
Abstract
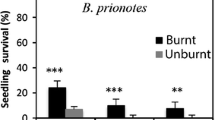
Syndromes are life history responses that are correlated to environmental regimes and are shared by a group of species (Stebbins, 1974). In the California chaparral there are two syndromes contrasted by the timing of seedling recruitment relative to wildfires. One syndrome, here called the fire-recruiter or refractory seed syndrome, includes species (both resprouting and non-resprouting) which share the feature that the timing of seedling establishment is specialized to the first rainy season after fire. Included are woody, suffrutescent and annual life forms but no geophytes have this syndrome. These species are linked by the characteristic that their seeds have a dormancy which is readily broken by environmental stimuli such as intense heat shock or chemicals leached from charred wood. Such seeds are referred to as “refractory” and dormancy, in some cases, is due to seed coat impermeability (such seeds are commonly called hardseeded), but in other cases the mechanism is unknown. Seeds of some may require cold stratification and/or light in addition to fire related stimuli. In the absence of fire related cues, a portion or all of a species’ seed pool remains dormant. Most have locally dispersed seeds that persist in the soil seed bank until the site burns. Dispersal of propagules is largely during spring and summer which facilitates the avoidance of flowering and fruiting during the summer and fall drought. Within a life form (e.g., shrub, suffrutescent, etc.), the seeds of these species have less mass than those of species with non-refractory seeds and this possibly reflects the environmental favorableness of the postfire environment for seedling establishment. Regardless of when fire occurs, germination is normally delayed until late winter or early spring. In the absence of fire, or other disturbance, opportunities for population expansion are largely lacking for species with this syndrome.
The other syndrome, here called the fire-resister or non-refractory seed syndrome, includes species that are resilient to frequent fires (mostly by vegetative resprouting), but require fire-free periods for recruiting new seedlings. Included are shrubs, subshrubs, suffrutescents, lianas, geophytes and annuals. All are linked by the characteristic that their seeds germinate in the absence of cues related to wildfires. In many cases no form of seed dormancy is present and the seeds germinate soon after dispersal; consequently these species do not accumulate a persistent seed bank. Germination and seedling establishment is independent of fire and thus opportunities for population expansion are also independent of fire. The demographic pattern of seedling recruitment varies with the life form. For shrubs, seedling recruitment may be restricted to sites free of fire for periods of a hundred years or more. Recruitment appears to require relatively mesic conditions and this may account for the patchy distribution of these species within the matrix of relatively arid sites. Finding such sites has selected for propagules specialized for wind or animal dispersal; the majority are bird dispersed. These shrub species all disperse fruits in fall and winter and this may have been selected to take advantage of migratory birds as well as to time dispersal to the winter rains typical of the mediterranean-climate. Germination typically occurs within several weeks of the first fall or winter rains. Maturation of flowers and fruits during the summer and fall drought may account for the distribution of these species on more mesic sites. Seed mass of these species is large and this may have been selected to provide an advantage to seedlings establishing under the canopy of this dense shrub community.
Resumen
Síndromes son las respuestas de ciclos biológicos correlacionados con régimenes ambientales y compartidos por un grupo de especies (Stebbins, 1974). En el Chaparral de California se encuentran dos síndromes, los cuales contrastan por el tiempo requerido para el restablecimiento de retonos en relación con los incendios forestales. Uno de estos síndromes, aquí denominado “síndrome fuego-restablecedor” o “semilla refractaria,” incluye especies (tanto retoāntes como no retonantes) que comparten la característica de que el tiempo para el establecimiento de plántulas está especializado en la primera estación de lluvias después del incendio Incluídas se encuentran formas de vida anuales, sufrutescentes y lenosas, sin embargo ninguna hierba perenne presenta este síndrome. Estas especies están unidas por la característica de poseer semillas con una latencia fácilmente interrumpida por estímulos ambientales, tales como un intenso shock de calor o productos químicos lixiviados de madera carbonizada. A este tipo de semillas se les conoce como refractarias. En ocasiones, su latencia se debe a la impermeabilidad de la capa de la semilla. En otros casos, el mecanismo es desconocido. Además de los estímulos relacionados con el fuego, las semillas de algunas especies pueden requerir de una estratificación fría y/o de luz. En ausencia de indicadores relacionados con el fuego, una parte o todo el grupo de semillas de una especie permanece latente. La mayoría cuenta con semillas dispersadas localmente, las cuales permanecen en el banco de semillas en el suelo, hasta que el lugar sufre de un incendio. La dispersión de semillas ocurre principalmente durante la primavera y el verano, lo cual impide el florecimiento y producción de frutos durante la sequía del verano y otono. Dentro de una forma de vida, las semillas de estas especies tienen una masa menor a la de aquellas especies con semillas no refractarias, lo cual refleja posiblemente el favorecimiento ambiental del ambiente posterior a un incendio en cuanto al establecimiento de plántulas. Sin tomar en cuenta cuando ocurre el incendio, la germinación se retrasa normalmente hasta finales del invierno o principios de la primavera. En ausencia de incendios o cualquier otra alteración, las oportunidades de expansión de la población son casi nulas para las especies con este síndrome.
