
Abstract
Semi-sessile Mytilus mussels are used as indicators of climate changes, but their geographic distribution is not sufficiently known in the Arctic. The aim of this study was to investigate the taxonomic status and genetic differentiation of Mytilus populations in a Northwest Greenlandic fjord at Maarmorilik, impacted by contaminations from a former mine. In this study, mussels were collected at three sites differing in exposure to environmental factors. A total of 54 polymorphic SNPs found in the Mytilus EST and DNA sequences analyzed were successfully applied to 256 individuals. The results provided the first evidence for the existence of M. trossulus in Greenland. The mussel from M. trossulus and M. edulis taxa are shown to coexist and hybridize in the fjord. The three studied sites were found to differ significantly in the distribution of taxa with a higher prevalence of M. trossulus in the inner fjord. The identified M. edulis × M. trossulus hybrids mostly had a hybrid index score of about 0.5, indicating a similar number of alleles characteristic for M. trossulus and M. edulis. There was a low number of backcrosses between ‘pure’ taxa and hybrids. This newly discovered hybrid zone between the two taxa is unique in comparison with the Canadian populations. As Mytilus mussels in Greenland hitherto have been regarded as the one taxon M. edulis, the results have importance for biogeography and future monitoring and environmental studies.
Similar content being viewed by others

Introduction
The blue mussel, Mytilus, is known to inhabit subarctic and arctic regions of Norway, Russia, Alaska, Canada and Greenland in addition to temperate areas of both the Northern and Southern hemispheres (Riginos and Henzler 2008; Väinölä and Strelkov 2011). In Greenland, populations of blue mussels are widely found in fjords in West and South Greenland, but they have not been found north of central east Greenland (Versteegh et al. 2012; Høgslund et al. 2014). The M. edulis species complex consists of three morphologically similar congeners: M. edulis, M. trossulus and M. galloprovincialis. Recently research has been intensified on the distribution of the species complex (e.g., Moreau et al. 2005; Shields et al. 2008; Riginos and Henzler 2008; Smietanka et al. 2009; Stuckas et al. 2009; Zbawicka et al. 2012; Smietanka et al. 2013, 2014; Zbawicka et al. 2014), which evidently reflects a history of inter- and transoceanic dispersal, secondary contact and hybridization, besides the effect brought about by human activities (Väinölä and Strelkov 2011). It is thought that the complex originated from M. trossulus in the Pacific followed by colonization in the Northern Atlantic giving rise to both M. edulis and M. galloprovincialis (Vermeij 1992; Riginos and Cunningham 2005). While until recently it has been believed that the distribution of M. trossulus was restricted to the North Pacific, eastern Canada and the Baltic Sea (Riginos and Cunningham 2005), more recent research has documented its occurrence on the coasts of Scotland, Iceland, the Barents Sea, the White Sea and Norway (Beaumont et al. 2008; Zbawicka et al. 2010; Kijewski et al. 2011; Väinölä and Strelkov 2011). The presence of M. edulis has been reported in the northern part of the Atlantic and European seas from the White and Barents Sea to the Atlantic coast of southern France and M. galloprovincialis in the Mediterranean Sea, Black Sea and along the Atlantic coastline of Western Europe including the British northern islands (e.g., Väinölä and Hvilsom 1991; Borsa et al. 1999; Bierne et al. 2003; Riginos and Cunningham 2005; Riginos and Henzler 2008; Zbawicka et al. 2012, 2014).
It is known that there is an overlap between M. edulis and M. trossulus throughout the North Atlantic with both co-occurrence and interbreeding (Riginos and Cunningham 2005; Gardner and Thompson 2009; Väinölä and Strelkov 2011; Zbawicka et al. 2012). It has been suggested that the colonization of Greenland by blue mussels occurred recently and exclusively from North America (Riginos and Henzler 2008). The occurrence of Mytilus in Greenland has until now been considered to consist of a single taxon, M. edulis. Few genetic studies have been performed on Greenlandic mussels. Versteegh et al. (2012) performed RFLP fingerprinting on mussels collected in an area restricted to Godthaabsfjord, central west Greenland. Wanamaker et al. (2007) sampled outside Sisimiut in open water and used genetic PCR-based markers, and Riginos and Henzler (2008), who applied mtDNA analysis of populations in the very south of Greenland, found that all sampled mussels belonged to M. edulis.
Mytilus edulis is important in the coastal areas in Greenland and has been applied in a range of ecology studies (e.g., Theisen 1973; Strand and Asmund 2003; Wanamaker et al. 2007; Blicher et al. 2013; Høgslund et al. 2014; Thyrring et al. 2015). In the mid-intertidal zone, blue mussel beds are found to dominate at wave-exposed steep sites. Such beds are highly productive, and besides being the primary food source for birds, fish and invertebrates, they provide habitat and refuge for numerous organisms (Blicher et al. 2013). Further, Mytilus has been used extensively in environmental monitoring studies, for example around mining-related activities to monitor for element dispersal (e.g., Riget et al. 1997, 2000; Søndergaard 2013; Søndergaard et al. 2014; Bach et al. 2014). Whether related to monitoring or to basic ecology, populations have been assumed to belong to this single taxon M. edulis only.
