
Abstract
Eastern Mediterranean silvopastoral oak woodlands have been greatly damaged through forest conversion, illegal lumbering, overgrazing, and forest fires. The aim of this study was to assess land-use changes and the legacies that they have imprinted on the forest structure of Quercus macrolepis and accompanying Quercus pubescens and Quercus cerris woodlands on Lesvos Island, Greece. The size structures of adult oak populations were analyzed as indicators of long-term oak regeneration, while short-term recruitment was determined by counting oak seedlings and saplings. The size structure of the adult Q. macrolepis population was similar to the inverse J-shaped distribution typical for natural Mediterranean oak forests, indicating continuous recruitment with a constant mortality rate of mature individuals. Seedling and sapling densities were highly variable, but generally low in relation to adult oak densities. Recruitment of oak seedlings and saplings was positively related to determinants such as forest cover, adult oak density and basal area, woody plant richness, and litter cover. Both seedling and sapling occurrence were negatively associated with dung frequency, which suggests that sheep grazing imposes a barrier to oak recruitment. The study outlines a comprehensive land-use transition from the 1950 to 1970s, during which a complex and multifunctional agrosilvopastoral land-use system was simplified to an intensive grazing system. The discrepancy between the successful long-term regeneration and the less successful short-term recruitment of oaks illustrates that intensified livestock grazing has been a major driver of vegetation change. Grazing impact is likely to interact with increasing drought conditions, which may trigger a negative feedback cycle that undermines the capacity of woodlands to sustain ecosystem services.
Similar content being viewed by others

Introduction
Since the 1960s, most dryland ecosystems of the Mediterranean Basin have experienced a comprehensive land-use transition from complex and multifunctional agrosilvopastoral land-use systems to simplified and intensified forms of livestock husbandry and agriculture (Naveh 1982; Pinto-Correia and Vos 2001; Hill et al. 2008). This manifested itself in two major land-use and land-cover changes: the large-scale abandonment of croplands and—regionally varying—an intensification or an abandonment of livestock grazing. Cropland abandonment has been observed throughout the Northern Mediterranean Basin, for example in Spain (Alados et al. 2004), France (Taillefumier and Piégay 2003), Portugal (Moreira et al. 2001), Italy (Vos 1993), and Greece (Kizos and Koulouri 2006). In many regions of the Mediterranean, land abandonment initiated an extensive increase of Mediterranean shrub- and woodland vegetation (Grove and Rackham 2001). This phenomenon has put into question the classical desertification dogma regarding the Mediterranean, which had predicted an irreversible decline of biological productivity following human disturbance (Hill et al. 2008). Nevertheless, the rapid development of shrublands has often also involved negative impacts on fire regimes, losses of biodiversity associated with traditional land uses, and degradation of cultural ecosystem services (MacDonald et al. 2000).
In many parts of the Eastern Mediterranean, the recovery of vegetation through abandonment of cultivation has been accompanied by a concomitant turn toward more intensive animal husbandry, with a major increase of sheep stocks (Giourga et al. 1998). Intensified livestock husbandry is believed to shift rangeland ecosystems from equilibrium states and to initiate degradation processes (Iosifides and Politidis 2005; Röder et al. 2007; Röder et al. 2008). In the Eastern Mediterranean, overgrazing has resulted in the removal of soil cover and the domination of undesirable plants, mostly Sarcopoterium spinosum (Bakker et al. 2005). These processes have been exacerbated by changes in the spatial configuration of grazing, which is now largely uncontrolled, continuous and all-season (Giourga et al. 1998). A remote-sensing survey showed that, between 1977 and 1996, 40% of rangelands on Crete (Greece) suffered declining vegetation cover in consequence of increased grazing pressure (Hostert et al. 2003). However, grazing pressure is highly heterogeneous, so that over- and undergrazing can be observed even in immediate proximity of each other (Röder et al. 2007). Both over- and underuse can modify ecosystem structure and functions, as Mediterranean drylands are tightly coupled human–environment systems (Aranzabal et al. 2008; Röder et al. 2008). For example, a comprehensive cessation of livestock may involve loss of biodiversity and devastating wildfires (Papanastasis 2009). The combined effects of cropland abandonment and intensified livestock husbandry on plant community composition and standing biomass have become the object of a few studies on Eastern Mediterranean drylands (Giourga et al. 1998; Papanastasis 2007), but gaps in knowledge remain concerning overall impacts on forest structure and regeneration. More importantly, it remains unclear whether land-use transitions have triggered a regime shift from a socio-ecological system characterized by decreasing forest cover to one characterized by expanding woodlands, a process commonly referred to as the forest transition (Lambin and Meyfroidt 2010).
(Agri-)silvopastoral woodlands are common Mediterranean vegetation complexes. They have been shaped by human uses and correspond to different stages of regressive succession of the Mediterranean climax forests, which have virtually disappeared (Scarascia-Mugnozza et al. 2000). Many of these complexes can be considered ‘legacies’ of past land uses. Land-use legacies persist and continue to influence ecosystem structures and functions, though former uses may have been abandoned decades or centuries ago. Land-use legacies express themselves in diverse ecological phenomena, ranging from biodiversity, vegetation structure, and soil properties to biogeochemical cycles (Foster et al. 2003). The legacy of traditional grazing and cultivation practices in Mediterranean woodlands is not well understood, but it is assumed to manifest itself in elevated morphological plasticity, stress tolerance, and ecological resilience (Bergmeier 2008). Traditionally, silvopastoral woodlands have delivered a large variety of ecosystem goods, including firewood, charcoal, food for humans and animals, gums, resins, dyes, pharmaceuticals, cork, and aromatic plants. Moreover, they provide intangible ecosystem services, such as soil protection, stabilization of soils, reduction of water runoff in mountainous and hilly watersheds, maintenance of landscape beauty, and microclimate amelioration (Palahi et al. 2009; Scarascia-Mugnozza et al. 2000). Loss of resilience in the silvopastoral systems in the Western Mediterranean has been expressed in a geographically widespread and profound lack of tree regeneration, accompanied by a gradual aging and dieback of existing forest stands (Moreno and Pulido 2008). In many Mediterranean oak woodlands, there seems to be a central trade-off: livestock grazing has enhanced biodiversity and many ecosystem services (e.g. prevention of forest fires), but may endanger the long-term stability of the systems (Plieninger 2007). Only limited knowledge is available on the regeneration processes of Mediterranean oaks, and most studies have focused exclusively on Q. ilex (Espelta et al. 1995; Lookingbill and Zavala 2000; Pulido and Díaz 2005) or Q. suber (Pausas et al. 2006; Pons and Pausas 2006; Plieninger et al. 2010). The lack of published evidence either on the status of oak regeneration or on the progression or retrogression of the heavily grazed Eastern Mediterranean oak woodlands has so far prevented the detection of potential regime shifts.