El otro síndrome, aquí denominado “resistidorde fuego” o “semilla no refractaria,” incluye especies resistentes a incendios fréquentes (por retono vegetativo en su mayoría), las cuales requieren, sin embargo, períodos libres de incendios para restablecer nuevas plántulas. Entre las especies incluídas se encuentran arbustos, subarbustos, sufrutescentes, lianas, hierbas perennes y anuales. Todas están unidas por la característica de tener semillas que germinan en ausencia de indicadores relacionados con incendios. En muchas ocasiones, las semillas no presentan latencia alguna y germinan poco después de su dispersión. Por consiguiente, estas especies no acumulan un banco de semillas persistente. La germinación y establecimiento de plántulas es independiente del fuego y, por lo tanto, las oportunidades de expansión para la población son asimismo independientes del fuego. El patrón demográfico de restablecimiento de plántulas varía de acuerdo a la forma de vida. En el caso de los arbustos, el restablecimiento de plántulas puede estar restringido a sitios libres de incendios por períodos de cien anos o más. El restablecimiento parece requérir condiciones relativamente mésicas, lo cual puede ser la causa de la distribución desigual de estas especies dentro de una matriz de sitios relativamente áridos. El encontrar estos sitios ha seleccionado en favor de las semillas especializadas en dispersión por viento o a través de animales. La mayoría son dispersadas por aves. Todas estas especies de arbustos dispersan sus frutos durante el otono e invierno, lo cual puede haber sido seleccionado para obtener ventaja de las aves migratorias, así como para programar la dispersión de acuerdo con las lluvias de invierno, típicas del clima mediterráneo. La germinacion ocurre típicamente por varias semanas durante las primeras lluvias de otono o invierno. La maduración de las flores y frutos durante la sequía del verano y otono puede explicar la distribución de estas especies en sitios mas mésicos. Las semillas de estas especies son de masa considerable, lo cual puede haber sido seleccionado para proporcionar una ventaja al establecimiento de plántulas bajo el follaje de esta densa comunidad de arbustos.
Similar content being viewed by others

Literature Cited
Ammirati, J. F. 1967. The occurrence of annual and perennial plants on chaparral burns. M.S. Thesis. San Francisco State University, San Francisco, California.
Angevine, M. W. &B. F. Chabot. 1979. Seed germination syndromes in higher plants. Pages 188–206in O. T. Solbrig, S. Jain, G. B. Johnson, and P. H. Raven (eds.), Topics in plant population biology. Columbia University Press, New York.
Atwater, B. R. 1980. Germination, dormancy and morphology of the seeds of herbaceous ornamental plants. Seed Sci. Tech.8: 523–573.
Bailey, S. F. 1978. Foraging strategies of frugivorous birds in relation to the availability of berries, with special reference to central California. Ph.D. Dissertation. University of California, Berkeley.
Baker, H. G. 1972. Seed weight in relation to environmental conditions in California. Ecology53: 997–1010.
Barrett, R. H. 1983. Food habits of coyotes,Canis latrans, in eastern Tehama County, California. California Fish and Game69: 184–186.
Baskin, C. C. &J. M. Baskin. 1988. Germination ecophysiology of herbaceous plant species in a temperate region. Amer. J. Bot.75: 286–305.
