
Abstract
Objective
The aim of this study was to evaluate the changes in follicular development, serum hormonal levels, and endometrium in the pre-implantation period of rats by using recombinant FSH (rFSH) without human chorionic gonadotropin (hCG).
Methods
Thirty female rats were studied in six groups of five specimens. Two groups determined as controls (groups 1 and 2). Two groups received constant doses of rFSH (groups 3 and 4) and other two decreasing doses (groups 5 and 6). One of the paired groups was mated. Uterus, ovaries, and blood samples were taken from non-mated groups (groups 1, 3, and 5) at the proestrus period and from mated groups (groups 2, 4, and 6) in the pre-implantation period.
Results
In non-mated groups antral follicles and corpus luteum periodicum and in mated groups antral follicles, corpus luteum periodicum, and corpus luteum graviditatis were increased in rFSH groups, especially in decreasing dose groups. Estradiol (E2) levels were increased and progesterone (P)/E2 ratio was significantly decreased in decreasing dose groups. Endometrium surface epithelium was columnar, irregular, and folded in rFSH groups. Endometrium glandular epithelium was cuboidal in all groups. In decreasing dose groups endometrial stroma was smooth and fibroblastic. Mitotic indices of endometrium surface, glandular epithelium, and stroma were significantly decreased in rFSH groups. Primary follicles and P levels showed no change.
Conclusion
It seems likely that decreasing doses of rFSH might be used in order to improve follicular development, although it has negative effects with E2 on endometrium in the pre-implantation period of rats.
Similar content being viewed by others

Introduction
In most of the assisted reproductive programs gonadotropins are used in order to have many follicles and to increase the fertilization rates [1]. Although the progress in ovulation induction and assisted reproductive technologies have increased the fertilization rates up to 60–70%, implantation rates are still detected to be the same at 10–15% [2] and this can be explained by an impaired uterine milieu [3]. In humans and rats endometrium was influenced consequently by E2 and progesterone (P) before implantation. For successful embryo attachment and implantation the synchronization between endometrium and embryo development is also important.
Exogenous gonadotropins disturb this synchronization by changing endogenous hormone levels and endometrium morphology. These changes can cause implantation failure [4, 5]. Uterine receptivity is also important for implantation but the factors effecting receptivity is not well understood yet. The human chorionic gonadotropin (hCG) can both effect follicular and endometrial development [6, 7]. Therefore, hCG was not administered in order not to mask pure rFSH effects. In this study our aim was to evaluate the ovarial, endometrial, and hormonal changes in the pre-implantation period of rats by using rFSH only.
Materials and methods
Animals
This study was carried out in the Medical Faculty Experiment and Research Laboratories of Trakya University, with the approval of the ethic committee and in agreement with the Guidelines for Care and Use of Experimental Animals. Thirty mature, healthy, 10 months old female rats of a Wistar strain having a 5 days regular estrous cycle with body weights between 180 and 250 g were included in the study. They were kept under conditions of 12 h light and 12 h dark periods, at a constant temperature of 21 ± 1°C and humidity of 40–60%. They had free access to standard dry pellets ad libitum and tap water until the end of the study. Rats followed at least three successive normal estrous cycles then they were randomized and studied in six groups of five specimens.
Estrous cycle determination
Estrous cycles of rats were determined by taking daily vaginal smears. Vaginal smears were taken by cotton rods, spread on lams, fixed in 5% alcohol, stained with 5% Giemsa, and than evaluated under the light microscope Olympus BH-2 (Tokyo, Japan). Estrous cycle were determined as follows; proestrus period (centrally nucleolated many epithelial cells), estrus period (cornified epithelial cells without nucleus), metestrus period (leukocyte, mucus, and a few cornified cells), diestrus period (various epithelial cells, mucus, and leukocyte) [8].
Gonadotropin treatment
Drug application period was defined as diestrus 1, 2, and 3 by uniting metestrus and diestrus periods as described by Van Cappelen et al. [9]. All drugs were administered every morning at 10 h Groups 1 and 2 were defined as controls and received 0.2 ml saline subcutaneously in diestrus 1, 2, and 3. Groups 3 and 4 were defined as constant dose groups and were given 16 IU rFSH (Puregon®, N.V. Organon, Oss, The Netherlands) subcutaneously in diestrus 1, 2, and 3. Groups 5 and 6 were defined as decreasing dose groups and were given 16, 8, and 4 IU rFSH subcutaneously consequently in diestrus 1, 2, and 3.