In Greenland, mussels must conform to extremely harsh environmental conditions. Mytilus is part of the intertidal and subtidal hard-bottom communities, where adults are attached by byssus threads to rocks or stones in shallow waters, from the tidal zone down to depths of several meters. In Greenland, the mussels may experience long periods during winter where the sea freezes and the mussels thus are enclosed by ice. They can survive long dormant periods and the shift between freezing and thaw in spring and autumn, but do not survive if the summer period is too short (Bennike and Wagner 2013). The fjord water forms in early summer after the ice break and disappears when the sea ice begins to form. Due to the influence of melt water, the salinity is reduced and temperature is elevated because of the solar insolation. Thus, temperatures in inner fjords may reach 5–8 °C in surface waters, while the temperature at a few meters depth is 2–4 °C. While M. edulis for example requires summer temperatures of a minimum of 4 °C (Bennike and Wagner 2013), studies on larval development have shown that M. trossulus is less tolerant to temperatures at 10 °C and above than M. edulis (Hayhurst and Rawson 2009). Further, M. trossulus has been found to inhabit lower salinity areas (Väinölä and Hvilsom 1991; Ridgway and Naevdal 2004; Zbawicka et al. 2010, 2014; Väinölä and Strelkov 2011). Experimental studies have shown that this taxon is less sensitive to lower salinities in the early ontogenetic stages than its congeners (Qiu et al. 2002). It is thus hypothesized that the distribution of M. edulis and M. trossulus is influenced greatly by temperature and salinity, with an expectation of a higher frequency of M. trossulus in cold and low salinity waters. However, no research has yet reported the presence of M. trossulus in Greenland.
The aim of this study was to investigate the taxonomic status and genetic differentiation of Mytilus populations in a Northwest Greenlandic fjord at Maarmorilik. The fjord system was impacted by elevated concentrations of contaminants, especially lead (Pb) and zinc (Zn), in seawater, sediments and surrounding biota discharged from a mine in the period 1973–1990. Mytilus mussels have been extensively used for monitoring studies in the fjord system, which is also influenced by changes in salinity during spring and summer as well as ice cover during winter time. This main aim is achieved by comparison of the Greenland Mytilus populations with European and North American M. edulis and M. trossulus populations using SNP markers. As Mytilus mussels in Greenland hitherto have been regarded as one taxon M. edulis, the results can thus have importance for biogeography, monitoring and environmental studies.
Methods
DNA sampling
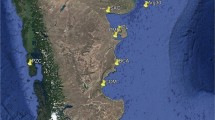
Mytilus spp. samples, consisting of 256 individuals in total, were collected from nine localities (Fig. 1; Table 1). Adult mussels of mixed age, sexes and size 15–50 mm were collected from two different geographic scales—from three localities in a fjord in Northwest Greenland in an area of a former lead-zinc mine, the Black Angel at Maarmorilik (GLL, GLS and GLD), and from reference samples from both sides of the Atlantic. Three reference samples of M. edulis were obtained, one from the Atlantic coast of North America at Indian River, Delaware (IRD), and two from the Atlantic coast of North Europe, at Loire, France (LOI), and Loch Etive, Scotland (LEE). Two reference samples of M. trossulus were obtained, one from the Atlantic coast of North America at Placentia Bay, Newfoundland (PBAT), and one from the Atlantic coast of North Europe at Loch Etive, Scotland (LET). Two reference samples of M. galloprovincialis were obtained, one from the Atlantic coast of North Europe at Bidasoa, Spain (BID), and one from the Adriatic Sea at Lido, Italy (LID). Most of the samples were collected in 2012, with four in the years 2006–2008 (Table 1), and stored frozen at −20 °C until the SNP analyses. Seven samples consisted of about 30 individuals each, and three reference samples from Scotland and Newfoundland had from 9 to 15 individuals. DNA was isolated from the mantle tissue using a modified CTAB method according to Hoarau et al. (2002) and suspended in sterile, filtered, distilled water.

Geographic location of ten Mytilis spp. sampling sites. See Table 1 for definition of site abbreviations
Identification and genotyping of SNPs
The EST sequences (Table 2) were aligned with appropriate GenBank EST sequences (mainly from M. galloprovincialis and M. edulis) using the ClustalX program version 1.83 (Thompson et al. 1997) with default settings. Multiple sequence alignments were used for SNP discovery using the Staden software programs (Staden et al. 2001). SNP polymorphisms were detected by visual inspection of aligned sequences. SNPs were scored as heterozygous when two peaks occurring in the sequence trace differed in intensity by <25 %. The possible effect of the SNPs, resulting in change or no change in amino acid sequence (non-synonymous or synonymous changes, respectively), was predicted on the basis of the open reading frames (ORF) identified (Table 2). An average of 21 EST sequences were used to characterize each SNP, with a minimum of four sequences. With a sufficient number of sequences, sequencing errors were disregarded by Staden software; other sequencing errors were identified during the validation. Additional SNPs were identified in Zbawicka et al. (2012, 2014). Genotyping of SNPs was performed using the Sequenom MassARRAY iPLEX platform (Gabriel et al. 2009). PCR and extension primers were designed using the Assay Design 3.1 program (Sequenom). Analysis and scoring were performed using Typer 3.4 software (Sequenom). SNPs were classified, based on manual inspection, as ‘failed assays’ (meaning that the majority of genotypes could not be scored and/or the samples did not cluster well according to genotype), ‘monomorphic SNPs’ (all studied individuals had the same genotype) or ‘polymorphic SNPs.’ Assays were designed for 79 candidate SNPs including 69 based on EST sequences and 10 identified by DNA sequences, selected from 385 putative SNPs (Zbawicka et al. 2012, 2014).