Silvopastoral woodland dominated by Q. macrolepis Kotschy is a particularly under-studied type of oak woodland (Fig. 1). In consequence of conversion of forests to agricultural land, illegal lumbering, overgrazing and forest fires, Q. macrolepis stands have become marginal and fragmented into small-forested units or isolated individuals in various locations (estimated remaining area in Greece: 29,600 ha; Pantera et al. 2008). Q. macrolepis is frequently mixed with Q. pubescens Willd. and Q. cerris L. Only very limited information is available on its distribution, ecological requirements, and phytogeography. However, being one of the few deciduous oak species in the Eastern Mediterranean zone, Q. macrolepis is being increasingly appreciated as a means for desertification control, for its ability to survive after wildfires and to thrive under conditions unfavorable to other oak species, and for an acorn mast that can support both wild fauna and domestic pigs (Pantera et al. 2008). Since the European Union has declared Q. macrolepis forests to be a natural habitat type of community interest (Habitat Directive 92/43EEC), efforts to protect, manage, and expand their populations by afforestation are underway. The overall aim of the following landscape-level study is to contribute toward conservation efforts by investigating land-use history, monitoring forest structure and regeneration of Quercus woodlands, and modeling ecological and management factors that determine habitats for oak regeneration on Lesvos Island, in the Eastern Aegean (Greece). In particular, we aim to answer the following research questions:

Different types of silvopastoral oak woodlands in Filia municipality, Greece: a Open stands established on formerly arable terraces, situated close to the village; b dense oak stands on steeper slopes at the outskirts of the village; and c stands heavily infested by Sarcopoterium spinosum
-
What land-use changes have taken place since the early twentieth century that may be influential on silvopastoral oak woodlands?
-
Does forest structure (especially long-term regeneration and short-term recruitment of Q. macrolepis and accompanying oak species) reflect local land-use legacies?
-
How are patterns of oak regeneration related to current grazing intensities, environmental site characteristics, and over- and understorey plant community structure?
We chose a small-scale, terrestrial monitoring approach because subtle or gradual land-cover change is often challenging to detect using remote sensing (Mulligan et al. 2004). Especially, the relation between livestock stocking rates and vegetation cover needs to be studied at fine-grained observation scales (Röder et al. 2008) to provide ‘early warning signals’ (Kefi et al. 2007) for proximity to thresholds inducing more comprehensive land-cover conversion (compare Plieninger and Schaar 2008).
Materials and methods
Study area
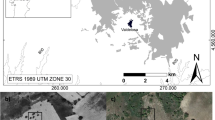
As study sites (Fig. 2), we selected silvopastoral oak woodlands in the municipality of Filia (39°15′–39°20′ N, 26°05′–26°10′ E; 2,190 ha area) in North-Central Lesvos Island. The area covers the valley of a small seasonal stream, and the village is located on a small plateau upwards from the outlet of the stream. The three hills that form the valley are 466, 682, and 785 m high; with the exception of the plateau and the pocket plain in the outlet of the stream, the area is sloping and steep. Geologically, the mountains are composed of acid volcanic parent rocks. Soils are stony, shallow, and severely eroded, described as Typic Yerochrept, Lithic Xerochrept, and Lithic Xerorthent. Typical for the Mediterranean region, the climate shows a strong seasonal variation of rainfall and high oscillations between minimum and maximum daily rainfall. Mild, humid winters alternate with hot and dry summers, with a mean annual air temperature of 17.7°C and an average rainfall of 670 mm. The estimated village population changed from 2,500 people in 1882 to 1,633 in 1920 and 1,817 in 1940, with the number continuously decreasing to 677 by 2001. Twenty-nine percent of villagers are older than 65 years and 42% older than 55 years. The agricultural sector is characterized by an increasing number of part-timers and hobby farmers and a decreasing number of professional farmers (Iosifides and Politidis 2005).

Map of Greece and location of the plots in the study area
Land-use statistics
Statistical information on land use and livestock for the Filia municipality was derived from the official data of the census for agriculture and animal husbandry, available at the settlement level from 1961 on ESYE 1964, 1978, 1994, 2003. Earlier data, not available at settlement level, were taken at the level of the entire island (ESYE 1958; GSYE 1934). In Greek agricultural statistics, cultivated area includes arable land, vines, tree crops, and fallow, while the utilized agricultural area (UAA) includes both cultivated and grazing lands.
Field measurements of forest structure and oak regeneration
As the sample for this study, we selected 70 parcels of silvopastoral woodland (a number of plots similar to that used in comparable regeneration studies, e.g. Espelta et al. 1995; Cierjacks and Hensen 2004; Dufour-Dror 2007) that were randomly distributed within five clusters across the municipality. The five clusters were marked by different land-use and environmental site characteristics in terms of distance to the village, slope, aspect, and altitude (see Fig. 2). The parcels included in each cluster were identified by a random-walk procedure (Kent and Coker 2000). The average size of the parcels, usually delimited by stone walls and/or fences, is 2.26 ha. Field measurements were performed in May and June 2009. We placed our sampling plot in the center of each parcel, with the criterion for a plot to enter the sample being the presence of at least one mature Q. macrolepis, Q. pubescens, or Q. cerris individual within a maximum distance of 15 m around the focus. Each plot consisted of three concentric circles. Mature trees were recorded within a 15-m radius (area: 706.9 m²), saplings and shrub species within a 5-m radius (area: 78.5 m²); and seedlings within a 2.5-m radius (area: 19.6 m²). For slopes >10°, we corrected the three plot radii according to the following equation:
The size structure of tree populations was used as an indicator of their age structure, as we assumed that environmental factors and competition would be distributed homogeneously within the given populations, an approach previously applied to oak silvopasture by Pulido et al. (2001) and Plieninger et al. (2003). For the analysis of forest stand structure, the number, species, and diameter at breast height (DBH) of all woody plants ≥10 cm DBH were recorded within the outer circle. The frequencies of stems in 5-cm-diameter classes were determined for the three oak species in each plot. Size-class frequency diagrams were evaluated to indirectly assess long-term regeneration, as recruitment failure translates into ‘gaps’ in the current age structures of tree populations (Ramírez and Díaz 2008).