Groups 2, 4, and 6 were mated with male rats of proven fertility at the proestrus period. Mating was confirmed with the observation of a vaginal plug or presence of spermatozoons in the vaginal smears [3]. Mating time was recorded as day 0.5 of pregnancy. After 48 h vaginal smears were taken again from the mated rats and if pregnancy type changes (abundant leukocyte and mucus) were seen pregnancy was confirmed [3]. Rats in groups 2, 4, and 6 were sacrificed on day 2.5 of pregnancy at the pre-implantation period. If pregnancy did not occur rats were pursued for maximum two more cycles. Rats in groups 1, 3, and 5 were sacrificed at the proestrus period.
General anesthesia was achieved by administering ketamine 35 mg/kg (Ketalar® 50 mg/ml Pfizer, Ortakoy, Istanbul, Turkey) and xylazine–hydrochloride 15 mg/kg (Rompun® %2 50 cm3 Bayer-Turk Ltd., Umraniye, Istanbul, Turkey) subcutaneously. A midline ventral laparotomy was performed. Uterus, bilateral ovaries, and uterine horns were removed. Rats were killed by taking out intracardiac blood. Uterus, ovaries, and uterine horns were fixed in formalin until examination.
Histological analysis
Ovaries, uterus, and uterine horns were studied according to the standard tissue procedures and embedded in paraffin wax. Fine sections of 5 μm were taken and stained with hematoxylin and eosin. Histological examination was performed for each of the slides under light microscope. Images were transferred to computer by using JVC Color Video Camera Head Model No: TK-890E (Tokyo, Japan), AverMedia, AverTV WDM Video Capture (Software) and digitally photographed. For ovarian detection serial sections were performed. Using UTHSCSA Image Tool for Windows Version 3.00 (Software) follicle (primary follicle; 30–400 μm, antral follicles; 650–700 μm) and corpora lutea (corpus luteum periodicum; about 700 μm, corpus luteum graviditatis; about 1,100 μm) dimensions were estimated according to Hebel and Stromberg [10] and counted stereological by physical dissector method. Uterine horns and uterus were detected with ten non-serial sections. Uterine horns were examined for widening and vascularization. Uteruses were evaluated for the general histological components of the surface and glandular epithelium and stroma. Mitosis in 1,000 cells were counted and mitotic indices for surface epithelium, glandular epithelium, and stroma were determined.
Hormonal analysis
Blood samples were centrifuged for 10 min at 3,000 g then serum was separated from plasma and stored at −70°C until measurement. E2 and P levels were measured by radioimmunoassay method by using commercially available radioimmunoassay kit Immulite 2000 (Diagnostic Products Corporation Los Angeles, CA, USA).
Statistical analysis
In the statistical analysis suitability of variables to normal distribution was measured using the Kolmogrov–Smirnov test. Variant analysis and the Kruskal–Wallis analysis were used in order to compare groups for variables, and chi-square was used in order to compare categorical variables. Pearson’s correlation analysis was used to compare the relationship between variables. Data are presented as mean ± SD if they were normally distributed and as mean rank if not. Statistical analysis was carried out using SPSS 13.0 (Statistical Package for Social Science Inc., Chicago, IL, USA). P < 0.05 was considered as statistically significant.
Results
No statistical difference was found between the mean number of primary follicles in the non-mated and mated groups. Among the non-mated groups the mean number of antral follicles and corpus luteum periodicum were significantly increased in rFSH groups (groups 3 and 5) (P = 0.03 and 0.02, respectively) and this was more evident between control group (group 1) and decreasing dose group (group 5) (P = 0.024 and 0.018, respectively) (Fig. 1). In the mated groups the mean number of antral follicles, corpus luteum periodicum, and corpus luteum graviditatis were significantly increased in rFSH groups (groups 4 and 6) (P = 0.002, 0.014, and 0.01, respectively) and this was more evident between control group (group 2) and decreasing dose group (group 6) (P = 0.002, 0.001, and 0.008, respectively) (Fig. 1). In the correlation analysis a positive correlation was detected between antral follicles and corpus luteum periodicum for all groups (P = 0.003) and in the mated groups a positive correlation was detected between antral follicles and corpus luteum graviditatis and between corpus luteum periodicum and corpus luteum graviditatis (P = 0.035 and 0.001, respectively).