Analysis of genotype data
SNP markers and populations were analyzed for genetic diversity, proportion of polymorphic SNP (P O), observed (H O) and expected (H E) heterozygosity, and inbreeding coefficient (F IS) using Arlequin v3.5 (Excoffier and Lischer 2010). The statistical significance of F IS (>0) was tested by 10,000 permutations of alleles between individuals. Departures from Hardy-Weinberg equilibrium (HWE) were tested by exact test and significance was determined by Markov chain Monte Carlo simulations. Pairwise analysis in Arlequin was used to calculate mean pairwise F ST values defining population differentiation. F ST values at individual SNPs were calculated using the AMOVA function of the same program. Permutation testing with 1000 iterations was used to calculate p values for mean and locus-by-locus F ST values. The program PopTree2 (Takezaki et al. 2010) was used to produce a neighbor-joining tree based on F ST distances from allele frequencies of the SNP loci. The robustness of the phylogenetic relationship calculated by the NJ tree was assessed using 1000 replication bootstrap analysis. Finally the NJ tree was constructed in MEGA version 4 (Tamura et al. 2007).
Correction for multiple tests of overall significance values was carried out using Benjamini-Yekutieli correction for multiple comparisons, a less conservative modified false discovery rate (FDR) method (Narum 2006). A list of loci with F ST ≥ 0.5, having the alleles that distinguish M. trossulus from M. edulis and from M. galloprovincialis, was prepared. The “M. trossulus characteristic allele” was defined as the allele characteristic for pure M. trossulus (about 90–100 %) for each marker. Fourteen SNP markers met the above condition (Table 2). The number of markers showing the “M. trossulus characteristic allele” was counted for each individual. The hybrid index score (HI) for each individual was calculated as the number of markers showing the “M. trossulus characteristic allele” divided by the number of selected markers (14).
A score of zero is a ‘pure’ M. edulis, whereas a score of one is a ‘pure’ M. trossulus. HI was calculated for all individuals and averaged for each population to assess the degree of introgression between M. edulis and M. trossulus. Contingency table analyses were used to compare inner and outer fjord groups for significant differences in species distribution.
GENEPOP 4.1 (Rousset 2008) was used to test for linkage disequilibrium (LD) between all pairwise combinations of 44 loci [one, the most differentiated SNP per fragment, marked in Table 2 for three groups of putatively ‘pure’ populations (3 M. edulis, 2 M. galloprovincialis and 2 M. trossulus)]. For a pair of diploid loci, no assumption was made about the gametic phase in double heterozygotes.
Correspondence analysis (CA; Benzécri 1992), implemented in GENETIX (Belkhir et al. 2003), was used for visualizing the genetic substructure at population and individual levels. The results have been presented as a scatter plot, with the axes representing the contribution of inertia of the data matrix in a way that can be considered analogous to the total variance in allelic frequency (Benzécri 1992).
Results
SNP validation
About 2300 Expressed Sequence Tags (ESTs) obtained from the Baltic M. trossulus × M. edulis hybrid were aligned with appropriate GenBank EST sequences and used to study SNPs. Three hundred eighty-five putative SNPs were found, distributed across 151 contigs.
SNP multiplexes were designed in silico and tested on a panel of 30 mussels from the reference samples. Finally, 79 candidate SNPs identified in 59 contigs and 5 DNA fragments (Zbawicka et al. 2012, 2014) were used to genotype 256 individual mussels from 9 localities. For 16 SNPs, the assay design failed, 6 did not provide an acceptable quality score, and 3 were monomorphic in all samples. The remaining 54 SNPs, found in 4 DNA fragments and 39 EST contigs, with one to three SNPs per fragment, were polymorphic with a high quality score in most of the sampled individuals. SNP annotation is presented in Table 2. Eight SNPs localized in DNA fragments and 25 SNPs identified in EST sequences (Table 2) have already been used (had been previously discovered and described) in the study of the European populations of the Mytilus mussels (Zbawicka et al. 2012, 2014). The remaining 21 SNPs were newly characterized in the present study.
Open reading frames were identified in 38 fragments; only 6 SNPs (marked NA in Table 2) were in fragments without ORFs. Of the 54 SNPs, 43 (79.6 %) were located in coding regions of which 40 (93 %) were synonymous and 3 (7 %) nonsynonymous. Of the SNPs used, five (9.3 %) were located in noncoding regions and six (11.1 %) had unknown location. Of the 54 SNPs, only two (BM21C and BM9C) had more than two alleles each.
Very little linkage disequilibrium between pairs of 44 markers in the three groups of Mytilus spp. populations was found. Only one pair of loci (BM201C vs. BM203B) out of a total of 1892 were in highly significant linkage disequilibrium (p < 0.0001) and concerned only M. galloprovincialis populations.
Hybrid index score and taxonomic status of mussels
Most individuals from Greenland were “pure” M. edulis (hybrid index HI near 0), “pure” M. trossulus (HI near 1) or hybrids with HI near 0.5 (Fig. 2). Only two individuals with HI between 0.7 and 0.9 were observed in one of the Greenlandic populations (GLS). The greatest variance of HI was observed for the population GLL located outside the fjord towards open waters, which was characterized by the highest number of loci with departures from HWE (Table 3). The two populations from the inner Maarmorilik Fjord (GLS and GLD) showed a significantly higher percentage of M. trossulus characteristic alleles (contingency table analysis, p < 0.05), and consequently up to 50 % of individuals had a ‘pure’ M. trossulus genotype. The ‘pure’ M. edulis genotype only was identified in 21 % of the mussels from those two sites.