Berg, A. R. 1974.Arctostaphylos Adans, manzanita. Pages 228–231 in C. S. Schopmeyer (ed.), Seeds of woody plants in the United States. USDA Forest Service, Agriculture Handbook No. 450.
Bewley, J. D. &M. Black. 1982. Physiology and biochemistry of seeds in relation to germination. Springer-Verlag, New York.
Boyd, R. 1985. The response to fire of three rare plant species from the Pine Hill Ecological Reserve. California Department of Fish and Game, unpublished report.
Bullock, S. 1982. Reproductive ecology ofCeanothus cordulatus. M.A. Thesis. California State University, Fresno.
Bullock, S. H. 1976. Comparison of the distribution of seed and parent-plant populations. Southw. Nat.21: 383–389.
—. 1978. Plant abundance and distribution in relation to types of seed dispersal in chaparral. Madroño25: 104–105.
—. 1989. Life history and seed dispersal of the short-lived chaparral shrubDendromecon rigida (Papaveraceae). Amer. J. Bot.76: 1506–1517.
Burk, J. H. 1978. Seasonal and diurnal water potentials in selected chaparral shrubs. Amer. Midl. Nat.99: 244–248.
Campbell, B. M. 1980. Some mixed hardwood forest communities of the coastal range of southern California. Phytocoenologia8: 297–320.
Capon, B., G. L. Maxwell &P. H. Smith. 1978. Germination responses to temperature pretreatment of seeds from ten populations ofSalvia columbariae in the San Gabriel Mountains and Mojave Desert, California. Aliso9: 365–373.
Carlson, J. R. &W. C. Sharp. 1975. Germination of high elevation manzanitas. USDA Forest Service, Tree Planters’ Notes26(3): 10–11, 25–26.
Carroll, M. C., L. L. Laughrin & A. C. Bromfield. Fire on the California islands: Does it play a role in chaparral and pine forest habitats?In E. Hochberg (ed.), Proceedings of the Third Symposium on the California Islands (in press).
Christensen, N. L. &C. H. Muller. 1975a. Effects of fire on factors controlling plant growth inAdenostoma chaparral. Ecol. Monogr.45: 29–55.
— & — 1975b. Relative importance of factors affecting germination and seedling survival inAdenostoma chaparral. Amer. Midl. Nat.93: 71–78.
Cohen, D. 1967. Optimizing reproduction in a randomly varying environment when a correlation may exist between the conditions at the time a choice has to be made and the subsequent outcome. J. Theoret. Biol.16: 1–14.
Critchfield, W. B. 1971. Profiles of California vegetation. USDA Forest Service Pacific Southwest Forest and Range Experiment Station, Research Paper PSW-76.
Cruden, R. W. 1974. The adaptive nature of seed germination inNemophila menziesii Aggr. Ecology5: 1295–1305.
Davey, J. R. 1982. Stand replacement inCeanothus crassifolius. M.S. Thesis. California State Polytechnic University, Pomona.
Davis, S. D. 1989. Patterns in mixed chaparral stands: Differential water status and seedling survival during summer drought. Pages 97–105in S. C. Keeley (ed.), The California chaparral paradigms reexamined. Natural History Museum of Los Angeles County, Science Series, No. 34.
DeBano, L. F., P. H. Dunn & C. E. Conrad 1977. Fire’s effect on physical and chemical properties of chaparral soils. Pages 65–74in H. A. Mooney & C. E. Conrad (eds.), Proceedings of the symposium on environmental consequences of fire and fuel management in mediterranean ecosystems. USDA Forest Service, General Technical Report WO-3.
DeSouza, J., P. A. Silka &S. D. Davis. 1986. Comparative physiology of burned and unburnedRhus laurina after chaparral wildfire. Oecologia71: 63–68.
Emery, D. E. 1988. Seed propagation of native California plants. Santa Barbara Botanic Garden, Santa Barbara, California.
Evans, R. A., H. H. Biswell &D. E. Palmquist. 1987. Seed dispersal inCeanothus cuneatus andC. leucodermis in a Sierran oak-woodland savanna. Madrono34: 283–293.