Mean numbers of primary follicle, antral follicle, corpus luteum periodicum, and corpus luteum graviditatis for each group
Among the non-mated groups and mated groups E2 levels were significantly increased in rFSH groups (P <, 0.01, respectively) and this significant difference was more evident in decreasing dose groups than the controls (P < 0.01 and 0.01, respectively) (Fig. 2). No significant difference was observed in P levels (P > 0.05). Among the non-mated and mated groups regarding the P/E2 ratio a significant difference was found (P = 0.003 and 0.009, respectively) and this was more evident between controls and decreasing dose groups (P = 0.01 and 0.04, respectively). P/E2 ratio was significantly decreased in decreasing dose groups (Fig. 3).

Mean values of estradiol and progestrone

Progestrone/estradiol ratio for each group
In the correlation analysis for all groups a positive correlation was detected between E2 and antral follicles and corpus luteum periodicum (P = 0.005) and in the mated groups a positive correlation was detected between E2 and corpus luteum graviditatis (P < 0.001 and P = 0.018, respectively).
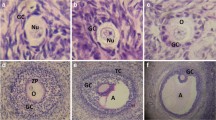
Uterine horns were found dilated and vascularized in mated groups. In controls endometrium was found cuboidal and smooth. In all rFSH groups endometrium surface epithelium was converted from cuboidal to columnar and an irregular and a folded structure was observed. Endometrium glandular epithelium was found cuboidal in all groups. Endometrial stromal decidual reaction was evident in controls and in constant dose groups but in decreasing dose groups the number of smooth and fibroblastic cells were elevated and no decidualization was observed (Fig. 4).

a Control group: Endometrium surface epithelium was cuboidal and smooth (arrow, endometrium cross-section × 40). b Decreasing dose group: Endometrium surface epithelium was converted to columnar and folded (arrowhead, endometrium cross-section × 40). c Stromal decidualization in the control group (arrow, endometrium cross-section × 40). d Decreasing dose group: Fibroblastic cells took place instead of decidualization in stroma (arrowhead, endometrium cross-section × 40)
Mitotic indices of endometrium surface epithelium, glandular epithelium, and stroma were significantly decreased in all rFSH groups compared with controls. As we compared the endometrium surface epithelium mitotic indices of the non-mated groups statistically significant difference was found, this was especially evident between the control group and the constant dose group. There was a manifest decrease in the constant dose group. No significant difference was found between the glandular epithelium mitotic indices of the non-mated groups. Statistically significant difference was found between the stromal mitotic indices of the non-mated groups, this difference was more evident between the control group and the constant dose group. There was a manifest decrease in the constant dose group. As we compare the endometrial surface epithelium and stroma mitotic indices of the mated groups no difference was detected but significant difference was found between the glandular epithelium mitotic indices of the mated groups. This difference was more evident between the control group and the constant dose group and between the control group and the decreasing dose group. There was a manifest decrease in the constant dose and the decreasing dose group (Table 1).
In the correlation analysis a negative correlation was detected between E2 and endometrium surface epithelium, glandular epithelium, and stroma mitotic indices in all groups (P = 0.009, 0.003, and 0.008, respectively) and a positive correlation was observed between the P/E2 ratio and endometrium surface epithelium, glandular epithelium, and stroma mitotic indexes for all groups (P = 0.03, 0.0043, and 0.005, respectively).
