
Abstract
Aims/hypothesis
A meta-analysis was performed to assess the association between the UCP2 −866G/A, UCP2 Ala55Val and UCP3 −55C/T polymorphisms and type 2 diabetes susceptibility.
Methods
A literature-based search was conducted to identify all relevant studies. The fixed or random effect pooled measure was calculated mainly at the allele level to determine heterogeneity bias among studies. Further analyses were performed that stratified for ethnicity.
Results
We examined 17 publications. Stratified analysis for ethnicity and sensitivity analysis revealed that there was no heterogeneity between studies for these variants. Using an additive model, no significant association of the UCP2 −866G/A polymorphism with type 2 diabetes risk was observed, either in participants of Asian (OR 1.05, 95% CI 0.96, 1.16) or of European (OR 1.03, 95% CI 0.99, 1.07) descent. Neither the UCP2 Ala55Val nor the UCP3 −55C/T polymorphism showed any significant association with type 2 diabetes risk in Europeans (OR 1.04, 95% CI 0.98, 1.09 for Ala55Val; OR 1.04, 95% CI 1.00, 1.09 for −55C/T). In contrast, a statistically significant association was observed for both polymorphisms in participants of Asian descent (OR 1.23, 95% CI 1.12, 1.36 for Ala55Val; OR 1.15, 95% CI 1.03, 1.28 for −55C/T).
Conclusions/interpretation
Our meta-analysis suggests that the UCP2 −866G/A polymorphism is unlikely to be associated with increased type 2 diabetes risk in the populations investigated. In contrast, our results indicate that the UCP2 Ala55Val and UCP3 −55C/T polymorphisms may indeed be risk factors for susceptibility to type 2 diabetes in individuals of Asian descent, but not in individuals of European descent. This conclusion warrants confirmation by further studies.
Similar content being viewed by others


Introduction
Uncoupling proteins (UCPs), a family of mitochondrial transporter proteins, uncouple the transport of protons across the inner mitochondrial membrane from electron transport and the synthesis of ATP from ADP [1]. Among the five UCP homologues (UCP1 to UCP5), UCP2 and UCP3 are located adjacent to one another on human chromosome 11q13 [2, 3] and are 73% identical to each other at the amino acid sequence level [4]. Studies indicate that UCP2, as a key component of the beta cell glucose-sensing mechanism that regulates glucose-stimulated insulin secretion [5–7], is a critical link between beta cell dysfunction and type 2 diabetes [8]. It has also been observed that lower UCP3 mRNA levels are present in the skeletal muscle of type 2 diabetes patients [9].
A number of studies have examined the association between genetic variability in the UCP2–UCP3 gene cluster and the risk of type 2 diabetes, with most studies focusing on three common single nucleotide polymorphisms (SNPs) [10]. These are a SNP located in a multifunctional cis-regulatory site of the UCP2 promoter region (−866G/A, rs659366), a missense variant in exon 4 of UCP2 (Ala55Val, rs660339) and a SNP 6 bp upstream from the TATA box in the core promoter region of UCP3 (−55C/T, rs1800849). The −866G/A polymorphism, which acts as a binding site for the pancreatic transcription factors Paired box-containing 6 and Insulin promoter factor 1 [11, 12], has been associated with higher UCP2 mRNA levels, reduced insulin secretion and increased type 2 diabetes risk [13–15]. The Ala55Val polymorphism has been associated with a lower degree of uncoupling, lower energy expenditure [16] and a higher risk of obesity, as well as a higher incidence of diabetes [17, 18]. Similarly, the T allele of the UCP3 −55C/T polymorphism has been associated with a reduced risk of type 2 diabetes and higher plasma total cholesterol and LDL-cholesterol [19].
