3-week post-treatment specimens (statistical power of 90% was attained to detect changes in diameter of ≥8%).
There was a significant sensory cell reduction in cristae at both 1 week and 3 weeks after intratympanic gentamicin treatment in comparison with controls. By 1 week, hair cell density was reduced by 16.8% (p < 0.0001). After correcting for the observed nuclear shrinkage, the reduction was estimated to be only 8.8% (p = 0.2). Examination of the histological sections demonstrated that this reduction mostly reflected damage to the central zone of the crista, where the cytoarchitecture was disrupted and the epithelium focally thinned relative to controls (Fig. 1). However, more substantial reduction occurred by 3 weeks after IT drug administration, when sensory cells were reduced by 68.3% (p < 0.0001), or 60.3% when corrected for cell shrinkage due to treatment (p < 0.0001). Type I and II hair cells could not be readily identified at 3 weeks because of the cytoarchi-tectural disruption of the epithelium that left only nuclei identifiable in the sensory layer. There were no cells observed to be intensely labeled throughout the cytoplasm with anti-gentamicin antibody as had been seen 1 week after treatment. There was also no tenascin staining of calyceal endings, whereas tenascin signal remained at the basement membrane. In correlation to the observation at 1 week of negligible fluorescent labeling within support cells with a nonsignificant reduction in linear density, at the 3-week time point these cells were reduced by only 5.33% (p = 0.0026).
A GLM analysis for the output of cell linear density was done using the following input factors: survival period (df = 2: time 0 (controls) vs 1-week treated vs 3-week treated), canal (df = 2: AC vs HC vs PC), and cell type (df = 1: sensory vs support) and their interactions. The model accounted for 82% of the adjusted sum square error from the mean. Survival period accounted for 44% of the model’s predicted error from the mean, survival period × cell type accounted for 34%, cell type for 18%, and survival period × canal for 5% (all p values < 0.0001). Three weeks after gentamicin treatment the sensory cell densities in the canals were reduced by 73% (AC), 69% (HC), and 63% (PC; p = 0.002, ANOVA).
DISCUSSION
It has previously been demonstrated that local application of gentamicin to the labyrinth causes greater loss of type I than type II vestibular hair cells (Lopez et al. 1997; Hirvonen et al. 2005). However, the reason for this selective damage has not been previously elucidated. The present study provides evidence that the selective loss of type I hair cells may occur because these cells preferentially accumulate gentamicin acutely after IT administration. Type II hair cells and support cells concentrate substantially less gentamicin (Fig. 2).
Imamura and Adams (2003) analyzed gentamicin concentration within inner ear tissues after IT administration and noted an accumulation within vestibular hair cells. However, differences between type I and type II hair cells were not evaluated. Hayashida et al. (1985) demonstrated that systemic administration of gentamicin led to greater accumulation of the drug in type I hair cells in the guinea pig. The present study is the first study to examine the differential uptake or retention of gentamicin after IT administration in the dose range used in clinical practice for the treatment of Ménière’s disease. The results therefore have clinical relevance in addition to basic implications for aminoglycoside ototoxicity.
Preferential loss of type I hair cells
We have previously demonstrated that IT gentamicin treatment in the chinchilla severely reduces vestibular afferent sensitivity to head acceleration but only mildly reduces spontaneous afferent firing rates (Hirvonen et al. 2005). That study demonstrated a 57% reduction in sensory cells 3 weeks to 3 months after treatment. However, the ultimate fate of type I hair cells could not be determined at the level of light microscopy (LM) because calyces, which are critical to identify these cells by LM, were virtually eliminated. The present study demonstrates an immunohistochemical correlate to the loss of type I hair cells in the loss of labeling for tenascin, a molecule that localizes to the space between the calyx and the basolateral membrane of the type I hair cell. Another piece of evidence that type I hair cells are lost by 3 weeks after intratympanic gentamicin treatment comes from the data derived with anti-GT antibody labeling. By 3 weeks after treatment, there are no longer any cells that diffusely label with anti-GT antibody. At 1 week after treatment, the only cells that were labeled in this fashion were type I hair cells. We conclude that type I hair cells are either lost or that they clear the drug by 3 weeks. However, given the decrease in the total number of sensory cell nuclei coupled with the loss of tenascin-associated calyx endings, the latter possibility seems less likely. Because type I cells normally comprise 50% of the hair cells in the chinchilla crista (Lysakowski and Goldberg 1997), the 68% loss observed here suggests that some type II cells are lost as well.