Everett, P. C. 1957. A summary of the culture of California plants at the Rancho Santa Ana Botanic Garden 1927–1950. Rancho Santa Ana Botanical Garden, Claremont, California.
Fiedler, P. L. 1987. Life history and population dynamics of rare and common Mariposa lilies (Calochortus Pursh: Liliaceae). J. Ecol.75: 977–995.
Flint, S. D. &I. G. Palmblad. 1978. Germination dimorphism and developmental flexibility in the ruderal weedHeterotheca grandiflora. Oecologia36: 33–43.
Geirsbach, J. 1937. Germination and seedling production inArctostaphylos uva-ursi. Contr. Boyce Thompson Inst. Pl. Res.9: 71–78.
Grant, V. 1949. Seed germination inGila capitata and its relatives. Madroño10: 87–93.
Gratkowski, H. J. 1962. Heat as a factor in germination of seeds ofCeanothus velutinus var.laevigatus T. & G. Ph.D. Dissertation. Oregon State University, Corvallis.
Grenfell, W. E., Jr. &A. J. Brody. 1983. Seasonal foods of black bears in Tahoe National Forest, California. California Fish and Game69: 132–150.
Griffin, J. R. 1977. Oak woodland. Pages 383–415in M. G. Barbour & J. Major (eds.), Terrestrial vegetation of California. John Wiley, New York.
Grinnell, J. 1936. Up-hill planters. Condor38: 80–82.
Hadley, E. B. 1961. Influence of temperature and other factors onCeanothus megacarpus seed germination. Madroño16: 132–138.
Hannan, G. L. 1980. Heteromericarpy and dual seed germination modes inPlatystemon californicus (Papaveraceae). Madroño27: 164–170.
Harper, J. L. 1977. Population biology of plants. Academic Press, New York.
Harrington, J. F. 1975. A study on the germination ofRomneya coulteri seed. Assoc. Off. Seed Anal. Newslet.49(3): 26–29.
Hellmers, H. &W. C. Ashby. 1958. Growth of native and exotic plants under controlled temperatures in the San Gabriel Mountains, California. Ecology39: 416–428.
Horn, S. 1984. Bird dispersal of toyon (Heteromeles arbutifolia). M.S. Thesis. California State University, Hayward.
Horton, J. S. &C. J. Kraebel. 1955. Development of vegetation after fire in the chamise chaparral of southern California. Ecology36: 244–262.
—. 1944. The wood rat as an ecological factor in southern California watersheds. Ecology25: 341–351.
Imamura, E. 1971. Floristic and ecological studies of chaparral communities in the Santa Barbara area, California. M.A. Thesis. San Francisco State University, San Francisco, California.
Jones, C. S. &W. H. Schlesinger. 1980.Emmenanthependuliflora (Hydrophyllaceae): Further consideration of germination response. Madroño27: 122–125.
Kaminsky, R. 1981. The microbial origin of the allelopathic potential ofAdenostomafasciculatum H. & A. Ecol. Monogr.51: 365–382.
Keeley, J. E. 1977. Seed production, seed populations in soil, and seedling production after fire for two congeneric pairs of sprouting and non-sprouting chaparral shrubs. Ecology58: 820–829.
—. 1984. Factors affecting germination of chaparral seeds. Bull. So. Cal. Acad. Sci.83: 113–120.
—. 1986. Seed germination patterns ofSalvia mellifera in fire-prone environments. Oecologia71: 1–5.
—. 1987a. Role of fire in seed germination of woody taxa in California chaparral. Ecology68: 434–443.
—. 1987b. Ten years of change in seed banks of the chaparral shrubs,Arctostaphylos glauca andA. glandulosa. Amer. Midl. Nat.117: 446–448.
—. 1986. Demographic structure of chaparral under extended fire-fire conditions. Pages 133–137in J. J. DeVries (ed.), Proceedings of the chaparral ecosystems research conference. California Water Resources Center, University of California, Davis, Report No. 62.
—. 1976. Differential seed predation on two species ofArctostaphylos (Ericaceae). Oecologia24: 71–81.
—. 1984. Posture recovery of California coastal sage scrub. Amer. Midl. Nat.111: 105–117.