Despite strong functional evidence for the involvement of these three SNPs in the regulation of uncoupling, the results of the genetic association studies on association with type 2 diabetes remain inconclusive. To further examine the potential role of these three SNPs in influencing type 2 diabetes susceptibility, we performed a meta-analysis on eligible case–control studies. Our aim was to estimate the effect of these SNPs in populations of Asian and European descent. Our results suggest that the UCP2 Ala55Val and UCP3 −55C/T polymorphisms may have a selective effect on the development of type 2 diabetes in individuals of Asian descent.
Methods
Search strategy
PubMed and Embase were searched systematically to identify all available relevant articles. The most-studied SNPs (UCP2 −866G/A, UCP2 Ala55Val and UCP3 −55C/T) were investigated using combinations of the following search terms: ‘diabetes and UCP2’, ‘UCP3’, ‘uncoupling protein 2’, ‘uncoupling protein 3’, ‘variant’, and ‘polymorphism’. The search was limited to English language papers and completed on June 10, 2011. We also used the PubMed option ‘Related Articles’ for each research article to retrieve additional potentially relevant articles. All of the included articles were also hand-searched to identify any other relevant citations. No restriction was set on the source of control participants (general population, clinic or hospital).
Inclusion and exclusion criteria
To determine whether an individual study was eligible for inclusion in the meta-analysis, all of the studies identified were carefully reviewed by two investigators working independently, any discrepancies being resolved by discussion and, when necessary, adjudicated by a third reviewer. The inclusion and exclusion criteria were as follows: First, each case–control study had to have been published as an original study designed to evaluate the association. Second, numbers in case and control groups had to be reported for each allele or genotype. Third, case–control studies had to have sufficient published data to estimate an OR with 95% CI or to provide raw data that allowed us to calculate them. Fourth, if the data were duplicated and had been published more than once, the most recent and complete study was chosen. Fifth, studies were excluded if the genotype distribution of the controls deviated from Hardy–Weinberg equilibrium (HWE). Sixth, the following were excluded: animal studies, review articles, abstracts, editorials, reports with incomplete data, studies based on pedigree data, studies on other type of diabetes (type 1 diabetes, gestational diabetes, etc.) and prospective studies.
Data extraction
Data were independently extracted by two investigators who reached a consensus on all of the items. Information extracted from each study was considered as follows: name of first author, publication year, ethnic origin of the population studied, number of participants in case and control groups, genotype and allele frequency by case/control status, and OR (95% CI). Not all papers reported the necessary statistics directly, so in some instances we transformed and estimated an OR from the reported data [20]. We did not define a minimum number of patients for a study to be included in our meta-analysis.
Statistical analysis
HWE of the genotype distribution of controls was tested by a goodness-of-fit χ 2 analysis. The distribution was considered to have deviated from HWE at p < 0.05. Pooled ORs with 95% CI were used to assess the strength of association in the additive, dominant and recessive models, respectively. Pooled estimates of the OR were obtained by calculating a weighted average of ORs from each study, with the statistical significance of the pooled OR being determined by the Z test.
To examine the possibility of heterogeneity across the studies, a statistical test for heterogeneity was performed. This was based on the χ 2-based Q statistic and I 2 metric, and quantifies between-study heterogeneity irrespective of the number of studies. Heterogeneity was considered significant at p < 0.05 for the Q statistic and I 2 > 50% for the I 2 metric. In the presence of substantial heterogeneity, the DerSimonian and Laird random effect model (REM) was adopted as the pooling method; otherwise the fixed effect model (FEM) was used [21, 22]. Meta-regression and sensitivity analysis were conducted to evaluate the key studies with a substantial impact on between-study heterogeneity. Influence analysis was performed to assess the stability of the results, with a single study in the meta-analysis being deleted each time to reflect the influence of the individual data set on the pooled OR.
The statistical power for each of the three SNPs was calculated by power and sample size software [23], and the false-positive report probability (FPRP) test of Wacholder et al. [24] was applied to address the issue of false-positive SNP associations. All genetic variants were analysed using the Begg and Egger tests for potential publication bias [25]. The significance of the intercept was determined by the t test suggested by Egger, with p < 0.10 considered representative of statistically significant publication bias. All statistical analyses were conducted using STATA version 11.0 (Stata, College Station, TX, USA).