A confounding factor in our study is the possible effect of nuclear shrinkage on estimates of cell density. Gentamicin treatment was associated with a decrease in the diameter of type I hair cell nuclei at 1 week. This may reflect the toxic effects of gentamicin on the cells that concentrate the drug the most. Whereas significant decreases were not detected in the diameter of nuclei of other cells, shrinkage of as much as 8% would not have been detected by our methods. Such shrinkage can affect estimates of linear cell density. Accounting for 8% shrinkage still does not explain the highly significant reduction in density of total sensory cells at 3 weeks. However, a small change such as the 16.8% reduction in sensory hair cells at 1 week is rendered potentially insignificant.
Evidence for greater concentration of gentamicin by type I hair cells
Both direct (anti-GT) and indirect (GTTR) localization methods demonstrated a greater intracellular concentration of gentamicin in type I hair cells when compared to type II cells in the treated crista 1 week after treatment. These results suggest that the preferential loss of type I hair cells after IT gentamicin treatment may be explained by a greater affinity that these cells have for the drug.
Secondary sources of variability in gentamicin localization were noted in the GLM analysis. For anti-GT and GTTR results, there was significant variability between animals. This may reflect variability in the penetration of gentamicin through the round window, pharmacokinetic differences, or differences in the temporal progression of toxicity between animals. Differences in the efficacy of antibody binding must always be considered as a limitation for quantification with immunohistochemistry. However, we controlled for this by expressing the ratio of fluorescence of type I to type II hair cells within each optical slice examined and still found a mean ratio significantly different than one.
Possible reasons for greater gentamicin concentration in type I hair cells
We found that apical concentration of gentamicin exceeded basal concentration in type I hair cells. This finding is consistent with entry of the drug either via apical endocytosis or stereociliary transduction channels (Myrdal et al. 2005). The latter mechanism raises two potential reasons for possibly greater uptake of gentamicin in type I hair cells as compared to type II cells. The first is structural. The hair bundle structure in type I hair cells differs from that of type II cells. Type I hair cell bundles are taller and have greater numbers of stereocilia that are larger in diameter (Morita et al. 1997; Moravec and Peterson 2004; Xue and Peterson 2006). Because each stereocilium is associated with one to two mechanotransduction channels, the greater number of stereocilia on type I hair cells can produce larger transduction current amplitudes (Denk et al. 1995). The second reason is physiological. There may be a greater electrochemical gradient driving gentamicin entry into type I hair cells. Chen and Eatock (2000) confirmed in isolated hair cells of the rat and other amniotes that the type I hair cells of the crista have more negative resting potentials than type II hair cells. When the transduction channel opens, there is therefore a greater electrochemical gradient driving gentamicin entry into the type I hair cell (Marcotti et al. 2005). Consistent with the possibility of greater entry of gentamicin through type I stereocilia, Walsh et al. (2000) demonstrated that type I mature hair bundles are more susceptible to structural damage after gentamicin exposure than type II mature hair bundles.
Gradual toxicity of gentamicin
Lopez et al. (1997) demonstrated a progressive loss of type I hair cells, first from the central zone and later from the peripheral zone, over 1 week after perilymphatic gentamicin treatment. Our findings are consistent with a similar temporal progression, albeit slower, probably because of the indirect (round window) application of the drug. Many type I hair cells persist 1 week after IT gentamicin but are lost at 3 weeks. In some specimens, there is diffuse and intense labeling of entire type I cells in the central zone but not in the peripheral zone (Fig. 3a). In other specimens, type I hair cells appear to be lost from the central zone, causing it to appear thinner, and labeling of those in the peripheral zone is diffuse and intense (Fig. 3b). Although a time series of multiple endpoints was not used in this study, the findings can be interpreted in light of the results of Lopez et al. (1997) to suggest that central zone type I hair cells first concentrate the drug, followed by those in the peripheral zone. Cell death may follow the same temporal progression, but that appears to be complete by 3 weeks.
It has been suggested that gradual hair cell toxicity may be due to the delayed release of aminoglycosides from intracellular stores where they are initially sequestered (Hashino et al. 1997). The possible progression in the anti-GT labeling of peripheral type I hair cells from punctate to diffuse may be germane to this hypothesis. Punctate labeling that appears to be an early pattern of gentamicin distribution may reflect sequestration in lysosomes, endoplasmic reticulum, mitochondria, or the Golgi apparatus (Sandoval et al. 2000; Steyger et al. 2003). The pan-cytoplasmic labeling that occurs later may reflect the rupture of these organelles. Alternatively, diffuse labeling of the cytoplasm may simply result from gentamicin entering through the stereocilia.