—. 1987. The role of fire in the germination of chaparral herbs and suffrutescents. Madroño34: 240–249.
— 1988. Chapter 6, Chaparral. Pages 165–207in M. G. Barbour & W. D. Billings (eds.), North American terrestrial vegetation. Cambridge University Press, New York.
—. 1989. Allelopathy and the fire induced herb cycle. Pages 65–72in S. C. Keeley (ed.), The California chaparral: Paradigms re-examined. Natural History Museum of Los Angeles County, California, Science Series, No. 34.
—. 1985. Role of allelopathy alleleopathy, heat, and charred wood in the germination of chaparral herbs and suffrutescents. J. Ecol.73: 445–458.
—. 1986. Postfire recovery of chaparral along an elevational gradient in southern California. Southw. Nat.31: 177–184.
—. 1984. Garden comparison of germination and seedling growth ofYucca whipplei subspecies. Madroño31: 24–29.
—. 1978. Reproduction of chaparral shrubs after fire: A comparison of sprouting and seeding strategies. Amer. Midl. Nat.99: 142–161.
Keeley, S. C. 1977. The relationship of precipitation to post-fire succession in the southern California chaparral. Pages 387–390in H. A. Mooney & C. E. Conrad (eds.), Proceedings of the symposium on environmental consequences of fire and fuel management in mediterranean ecosystems. USDA Forest Service, General Technical Report WO-3.
—. 1981. Postfire succession of the herbaceous flora in southern California chaparral. Ecology62: 1608–1621.
—. 1986. Charred wood stimulated germination of two fire-following herbs of the California chaparral and the role of hemi-cellulose. Amer. J. Bot.73: 1289–1297.
Kelly, V. R. 1986. Seed banks and reproductive life histories in sprouting and non-sproutingArctostaphylos species. M.S. Thesis. San Francisco State University, San Francisco, California.
Koller, D. 1972. Environmental control of seed germination. Pages 1–107in T. T. Kozlowski (ed.), Seed biology. Vol. II. Academic, New York.
Krause, D. &J. Kummerow. 1977. Xeromorphic structure and soil moisture in the chaparral. Oecol. Plant.12: 133–148.
Kruse, W. H. 1970. Temperature and moisture stress affect germination ofGutierrezia sarothrae. J. Range Mgmt.23: 143–144.
Kummerow, J., G. Montenegro &D. Krause. 1981. Biomass, phenology and growth. Pages 69–96in P. C. Miller (ed.), Resource use by chaparral and matorral. Springer-Verlag, New York.
Martin, A. C. 1946. The comparative internal morphology of seeds. Amer. Midl. Nat.36: 513–660.
Matsuda, K. &J. R. McBride. 1989. Germination characteristics of selected California oak species. Amer. Midl. Nat.122: 66–76.
McPherson, J. K. &C. H. Muller. 1969. Allelopathic effects ofAdenostoma fasciculatum, “chamise,” in the California chaparral. Ecol. Monogr.39: 177–198.
Mills, J. N. 1986. Herbivores and early postfire succession in southern California chaparral. Ecology67: 1637–1649.
—. 1989. Herbivores, seed predators and chaparral succession. Pages 49–55 in S. C. Keeley (ed.), The California chaparral: Paradigms re-examined. Natural History Museum of Los Angeles County, California, Science Series, No. 34.
Minnich, R. A. 1982. Grazing, fire and the management of vegetation of Santa Catalina Island, California. Pages 444–449in C. E. Conrad & W. C. Oechel (eds.), Proceedings of the symposium on dynamics and management of mediterranean-type ecosystems. USDA Forest Service, Pacific Southwest Forest and Range Experiment Station, General Technical Report PSW-58.
Mirov, N. T. 1936. Germination behaviour of some California plants. Ecology17: 667–672.
Mooney, H. A., J. Kummerow, A. W. Johnson, D. J. Parsons, S. Keeley, A. Hoffmann, R. I. Hays, J. Giliberto &C. Chu. 1977. The producers—Their resources and adaptive resources. Pages 85–143in H. A. Mooney (ed.), Convergent evolution of Chile and California mediterranean climate ecosystems. Dowden, Hutchinson and Ross, Stroudsburg, Pennsylvania.