Results
Characteristics of study
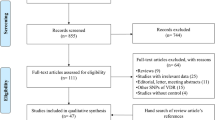
The trial flow is summarised in Fig. 1 of the electronic supplementary material (ESM). A total of 17 published articles [11, 13, 14, 19, 26–38] with 28 outcomes met the inclusion and exclusion criteria. All were case–control studies and most were population-based. The allele and genotype distributions in the studies included are summarised in Tables 1, 2, and 3 for the UCP2 −866G/A, UCP2 Ala55Val and UCP3 −55C/T polymorphisms respectively. The association of the UCP2 −866G/A polymorphism, the UCP2 Ala55Val polymorphism and the UCP3 −55C/T polymorphism with type 2 diabetes risk was examined in 13, 7 and 8 studies respectively. Other characteristics (sex, age, etc.) are summarised in ESM Table 1.
Quantitative synthesis
Results of pooled analyses are summarised in detail in Table 4. Our meta-analysis showed no significant association between the UCP2 −866G/A polymorphism and risk of type 2 diabetes, either by additive (REM OR 1.03, 95% CI 0.95, 1.11), dominant (FEM OR 1.03, 95% CI 0.90, 1.18) or recessive (REM OR 1.00, 95% CI 0.84, 1.18) models. Moreover, no significant association was observed when an additive model was used after stratification for ethnicity (Asian descent FEM OR 1.05, 95% CI 0.96, 1.16; European descent REM OR 1.04, 95% CI 0.92, 1.17) (Fig. 1).

Stratified analysis pooled ORs for the association between the UCP2 −866G/A polymorphism and susceptibility to type 2 diabetes by ethnicity. The area of the squares reflects the study-specific weight. The diamond shows the summary fixed-effects OR estimate from 12 studies
For the UCP2 Ala55Val polymorphism, the C allele was found to be significantly associated with an increased risk of type 2 diabetes when using a recessive model (FEM OR 1.39, 95% CI 1.16, 1.66), but not when using additive (REM OR 1.11, 95% CI 0.98, 1.26) or dominant (REM OR 1.38, 95% CI 0.80, 2.37) models. However, after stratification by ethnicity, a significant association was revealed by an additive model in populations of Asian descent (FEM OR 1.23, 95% CI 1.12, 1.36), but not in those of European descent (FEM OR 1.04, 95% CI 0.98, 1.09) (Table 4 and Fig. 2).

Stratified analysis pooled ORs for the association between the UCP2 Ala55Val polymorphism and susceptibility to type 2 diabetes mellitus by ethnicity. The area of the squares reflects the study-specific weight. The diamond shows the summary fixed-effects OR estimate from seven studies
Our meta-analysis also showed a significant overall association between the UCP3 −55C/T polymorphism and increased risk of type 2 diabetes in all models (additive FEM OR 1.05, 95% CI 1.01, 1.10; dominant FEM OR 1.33, 95% CI 1.02, 1.73; recessive FEM OR 1.19, 95% CI 1.04, 1.36). Surprisingly, when stratified by ethnicity, the significant association between the UCP3 −55C/T polymorphism and risk of type 2 diabetes was most evident in individuals of Asian descent (FEM OR 1.15, 95% CI 1.03, 1.28), with only a marginal significance observed in persons of European descent (FEM OR 1.04, 95% CI 1.00, 1.09) (Table 4 and Fig. 3).