Pan-cytoplasmic labeling was not observed in our GTTR-treated specimens. Such diffuse labeling is only seen after delipidation, which is thought to unveil GTTR fluorescence otherwise quenched by intracellular phopholipids (Myrdal et al. 2005; Dai et al. 2006). While our specimens were treated with Triton, fixation with acrolein may have crosslinked molecules and prevented effective delipidation. However, this methodological limitation would not apply to the anti-GT technique, in which both diffuse and punctate labeling can be seen in cells within the same specimen (Fig. 3a).
Differences between canals
Hair cells with type I morphology were eliminated from all of the canals by 3 weeks after IT gentamicin treatment. Loss of remaining cells in the sensory layer—presumably type II hair cells—at this time was greater in the AC and HC than in the PC. However, 1 week after treatment, the intensity of anti-GT antibody label in type II cells was greater in the PC and AC than in the HC. This unexpected finding raises the possibility that there are biological differences in the ototoxic potential of gentamicin for type II hair cells independent of the degree of concentration of the drug.
Physiological implications
Irregularly discharging units in the normal crista invariably have calyx endings. High-gain units also have bouton endings (dimorphic units), and a group of low-gain irregular afferents in the crista have only calyx endings (Baird et al. 1988). We have previously shown that irregular afferent units persist after IT gentamicin treatment (Hirvonen et al. 2005). This study suggests that IT gentamicin treatment eliminates type I hair cells and therefore their inputs to calyx endings. Why do these irregular units persist? One explanation has been offered by Smith and Goldberg (1986), who hypothesized that discharge regularity is a feature determined by the membrane properties of the afferent neuron, not its complement of hair cell input. Another explanation may be that calyx endings persist in a retracted form, corresponding to the large globular structures observed in the neural layer after IT gentamicin treatment (Hirvonen et al. 2005). These afferent endings could obtain synaptic inputs from outer face synapses on remaining type II cells. Electron microscopy and afferent immunohistochemical labeling work is ongoing in our laboratory to help elucidate whether such globular endings correspond to former calyceal endings and whether they receive synaptic input.
Relevance to human IT gentamicin treatment
Can we extrapolate that the lesion seen in the chinchilla applies to the human labyrinth treated with intratympanic gentamicin? Ishiyama et al. (2007) recently reported histological data from after labyrinthectomy in a patient with Ménière’s disease who continued to have vertigo attacks after six IT gentamicin injections. Histology indicated near complete ablation of the vestibular neuroepithelium of the crista ampullares and a severe loss of both type I and type II hair cells of the utricular macula. This degree of loss likely reflects greater damage than would occur after a single IT gentamicin injection. Human physiological testing suggests that the lesion created by low-dose IT gentamicin treatment is a partial one. The gain of the angular vestibuloocular (AVOR) reflex in humans after IT gentamicin treatment is reduced, but not as severely as after labyrinthectomy or surgery to sever the vestibular nerve (Hirvonen et al. 2005). However, IT gentamicin does abolish the ability to augment the AVOR gain for viewing near targets (Migliaccio et al. 2004). This result matches that obtained in the rhesus monkey when galvanic currents were applied to the vestibular nerve so as to silence irregularly discharging afferents (Chen-Huang and McCrea 1998). Irregularly discharging units receive hair cell input from type I hair cells alone (calyx units) or from both type I and II hair cells (dimorphic units); very regularly discharging afferents receive only input from boutons on type II hair cells (Goldberg 2000). Thus, silencing the pool of afferents that carry type I hair cell input in monkeys has the same effect as IT gentamicin does in humans. It is therefore tempting to speculate that the human labyrinth treated with low-dose IT gentamicin has a selective lesion of type I hair cells. The present study lends some strength to this hypothesis, as type I cells preferentially accumulate gentamicin and are then eliminated from the crista in this animal model. However, in this model, both regular and irregular afferents show marked reductions in gain for head rotations. Although type II hair cells appear to be preserved, they may lose their stereociliary bundles and the ability to transduce head acceleration (Hirvonen et al. 2005). The gentamicin lesion in the chinchilla may be more severe than that in humans due to the thinner round window membrane in the chinchilla (Sahni et al. 1987). Further studies delivering less of the drug to the chinchilla labyrinth might demonstrate a lesion that is more physiologically restricted to irregular units.
References
Baird RA, Desmadryl G, Fernandez C, Goldberg JM. The vestibular nerve of the chinchilla. II. Relation between afferent response properties and peripheral innervation patterns in the semicircular canals. J. Neurophysiol. 60:182–203, 1988.