Munz, P. A. 1968. A California flora and supplement. University of California Press, Los Angeles.
Newton, W. C. 1979. Physiological ecology ofSalvia columbariae seed germination. M.S. Thesis. California State University, Los Angeles.
O’Leary, J. F. &W. E. Westman. 1988. Regional disturbance effects on herb succession patterns in coastal sage scrub. J. Biogeogr.15: 775–786.
Parker, V. T. 1984. Correlation of physiological divergence with reproductive mode in chaparral shrubs. Madroño31: 231–242.
—. 1987. Effects of wet-season management burns on chaparral vegetation: Implications for rare species. Pages 233–237in T. S. Elias (ed.), Conservation and management of rare and endangered plants. California Native Plant Society, Sacramento, California.
—. 1989. Seed banks in California chaparral and other mediterranean climate shrublands. Pages 231–255 in M. A. Leck, V. T. Parker & R. L. Simpson (eds.), Ecology of soil seed banks. Academic Press, San Diego.
Parrish, M. C. 1927. A study of the germination of the seeds of some native California plants. M.A. Thesis. University of California, Berkeley.
Pase, C. P. 1965. Shrub seedling regeneration after controlled burning and herbicidal treatment of dense pringle manzanita chaparral. USDA Forest Service, Rocky Mountain Forest and Range Experiment Station, Research Note RM-56.
Patric, J. H. &T. L. Hanes. 1964. Chaparral succession in a San Gabriel Mountain area of California. Ecology45: 353–360.
Pelton, J. 1962. Factors influencing survival and growth of a seedling populationof Arbutus menziesii in California. Madroño16: 237–256.
Plumb, T. R. 1982. Factors affecting germination of southern California oaks. Page 625in C. E. Conrad & W. C. Oechel (eds.), Proceedings of the symposium on dynamics and management of mediterranean-type ecosystems. USDA Forest Service, Pacific Southwest Forest and Range Experiment Station, General Technical Report PSW-58.
Poole, D. K., S. W. Roberts &P. C. Miller. 1981. Water utilization. Pages 123–149in P. C. Miller (ed.), Resource use by chaparral and matorral. Springer-Verlag, New York.
Primack, R. B. 1987. Relationships among flowers, fruits, and seeds. Annual Rev. Ecol. Syst.18: 409–430.
Quick, C. R. 1935. Notes on the germination ofCeanothus seeds. Madroño3: 135–140.
—. 1947. Germination ofPhacelia seeds. Madroño9: 17–20.
—. 1961. Germination ofCeanothus seeds. Madroño16: 23–30.
Roach, D. A. &R. D. Wulff. 1987. Maternal effects in plants. Ann. Rev. Ecol. Syst.18: 209–235.
Roberts, S. W., P. C. Miller &A. Valamanesh. 1981. Comparative field water relations of four co-occurring chaparral shrub species. Oecologia48: 360–363.
Rogers, B. 1949. Effects of fire on germination of seeds ofArctostaphylos viscida. M.S. Thesis. University of California, Berkeley.
Roof, J. 1988. Germination studies of someEriodictyon species. Four Seasons8(1): 19–34.
Rundel, P. W., G. A. Baker, D. J. Parsons &T. J. Stohlgren. 1987. Postfire demography of resprouting and seedling establishment byAdenostoma fasciculatum in the California chaparral. Pages 575–596in J. D. Tenhunen, F. M. Catarino, O. L. Lange & W. C. Oechel (eds.), Plant response to stress, functional analysis in mediterranean ecosystems. Springer-Verlag, New York.
Sampson, A. W. 1944. Plant succession and burned chaparral lands in northern California. Agricultural Experiment Station, University of California, Berkeley, Bulletin 685.
Sawyer, J. O., D. A. Thornburgh &J. R. Griffin. 1977. Mixed evergreen forest. Pages 359–381in M. G. Barbour & J. Major (eds.), Terrestrial vegetation of California. John Wiley, New York.
Schlesinger, W. H., J. T. Gray, D. S. Gill &B. E. Mahall. 1982.Ceanothus megacarpus chaparral: A synthesis of ecosystem properties during development and annual growth. Bot. Rev.48: 71–117.