Stratified analysis pooled ORs for the association between the UCP3 −55C/T polymorphism and susceptibility to type 2 diabetes mellitus by ethnicity. The area of the squares reflects the study-specific weight. The diamond shows the summary fixed-effects OR estimate from eight studies
Heterogeneity and sensitivity analyses
As shown in Table 4, significant heterogeneity was observed among studies of the UCP2 −866G/A and Ala55Val polymorphisms in the overall populations, but no heterogeneity was found in the inherited models for the UCP3 −55C/T polymorphism. To investigate this further, the following covariates were considered: publication year, sex (ratio of males in cases to that in controls), age (ratio of the mean age in cases to that in controls) and sample size. However, univariate meta-regression analysis showed that none of the tested covariates could by themselves explain the observed between-study heterogeneity. To identify the studies with the greatest impact on the overall between-study heterogeneity, sensitivity analyses were conducted in the overall population. The results indicated that two studies [11, 35] were mainly responsible for the observed heterogeneity. Moreover, when the data were stratified by ethnicity and an additive model used, the heterogeneity between the studies of the UCP2 Ala55Val polymorphism was significantly decreased or eliminated in populations of Asian and European descent (Table 4). Similarly, the heterogeneity was also effectively removed from the studies of the UCP2 −866G/A polymorphism in participants of Asian descent, but still existed in studies investigating individuals of European descent.
Influence analysis
To assess the degree to which each individual study affected the overall OR estimates, influence analysis was conducted by repeating the meta-analysis sequentially excluding one study at a time. As shown in Table 4, only one study [35] was found to have an excessive influence on the pooled effect. This was limited to analysis of the UCP2 Ala55Val polymorphism in the overall population using an additive model (FEM OR 1.05, 95% CI 1.00, 1.10). Otherwise no single study excessively influenced the analyses.
Publication bias
As expected, no significant publication bias was detected in the inherited models for any of the polymorphisms examined (ESM Table 2), confirming that our results are statistically robust.
Discussion
Results from several genome-wide association studies (GWAS) in a variety of populations have identified 37 replicating type 2 diabetes susceptibility loci [34, 39–44]. However, the biological pictures revealed by GWAS remain incomplete. Thus, many of the associations identified by GWAS do not involve previously identified type 2 diabetes candidate genes, and many of the associated markers are in genomic locations containing genes whose function is currently unknown. Recently, several studies suggested an association between the UCP2 −866G/A, UCP2 Ala55Val and UCP3 −55C/T polymorphisms and type 2 diabetes risk. Despite strong functional evidence for the relevance of these three SNPs, the results for association with type 2 diabetes show significant between-study variation. To obtain a more definitive conclusion, we conducted a meta-analysis of 17 published articles with 28 outcomes from populations of different ethnic origins [11, 13, 14, 19, 26–38]. We believe such a meta-analysis has a much greater possibility of reaching reasonably strong conclusions.
Heterogeneity is potentially a significant problem when interpreting the results of any meta-analysis of genetic association studies [45]. Our meta-analysis also showed significant between-study heterogeneity in most of the models that we used to examine the associations of the UCP2 −866G/A and Ala55Val polymorphisms. Many of the variables that varied between the various studies might be responsible for this observed heterogeneity, including the source of the controls, sex bias, ethnicity, etc. Initial inspection of the data did not immediately identify any likely candidate variable or study that was significantly impacting on our overall results. Thus, to explore this matter further, meta-regression and ‘leave one out’ sensitivity analyses were performed [46], revealing that ethnicity was the only covariate likely to have made an important contribution to the overall between-study heterogeneity. The reason for this is unclear, but it may be that populations of different ethnicity also have environmental differences that affect their sensitivity to particular genomic variants. Similarly, based on sensitivity analyses using I 2 > 50% as the cut-off criteria, two studies [11, 35] were identified as the principal outliers in our analyses.