Carey JP, Hirvonen T, Peng GC, Della Santina CC, Cremer PD, Haslwanter T, Minor LB. Changes in the angular vestibulo-ocular reflex after a single dose of intratympanic gentamicin for Meniere’s disease. Ann. N. Y. Acad. Sci. 956:581–584, 2002.
Chen JW, Eatock RA. Major potassium conductance in type I hair cells from rat semicircular canals: characterization and modulation by nitric oxide. J Neurophysiol 84:139–151, 2000.
Chen-Huang C, McCrea RA. Contribution of vestibular nerve irregular afferents to viewing distance-related changes in the vestibulo-ocular reflex. Exp Brain Res 119:116–130, 1998.
Chia SH, Gamst AC, Anderson JP, Harris JP. Intratympanic gentamicin therapy for Meniere’s disease: a meta-analysis. Otol. Neurotol. 25:544–552, 2004.
Clerici WJ, Hensley K, DiMartino DL, Butterfield DA. Direct detection of ototoxicant-induced reactive oxygen species generation in cochlear explants. Hear. Res. 98:116–124, 1996.
Cohen-Kerem R, Kisilevsky V, Einarson TR, Kozer E, Koren G, Rutka JA. Intratympanic gentamicin for Meniere’s disease: a meta-analysis. Laryngoscope 114:2085–2091, 2004.
Dai CF, Mangiardi D, Cotanche DA, Steyger PS. Uptake of fluorescent gentamicin by vertebrate sensory cells in vivo. Hear. Res. 213:64–78, 2006.
Denk W, Holt JR, Shepherd GM, Corey DP. Calcium imaging of single stereocilia in hair cells: localization of transduction channels at both ends of tip links. Neuron. 15:1311–1321, 1995.
Dulon D, Hiel H, Aurousseau C, Erre JP, Aran JM. Pharmacokinetics of gentamicin in the sensory hair cells of the organ of Corti: rapid uptake and long term persistence. C. R. Acad. Sci. III 316:682–687, 1993.
Forge A, Li L. Apoptotic death of hair cells in mammalian vestibular sensory epithelia. Hear. Res. 139:97–115, 2000.
Goldberg JM. Afferent diversity and the organization of central vestibular pathways. Exp. Brain Res. 130:277–297, 2000.
Govaerts PJ, Claes J, van de Heyning PH, Jorens PhG, Marquet J, De Broe ME. Aminoglycoside-induced ototoxicity. Toxicol. Lett. 52:227–251, 1990.
Hashino E, Shero M, Salvi RJ. Lysosomal targeting and accumulation of aminoglycoside antibiotics in sensory hair cells. Brain Res. 777:75–85, 1997.
Hayashida T, Nomura Y, Iwamori M, Nagai Y, Kurata T. Distribution of gentamicin by immunofluorescence in the guinea pig inner ear. Arch. Otorhinolaryngol. 242:257–264, 1985.
Hinshow HC, Feldmann WH. Streptomycin in the treatment of clinical tuberculosis: a preliminary report. Proc Mayo Clin 20:313–318, 1945.
Hirose K, Westrum LE, Stone JS, Zirpel L, Rubel EW. Dynamic studies of ototoxicity in mature avian auditory epithelium. Ann. N. Y. Acad. Sci. 884:389–409, 1999.
Hirvonen TP, Minor LB, Hullar TE, Carey JP. Effects of intratympanic gentamicin on vestibular afferents and hair cells in the chinchilla. J. Neurophysiol. 93:643–655, 2005.
Imamura S, Adams JC. Distribution of gentamicin in the guinea pig inner ear after local or systemic application. J. Assoc. Res. Otolaryngol. 4:176–195, 2003.
Ishiyama G, Lopez I, Baloh RW, Ishiyama A. Histopathology of the vestibular end organs after intratympanic gentamicin failure for Meniere’s disease. Acta. Otolaryngol. 127:34–40, 2007.
Li L, Nevill G, Forge A. Two modes of hair cell loss from the vestibular sensory epithelia of the guinea pig inner ear. J. Comp. Neurol. 355:405–417, 1995.
Lindeman HH. Regional differences in sensitivity of the vestibular sensory epithelia to ototoxic antibiotics. Acta Otolaryngol. (Stockh.) 67:177–189, 1969.
Lopez I, Honrubia V, Lee SC, Schoeman G, Beykirch K. Quantification of the process of hair cell loss and recovery in the chinchilla crista ampullaris after gentamicin treatment. Int. J. Dev. Neurosci. 15:447–461, 1997.