Discussion
It is a well known and a frequently used method to stimulate ovaries with exogenous gonadotropins in order to have many follicles [11]. The hCG has been used for a long time to trigger luteinizing hormone (LH) peak to induce ovulation with rFSH. It is also known that hCG has direct effects on endometrium via the endometrial hCG/LH receptors [7] and that hCG can induce ovarian follicle development via the granulose cell hCG/LH receptor [6]. The hCG can induce the transformation of endometrial stromal cells to decidua and this effect can be imitated by LH but not by FSH [12]. Because hCG can possibly mask or augment unfavorable or positive effects of FSH no hCG was performed to measure the-only FSH effects.
In superovulation procedures dose regimen is also important to make the best stimulation [9]. Therefore we applied constant and decreasing doses of rFSH. The beginning of follicle development and depletion of the resting follicle pool was independent of gonadotropin stimulation [13]. When we compared all groups we found that the number of primary follicles was similar and this supports that primary follicle development is independent of FSH. As opposed to the development of early follicles, FSH was necessary for large antral preovulatory follicle development [13]. In our study the number of antral follicles, corpus luteum periodicum, and corpus luteum graviditatis was elevated in all rFSH groups. These findings also pointed out that advanced follicle development was depending on FSH stimulation. In our study the number of antral follicles, corpus luteum periodicum, and corpus luteum graviditatis were elevated particularly in the decreasing dose group. This evident increase in the decreasing dose group seems to be associated with the FSH threshold concept. In the normal menstrual cycle surpass of threshold level is limited to a certain period with the decrease in FSH [14]. Thus, application of decreasing doses of rFSH mimics the normal cycle and the number of selected oocytes increases.
The E2 levels were elevated in all rFSH groups and this is concordant with the two cell two gonadotropin theory [13]. A positive correlation was detected between E2 and antral follicles and corpus luteum periodicum in all groups and between E2 and corpus luteum graviditatis in the mated groups. These results supported the relation between E2 levels and increasing number of advanced stage follicles related to rFSH application. No difference was observed in P levels between the experiment groups and this supports that rFSH treatment does not affect P levels, which was also similarly reported by Kramer et al. in rats [3]. When compared with controls P levels were same despite the increase in follicle numbers in all rFSH groups, which suggested that P had no role in follicle development. Our results also supported by Van Dessel et al. [15].
The endometrium plays an important physiologic role for the implantation of embryos [1]. Development of enough receptive endometrium is the main factor determining the outcome in IVF [16]. It was shown that the stimulation related to gonadotropin application can disturb the endometrial receptivity in IVF [2]. It was also reported that supraphysiologic levels of hormones had a negative impact on endometrial development and differentiation [17–21]. In our study endometrium surface epithelium was cuboidal and smooth in control groups while it was converted to columnar with an irregular and folded structure in all rFSH groups. These findings suggest that rFSH application can disturb the endometrium surface epithelium and might have a negative impact on endometrium whether pregnancy takes place or not. This corruption in the pre-implantation period can possibly cause a failure in the implantation.
Decidualization of stroma is an important pre-condition for successful maintenance of pregnancy and the decrease in stromal decidualization is a negative factor for fertility [4]. We found out that endometrial stromal decidual reaction was evident in constant dose and control groups and additionally smooth and fibroblastic cells were observed instead of decidualization in decreasing dose groups. Although the decidualization of stroma was detected in constant dose groups, no decidualization was observed in decreasing dose groups and this cannot be explained by the direct negative impact of FSH on stroma. This condition can be explained by the negative impact of high E2 levels in the decreasing dose groups.
In all rFSH groups compared with controls mitotic indices of endometrium surface epithelium, glandular epithelium, and stroma were detected to be decreased. In non-mated groups the decrease in mitotic indices of endometrium surface epithelium, glandular epithelium, and stroma was more evident in the constant dose groups. It was concluded that in non-mated groups rFSH had a negative impact on mitotic indices. In mated groups the decrease in mitotic indices of endometrium surface epithelium and glandular epithelium was more evident in the constant dose group and the decrease in mitotic index of stroma was more evident in decreasing dose group. Also in mated groups it was shown that rFSH had a negative impact on mitotic indices. Both in mated and non-mated groups the significant decrease in the mitotic indices of endometrium surface epithelium and glandular epithelium can be explained by direct negative effect of rFSH on endometrium surface epithelium and glandular epithelium.