The study by Voight et al. [34], which is a Diabetes Genetics Replication and Meta-analysis (DIAGRAM) study and includes eight GWAS on type 2 diabetes, also met our inclusion criteria. After confirming by sensitivity analysis that it would not contribute to overall heterogeneity, we combined this with the other studies included. This additional analysis indicated that the UCP2 Ala55Val and UCP3 −55C/T polymorphisms, but not the UCP2 −866G/A polymorphism were significantly associated with type 2 diabetes risk in the overall population. As heterogeneity still existed and the DIAGRAM study was from populations of European descent, we again stratified our analysis by ethnicity. The results indicated that no obvious heterogeneities among the stratified studies existed and that the UCP2 −866G/A, UCP2 Ala55Val and UCP3 −55C/T polymorphisms had no significant association with type 2 diabetes risk in populations of European descent, a finding consistent with the conclusions of the DIAGRAM study. Interestingly, the results from the studies examining populations of Asian descent conflicted with this conclusion and indicated that the association with type 2 diabetes was statistically significant for the UCP2 Ala55Val and UCP3 polymorphisms, but not for the UCP2 −866G/A polymorphism. Although our analysis of Asian populations had a relatively small sample size, we nevertheless had 80% power at a 0.05 significance level to detect an OR of 1.5 or greater (statistical power 0.996 and 0.793 for UCP2 Ala55Val and UCP3 −55C/T respectively). The FPRP value for the UCP3 −55C/T polymorphism suggested a <20% chance of the result being a false positive when assigned a relatively high prior probability range (i.e. 0.01–0.1) (data not shown). In contrast, the FPRP value for the UCP2 Ala55Val polymorphism remained below 0.2 even for a prior probability of 0.001, suggesting that the FPRP value is quite robust and that UCP2 may contain one or more genetic variants that increase type 2 diabetes risk in individuals of Asian descent.
The results of the present meta-analysis should also be interpreted within the context of its limitations. Thus previous studies have also indicated that the UCP2 −866G/A, UCP2 Ala55Val and UCP3 −55C/T polymorphisms are associated with obesity [10], and that the UCP2 −866G/A and Ala55Val polymorphisms are associated with proliferative diabetic retinopathy in type 2 diabetes patients or decreased risk of coronary artery disease in men with type 2 diabetes [47, 48]. However, the number of studies providing this clinical information was too low for us to take these covariates into account by meta-regression. Similarly, besides ethnicity, other potential environment × gene interactions may well be contributors to the observed disease-effect unconformity, but we had insufficient data to perform an evaluation of such interactions. Furthermore, one single study found that overweight white women with a potential high-risk haplotype (in high linkage disequilibrium with 866A- and 55T- alleles) had a 3.8-fold increased type 2 diabetes risk [30]. We again had insufficient data to confirm this association, but on the basis of our meta-analysis, we propose that this may be due to some other, as yet unidentified variants also contained within this diabetes-associated haplotype.
In conclusion, our results indicate that the UCP2 −866G/A polymorphism is not a candidate for susceptibility to type 2 diabetes in any ethnic population. However, our results do support the hypothesis that the UCP2 Ala55Val and UCP3 −55C/T polymorphisms are type 2 diabetes susceptibility loci in populations of Asian, but not European descent. We suggest that additional larger studies allowing stratification for other gene × environment interactions should be performed to further clarify the possible roles of the three UCP2 and UCP3 genetic variants in the aetiology of type 2 diabetes.