Short-term recruitment was assessed directly by measuring two juvenile life stages of oaks delimited by size: seedlings (base diameter <1 cm) and saplings (base diameter: 1 to <10 cm). We did not separate the generative and vegetative origins of young oaks. We counted the number of all saplings of the three oak species within the 5-m radius. For each sapling, we measured base diameter of the main stem, height, and the number of stems in cases of stem aggregations. Within the inner circle, the numbers of both individuals and aggregations of oak seedlings were counted. We pooled Q. macrolepis, Q. pubescens, and Q. cerris seedlings and saplings, as it was often impossible to distinguish the species at these early life stages.
Statistical models
It was assumed that the probability of occurrence of oak seedlings and saplings was influenced by livestock grazing intensity, environmental and plant community variables. Cover of canopy, shrubs (separately for each species), forbs, rocks, litter, and bare soil were estimated by the step-point method (Evans and Love 1957). Ground and canopy cover was noted down for 100 points, situated at 1-step intervals in a 50-step transect perpendicular to the slope and another 50-step transect parallel to contours. Canopy cover was estimated by looking upward at each step and identifying whether the point was covered by an adult tree canopy or not. The two transects crossed at the center of the plot. Cover data are expressed in percent. Short-term grazing intensity was estimated by counting the number of observations of feces pellets along the steps of the two transects and is also expressed in percent of observations. Dung deposits are frequently used to estimate current grazing pressure (e.g. Posse et al. 2000; Cierjacks and Hensen 2004), and strong correlations between dung density and actual livestock presence have been experimentally confirmed for sheep by Lange and Willcocks (1978). Presence of shrub species within the 5-m radius was also noted down. Geographic position and altitude were measured with a GPS receiver. Slope was determined at each sampling point with a clinometer.
χ² tests were used for analyzing the relationship between oak seedling and sapling occurrence and the occurrence of dominant shrub species. We applied bivariate logistic regression to explore simple patterns of oak recruitment and thus potential drivers among site conditions, vegetation structure, and grazing intensity parameters. Logistic regression allows the use of a binary dependent variable and continuous, nominal or binary independent variables, none of which need to have a normal distribution (Peng et al. 2002). The presence on a plot of at least one seedling or sapling (coded as 1) or no presence at all (coded as 0) was used as the dependent variable. We analyzed oak recruitment as a binary variable rather than as a continuous one, as recruitment density did not follow a normal distribution and no transformation was found to normalize it. Hence, we followed Pausas et al. (2006) and modeled recruitment occurrence rather than recruitment density. Moreover, the occurrence of oak recruitment may be similarly indicative as recruitment density, as only a few seedlings or saplings may be required to safeguard oak regeneration. An array of continuous parameters describing site conditions (slope [°], northness,Footnote 1 bare soil cover [%], rock cover [%]), vegetation structure (forest cover [%], oak density [n ha−1], oak basal area [m² ha−1], woody plants richness [n], shrub cover [%], herbaceous cover [%], litter cover [%]), and livestock grazing (dung frequency [%]) were used as independent variables. Effect size of each single-predictor model was assessed by Nagelkerke R².
Results
Land-use change
In the early 20th century, most of the cultivated area on Western Lesvos was covered by cereals and arable crops, followed by olives and vines. An increase of cultivated land followed the population increases of the 1930 and 1940s up until the 1950s; in the Mithimna district, where Filia is located, this increase led to arable land taking more than 70% of the cultivated area, 26% of which was in areas with ‘scattered trees’ (ESYE 1958). After 1961, the utilized agricultural area in Filia increased slightly, due to the increase of grazing lands by 44% and that of tree crops (practically speaking, olives) by 81% until 2001 (Table 1). All arable uses—which were associated with extensive terracing to accommodate plowing—have decreased, practically disappearing from the 1991 and 2001 statistics. The decrease of arable crops seems to have started in 1961, when the proportion of fallow land was slightly greater than arable land. In 1971, it was ten times greater—which indicates abandonment and a shift toward grazing land. In the 1991 and 2001 statistics, fallow land was re-declared as grazing land or not declared at all. The increase of grazing land by approximately 162 ha more than the decrease of cultivated land indicates that land additional to what was officially declared ‘grazing land’ was used by livestock from 1961 to 2001, probably comprised of forest and barren lands. Grazing lands include treeless rangelands and scattered or denser oak woodlands. The period from the 1950 to 2000s has been characterized by a more than twofold increase of the sheep population, while the number of sheep farmers has sharply decreased (Table 1). Even more dramatic livestock increases are to be found for the whole of Lesvos. In 1918, the total number of sheep was 70,000, a figure that remained more or less constant until the 1950s, when it went up to 93,000; but after that the sheep population effectively tripled on the island (Fig. 3). In Filia, the decrease of the number of sheep farms was lower than that of all farms. The structural characteristics of the farms have changed as well: average farm size was five times greater in 2001 than 1961, and average plot size increased from 0.55 to 2.26 ha in the same period (Table 1), as existing farms were probably able to buy or rent additional land and increase their plots. The average number of 4 plots per farm has not changed significantly, though.