The E2 levels increased both in constant and decreasing dose groups and although this increase was more evident in decreasing dose groups, the decrease in mitotic indices of endometrium surface and glandular epithelium was much more in the constant dose groups. Therefore, a direct negative impact of E2 on endometrium surface and glandular epithelium does not seem to be likely. It is also known that E2 increase the mitotic indices of endometrium surface epithelium, glandular epithelium, and stroma [3]. Nevertheless, the cumulative rFSH dose was higher in the constant dose group than the decreasing dose group. More significant decrease in the mitotic indices of endometrium surface and glandular epithelium in the constant dose group supports that the endometrium surface and glandular epithelium was negatively affected in relation with rFSH dose.
The significant decrease in stromal mitotic indices in the constant and decreasing dose groups (mated groups) can be explained by both direct negative effects of rFSH and in the decreasing dose group by increased E2 levels. Stromal decidualization was insufficient in decreasing dose groups and in mated groups particularly in the decreasing dose group the significant decrease in the mitotic indices appears to be due to the level of E2 outcome rather than FSH’s negative effect. These findings suggest that high doses of rFSH have a direct negative impact on the endometrium [4, 22]. Elevated E2 levels seem to initially produce a negative impact on stroma [18, 19]. However, as the impact level and impact duration of E2 increase it can cause damage on the other layers of endometrium [3]. These are supported by the negative correlation between E2 and endometrium surface epithelium, glandular epithelium, and stroma mitotic indices and the positive correlation between P/E2 ratio and endometrium surface epithelium, glandular epithelium, and stroma mitotic indices.
Gidley-Baird et al. [23] demonstrated that P/E2 ratio is important and P prevents the inhibiting effect of E2 on implantation in humans and mice. We also emphasize the importance of P/E2 ratio but we did not measure any protective effect of P. To augment uterine response in patients that give high response to controlled hyperstimulation, Simon et al. [24] applied decreasing doses of FSH on the following cycle to those that gave high response and they detected decreased pre-implantation E2 levels and increased implantation and pregnancy rates. But in our study in decreasing dose groups (due to high E2 levels) in the pre-implantation period stromal decidualization did not occur. This difference between two studies could be the result of not using hCG in the present study.
In conclusion, the clinical effect of drugs used in experimental studies carried out on animals may be expected to differ between subjects with different tissue characteristics, however if translated to its model, variations of responses in individual experiments may be masked in analysis of the whole group. Additionally, due to the limited number of sample size in the present study, it is not possible to make a strict conclusion but it seems likely that decreasing doses of rFSH might be used in order to improve ovarian follicular development although it might cause inappropriate endometrial development.
In this experimental study we aimed to determine the-only rFSH effects without hCG priming in the pre-implantation period of rats. Addition of hCG to rFSH might change its effect (deteriorate, improve or no change) therefore, studies investigating this effect would be interesting to know which warrants further research.
References
Basir GS, WS O, Ng EHY, Ho PC (2001) Morphometric analysis of peri-implantation endometrium in patients having excessively high oestrodiol concentrations after ovarian stimulation. Hum Reprod 16(3):435–440
Ng EHY, Yeung WSB, Lau EYL, So WWK, Ho PC (2000) High serum oestrodiol concentrations in fresh IVF cycles do not impair implantation and pregnancy rates in subsequent frozen-thawed embryo transfer cycles. Hum Reprod 15(2):250–255
Kramer B, Stein BA, Van Der Walt LA (1990) Exogenous gonadotropins- serum oestrogen and progestrone and the effect on endometrial morphology in the rat. J Anat 173:177–186