Abbreviations
- DIAGRAM:
-
Diabetes Genetics Replication and Meta-analysis
- FEM:
-
Fixed effect model
- FPRP:
-
False-positive report probability
- GWAS:
-
Genome-wide association study/studies
- HWE:
-
Hardy–Weinberg equilibrium
- REM:
-
Random effect model
- SNP:
-
Single nucleotide polymorphism
- UCP:
-
Uncoupling protein
References
Dalgaard LT, Pedersen O (2001) Uncoupling proteins: functional characteristics and role in the pathogenesis of obesity and type II diabetes. Diabetologia 44:946–965
Fleury C, Neverova M, Collins S et al (1997) Uncoupling protein-2: a novel gene linked to obesity and hyperinsulinemia. Nat Genet 15:269–272
Solanes G, Vidal-Puig A, Grujic D, Flier JS, Lowell BB (1997) The human uncoupling protein-3 gene. Genomic structure, chromosomal localization, and genetic basis for short and long form transcripts. J Biol Chem 272:25433–25436
Krauss S, Zhang CY, Lowell BB (2005) The mitochondrial uncoupling-protein homologues. Nat Rev Mol Cell Biol 6:248–261
Chan CB, Kashemsant N (2006) Regulation of insulin secretion by uncoupling protein. Biochem Soc Trans 34:802–805
de Souza CT, Araujo EP, Stoppiglia LF et al (2007) Inhibition of UCP2 expression reverses diet-induced diabetes mellitus by effects on both insulin secretion and action. FASEB J 21:1153–1163
Chan CB, DeLeo D, Joseph JW et al (2001) Increased uncoupling protein-2 levels in beta-cells are associated with impaired glucose-stimulated insulin secretion: mechanism of action. Diabetes 50:1302–1310
Zhang CY, Baffy G, Perret P et al (2001) Uncoupling protein-2 negatively regulates insulin secretion and is a major link between obesity, beta cell dysfunction, and type 2 diabetes. Cell 105:745–755
Krook A, Digby J, O’Rahilly S, Zierath JR, Wallberg-Henriksson H (1998) Uncoupling protein 3 is reduced in skeletal muscle of NIDDM patients. Diabetes 47:1528–1531
Jia JJ, Zhang X, Ge CR, Jois M (2009) The polymorphisms of UCP2 and UCP3 genes associated with fat metabolism, obesity and diabetes. Obes Rev 10:519–552
Krempler F, Esterbauer H, Weitgasser R et al (2002) A functional polymorphism in the promoter of UCP2 enhances obesity risk but reduces type 2 diabetes risk in obese middle-aged humans. Diabetes 51:3331–3335
Esterbauer H, Schneitler C, Oberkofier H et al (2001) A common polymorphism in the promoter of UCP2 is associated with decreased risk of obesity in middle-aged humans. Nat Genet 28:178–183
D’Adamo M, Perego L, Cardellini M et al (2004) The −866A/A genotype in the promoter of the human uncoupling protein 2 gene is associated with insulin resistance and increased risk of type 2 diabetes. Diabetes 53:1905–1910
Sasahara M, Nishi M, Kawashima H et al (2004) Uncoupling protein 2 promoter polymorphism −866G/A affects its expression in beta-cells and modulates clinical profiles of Japanese type 2 diabetic patients. Diabetes 53:482–485
Sesti G, Cardellini M, Marini MA et al (2003) A common polymorphism in the promoter of UCP2 contributes to the variation in insulin secretion in glucose-tolerant subjects. Diabetes 52:1280–1283
Astrup A, Toubro S, Dalgaard LT, Urhammer SA, Sorensen TI, Pedersen O (1999) Impact of the v/v 55 polymorphism of the uncoupling protein 2 gene on 24-h energy expenditure and substrate oxidation. Int J Obes Relat Metab Disord 23:1030–1034
Yu X, Jacobs DR, Schreiner PJ, Gross MD, Steffes MW, Fornage M (2005) The uncoupling protein 2 Ala55Val polymorphism is associated with diabetes mellitus: the CARDIA Study. Clin Chem 51:1451–1456
Walder K, Norman RA, Hanson RL et al (1998) Association between uncoupling protein polymorphisms (UCP2-UCP3) and energy metabolism/obesity in Pima Indians. Hum Mol Genet 7:1431–1435
Meirhaeghe A, Amouyel P, Helbecque N et al (2000) An uncoupling protein 3 gene polymorphism associated with a lower risk of developing type II diabetes and with atherogenic lipid profile in a French cohort. Diabetologia 43:1424–1428