The frequency distribution of the score for a hybrid index giving the percentage of M. trossulus characteristic alleles (marked in Table) in three populations from Greenland. A score of zero is a pure M. edulis, whereas a score of one is a pure M. trossulus. Data are presented for three populations from Greenland and reference populations of M. trossulus and M. edulis
An estimated 30 and 39 % of individuals were hybrids in the GLD and GLS populations, respectively. Most hybrids (25 individuals) had the hybrid index 0.42–0.59. HI for ten hybrids from GLD populations ranged from 0.50–0.57, for ten hybrids from GLS ranged from 0.42–0.59 and for five hybrids found in GLL ranged from 0.47–0.53. An average of 85 % of the loci in each of the 25 described hybrids (range 64–100 %) was heterozygous for one M. trossulus allele and one M. edulis allele. The presence of a large number of multiple heterozygotes indicates the existence of both the F1 hybrids as well as a certain number of F2 hybrids, which are backcrosses between ‘pure’ taxa and F1 hybrids.
In locus-specific analyses performed for the Greenland populations, different levels of introgressions were observed at the same loci. Fifteen SNPs characteristic of M. trossulus populations were observed in the inner fjord populations with at least 59 % frequency, including three SNPs with at least 75 % frequency (Online Resource 1). However, in the GLL population the frequency of the same loci was about 40 %. A neighbor-joining tree was constructed using only pure individuals identified by HI (near 0 and 1). The Greenland samples grouped respectively within the reference samples of pure M. edulis and M. trossulus from North America (Fig. 3).

Neighbor-joining tree of Mytilus spp. samples from Greenland constructed using only pure individuals identified by HI and reference samples of M. edulis, M. trossulus and M. galloprovincialis, based on the F ST distance from allele frequencies of the SNP loci. The numbers at the nodes represent bootstrap values estimated from 1000 replications. e, individuals with “pure” M. edulis genotype (HI near 0); t, individuals with “pure” M. trossulus genotype (HI near 1)
Genetic diversity and Hardy-Weinberg equilibrium
The highest percentage of polymorphic SNP (35–40) was identified in Greenland populations compared to the reference samples of Mytilus. The most heterogeneous of the Greenlandic samples was the GLL population (Table 3). H O for 54 loci among all populations was in general lower than H E. The greatest difference can be seen in the Greenland populations, especially in GLL where H O is two times lower than H E. Most loci were in HWE in the reference samples from both sides of the Atlantic, while many loci were not in HWE in Greenland populations. The biggest fraction of SNPs that were not in HWE (p < 0.01) was observed in the outer fjord population GLL (43 %). Twenty-three loci showed significant departures from HWE after a Benjamini-Yekutieli correction. BM115B, BM147A (unknown location), BM54A (ETC complex) and BM17B, BM44B, BM67C (ribosomal proteins) were not in HWE in the three Greenland populations.
The positive values of the within-population fixation index F IS (averaged over all polymorphic loci in each population) indicate the most significant excess of homozygotes (deficiency of heterozygotes) in three populations from Greenland and in a Newfoundland sample of M. trossulus (Table 3).
In the Greenland populations, 16 % (GLD, GLS) to 37 % (GLL) of the loci had very large (>0.6) F IS values (Online Resource 2). Most of these loci were nearly homozygous for one allele in a population, with the absence of any heterozygotes or a single heterozygote only, and a few individuals homozygous for the alternative allele. In a few cases, F IS had values of 1, indicating the existence of two alternative homozygotes without any heterozygotes (e.g., BM17B, BM203C and BM44B). ‘Pure’ M. edulis populations were mostly homozygous for this alternative allele, and the M. trossulus sample from Scotland and Newfoundland was predominately homozygous for the second allele.
Genetic differentiation among populations
The neighbor-joining tree (Fig. 4) revealed the three main groups representing M. edulis, M. trossulus and M. galloprovincialis and the Greenland samples, which contained mixed Mytilus populations that are located between the M. trossulus and a group of M. edulis and M. galloprovincialis. Further, it can be observed that the populations from the inner fjord (GLD and GLS) are grouped more with M. trossulus samples than the more open water Greenland (GLL) population.

Neighbor-joining tree of three Mytilus spp. populations from Greenland and reference samples of M. edulis, M. trossulus and M. galloprovincialis from North America and Europe based on the F ST distance from allele frequencies of the SNP loci. The numbers at the nodes represent bootstrap values estimated from 1000 replications. The population abbreviations and descriptions are given in Table 1
The three Greenlandic sites were found to differ in the taxa distribution with a significantly higher proportion of M. trossulus (p < 0.05, 2 × 2 contingency table analysis) in the inner fjord (GLD and GLS). M. edulis and M. trossulus numbers of specimens at both stations were respectively 7 and 16, 7 and 13 compared to the outer fjord (GLL) with 16 M. edulis and 8 M. trossulus. The average genetic level of differentiation among all studied populations was very high (F ST = 0.426) (Table 4). F ST values were significantly greater than zero between most of the pairs of samples (reaching values as high as 0.787 for M. trossulus and M. edulis populations). All this means is that the inner fjord populations had more M. trossulus specimens and consequently fewer M. edulis specimens than the outer (GLL) population.