Forest structure
In total, 1,312 trees ≥10 cm DBH belonging to 14 species were recorded. Q. macrolepis (44.4% of all trees) and Q. pubescens (30.1%) were by far the most frequent tree species. Q. macrolepis occurred in 87% of the plots, Q. pubescens in 91%, and Q. cerris in 36%. Q. macrolepis and Q. pubescens species coexisted on 78.6% of the plots. Total tree densities ranged from 28.3 to 820.5 trees ha−1. Basal area extended from 2.2 to a maximum of 24.6 m² ha−1. The densities and basal areas of individual tree species are given in Table 2. Mean forest cover was 45.2% (±2.3%), ranging from very open (3% cover) to almost fully closed (92%) stands. Between one and seven tree species ≥10 cm DBH were found within one plot, resulting in a mean of 3.4 species (±1.4) per plot. In total, 24 woody species (above and below 10 cm DBH) were recorded, with Asparagus acutifolius, Phillyrea latifolia, Prunus domestica, Q. macrolepis, and Q. pubescens being present on >50% of the plots (Table 2). However, besides the dominant oaks only two species—Cistus creticus and Sarcopoterium spinosum—covered on average more than 10% of the surface in the plots where they appeared (Table 2). Forest cover was significantly related to litter cover (Spearman’s r = 0.720, P < 0.001, n = 70) and bare soil cover (Spearman’s r = −0.443, P < 0.001), but unrelated to shrub and forbs cover. In contrast, shrub and forbs cover were correlated negatively (Spearman’s r = −0.457, P < 0.001).
Long-term regeneration of oak populations
The three populations of oaks were composed of 583 Q. macrolepis, 395 Q. pubescens, and 150 Q. cerris trees. Of these, 80.8% (Q. macrolepis), 80.3% (Q. pubescens), and 71.3% (Q. cerris) were composed of one major stem, whereas 19.2, 19.7, and 28.7% were multi-stemmed associations, indicating coppice-like stand structures with a high extent of vegetative regeneration. Mean stand density was 135.21 trees ha−1 (Q. macrolepis), 87.30 trees ha−1 (Q. pubescens), and 85.84 trees ha−1 (Q. cerris), that is, in the plots where these species actually occurred (Table 2). Q. macrolepis populations showed an inverse J-shaped distribution (Fig. 4). Among all mature oaks recorded, the 10-cm and 15-cm size classes were most frequent, comprising on average 61.1% (Q. macrolepis), 52.1% (Q. pubescens), and 62.5% (Q. cerris) of all trees. The three distributions departed significantly from normality (Z = 3.872, P < 0.001 for Q. macrolepis; Z = 2.685, P < 0.001 for Q. pubescens; Z = 3.083, P < 0.001 for Q. cerris) and were positively skewed (g1 = 2.064, Q. macrolepis; g1 = 1.962, Q. pubescens; g1 = 2.008, Q. cerris). In Q. pubescens and Q. cerris stands, the 15-cm class was more strongly represented (26.5 and 34.9%) than the 10-cm class (25.6 and 27.6%), indicating a potential transition from an inverse J-shaped toward a bell-shaped size distribution. The abundance of adult size classes was positively correlated for most classes of the three oak species, but most significantly among the smaller size classes (Table 3).

Size structure of mature a Q. macrolepis (n = 61), b Q. pubescens (n = 64), and c Q. cerris (n = 25) stands (mean ± SD)
Short-term oak recruitment
All in all, 353 oak seedlings were counted in the sampling plots, of which 276 could be categorized as isolated shoots and 77 as seedling aggregations. Seedlings were absent from 31% of the plots. In those plots where seedlings occurred, densities ranged from 509.2 to 20,366.8 seedlings ha−1 (mean: 2,567.7, SD: 4,080.2). We counted 163 oak saplings in the 70 plots, with sapling densities ranging from 127.3 to 4,456.3 saplings ha−1 (mean: 296.5, SD: 605.7), where they occurred at all. Oak saplings were completely absent from 44% of the plots (Fig. 5). Sapling stem diameters ranged widely: from 1 cm to a maximum of 21 cm (mean: 4.44 cm, SD: 4.76 cm). This distribution was skewed, with 47.2% of oak saplings being in the 1-cm size class. Stem diameter and height of oak saplings were positively correlated (Spearman’s r = 0.863, P < 0.001, n = 163). Between stem diameter and the number of stems, there was a significant negative correlation (Spearman’s r = −0.250, P = 0.001, n = 163), indicating that the larger stem diameters are related to a smaller number of stems. Sapling density was positively correlated to seedling density (Spearman’s r = 0.424, P < 0.001, n = 70).

Percentage of plots containing 0 individuals, 1–5 individuals, 6–10 individuals, and >10 individuals of oak seedlings and saplings (n = 70)
Influences of grazing and other ecological factors
The occurrence of oak seedlings and saplings was significantly associated with several site, vegetation, and management parameters (Table 4). Dung frequency was negatively associated with both size classes. Forest cover, density and basal area of oak stands, woody plant richness, and litter cover showed significant positive relationships for both seedlings and saplings. We did not detect any significant associations (threshold: P < 0.05) for slope, northness, rockiness, or shrub cover. Seedlings were negatively related to bare soil, and saplings were negatively related to herbaceous cover. Occurrence of Cistus creticus shrubs was significantly and positively associated with both seedlings and saplings. Asparagus acutifolius was positively associated only with saplings. Associations with other shrubs were insignificant (Table 5).
Discussion
Land-use transition
Available statistical data on land-use and human population changes reveal the great rupture that most Mediterranean land uses experienced from the 1950 to 1970s (Pinto-Correia and Vos 2001): A complex and multifunctional agrosilvopastoral land-use system was simplified into a pure livestock-raising system. In landscape terms, the former mosaic of terraced arable land, tree crops, and pastures was replaced by more homogeneous grazed woodland. The abandonment of cereal cultivation was especially widespread on the terraces in the hill and mountain zones of the study area (Papanastasis 2007). In a wider context, this land-use transition can be interpreted as a regional specification of a global land-use trend: from subsistence agriculture and small-scale farms toward intensive agriculture (Foley et al. 2005). The transition is the result of several parallel social and economic developments. The opening of local markets to competition from more productive areas had made most extensive cultivation of the Aegean islands unprofitable since the 1950s (Papanastasis 2007). A rural exodus reduced the population of the study area by more than 60% (1940–2000s) and the number of farms by around 50% (1960–2000s). However, an increased nationwide demand for dairy and meat products provided a powerful incentive for intensified livestock raising in the 1970 and 1980s. European agricultural policies that granted per-capita subsidies for sheep and goats in the 1980 and 1990s additionally contributed toward increasing herd sizes. However, the current ongoing intensification of livestock husbandry cannot be attributed to any of these factors, since European agricultural policies have been reformed, and the demand for milk and meat products is now being satisfied by cheaper imports. Rather, it seems to be the consequence of the reduced profitability of animal production: those that choose to keep their herds face diminishing per-head profit (if any) and increasing dependence on imported feed. To maintain total revenues, these farmers have been tending to further increase their herds and, thus, have more and more decoupled livestock raising from the land (Beopoulos and Vlahos 2004; Iosifides and Politidis 2005). In consequence, a standardized and largely uncontrolled form of land use has been introduced, while cultural legacies and localized ecological knowledge have been lost (compare Hostert et al. 2003; Röder et al. 2007 for similar insight found for Northern Greece and Crete).