Parmar MKB, Torri V, Stewart L (1998) Extracting summary statistics to perform meta-analyses of the published literature for survival endpoints. Stat Med 17:2815–2834
Higgins JP, Thompson SG (2002) Quantifying heterogeneity in a meta-analysis. Stat Med 21:1539–1558
Higgins JP, Thompson SG, Deeks JJ, Altman DG (2003) Measuring inconsistency in meta analyses. BMJ 327:557–560
Dupont WD, Plummer WD Jr (1990) Power and sample size calculations. A review and computer program. Control Clin Trials 11:116–128
Wacholder S, Chanock S, Garcia-Closas M, ElGhormli L, Rothman N (2004) Assessing the probability that a positive report is false: an approach for molecular epidemiology studies. J Natl Cancer Inst 96:434–442
Harbord RM, Egger M, Sterne JA (2006) A modified test for small-study effects in meta-analyses of controlled trials with binary endpoints. Stat Med 25:3443–3457
Wang H, Chu WS, Lu T, Hasstedt SJ, Kern PA, Elbein SC (2004) Uncoupling protein-2 polymorphisms in type 2 diabetes, obesity, and insulin secretion. Am J Physiol Endocrinol Metab 286:E1–E7
Ji Q, Ikegami H, Fujisawa T et al (2004) A common polymorphism of uncoupling protein 2 gene is associated with hypertension. J Hypertens 22:97–102
Bulotta A, Ludovico O, Coco A et al (2005) The common −866G/A polymorphism in the promoter region of the UCP-2 gene is associated with reduced risk of type 2 diabetes in Caucasians from Italy. J Clin Endocrinol Metab 90:1176–1180
Pinelli M, Giacchetti M, Acquaviva F et al (2006) Beta2-adrenergic receptor and UCP3 variants modulate the relationship between age and type 2 diabetes mellitus. BMC Med Genet 7:85
Hsu YH, Niu T, Song Y, Tinker L, Kuller LH, Liu S (2008) Genetic variants in the UCP2-UCP3 gene cluster and risk of diabetes in the women’s health initiative observational study. Diabetes 57:1101–1107
Lee HJ, Ryu HJ, Shin HD et al (2008) Associations between polymorphisms in the mitochondrial uncoupling proteins (UCPs) with T2DM. Clin Chim Acta 398:27–33
Beitelshees AL, Finck BN, Leone TC et al (2010) Interaction between the UCP2 −866G>A polymorphism, diabetes, and beta-blocker use among patients with acute coronary syndromes. Pharmacogenet Genomics 20:231–238
Heidari J, Akrami SM, Heshmat R, Amiri P, Fakhrzadeh H, Pajouhi M (2010) Association study of the −866G/A UCP2 gene promoter polymorphism with type 2 diabetes and obesity in a Tehran population: a case control study. Arch Iran Med 13:384–390
Voight BF, Scott LJ, Steinthorsdottir V et al (2010) Twelve type 2 diabetes susceptibility loci identified through large-scale association analysis. Nat Genet 42:579–589
Vimaleswaran KS, Radha V, Ghosh S, Majumder PP, Sathyanarayana Rao MR, Mohan V (2011) Uncoupling protein 2 and 3 gene polymorphisms and their association with type 2 diabetes in Asian Indians. Diabetes Technol Ther 13:19–25
Kubota T, Mori H, Tamori Y et al (1998) Molecular screening of uncoupling protein 2 gene in patients with noninsulin-dependent diabetes mellitus or obesity. J Clin Endocrinol Metab 83:2800–2804
Cho YM, Ritchie MD, Moore JH et al (2004) Multifactor-dimensionality reduction shows a two-locus interaction associated with type 2 diabetes mellitus. Diabetologia 47:549–554
Dalgaard LT, Hansen T, Urhammer SA, Drivsholm T, Borch-Johnsen K, Pedersen O (2001) The uncoupling protein 3–55 C>T variant is not associated with Type II diabetes mellitus in Danish subjects. Diabetologia 44:1065–1067
Saxena R, Voight BF, Lyssenko V et al (2007) Genome-wide association analysis identifies loci for type 2 diabetes and triglyceride levels. Science 316:1331–1336
Scott LJ, Mohlke KL, Bonnycastle LL et al (2007) A genome-wide association study of type 2 diabetes in Finns detects multiple susceptibility variants. Science 316:1341–1345