Population structure
Correspondence analysis was carried out for individuals to characterize the Greenlandic mixed Mytilus populations (Fig. 5). The first two axes accounted for 90 % of the total variation, with axis 1 accounting for 69.43 % showing a clear separation of M. edulis and M. trossulus populations. Individuals from Greenland either grouped with M. edulis or M. trossulus samples or formed a separate group between them. Hardly any individuals were observed between the group of hybrids and ‘pure’ taxa (either M. edulis or M. trossulus).

The first two axes of the correspondence analysis (CA) computed from the SNP data on three populations of M. edulis/M. trossulus from Greenland and reference samples of M. edulis, M. trossulus and M. galloprovincioalis from North America and Europe. Each dot (point) is an individual
Discussion
Characteristics of Greenland Mytilus populations and interpopulation differentiation
This study is the first presenting evidence for existence of M. trossulus in Greenland. Knowledge of the life-history differences between the M. edulis and M. trossulus taxa is needed to understand the dynamics of mussel taxa coexistence and hybrid zones as well as the environmental and genetic factors that can explain not only the occurrence of hybrids but also the mechanisms that keep the species distinct despite hybridization (McDonald et al. 1991; Toro et al. 2004). The two taxa, M. edulis and M. trossulus, hold the same habitat, where individuals are attached to the substrate by byssus threads and filter food particles from the water column. They are almost morphologically indistinguishable, and results of ongoing studies debate whether small shell differences are caused by environmental impacts or genetic differences (e.g., McDonald et al. 1991; Innes and Bates 1999).
The mussels M. trossulus and M. edulis are here shown to coexist and hybridize in a fjord in the Maarmorilik area, West Greenland. This finding is further documented by comparison of F ST values between M. edulis and M. trossulus populations, which revealed that the GLD and GLS samples were found more introgressed than the GLL population. While the GLL (outer fjord site) was mostly composed of M. edulis mussels (53 %), and to a lesser degree M. trossulus (27 %) and hybrids (20 %), the GLS and GLD (inner fjord sites) both had a much higher occurrence of both M. trossulus (about 40 and 50 %, respectively) and hybrids (39 and 30 %) than of M. edulis (both 21 %).
All Greenlandic mussels were sampled in the intertidal zone, where the mussels were collected during low tide from underneath or in crevices in the rocks. While the open water site (GLL) was a true marine site, the GLS and GLD were collected in a fjord that can be described as a stratified estuary during summer. The stratification is caused by melt water from the rivers and glacier runoff of approximately 110 × 106 m3 freshwater, which flows on top of the heavier seawater, creating a stagnant water mass in the bottom of the fiords and thus salinities from 34 down to 26 psu (Møller 1984). Based on findings of M. trossulus in areas of lower salinities, studies relate this to a higher tolerance to lower salinities compared to M. edulis, a theory that is further backed up by experiments documenting a higher tolerance for M. trossulus at early life history stages (e.g., Qiu et al. 2002; Ridgway and Naevdal 2004; Przytarska et al. 2010; Zbawicka et al. 2014). While the two taxa in most studies are suggested to be physiologically differentiated with respect to salinity, one must bear in mind that areas characterized by lower salinity are also often subjected to significant variation in other environmental factors such as pH, temperature, dissolved oxygen and particulate loadings (Gardner and Thompson 2001), and the proportion of M. edulis vs. M. trossulus may not be solely attributed to a single environmental factor such as salinity.
In Greenland, the harsh Arctic climate challenges the intertidal mussels as in winter they may be exposed to extreme cold air temperatures (down to −25 °C) twice daily if not covered by a stable ice foot (Scrosati and Eckersley 2007). The taxa distribution with a tendency to more M. trossulus in the inner fjord may be affected by the fact that the ice breaks up later in the spring in the inner fjord than at open sea, possibly affecting the time set for mussel reproduction and settlement of larvae. While M. edulis has been shown to start their spawning in late spring, M. trossulus generally starts later in the summer (Toro et al. 2002, 2006), paving a possibility for M. trossulus to dominate areas with longer-lasting sea ice. M. trossulus is more robust compared to its congener, M. edulis, not only in relation to salinity, but also temperature (Hayhurst and Rawson 2009). While this study documented occurrences of M. trossulus in Greenland, other studies in other parts of Greenland by Wanamaker et al. (2007), Riginos and Henzler (2008) and Versteegh et al. (2012) failed to document the existence of other Mytilus taxa than M. edulis. Mytilus mussels are spawners with an extended pelagic larval phase lasting up to several months (Bayne 1965), and with the short distance across the David Strait between Greenland and Canada, the introduction could have occurred after the end of the last glacial period approximately 10,000 years ago.
Hybridization
The genetic structure of Greenland populations can be compared with Scotland, Baltic and North American populations where M. trossulus and M. edulis individuals coexist and hybridize (Riginos and Cunningham 2005; Zbawicka et al. 2010, 2012, 2014). However, while European populations are characterized by large numbers of hybrids, including some with intermediate hybrid index values (backcrosses), Riginos and Cunningham (2005) and Toro et al. (2002) described North American populations as having a low frequency of hybrid mussels.