Land-use legacies in forest structure and regeneration
In this study, we asked whether the land-use transition outlined above has left specific legacies in terms of forest vegetation patterns, as these have also been found in deforestation–reforestation sequences elsewhere (Foster 1992; Bellemare et al. 2002; Foster et al. 2003). In general, legacies may be imprinted on several components of forest structure: First, tree species composition may be modified compared to ancient forests (Hermy and Verheyen 2007). Second, agricultural legacies may leave biotic and abiotic characteristics, such as multiple-stemmed trees, whereas elements of ancient forests, such as large trees, dead snags, and uproot mounds and pits, are rare (Foster et al. 2003). Third, and most importantly, agricultural abandonment may generate specific age and size structures of forests (Marks and Gardescu 2001). We recorded many species and elements in our plots that bear witness to an agrosilvopastoral past, for example cultivated tree species such as Olea europea, Prunus domestica, Pyrus communis, and Prunus dulcis. Indeed, even the wide distribution of Quercus macrolepis can be considered an enduring land-use legacy, as oaks populations were deliberately expanded by humans at the end of the 19th century to use oak cupula for extraction of tanning agents (Bergmeier 2008). Frequent terraces (Fig. 1a), threshing floors, stone walls, farm infrastructural elements, and a proportion of 19–29% multi-stemmed mature trees present further evidence of past land uses.
We have documented a major land-use legacy in the diameter structures of silvopastoral oak woodlands, which we interpret as an indicator for long-term regeneration: The size structure of the Q. macrolepis population is similar to the inverse J-shaped distribution typical for natural Mediterranean oak forests (Pulido et al. 2001). Assuming a close relationship between tree rings and diameter—as has been demonstrated for Q. ilex stands in the Mediterranean for homogeneous environments (Pulido et al. 2001)—the population structures of both oak species correspond to those of natural multi-age forest stands. A positively skewed age distribution indicates continuous recruitment with a constant mortality rate of mature individuals, resulting in long-term persistence of the tree populations in the absence of exogenous disturbance (Oliver and Larson 1996). The size distribution of the studied oak populations suggests that the traditional land-use system has supported continuous regeneration. Our data further show that short-term recruitment, as determined by seedling and sapling counts, is highly variable, but generally low in relation to adult oak densities. Seedling densities seem similar to the mean values of Q. pubescens (700 seedlings ha−1) and Q. ilex (1,600 seedlings ha−1) in a less intensively used mixed stand in Northern Spain (Lookingbill and Zavala 2000) and to those in a mixed marginal stand of Q. suber and Pinus pinaster in Eastern Spain (mean: 774 individuals ha−1; Pausas et al. 2006). Sapling densities are clearly lower than those of Q. petraea (1,760.8 saplings ha−1) and Q. pyrenaica (1,023.3 saplings ha−1) in wood pastures of Central Spain (Pardo et al. 2004). In conclusion, the legacy of the land-use transition that we studied becomes evident in a major discrepancy between the successful long-term regeneration (as indicated by size-structure analysis) and the less successful short-term recruitment of oaks. Oak trees are mere legacies of the former, rather than active components of the current, land-use system.
Grazing and other ecological factors related to recruitment
Both oak seedlings and saplings are in consistent negative association with dung frequency, confirming the barrier that grazing can impose on oak regeneration. The significant impact of grazing on regeneration of Q. ithaburensis—a near relative species to Q. macrolepis—has been demonstrated by a study in Israel (Dufour-Dror 2007). In a comparison between a grazed and an ungrazed oak stand, the densities of seedlings were 61% and of sapling 67% lower in the grazed treatment. A similar relationship has been found for Q. ilex in South-East Spain (Cierjacks and Hensen 2004). This finding shows that the intensification of livestock grazing has been influential on forest regeneration and supports the view that Mediterranean vegetation is generally resilient against light and moderate grazing, but that impacts increase sharply beyond a certain threshold of grazing intensity (Plieninger 2007; Köchy et al. 2008). Recruitment of oak seedlings and saplings is also related to determinants such as forest cover, adult oak density and basal area, woody plant richness, and litter cover. A relationship of basal area with occurrence or density of recruitment has also been found for other oak species in the Western Mediterranean (Lookingbill and Zavala 2000; Pausas et al. 2006). On the one hand, this can be interpreted as an effect of the elevated acorn input from mature oaks. On the other hand, it could be a consequence of more favorable microclimatic conditions in terms of the radiation and soil moisture of dense stands. The association of higher seedling and sapling densities with elevated woody plant diversity is difficult to interpret. Both may be related to a common cause, such as livestock grazing, which may hinder oak recruitment and the establishment of grazing-sensitive woody species at the same time. Together with the finding that forest cover (and not only oak cover) is a significant variable, this indicates that mixed woodlands are appropriate or even better habitats for oak recruitment than monospecific oak woodlands. Litter cover seems to support recruitment by increasing soil moisture and, thus, water availability for oak seedlings (Espelta et al. 1995). Bare soil is negatively associated with seedling recruitment, while herbaceous cover negatively impacts sapling occurrence. Thus, it seems that seedlings are more sensitive to the adverse conditions of open microsites, such as livestock trampling and insulation, whereas saplings suffer more from the competition of herbs. We cannot confirm that rocky outcrops and shrubs facilitate or inhibit oak recruitment. An exception is the common Cistus creticus shrub, which seems to exert net facilitative effects on oak recruitment.