Schlising, R. A. 1969. Seedling morphology inMarah (Cucurbitaceae) related to the California mediterranean climate. Amer. J. Bot.56: 552–561.
—. 1976. Reproductive proficiency inPaeonia californica (Paeoniaceae). Amer. J. Bot.63: 1095–1103.
Schopmeyer, C. S. (ed). 1974. Seeds of woody plants in the United States. USDA Forest Service, Agriculture Handbook No. 450.
Schroeder, E. M. &L. V. Barton. 1939. Germination and growth of some rock garden plants. Contr. Boyce Thompson Inst. Pl. Res.10: 235–255.
Shmida, A. &R. H. Whittaker. 1981. Pattern and biological microsite effects in two shrub communities, southern California. Ecology62: 234–251.
Smith-Huerta, N. L. 1984. Seed germination in related diploid and allotetraploidClarkia species. Bot. Gaz.145: 246–252.
Stebbins, G. L. 1974. Flowering plants. Evolution above the species level. Belknap Press of Harvard University Press, Cambridge, Massachusetts.
Stocking, S. K. 1966. Influences of fire and sodium-calcium borate on chaparral vegetation. Madroño18: 193–203.
Stone, E. C. 1951. The stimulative effect of fire on the flowering of the golden brodiaea (Brodiaea ixiodes Wats. var.lugens Jeps.). Ecology32: 534–537.
—. 1951. The effect of fire on the germination of the seed ofRhus ovata Wats. Amer. J. Bot.38: 368–372.
—. 1953. Fire stimulated germination. Calif. Agric. 7(9): 13–14.
Sweeney, J. R. 1956. Responses of vegetation to fire. A study of the herbaceous vegetation following chaparral fires. University Calif. Publ. Bot.28: 143–216.
Vankat, J. L. 1989. Water stress in chaparral shrubs in summer-rain versus summer-drought climates—Wither the mediterranean-type climate paradigm? Pages 117–124in S. C. Keeley (ed.), The California chaparral: Paradigms re-examined. Natural History Museum of Los Angeles County, California, Science Series, No. 34.
Wells, P. V. 1962. Vegetation in relation to geological substratum and fire in the San Luis Obispo quadrangle, California. Ecol. Monogr.32: 79–103.
—. 1969. The relation between mode of reproduction and extent of speciation in woody genera of the California chaparral. Evolution23: 264–267.
Went, F. W., G. Juhren &M. C. Juhren. 1952. Fire and biotic factors affecting germination. Ecology33: 351–364.
Westermeier, L. J. 1978. Effects of dry and moist heat shocks on seed viability and germinationof Lotus strigosus andLupinus excubitus var.hallii. M.A. Thesis. California State University, Fullerton.
Wicklow, D. T. 1966. Further observations on serpentine response inEmmenanthe. Ecology47: 864–865.
—. 1977. Germination response inEmmenanthe penduliflora (Hydrophyllaceae). Ecology58: 201–205.
Wright, E. 1931. The effect of high temperature on seed germination. J. For.29: 679–687.
Zammit, C. A. &P. H. Zedler. 1988. The influence of dominant shrubs, fire, and time since fire on soil seed banks in mixed chaparral. Vegetatio75: 175–187.
Zedler, P. H. 1981. Vegetation change in chaparral and desert commmunities in San Diego county, California. Pages 406–430in D. C. West, H. H. Shugart & D. Botkin (eds.), Forest succession. Concepts and applications. Springer, New York.
-. 1982. Demography and chaparral management in southern California. Pages 123–127in C. E. Conrad & W. C. Oechel (eds.), Proceedings of the symposium on environmental consequences of fire and fuel management in mediterranean ecosystems. USDA Forest Service, General Technical Report WO-3.
—. 1983. Vegetation change in response to extreme events: The effect of a short interval between fires in California chaparral and coastal scrub. Ecology64: 809–818.
Author information
Authors and Affiliations
Rights and permissions
About this article
Cite this article
Keeley, J.E. Seed germination and life history syndromes in the California chaparral. Bot. Rev 57, 81–116 (1991). https://doi.org/10.1007/BF02858766
Issue Date:
DOI: https://doi.org/10.1007/BF02858766