This study found low numbers of hybrids between ‘pure’ species and F1 hybrids, and such backcrossing could only be assigned to one individual from GLL and three from GLS biased toward both M. trossulus and M. edulis. This situation is completely different from that at the Danish strait (Zbawicka et al. 2014) and Scotland (Comesaña et al. 1999; Zbawicka et al. 2010, 2012) where hybridization between ‘pure’ taxa and hybrids is frequent.
The deficiency of heterozygotes observed in Greenland populations was observed also in the Scottish and Norwegian populations of Mytilus spp. (European populations from hybridization regions), where 25–30 % of SNP loci also had very large (>0.6) F IS values compared to the reference population of Mytilus. The possible impact of recent mixing of M. trossulus with M. edulis was discussed by Zbawicka et al. (2012). Structures in hybrid zones have been studied with respect to reproductive barriers, and results have indicated higher conspecific than heterospecific fertilization rates and greater survival of larvae from conspecific than heterospecific crosses (e.g., Miranda et al. 2010; Toro et al. 2004; Klibansky and McCartney 2014). There are also indications of differences in life-history traits, where for example M. edulis have been found to initiate spawning before the time of M. trossulus. Thus, the maintenance of the hybrid zone may at least partly be caused by reproductive fertilization isolating mechanisms in addition to differences in life history traits, limiting the production of hybrids. Unfitness/fitness of hybrids compared to parent taxa is an ongoing discussion, and for the M. edulis-M. trossulus hybrid zones, examples of both decreased fitness and no differences exist in the literature when adult physiological fitness is compared (Gardner and Thompson 2001).
Implications related to monitoring programs
The finding of M. trossulus might have implications not only for the ecological understanding of the Greenlandic ecosystem, but indeed also in the context of biomonitoring of marine impacts. Mussels are commonly used across Greenland (and worldwide) as bioindicators for industries such as mining, where a sample usually consists of approximately 20–30 pooled individuals to represent a specific site (e.g., Riget et al. 1997, 2000; Søndergaard et al. 2014; Bach et al. 2014). As there are indications that individuals of M. trossulus accumulate some elements to a higher extent than M. edulis (Lobel et al. 1990; Blackmore and Wang 2003), the taxa composition in a sample could thus have an impact on elemental monitoring results. As a result of this study, it clearly calls for further research on the distribution of Mytilus taxa across Greenland and studies of differences among them and hybrids in uptake, accumulation and effects of heavy metals among other contaminant stressors. Such research would be relevant for monitoring perspectives within both national and international frameworks.
Conclusion
This study is the first presenting evidence for the existence of M. trossulus in Greenland. The mussels M. trossulus and M. edulis taxa are here shown to coexist and hybridize in a fjord in the Maarmorilik area, West Greenland. The discovered hybrid zone is characterized by particular features that are different from those of the hybrid zones of both taxa found in the North American Atlantic areas. This finding has implications for future monitoring programs related to climate changes and understanding of the evolution of marine benthic communities in Greenland.
References
Bach L, Olsen M, Asmund G (2014) Environmental Monitoring at the Nalunaq Gold Mine, South Greenland, 2013. Aarhus University, DCE—Danish Centre for Environment and Energy, 44 pp. Scientific Report from DCE—Danish Centre for Environment and Energy No. 97. http://dce2.au.dk/pub/SR97.pdf
Bayne BL (1965) Growth and delay of metamorphosis of the larvae of Mytilus edulis (L.). Ophelia 2:1–47
Beaumont AR, Hawkins MP, Doig FL, Davies IM, Snow M (2008) Three species of Mytilus and their hybrids identified in a Scottish Loch: natives, relicts and invaders? J Exp Mar Biol Ecol 367:100–110
Belkhir K, Borsa P, Chikhi L, Raufaste N, Bonhomme F (2003) GENETIX version 4.04, logiciel sous Windows™ pour la genetique des populations. Laboratoire Genome, Populations, Interactions: CNRS UMR 5000, Université de Montpellier II, Montpellier
Bennike O, Wagner B (2013) Holocene range of Mytilus edulis in central East Greenland. Polar Rec 49:291–296
Benzécri JP (1992) Correspondence analysis handbook. In: Balakrishnan N, Schucany WR, Garvey PR (eds) Statistics: a series of textbooks and monographs, vol 125. Marcel Dekker, New York, NY
Bierne N, Borsa P, Daguin C, Jollivet D, Viard F, Bonhomme F, David P (2003) Introgression patterns in the mosaic hybrid zone between Mytilus edulis and M. galloprovincialis. Mol Ecol 12:447–461
Blackmore G, Wang WX (2003) Comparison of metal accumulation in mussels at different local and global scales. Environ Toxicol Chem 22:388–395