Lysakowski A, Goldberg JM. A regional ultrastructural analysis of the cellular synaptic architecture in the chinchilla cristae ampullares. J. Comp. Neurol. 389:419–443, 1997.
Marcotti W, van Netten SM, Kros CJ. The aminoglycoside antibiotic dihydrostreptomycin rapidly enters mouse outer hair cells through the mechano-electrical transducer channels. J. Physiol. 567:505–521, 2005.
Matsui JI, Ogilvie JM, Warchol ME. Inhibition of caspases prevents ototoxic and ongoing hair cell death. J. Neurosci. 22:1218–1227, 2002.
Migliaccio AA, Minor LB, Carey JP. Vergence-mediated modulation of the human angular vestibulo-ocular reflex is eliminated by a partial peripheral gentamicin lesion. Exp. Brain Res. 159:92–98, 2004.
Moravec WJ, Peterson EH. Differences between stereocilia num-bers on type I and type II vestibular hair cells. J. Neurophysiol. 92:3153–3160, 2004.
Morita I, Komatsuzaki A, Tatsuoka H. The morphological differences of stereocilia and cuticular plates between type-I and type-II hair cells of human vestibular sensory epithelia. ORL J Otorhinolaryngol. Relat. Spec. 59:193–197, 1997.
Myrdal SE, Johnson KC, Steyger PS. Cytoplasmic and intra-nuclear binding of gentamicin does not require endocytosis. Hear. Res. 204:156–169, 2005.
Nakagawa T, Yamane H, Shibata S, Nakai Y. Gentamicin ototoxicity induced apoptosis of the vestibular hair cells of guinea pigs. Eur. Arch. Otorhinolaryngol. 254:9–14, 1997.
Sahni RS, Paparella MM, Schachern PA, Goycoolea MV, Le CT. Thickness of the human round window membrane in different forms of otitis media. Arch. Otolaryngol Head Neck Surg. 113:630–634, 1987.
Sandoval RM, Dunn KW, Molitoris BA. Gentamicin traffics rapidly and directly to the Golgi complex in LLC-PK(1) cells. Am. J. Physiol. Renal Physiol. 279:F884–F890, 2000.
Schacht J. Biochemical basis of aminoglycoside ototoxicity. Otolaryngol. Clin. North Am. 26:845–856, 1993.
Smith CE, Goldberg JM. A stochastic afterhyperpolarization model of repetitive activity in vestibular afferents. Biol. Cybern. 54:41–51, 1986.
Song BB, Sha SH, Schacht J. Iron chelators protect from aminoglycoside-induced cochleo- and vestibulo-toxicity. Free Radic. Biol. Med. 25:189–195, 1998.
Steyger PS, Peters SL, Rehling J, Hordichok A, Dai CF. Uptake of gentamicin by bullfrog saccular hair cells in vitro. J Assoc Res Otolaryngol 4:565–578, 2003.
Swartz DJ, Santi PA. Immunolocalization of tenascin in the chinchilla inner ear. Hear. Res. 130:108–114, 1999.
Urbaniak G, Plous S. Research Randomizer. 2006. Social Psychology Network. 2006.
Walsh RM, Hackney CM, Furness DN. Regeneration of the mammalian vestibular sensory epithelium following gentamicin-induced damage. J. Otolaryngol. 29:351–360, 2000.
Webster JC, McGee TM, Carroll R, Benitez JT, Williams ML. Ototoxicity of gentamicin. Histopathologic and functional results in the cat. Trans. Am. Acad. Opthalmol. Otolaryngol. 74:1155–1165, 1970.
Xue J, Peterson EH. Hair bundle heights in the utricle: differences between macular locations and hair cell types. J. Neurophysiol. 95:171–186, 2006.
Acknowledgement
Support was provided by a grant from the Research Fund of the American Otological Society (Carey) and by grants from the National Institute on Deafness and Other Communication Disorders: K23 DC00196 and R03 DC05700 (Carey), K08 DC0062160 (Della Santina), T32 DC00027 (Lloyd Minor), and P30 DC005211 (Paul Fuchs). Confocal microscopy was made possible by a grant from the National Eye Institute: P30EY001765 (The Wilmer Eye Institute Microscopy and Imaging Core Facility). We would like to thank Mohamed Lehar, Mary Ellen Pease and Rhonda Grebe and Anna Lysakowski’s laboratory for technical support.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Lyford-Pike, S., Vogelheim, C., Chu, E. et al. Gentamicin is Primarily Localized in Vestibular Type I Hair Cells after Intratympanic Administration. JARO 8, 497–508 (2007). https://doi.org/10.1007/s10162-007-0093-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10162-007-0093-8