Stein B, Kramer B (1989) The effect of exogenous gonadotropic hormones on the endometrium of the rat. J Anat 164:123–130
Kramer B (1997) Changes in vascular permeability and deciduoma formation during the peri-implantation period of the rat in response to exogenous gonadotropins. Anat Rec 247:20–24
Filicori M, Cognigni GE, Taraborrelli S, Parmegiani L, Bernardi S, Ciampaglia W (2002) Intracytoplasmic sperm injection pregnancy after low-dose human chorionic gonadotropin alone to support ovarian folliculogenesis. Fertil Steril 78(2):414–416
Fanchin R, Peltier E, Frydman R, de Ziegler D (2001) Human chorionic gonadotropin: does it affect human endometrial morphology in vivo? Semin Reprod Med 19(1):31–35
Waynforth HB, Flecknell PA (1991) Experimental and surgical technique in the rat. Jovanovich Publishers, London, pp 346–357
Van Cappellen WA, Kramer P, Van Leeuwen ECM, De Leeuw R, De Jong FH (1997) Induction of superovulation in cyclic rats by administration of decreasing doses of recombinant follicle stimulating hormone (Org32489). Hum Reprod 12(2):224–230
Hebel R, Stromberg MW (1976) Anatomy of the laboratory rat. The Williams & Wilkins Company, Baltimore, pp76–82
Daya S, Gunby J (1999) Recombinant versus urinary follicle stimulating hormone for ovarian stimulation in assisted reproduction. Hum Reprod 14(9):2207–2215
Han SW, Lei ZM, Rao CV (1999) Treatment of human endometrial stromal cells with chorionic gonadotropin promotes their morphological and functional differentiation into decidua. Mol Cell Endocrinol 147(1–2):7–16
Macklon NS, Fauser BCJM (2001) Follicle-stimulating hormone and advanced follicle development in the human. Arch Med Res 32:595–600
Macklon NS, Fauser BCJM (1998). Follicle development during the normal menstrual cycle. Maturitas 30:181–188
Van dessel HJ, Schipper I, Pache TD, Van Geldrop H, De Jong FH, Fauser BCJM (1996) Normal human follicle development: an evaluation of corelations with oestradiol, androstenedione and progestrone levels in individual follicles. Clin Endocrinol 44:191–198
Ben-Nun I, Jaffe R, Fejing MD, Beyth Y (1992) Therapeutic maturation of endometrium in in vitro fertilization and embryo transfer. Fertil Steril 57(5):953–962
Paulson RJ, Sauer MV, Lobo RA (1990) Embryo implantation after human in vitro fertilization: importance of endometrial receptivity. Fertil Steril 53(5):870–874
Bonhoff A, Johannisson E, Bohnet HG (1990) Morphometric analysis of the endometrium of infertile patients in relation to peripheral hormone levels. Fertil Steril 54(1):84–89
Bonhoff A, Naether O, Johannisson E, Bohnet HG (1993) Morphometric characteristics of endometrial biopsies after different types of ovarian stimulation for infertility treatment. Fertil Steril 59(3):560–566
Pellicer A, Remohi J, Valbuena D, Simon C, Cano F (1996) Lower implantation rates in high responders: evidence for an altered endocrine milieu during the preimplantaion period. Fertil Steril 65(6):1190–1195
Noci I, Borri P, Coccia ME, Criscuoli L, Scarselli G, Messeri G, et al (1977) Hormonal patterns,steroid receptors and morphological pictures of endometrium in hyperstimulated IVF cycles. Eur J Obstet Gynecol Reprod Biol 75(2):215–220
Stein BA, Kramer B, De Wet G, Van Der Walt LA (1993) Dose-dependent effects of exogenous gonadotropins on the endometrium of the rat. S Afr Med J 83(2):122–125
Gidley-Baird AA, O’Neill C, Sinosich MJ, Porter RN, Pike IL, Saunders DM (1986) Failure of implantation in human in vitro fertilization and embryo transfer patients: the effects of altered progestrone/estrogen ratios in humans and mice. Fertil Steril 45(1):69–74
Simon C, Velasco JJG, Valbuena D, Peinado JA, Moreno C, Remohi J, et al (1998) Increasing uterine receptivity by decreasing estradiol levels during the preimplantation period in high responders with the use of a follicle-stimulating hormone step-down regimen. Fertil Steril 70(2):234–239
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Canda, M.T., Kucuk, M., Bagriyanik, H.A. et al. The only effects of recombinant follicle-stimulating hormone for ovulation induction in the pre-implantation period of rats. Arch Gynecol Obstet 275, 99–105 (2007). https://doi.org/10.1007/s00404-006-0236-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00404-006-0236-x