Blicher ME, Sejr MK, Hogslund S (2013) Population structure of Mytilus edulis in the intertidal zone in a sub-Arctic fjord, SW Greenland. Mar Ecol Prog Ser 487:89–100
Borsa P, Daguin C, Caetano SR, Bonhomme F (1999) Nuclear-DNA evidence that northeastern Atlantic Mytilus trossulus mussels carry Mytilus edulis genes. J Molluscan Stud 65:504–507
Comesaña AS, Toro JE, Innes DJ, Thompson RJ (1999) A molecular approach to the ecology of a mussel (Mytilus edulis–M. trossulus) hybrid zone on the east coast of Newfoundland, Canada. Mar Biol 133:213–221
Excoffier L, Lischer HEL (2010) Arlequin suite ver 3.5: a new series of programs to perform population genetics analyses under Linux and Windows. Mol Ecol Resour 10:564–567
Gabriel S, Ziaugra L, Tabbaa D (2009) SNP genotyping using the Sequenom MassARRAY iPLEX platform. Curr Protoc Hum Genet 60:12.1.1–12.1.18
Gardner JPA, Thompson RJ (2001) The effects of coastal and estuarine conditions on the physiology and survivorship of the mussels Mytilus edulis, M. trossulus and their hybrids. J Exp Mar Biol Ecol 265:119–140
Gardner JPA, Thompson RJ (2009) Influence of genotype and geography on shell shape and morphometric trait variation among North Atlantic blue mussel (Mytilus spp.) populations. Biol J Linn Soc 96:875–897
Hayhurst S, Rawson PD (2009) Species-specific variation in larval survival and patterns of distribution for the blue mussels Mytilus edulis and Mytilus trossulus in the Gulf of Maine. J Mollusca Stud 75:215–222
Hoarau G, Rijnsdorp AD, Van Der Veer HW, Stam WT, Olsen JL (2002) Population structure of plaice (Pleuronectes platessa L.) in northern Europe: microsatellites revealed large-scale spatial and temporal homogeneity. Mol Ecol 11:1165–1176
Høgslund S, Sejr MK, Wiktor J Jr, Blicher ME, Wegeberg S (2014) Intertidal community composition along rocky shores in Southwest Greenland: a quantitative approach. Polar Biol 37:1549–1561
Innes DJ, Bates JA (1999) Morphological variation of Mytilus edulis and Mytilus trossulus in eastern Newfoundland. Mar Biol 133:691–699
Kijewski T, Smietanka B, Zbawicka M, Gosling E, Hummel H, Wenne R (2011) Distribution of Mytilus taxa in European coastal areas as inferred from molecular markers. J Sea Res 65:224–234
Klibansky LKJ, McCartney MA (2014) Conspecific sperm precedence is a reproductive barrier between free-spawning marine mussels in the Northwest Atlantic Mytilus Hybrid Zone. PLoS One 9:e108433
Lobel PB, Belkhode SP, Jackson SE, Longerich HP (1990) Recent taxonomic discoveries concerning the mussel Mytilus: implications for biomonitoring. Arch Environ Contam Toxicol 19:508–512
McDonald JH, Seed R, Koehn RK (1991) Allozymes and morphometric characters of three species of Mytilus in the Northern and Southern Hemispheres. Mar Biol 111:323–333
Miranda MBB, Innes DJ, Thompson RJ (2010) Incomplete reproductive isolation in the blue mussel (Mytilus edulis and M. trossulus) hybrid zone in the Northwest Atlantic: role of gamete interactions and larval viability. Biol Bull 218:266–281
Møller JS (1984) Hydrodynamics of an Arctic Fiord. Thesis: Institute of Hydrodynamics and Hydraulic Engineering (ISVA). Technical University of Denmark, Lyngby, p. 197
Moreau V, Tremblay R, Bourget E (2005) Distribution of Mytilus edulis and M. trossulus on the Gaspe coast in relation to spatial scale. J Shellfish Res 24:545–551
Narum SR (2006) Beyond Bonferroni: less conservative analyses for conservation genetics. Conserv Genet 7:783–787
Przytarska J, Sokołowski A, Wołowicz M, Hummel H, Jansen J (2010) Comparison of trace metal bioavailabilities in European coastal waters using mussels from Mytilus edulis complex as biomonitors. Environ Monit Assess 166:461–476
Qiu JW, Tremblay R, Bourget E (2002) Ontogenetic changes in hyposaline tolerance in the mussels Mytilus edulis and M. trossulus: implications for distribution. Mar Ecol Prog Ser 228:143–152
Ridgway G, Naevdal GN (2004) Genotypes of Mytilus from waters of different salinity around Bergen, Norway. Helgol Mar Res 58:104–109
Riget F, Johansen P, Asmund G (1997) Uptake and release of lead and zinc by blue mussels: experience from transplantation experiments in Greenland. Mar Pollut Bull 34:805–815
Riget F, Dietz R, Johansen P, Asmund G (2000) Lead, cadmium, mercury and selenium in Greenland marine biota and sediments during AMAP phase 1. Sci Total Environ 245:3–14
Riginos C, Cunningham CW (2005) Local adaptation and species segregation in two mussel (Mytilus edulis × Mytilus trossulus) hybrid zones. Mol Ecol 14:381–400
Riginos C, Henzler C (2008) Patterns of mtDNA diversity in North Atlantic populations of the mussel Mytilus edulis. Mar Biol 155:399–412
Rousset F (2008) GENEPOP’007: a complete re-implementation of the GENEPOP software for Windows and Linux. Mol Ecol Resour 8:103–106
Scrosati R, Eckersley LK (2007) Thermal insulation of the intertidal zone by the ice foot. J Sea Res 58:331–334
Shields J, Barnes P, Heath D (2008) Growth and survival differences among native, introduced and hybrid blue mussels (Mytilus spp.): genotype, environment and interaction effects. Mar Biol 154:919–928