Representativeness of the study and remaining uncertainties
This study breaks new ground as it provides, to our knowledge, the first quantitative description of the diameter structures of silvopastoral oak woodlands and their relationship to the land-use transition undergone by the Eastern Mediterranean in the latter half of the 20th century. Therefore, the representativeness and remaining uncertainties of the study need to be scrutinized. In terms of representativeness, our study area covered around 1,550 ha of Q. macrolepis woodland, which represents a third of the Q. macrolepis population on Lesvos Island (5,000 ha) and ~5% of the total distribution area of the species in Greece (ca. 29,600 ha) (Pantera et al. 2008). We focused on the core area of oak populations in North-Central Lesvos (where contiguous and closed woodlands prevail), rather than on those at the edge of the species distribution (where single oaks are scattered into open rangelands), as we were mostly interested in the impact of different land uses and topographic situations and intended to exclude other factors prevailing in more extreme environmental conditions, such as a semi-arid climatic regime in the Western part of the island. Therefore, our study may rather underestimate the failure of regeneration. Uncertainties remain about the accuracy of the use of tree size structure as a surrogate for age structure, as equating these parameters implies that there are no other significant effects on tree size than age. Unfortunately, studies of direct age determination of Q. macrolepis are as of yet unavailable. Thus, further research, possibly with the use of remote sensing, is needed to confirm the long-term oak population trends that we observed and to fully understand the forest transition of Mediterranean oak woodlands. Another uncertainty persists as to the relationship between the land-use transition outlined and oak regeneration. In this study, we reveal a temporal match between intensified grazing and retrogressive oak regeneration. But our approach does not allow the establishment of a firm causality, as we contrasted land-use and vegetation data at a relatively coarse spatial scale—the level of a municipality. Spatially explicit integration of socioeconomic and vegetation data at plot level may contribute further evidence and corroborate a causality between land-use transition and forest regeneration.
Conservation and management implications
Low recruitment rates and current land-use trends indicate that the long-term persistence of silvopastoral oak woodlands may be at risk. Socioeconomic developments, such as increasing production costs (mostly due to high forage prices) and decreasing sales prices for livestock, have been leading to further increases in grazing intensities. An additional stress that will be testing the resilience of Eastern Mediterranean grazing systems is increasing drought conditions (as predicted by IPCC scenarios). The past decades have shown a gradual reduction in rainfall and a subsequent reduction of tree growth in the Aegean Islands (Körner et al. 2005). The predicted climate changes are likely to interact with grazing impacts, which may trigger a negative feedback cycle that increases soil erosion (Köchy et al. 2008), reduces carrying capacity, and in the end undermines the capacity of rangelands to sustain ecosystem services (Körner et al. 2005). The inclusion of Q. macrolepis forests into the European Union’s Habitat Directive as a habitat type of community interest entails the ensuring of favorable conservation status through the formulation of a sound management plan. Our results suggest that oak regeneration can be enhanced by adaptive management of livestock grazing. For Q. ithaburensis, Dufour-Dror (2007) proposes a stocking rate limit of 0.7 livestock units ha−1. Livestock management could also be improved by a controlled rotational system of grazing over several parcels and a seasonal steering of herds according to forage availability and depletion (Gutman et al. 1999). A highly important task, however, will be to restore lost traditional knowledge on oak regeneration in cultivated and grazed landscapes through employing participatory approaches among resource users (Berkes et al. 2000). This knowledge may help to foster resilience by promoting appropriate resource usage and oak regeneration at the same time.
Notes
This is the degree to which an aspect is north: values from −1 (south) to 1 (north), calculated as cos(aspect*π/180).
References
Alados CL, Pueyo Y, Barrantes O, Escos J, Giner L, Robles AB (2004) Variations in landscape patterns and vegetation cover between 1957 and 1994 in a semiarid Mediterranean ecosystem. Landscape Ecol 19:543–559. doi:10.1023/B:LAND.0000036149.96664.9a
Aranzabal I, Schmitz MF, Aquilera P, Pineda FD (2008) Modelling of landscape changes derived from the dynamics of socio-ecological systems: a case of study in a semiarid Mediterranean landscape. Ecol Indic 8:672–685. doi:10.1016/j.ecolind.2007.11.003
Bakker MM, Govers G, Kosmas C, Vanacker V, van Oost K, Rounsevell M (2005) Soil erosion as a driver of land-use change. Agr Ecosyst Environ 105:467–481. doi:10.1016/j.agee.2004.07.009
Bellemare J, Motzkin G, Foster DR (2002) Legacies of the agricultural past in the forested present: an assessment of historical land-use effects on rich mesic forests. J Biogeogr 29:1401–1420. doi:10.1046/j.1365-2699.2002.00762.x
Beopoulos N, Vlahos G (2004) Exploitation of pastures in a sensitive natural environment: the case of Western Lesvos. Adv Geoecol 37:183–194
Bergmeier E (2008) Xero-thermophilous broadleaved forests and wooded pastures in the EU Habitats Directive: what is a favourable conservation status? Berichte der Reinhold-Tüxen-Gesellschaft 20:108–124
Berkes F, Colding J, Folke C (2000) Rediscovery of traditional ecological knowledge as adaptive management. Ecol Appl 10:1251–1262. doi:10.1890/1051-0761(2000)010[1251:ROTEKA]2.0.CO;2