Wellcome Trust Case Control Consortium (2007) Genome-wide association study of 14,000 cases of seven common diseases and 3,000 shared controls. Nature 447:661–678
Zeggini E, Weedon MN, Lindgren CM et al (2007) Replication of genome-wide association signals in UK samples reveals risk loci for type 2 diabetes. Science 316:1336–1341
Sladek R, Rocheleau G, Rung J et al (2007) A genome-wide association study identifies novel risk loci for type 2 diabetes. Nature 445:881–885
McCarthy MI (2010) Genomics, type 2 diabetes, and obesity. N Engl J Med 363:2339–2350
Munafo MR, Flint J (2008) Meta-analysis of genetic association studies. Trends Genet 20:439–444
Patsopoulos NA, Evangelou E, Ioannidis JP (2008) Sensitivity of between-study heterogeneity in meta-analysis: proposed metrics and empirical evaluation. Int J Epidemiol 37:1148–1157
Crispim D, Fagundes NJ, dos Santos KG et al (2010) Polymorphisms of the UCP2 gene are associated with proliferative diabetic retinopathy in patients with diabetes mellitus. Clin Endocrinol (Oxf) 72:612–619
Cheurfa N, Dubois-Laforgue D, Ferrarezi DA et al (2008) The common −866G>A variant in the promoter of UCP2 is associated with decreased risk of coronary artery disease in type 2 diabetic men. Diabetes 57:1063–1068
Acknowledgements
The study was supported by grants from the National Natural Science Foundation of China (numbers 30400219, 30671010 and 30971405). We would like to thank M. McCarthy and A. Morris from the Oxford Centre for Diabetes, Endocrinology and Metabolism (OCDEM), Oxford, UK. Thanks also go to: R. Venkatesan from the Madras Diabetes Research Foundation and Mohan’s Diabetes Specialities Centre, India; K. S. Park and Y. M. Cho from the Department of Internal Medicine, Seoul National University College of Medicine, Korea for their data about UCP2 and UCP3 variants; and J. Congqi from the Department of Epidemiology and Health Statistics, Shandong University, China and Z. Zhang from the Department of Molecular and Genetic Toxicology, School of Public Health, Nanjing Medical University, China for their helpful comments on our revised manuscript. We also thank H. Davidson at the Barbara Davis Center for Childhood Diabetes, University of Colorado Denver, Aurora, CO, USA for reviewing and editing a late version of the manuscript.
Contribution statement
K.X. was involved in conception and design, analysis and interpretation of data, drafting of the article and revising it critically for important intellectual content. M.Z., D.C., Y.F., L.Q. and R.G. worked on collection and interpretation of data, and critical revision of the manuscript for important intellectual content. M.W., C.S. and R.Y. were involved in analysis and interpretation of data and critical revision of the manuscript for important intellectual content. T.Y. was involved in conception and design, and critical revision of the manuscript for important intellectual content. All the co-authors gave final approval of the version to be published.
Duality of interest
The authors declare that there is no duality of interest associated with this manuscript.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
ESM Table 1
Summary of manuscripts included in the meta-analysis (PDF 20.8 kb)
ESM Table 2
Egger’s publication bias test for the UCP2 -866G/A, Ala55Val C/T, and UCP3 -55C/T polymorphisms in type 2 diabetes (PDF 9.71 kb)
ESM Fig. 1
Systematic review flow diagram. n, number of studies. (PDF 17.5 kb)
Rights and permissions
About this article
Cite this article
Xu, K., Zhang, M., Cui, D. et al. UCP2 −866G/A and Ala55Val, and UCP3 −55C/T polymorphisms in association with type 2 diabetes susceptibility: a meta-analysis study. Diabetologia 54, 2315–2324 (2011). https://doi.org/10.1007/s00125-011-2245-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00125-011-2245-y