Smietanka B, Burzynski A, Wenne R (2009) Molecular population genetics of male and female mitochondrial genomes in European mussels Mytilus. Mar Biol 156:913–925
Smietanka B, Zbawicka M, Sanko T, Wenne R, Burzynski A (2013) Molecular population genetics of male and female mitochondrial genomes in subarctic Mytilus trossulus. Mar Biol 160:1709–1721
Smietanka B, Burzynski A, Hummel H, Wenne R (2014) Glacial history of the European marine mussels Mytilus, inferred from distribution of mitochondrial DNA lineages. Heredity 113:250–258
Søndergaard J (2013) Dispersion and bioaccumulation of elements from an open-pit olivine mine in Southwest Greenland assessed using lichens, seaweeds, mussels and fish. Environ Monit Assess 185:7025–7035
Søndergaard J, Bach L, Gustavson K (2014) Measuring bioavailable metals using diffusive gradients in thin films (DGT) and transplanted seaweed (Fucus vesiculosus), blue mussels (Mytilus edulis) and sea snails (Littorina saxatilis) suspended from monitoring buoys near a former lead-zinc mine in West Greenland. Mar Pollut Bull 78:102–109
Staden R, Judge DP, Bonfield JK (2001) Sequence assembly and finishing methods. In: Baxevanis AD, Ouellette BFF (eds) Bioinformatics: a practical guide to the analysis of genes and proteins, 2nd edn. Wiley, New York, pp 303–322
Strand J, Asmund G (2003) Tributyltin accumulation and effects in marine molluscs from West Greenland. Environ Pollut 123:31–37
Stuckas H, Stoof K, Quesada H, Tiedemann R (2009) Evolutionary implications of discordant clines across the Baltic Mytilus hybrid zone (Mytilus edulis and Mytilus trossulus). Heredity 103:146–156
Takezaki N, Nei M, Tamura K (2010) POPTREE2: software for constructing population trees from allele frequency data and computing other population statistics with windows interface. Mol Biol Evol 27:747–752
Tamura K, Dudley J, Nei M, Kumar S (2007) MEGA4: molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol Biol Evol 24:1596–1599
Theisen BF (1973) The growth of Mytilus edulis L. (bivalva) from Disko and Thule district,Greenland. Ophelia 12:59–77
Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins DG (1997) The CLUSTAL_X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res 25:4876–4882
Thyrring J, Rysgaard S, Blicher ME, Sejr MK (2015) Metabolic cold adaptation and aerobic performance of blue mussels (Mytilus edulis) along a temperature gradient into the High Arctic region. Mar Biol 162:235–243
Toro JE, Thompson RJ, Innes DJ (2002) Reproductive isolation and reproductive output in two sympatric mussel species (Mytilus edulis, M. trossulus) and their hybrids from Newfoundland. Mar Biol 141:897–909
Toro J, Innes DJ, Thompson RJ (2004) Genetic variation among life-history stages of mussels in a Mytilus edulis–M. trossulus hybrid zone. Mar Biol 145:713–725
Toro JE, Thompson RJ, Innes DJ (2006) Fertilization success and early survival in pure and hybrid larvae of Mytilus edulis (Linnaeus, 1758) and M. trossulus (Gould, 1850) from laboratory crosses. Aquac Res 37:1703–1708
Väinölä R, Hvilsom MM (1991) Genetic-divergence and a hybrid zone between Baltic and North-Sea Mytilus populations (mytilidae, mollusca). Biol J Linn Soc 43:127–148
Väinölä R, Strelkov P (2011) Mytilus trossulus in Northern Europe. Mar Biol 158:817–833
Vermeij GJ (1992) Trans-equatorial connections between biotas in the temperate Eastern Atlantic. Mar Biol 112:343–348
Versteegh EAA, Blicher ME, Mortensen J, Rysgaard S, Als TD, Wanamaker AD (2012) Oxygen isotope ratios in the shell of Mytilus edulis: archives of glacier meltwater in Greenland? Biogeosciences 9:5231–5241
Wanamaker AD, Kreutz KJ, Borns HW, Introne DS, Feindel S et al (2007) Experimental determination of salinity, temperature, growth, and metabolic effects on shell isotope chemistry of Mytilus edulis collected from Maine and Greenland. Paleoceanography 22:PA2217
Zbawicka M, Burzynski A, Skibinski D, Wenne R (2010) Scottish Mytilus trossulus mussels retain ancestral mitochondrial DNA: complete sequences of male and female mtDNA genomes. Gene 456:45–53
Zbawicka M, Drywa A, Smietanka B, Wenne R (2012) Identification and validation of novel SNP markers in European populations of marine Mytilus mussels. Mar Biol 159:1347–1362
Zbawicka M, Sanko T, Strand J, Wenne R (2014) New SNP markers reveal largely concordant clinal variation across the hybrid zone between Mytilus spp. in the Baltic Sea. Aquat Biol 21:25–36
Acknowledgments
This study was partially funded by the 2011/01/B/NZ9/04352 NCN project for R.W., the Leading National Research Centre (KNOW)—the Centre for Polar Studies for the period 2014–2018 and statutory topic IV in the IOPAS.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Wenne, R., Bach, L., Zbawicka, M. et al. A first report on coexistence and hybridization of Mytilus trossulus and M. edulis mussels in Greenland. Polar Biol 39, 343–355 (2016). https://doi.org/10.1007/s00300-015-1785-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-015-1785-x