Cierjacks A, Hensen I (2004) Variation of stand structure and regeneration of Mediterranean holm oak along a grazing intensity gradient. Plant Ecol 173:215–223. doi:10.1023/B:VEGE.0000029322.75004.ad
Dufour-Dror JM (2007) Influence of cattle grazing on the density of oak seedlings and saplings in a Tabor oak forest in Israel. Acta Oecol 31:223–228. doi:10.1016/j.actao.2006.11.003
Espelta JM, Riba M, Retana J (1995) Patterns of seedling recruitment in West-Mediterranean Quercus ilex forests influenced by canopy development. J Veg Sci 6:465–472. doi:10.2307/3236344
ESYE (1958) Apotelesmata Georgikis Apografis tis Ellados 1950 (Results of the agricultural census 1950). National Print, Athens
ESYE (1964) Apotelesmata tis Apografis Georgias—Ktinotrofias tis Ellados 1961 (Results of the census for agriculture-animal husbandry 1961). ESYE, Athens
ESYE (1978) Apotelesmata Apografis Georgias—Ktinotrofias tis 14 Martiou 1971 (Results of the census for agriculture–animal husbandry of 14 March 1971). ESYE, Athens
ESYE (1994) Apotelesmata Apografis Georgias—Ktinotrofias 1991 (Results of the census for agriculture–animal husbandry of 1991). ESYE, Athens
ESYE (2003) Apotelesmata Apografis Georgias—Ktinotrofias 2001 (Results of the census for agriculture-animal husbandry of 2001). Available at http://www.statistics.gr. Accessed 20/10/09. ESYE, Athens
Evans RA, Love RM (1957) The step-point method of sampling: a practical tool in range research. J Range Manage 10:208–212. doi:10.2307/3894015
Foley JA, DeFries R, Asner GP, Barford C, Bonan G, Carpenter SR, Chapin FS, Coe MT, Daily GC, Gibbs HK, Helkowski JH, Holloway T, Howard EA, Kucharik CJ, Monfreda C, Patz JA, Prentice IC, Ramankutty N, Snyder PK (2005) Global consequences of land use. Science 309:570–574. doi:10.1126/science.1111772
Foster DR (1992) Land-use history (1730–1990) and vegetation dynamics in Central New-England, USA. J Ecol 80:753–772. doi:10.2307/2260864
Foster D, Swanson F, Aber J, Burke I, Brokaw N, Tilman D, Knapp A (2003) The importance of land-use legacies to ecology and conservation. Bioscience 53:77–88. doi:10.1641/0006-3568(2003)053[0077:TIOLUL]2.0.CO;2
Giourga H, Margaris NS, Vokou D (1998) Effects of grazing pressure on succession process and productivity of old fields on Mediterranean Islands. Environ Manage 22:589–596. doi:10.1007/s002679900130
Grove AT, Rackham O (2001) The nature of Mediterranean Europe: an ecological history. Yale University Press, New Haven, CT, London
GSYE (1934) Georgiki kai Ktinotrofiki Apografi tis Ellados 1929 (Agricultural and livestock census 1929). Hellenic Republic, Athens
Gutman M, Holzer Z, Baram H, Noy-Meir I, Seligman NG (1999) Heavy stocking of beef cattle and early season deferment of grazing on Mediterranean-type grassland. J Range Manage 52:590–599
Hermy M, Verheyen K (2007) Legacies of the past in the present-day forest biodiversity: a review of past land-use effects on forest plant species composition and diversity. Ecol Res 22:361–371. doi:10.1007/s11284-007-0354-3
Hill J, Stellmes M, Udelhoven T, Röder A, Sommer S (2008) Mediterranean desertification and land degradation: mapping related land use change syndromes based on satellite observations. Global Planet Change 64:146–157. doi:10.1016/j.gloplacha.2008.10.005
Hostert P, Röder A, Hill J, Udelhoven T, Tsiourlis G (2003) Retrospective studies of grazing-induced land degradation: a case study in Central Crete, Greece. I J Remote Sens 24:4019–4034. doi:10.1080/0143116031000103844
Iosifides T, Politidis T (2005) Socio-economic dynamics, local development and desertification in Western Lesvos, Greece. Local Environ 10:487–499. doi:10.1080/13549830500203162
Kefi S, Rietkerk M, Alados CL, Pueyo Y, Papanastasis VP, ElAich A, de Ruiter PC (2007) Spatial vegetation patterns and imminent desertification in Mediterranean arid ecosystems. Nature 449:U213–U215. doi:10.1038/nature06111
Kent M, Coker P (2000) Vegetation description and analysis: a practical approach. Wiley, New York, NY
Kizos T, Koulouri M (2006) Agricultural landscape dynamics in the Mediterranean: Lesvos (Greece) case study using evidence from the last three centuries. Environ Sci Policy 9:330–342. doi:10.1016/j.envsci.2006.02.002
Köchy M, Mathaj M, Jeltsch F, Malkinson D (2008) Resilience of stocking capacity to changing climate in arid to Mediterranean landscapes. Reg Environ Change 8:73–87. doi:10.1007/s10113-008-0048-6
Körner C, Sarris D, Christodoulaiks D (2005) Long-term increase in climatic dryness in the East-Mediterranean as evidenced for the island of Samos. Reg Environ Change 5:27–36. doi:10.1007/s10113-004-0091-x
Lambin EF, Meyfroidt P (2010) Land use transitions: socio-ecological feedback versus socio-economic change. Land Use Policy 27:108–118. doi:10.1016/j.landusepol.2009.09.003
Lange RT, Willcocks MC (1978) Relation between sheep-time spent and egesta accumulated within an arid zone paddock. Aust J Exp Agr 18:764–767. doi:10.1071/EA9780764
Lookingbill TR, Zavala MA (2000) Spatial pattern of Quercus ilex and Quercus pubescens recruitment in Pinus halepensis dominated woodlands. J Veg Sci 11:607–612. doi:10.2307/3246590
MacDonald D, Crabtree JR, Wiesinger G, Dax T, Stamou N, Fleury P, Lazpita JG, Gibon A (2000) Agricultural abandonment in mountain areas of Europe: environmental consequences and policy response. J Environ Manage 59:47–69. doi:10.1006/jema.1999.0335
Marks PL, Gardescu S (2001) Inferring forest stand history from observational field evidence. In: Egan D, Howell EA (eds) The historical ecology handbook. Island Press, Washington D.C., pp 177–198
Moreira F, Rego FC, Ferreira PG (2001) Temporal (1958–1995) pattern of change in a cultural landscape of northwestern Portugal: implications of fire occurence. Landscape Ecol 16:557–567. doi:10.1023/A:1013130528470
Moreno G, Pulido FJ (2008) The function, management and persistence of dehesas. In: Rigueiro A, Mosquera MR, McAdam J (eds) Agroforestry systems in Europe current status and future prospects. Springer, Heidelberg, Berlin, New York, pp 127–160
Mulligan M, Burke SM, Ramos C (2004) Climate change, land-use change and the “desertification” of Mediterranean Europe. In: Mazzoleni S, Di Pasquale G, Mulligan M, Di Martino P, Rego F (eds) Recent dynamics of Mediterranean vegetation and landscape. Wiley & Son, New York, pp 259–280
Naveh Z (1982) Mediterranean landscape evolution and degradation as multivariate biofunctions—theoretical and practical implications. Landscape Plan 9:125–146. doi:10.1016/0304-3924(82)90003-X
Oliver CD, Larson BC (1996) Forest stand dynamics. Wiley, New York, NY
Palahi M, Birot Y, Bravo F, Gorriz E (2009) Modelling, valuing and managing Mediterranean forest ecosystems for non-timber goods and services. In: EFI Proceedings 57. European Forest Institute, Joensuu
Pantera A, Papadopoulos AM, Fotiadis G, Papanastasis VP (2008) Distribution and phytogeographical analysis of Quercus ithaburensis ssp. macrolepis in Greece. Ecol Mediterr 34:73–82
Papanastasis VP (2007) Land abandonment and old field dynamics in Greece. In: Cramer VA, Hobbs RJ (eds) Old fields: dynamics and restoration of abandoned farmland. Island Press, Washington DC, pp 225–246
Papanastasis VP (2009) Restoration of degraded grazing lands through grazing management: can it work? Restor Ecol 17:441–445. doi:10.1111/j.1526-100X.2009.00567.x
Pardo F, Gil L, Pardos JA (2004) Structure and composition of pole-stage stands developed in an ancient wood pasture in central Spain. Forestry 77:67–74. doi:10.1093/forestry/77.1.67
Pausas JG, Ribeiro E, Dias SG, Pons J, Beseler C (2006) Regeneration of a marginal Quercus suber forest in the Eastern Iberian Peninsula. J Veg Sci 17:729–738. doi:10.1658/1100-9233(2006)17[729:ROAMQS]2.0.CO;2
Peng CYJ, Lee KL, Ingersoll GM (2002) An introduction to logistic regression analysis and reporting. J Educ Res 96:3–14. doi:10.1080/00220670209598786
Pinto-Correia T, Vos W (2001) Multifunctionality in Mediterranean landscapes—past and future. In: Jongman RHG (ed) The new dimensions of the European landscape. Springer, Dordrecht, pp 135–164
Plieninger T (2007) Compatibility of livestock grazing with stand regeneration in Mediterranean holm oak parklands. J Nat Conserv 15:1–9. doi:10.1016/j.jnc.2005.09.002
Plieninger T, Schaar M (2008) Modification of land cover in a traditional agroforestry system in Spain: processes of tree expansion and regression. Ecol Soc 13:25. http://www.ecologyandsociety.org/vol13/iss2/art25/
Plieninger T, Pulido FJ, Konold W (2003) Effects of land use history on size structure of holm oak stands in Spanish dehesas: implications for conservation and restoration. Environ Conserv 30:61–70. doi:10.1017/S0376892903000055
Plieninger T, Rolo V, Moreno G (2010) Large-scale patterns of Quercus ilex, Quercus suber, and Quercus pyrenaica regeneration in Central-Western Spain. Ecosystems 13:644–660. doi:10.1007/s10021-010-9345-2
Pons J, Pausas JG (2006) Oak regeneration in heterogeneous landscapes: the case of fragmented Quercus suber forests in the Eastern Iberian Peninsula. Forest Ecol Manag 231:196–204. doi:10.1016/j.foreco.2006.05.049
Posse G, Anchorena J, Collantes MB (2000) Spatial micro-patterns in the steppe of Tierra del Fuego induced by sheep grazing. J Veg Sci 11:43–50. doi:10.2307/3236774
Pulido FJ, Díaz M (2005) Regeneration of a Mediterranean oak: a whole cycle approach. Ecoscience 12:92–102. doi:10.2980/i1195-6860-12-1-92.1
Pulido FJ, Díaz M, Hidalgo de Trucios S (2001) Size-structure and regeneration of Spanish holm oak Quercus ilex forests and dehesas: effects of agroforestry use on their long-term sustainability. Forest Ecol Manag 146:1–13. doi:10.1016/S0378-1127(00)00443-6
Ramírez JA, Díaz M (2008) The role of temporal shrub encroachment for the maintenance of Spanish holm oak Quercus ilex dehesas. Forest Ecol Manag 255:1976–1983. doi:10.1016/j.foreco.2007.12.019
Röder A, Kuemmerle T, Hill J, Papanastasis VP, Tsiourlis GM (2007) Adaptation of a grazing gradient concept to heterogeneous Mediterranean rangelands using cost surface modelling. Ecol Model 204:387–398. doi:10.1016/j.ecolmodel.2007.01.022
Röder A, Udelhoven T, Hill J, del Barrio G, Tsiourlis G (2008) Trend analysis of Landsat-TM and -ETM + imagery to monitor grazing impact in a rangeland ecosystem in Northern Greece. Remote Sens Environ 112:2863–2875. doi:10.1016/j.rse.2008.01.018
Scarascia-Mugnozza G, Oswald H, Piussi P, Radoglou K (2000) Forests of the Mediterranean region: gaps in knowledge and research needs. Forest Ecol Manag 132:97–109. doi:10.1016/S0378-1127(00)00383-2
Taillefumier F, Piégay H (2003) Contemporary land use changes in prealpine Mediterranean mountains: a multivariate GIS-based approach applied to two municipalities in the Southern French Prealps. Catena 51:267–296. doi:10.1016/S0341-8162(02)00168-6
Vos W (1993) Recent landscape transformation in the Tuscan Apennines caused by changing land-use. Landsc Urban Plan 24:63–68. doi:10.1016/0169-2046(93)90084-Q
Acknowledgments
This study has been financed by IKYDA, an integrated action program coordinated between the German Academic Exchange Service (DAAD) and the Greek State Scholarship Foundation (IKY). T.P.’s contribution was funded by the German Ministry of Education and Research (FKZ 01UU0904A). We are grateful to two anonymous reviewers for helpful comments on this manuscript and to Christopher Hank for improving the language of this paper.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Plieninger, T., Schaich, H. & Kizos, T. Land-use legacies in the forest structure of silvopastoral oak woodlands in the Eastern Mediterranean. Reg Environ Change 11, 603–615 (2011). https://doi.org/10.1007/s10113-010-0192-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10113-010-0192-7